
Beneficial Effects of Indigenous Probiotics in High-Cholesterol Diet-Induced Hypercholesterolemic Rats

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Probiotic Strains and Culture Condition
2.2. Animal Housing and Feeding
- (Control), control group received 2 mL of PBS;
- (Probiotics), probiotics control group received 1 mL of 1 × 109 CFU/mL mixed three probiotic strains and 1 mL of PBS;
- (HCD), HCD group received 1 mL of HCD and 1 mL of PBS;
- (HCD-L), HCD supplemented with L. reuteri TF-7 group received 1 mL of HCD and 1 mL of 1 × 109 CFU/mL L. reuteri TF-7;
- (HCD-E), HCD supplemented with E. faecium TF-18 group received 1 mL of HCD and 1 mL of 1 × 109 CFU/mL E. faecium TF-18;
- (HCD-B), HCD supplemented with B. animalis TA-1 group received 1 mL of HCD and 1 mL of 1 × 109 CFU/mL B. animalis TA-1;
- (HCD-mix), HCD supplemented with mixed three probiotic strains group received 1 mL of HCD and 1 mL of 1 × 109 CFU/mL mixed three probiotic strains.
2.3. Sample Collection
2.4. Biochemical Analysis
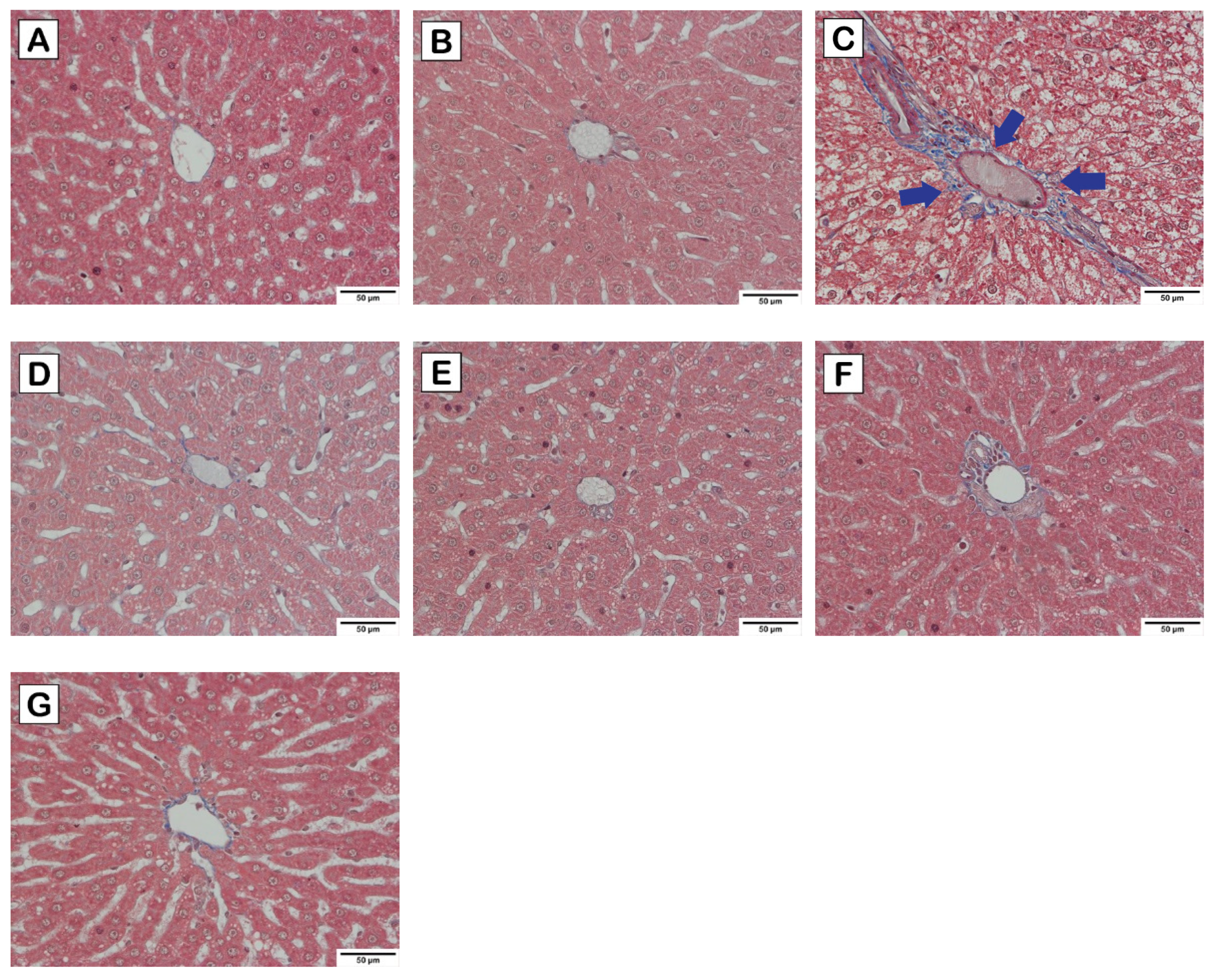
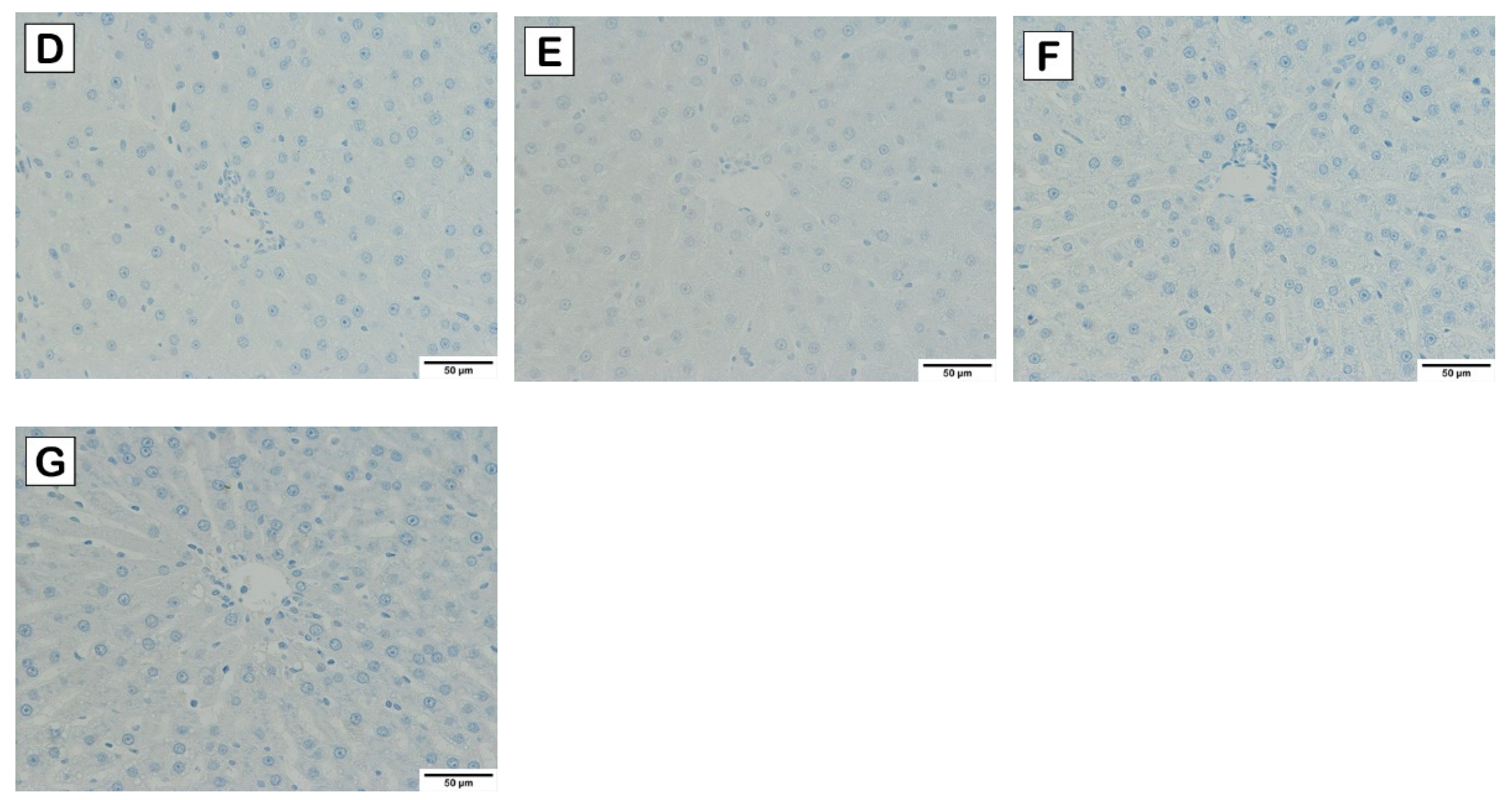
2.5. Liver Histological Analysis
2.6. Fecal Microbiota Analysis
2.7. Statistical Analysis
3. Results
3.1. Effects of Cholesterol-Lowering Probiotics on Body Weight Gain, Food Intake, Food Efficiency, and Visceral Organ Indexes of Rats
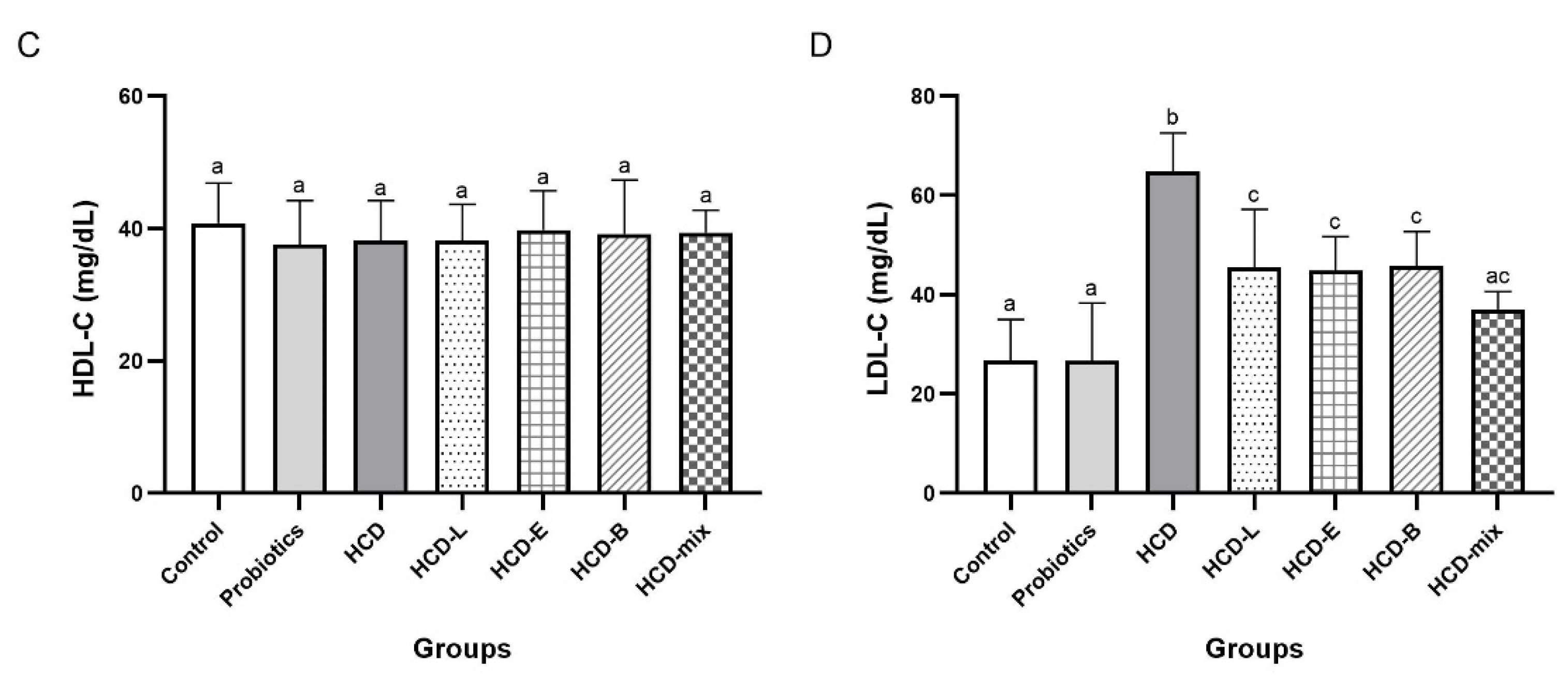
3.2. Effects of Cholesterol-Lowering Probiotics on Lipid Profiles of Rats
3.3. Effects of Cholesterol-Lowering Probiotics on Hepatic Steatosis of Rats
3.4. Effects of Cholesterol-Lowering Probiotics on Modulating Fecal Microbiota of Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jeong, M.J.; Kim, S.R.; Jung, U.J. Schizandrin A supplementation improves nonalcoholic fatty liver disease in mice fed a high-fat and high-cholesterol diet. Nutr. Res. 2019, 64, 64–71. [Google Scholar] [CrossRef] [PubMed]
- WHO. Cardiovascular Disease; WHO: Geneva, Switzerland, 2009; p. 317. [Google Scholar]
- Dai, F.J.; Hsu, W.H.; Huang, J.J.; Wu, S.C. Effect of pigeon pea (Cajanus cajan L.) on high-fat diet-induced hypercholesterolemia in hamsters. Food Chem. Toxicol. 2013, 53, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Bellosta, S.; Paoletti, R.; Corsini, A. Safety of statins: Focus on clinical pharmacokinetics and drug interactions. Circulation 2004, 109, 50–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The international scientific association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Xavier-Santos, D.; Bedani, R.; Lima, E.D.; Saad, S.M.I. Impact of probiotics and prebiotics targeting metabolic syndrome. J. Funct. Foods 2020, 64, e103666. [Google Scholar] [CrossRef]
- Wang, P.; Gao, X.; Li, Y.; Wang, S.; Yu, J.; Wei, Y. Bacillus natto regulates gut microbiota and adipose tissue accumulation in a high-fat diet mouse model of obesity. J. Funct. Foods 2020, 68, e103923. [Google Scholar] [CrossRef]
- Shehata, M.G.; El Sohaimy, S.A.; El-Sahn, M.A.; Youssef, M.M. Screening of isolated potential probiotic lactic acid bacteria for cholesterol lowering property and bile salt hydrolase activity. Ann. Agric. Sci. 2016, 61, 65–75. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.B.; Lew, L.C.; Yeo, S.K.; Nair Parvathy, S.; Liong, M.T. Probiotics and the BSH-related cholesterol lowering mechanism: A Jekyll and Hyde scenario. Crit. Rev. Biotechnol. 2015, 35, 392–401. [Google Scholar] [CrossRef]
- Mainville, I.; Arcand, Y.; Farnworth, E.R. A dynamic model that simulates the human upper gastrointestinal tract for the study of probiotics. Int. J. Food Microbiol. 2005, 99, 287–296. [Google Scholar] [CrossRef]
- Jarocki, P.; Podlesny, M.; Glibowski, P.; Targonski, Z. A new insight into the physiological role of bile salt hydrolase among intestinal bacteria from the genus Bifidobacterium. PLoS ONE 2014, 9, e114379. [Google Scholar] [CrossRef] [Green Version]
- Lye, H.S.; Rahmat-Ali, G.R.; Liong, M.T. Mechanisms of cholesterol removal by lactobacilli under conditions that mimic the human gastrointestinal tract. Int. Dairy J. 2010, 20, 169–175. [Google Scholar] [CrossRef]
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, B.; Delgado, S.; Blanco-Miguez, A.; Lourenco, A.; Gueimonde, M.; Margolles, A. Probiotics, gut microbiota, and their influence on host health and disease. Mol. Nutr. Food Res. 2017, 61, e1600240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puttarat, N.; Ladda, B.; Kasorn, A.; Tanasupawat, S.; Taweechotipatr, M. Cholesterol-lowering activity and functional characterization of lactic acid bacteria isolated from traditional Thai foods for their potential used as probiotics. SJST 2021, 43, 1283–1291. [Google Scholar]
- Paigen, B.; Morrow, A.; Brandon, C.; Mitchell, D.; Holmes, P. Variation in susceptibility to atherosclerosis among inbred strains of mice. Atherosclerosis 1985, 57, 65–73. [Google Scholar] [CrossRef]
- Ding, W.; Shi, C.; Chen, M.; Zhou, J.; Long, R.; Guo, X. Screening for lactic acid bacteria in traditional fermented Tibetan yak milk and evaluating their probiotic and cholesterol-lowering potentials in rats fed a high-cholesterol diet. J. Funct. Foods 2017, 32, 324–332. [Google Scholar] [CrossRef]
- Karimi, G.; Jamaluddin, R.; Mohtarrudin, N.; Ahmad, Z.; Khazaai, H.; Parvaneh, M. Single-species versus dual-species probiotic supplementation as an emerging therapeutic strategy for obesity. Nutr. Metab. Cardiovasc. Dis. 2017, 27, 910–918. [Google Scholar] [CrossRef] [Green Version]
- Khani, S.; Hosseini, H.M.; Taheri, M.; Nourani, M.R.; Fooladi, A.I. Probiotics as an alternative strategy for prevention and treatment of human diseases: A review. Inflamm. Allergy Drug Targets 2012, 11, 79–89. [Google Scholar] [CrossRef]
- Kim, S.J.; Park, S.H.; Sin, H.S.; Jang, S.H.; Lee, S.W.; Kim, S.Y.; Kwon, B.; Yu, K.Y.; Kim, S.Y.; Yang, D.K. Hypocholesterolemic effects of probiotic mixture on diet-induced hypercholesterolemic rats. Nutrients 2017, 9, 293. [Google Scholar] [CrossRef] [Green Version]
- Al-Muzafar, H.M.; Amin, K.A. Probiotic mixture improves fatty liver disease by virtue of its action on lipid profiles, leptin, and inflammatory biomarkers. BMC Complement. Altern. Med. 2017, 17, 43. [Google Scholar] [CrossRef] [Green Version]
- Saikia, D.; Manhar, A.K.; Deka, B.; Roy, R.; Gupta, K.; Namsa, N.D.; Chattopadhyay, P.; Doley, R.; Mandal, M. Hypocholesterolemic activity of indigenous probiotic isolate Saccharomyces cerevisiae ARDMC1 in a rat model. J. Food Drug Anal. 2018, 26, 154–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sangwan, S.; Singh, R. Synergistic effect of oats and LGG fermented milk on lowering hypercholesterolemia in rats. J. Cereal Sci. 2018, 82, 164–169. [Google Scholar] [CrossRef]
- Mallappa, R.H.; Rokana, N.; Duary, R.K.; Panwar, H.; Batish, V.K.; Grover, S. Management of metabolic syndrome through probiotic and prebiotic interventions. Indian J. Endocrinol. Metab. 2012, 16, 20–27. [Google Scholar] [PubMed]
- Rafiq, N.; Younossi, Z.M. Nonalcoholic fatty liver disease: A practical approach to evaluation and management. Clin. Liver Dis. 2009, 13, 249–266. [Google Scholar] [CrossRef]
- Nabavi, S.; Rafraf, M.; Somi, M.H.; Homayouni-Rad, A.; Asghari-Jafarabadi, M. Effects of probiotic yogurt consumption on metabolic factors in individuals with nonalcoholic fatty liver disease. J. Dairy Sci. 2014, 97, 7386–7393. [Google Scholar] [CrossRef]
- Kunnen, S.; Van, E.M. Lecithin: Cholesterol acyltransferase: Old friend or foe in atherosclerosis? J. Lipid Res. 2012, 53, 1783–1799. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Qiu, L.; Xu, X.; Liu, Z.; Zhan, H.; Tao, X.; Shah, N.P.; Wei, H. Beneficial effects of probiotic cholesterol-lowering strain of Enterococcus faecium WEFA23 from infants on diet-induced metabolic syndrome in rats. J. Dairy Sci. 2017, 100, 1618–1628. [Google Scholar] [CrossRef] [Green Version]
- Tsuru, H.; Osaka, M.; Hiraoka, Y.; Yoshida, M. HFD-induced hepatic lipid accumulation and inflammation are decreased in Factor D deficient mouse. Sci. Rep. 2020, 10, 17593. [Google Scholar] [CrossRef]
- Riezu-Boj, J.I.; Barajas, M.; Pérez-Sánchez, T.; Pajares, M.J.; Araña, M.; Milagro, F.I.; Urtasun, R. Lactiplantibacillus plantarum DSM20174 attenuates the progression of non-alcoholic fatty liver disease by modulating gut microbiota, improving metabolic risk factors, and attenuating adipose inflammation. Nutrients 2022, 14, 5212. [Google Scholar] [CrossRef]
- Kwoji, I.D.; Aiyegoro, O.A.; Okpeku, M.; Adeleke, M.A. Multi-strain probiotics: Synergy among isolates enhances biological activities. Biology 2021, 10, 322. [Google Scholar] [CrossRef]
- Wa, Y.; Yin, B.; He, Y.; Xi, W.; Huang, Y.; Wang, C.; Guo, F.; Gu, R. Effects of single probiotic- and combined probiotic-fermented milk on lipid metabolism in hyperlipidemic rats. Front. Microbiol. 2019, 10, 1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Sheraji, S.H.; Amin, I.; Azlan, A.; Manap, M.Y.; Hassan, F.A. Effects of Bifidobacterium longum BB536 on lipid profile and histopathological changes in hypercholesterolaemic rats. Benef. Microbes 2015, 6, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Jeun, J.; Kim, S.; Cho, S.Y.; Jun, H.J.; Park, H.J.; Seo, J.G.; Chung, M.J.; Lee, S.J. Hypocholesterolemic effects of Lactobacillus plantarum KCTC3928 by increased bile acid excretion in C57BL/6 mice. Nutrition 2010, 26, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Aron-Wisnewsky, J.; Warmbrunn, M.V.; Nieuwdorp, M.; Clement, K. Nonalcoholic fatty liver disease: Modulating gut microbiota to improve severity? J. Gastroenterol. 2020, 158, 1881–1898. [Google Scholar] [CrossRef] [PubMed]
- Abenavoli, L.; Scarpellini, E.; Colica, C.; Boccuto, L.; Salehi, B.; Sharifi-Rad, J.; Aiello, V.; Romano, B.; De Lorenzo, A.; Izzo, A.A.; et al. Gut microbiota and obesity: A role for probiotics. Nutrients 2019, 11, 2690. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Lin, S.; Vanhoutte, P.M.; Woo, C.W.; Xu, A. Akkermansia muciniphila protects against atherosclerosis by preventing metabolic endotoxemia-induced inflammation in Apoe−/− mice. Circulation 2016, 133, 2434–2446. [Google Scholar] [CrossRef] [Green Version]
- Jie, Z.; Xia, H.; Zhong, S.L.; Feng, Q.; Li, S.; Liang, S.; Zhong, H.; Liu, Z.; Gao, Y.; Zhao, H.; et al. The gut microbiome in atherosclerotic cardiovascular disease. Nat. Commun. 2017, 8, 845. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Xiao, S.; Gong, Z.; Zhu, X.; Yang, Q.; Li, Y.; Gao, S.; Dong, Y.; Shi, Z.; Wang, Y.; et al. Wuji wan formula ameliorates diarrhea and disordered colonic motility in post-inflammation irritable bowel syndrome rats by modulating the gut microbiota. Front. Microbiol. 2017, 8, 2307. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Body Weight Gain (g) | |||||||
|---|---|---|---|---|---|---|---|---|
| Week 1 | Week 2 | Week 3 | Week 4 | Week 5 | Week 6 | Week 7 | Week 8 | |
| Control | 61.60 ± 17.34 aA | 125.00 ± 19.70 aB | 159.80 ± 13.52 aC | 219.00 ± 13.93 aD | 260.20 ± 8.29 aE | 316.40 ± 13.99 aF | 348.20 ± 12.40 aG | 305.25 ± 13.79 aH |
| Probiotics | 69.80 ± 12.52 aA | 122.00 ± 9.00 aB | 162.20 ± 11.76 abC | 233.00 ± 15.05 abD | 271.00 ± 13.34 aE | 325.20 ± 8.20 aF | 352.00 ± 14.68 aF | 315.52 ± 19.24 aG |
| HCD | 61.40 ± 21.62 aA | 135.40 ± 17.95 aB | 183.40 ± 4.98 bC | 251.80 ± 9.52 bD | 292.40 ± 14.74 cE | 350.00 ± 11.77 bF | 393.00 ± 15.02 bG | 408.22 ± 20.14 cG |
| HCD-L | 76.60 ± 33.58 aA | 138.80 ± 26.95 aB | 179.80 ± 16.45 acB | 239.00 ± 26.43 abC | 286.60 ± 13.37 acD | 338.40 ± 15.01 abE | 370.40 ± 23.93 abE | 361.18 ± 8.85 bE |
| HCD-E | 60.80 ± 23.33 aA | 122.60 ± 16.20 aB | 161.60 ± 19.30 abC | 236.80 ± 4.82 abD | 269.60 ± 12.50 acD | 334.60 ± 6.19 abE | 356.40 ± 17.13 aE | 354.41 ± 21.25 bE |
| HCD-B | 70.00 ± 4.90 aA | 136.20 ± 22.16 aB | 174.60 ± 21.87 abC | 236.00 ± 19.94 abD | 268.20 ± 22.42 acD | 335.20 ± 14.96 abE | 354.20 ± 14.31 aE | 364.87 ± 15.18 bE |
| HCD-mix | 62.00 ± 16.63 aA | 122.80 ± 11.95 aB | 162.40 ± 19.36 abC | 238.00 ± 14.68 abD | 264.20 ± 7.12 abD | 329.70 ± 9.54 abE | 351.20 ± 15.22 aE | 333.99 ± 16.19 aE |
| Groups | Food Intake (g) | |||||||
|---|---|---|---|---|---|---|---|---|
| Week 1 | Week 2 | Week 3 | Week 4 | Week 5 | Week 6 | Week 7 | Week 8 | |
| Control | 143.68 ± 3.32 aA | 287.36 ± 6.63 aB | 431.05 ± 9.95 aC | 578.37 ± 17.84 aD | 728.91 ± 28.96 aE | 862.09 ± 19.89 aF | 1033.70 ± 59.78 aG | 1149.46 ± 26.52 aH |
| Probiotics | 145.98 ± 4.60 aA | 291.96 ± 9.20 aB | 437.93 ± 13.80 aC | 583.91 ± 18.41 aD | 729.89 ± 23.01 aE | 875.87 ± 27.61 aF | 1032.82 ± 27.82 aG | 1167.82 ± 36.81 aH |
| HCD | 148.01 ± 2.86 aA | 296.02 ± 5.73 aB | 465.32 ± 21.51 aC | 602.84 ± 28.07 aD | 753.69 ± 28.86 aE | 907.12 ± 35.08 aF | 1044.68 ± 22.47 aG | 1184.06 ± 22.90 aH |
| HCD-L | 146.73 ± 8.13 aA | 293.47 ± 16.25 aB | 440.20 ± 24.38 aC | 586.94 ± 32.50 aD | 733.67 ± 40.63 aE | 880.40 ± 48.76 aF | 1014.40 ± 51.44 aG | 1173.87 ± 65.01 aH |
| HCD-E | 144.45 ± 5.25 aA | 288.90 ± 10.50 aB | 433.36 ± 15.75 aC | 582.12 ± 23.33 aD | 722.26 ± 26.25 aE | 866.71 ± 31.50 aF | 1011.16 ± 36.75 aG | 1155.62 ± 42.01 aH |
| HCD-B | 145.40 ± 5.51 aA | 290.81 ± 11.02 aB | 436.21 ± 16.53 aC | 581.62 ± 22.04 aD | 729.33 ± 28.06 aE | 891.16 ± 33.51 aF | 1017.83 ± 38.58 aG | 1163.23 ± 44.09 aH |
| HCD-mix | 144.05 ± 6.39 aA | 288.09 ± 12.79 aB | 432.14 ± 19.18 aC | 578.03 ± 23.83 aD | 720.23 ± 31.97 aE | 864.28 ± 38.36 aF | 1011.56 ± 41.71 aG | 1152.37 ± 51.15 aH |
| Groups | Food Efficiency (%) | |||||||
|---|---|---|---|---|---|---|---|---|
| Week 1 | Week 2 | Week 3 | Week 4 | Week 5 | Week 6 | Week 7 | Week 8 | |
| Control | 42.88 ± 12.31 aA | 43.48 ± 6.68 aA | 37.04 ± 2.46 aAB | 37.85 ± 1.75 aA | 35.72 ± 0.99 aAB | 36.71 ± 1.57 aAB | 33.76 ± 2.09 aAB | 26.55 ± 0.75 aB |
| Probiotics | 47.97 ± 9.46 aA | 41.86 ± 3.89 aA | 37.08 ± 3.13 aB | 39.94 ± 2.97 aA | 37.14 ± 1.82 aB | 37.15 ± 1.07 aB | 34.13 ± 2.24 aBC | 27.03 ± 1.64 abC |
| HCD | 41.44 ± 14.48 aA | 45.73 ± 5.89 aA | 39.82 ± 4.68 aA | 41.79 ± 1.14 aA | 38.82 ± 1.94 aA | 38.60 ± 1.22 aA | 37.64 ± 1.74 aA | 34.48 ± 1.59 cA |
| HCD-L | 51.97 ± 22.15 aA | 43.25 ± 8.43 aAB | 38.94 ± 4.44 aAB | 40.52 ± 5.01 aAB | 37.14 ± 2.49 aAB | 38.26 ± 3.15 aAB | 36.58 ± 2.87 aAB | 30.87 ± 2.27 cdB |
| HCD-E | 41.88 ± 15.29 aA | 42.52 ± 6.17 aA | 37.42 ± 5.38 aA | 40.72 ± 1.40 aA | 37.36 ± 2.04 aA | 37.49 ± 1.46 aA | 35.30 ± 2.38 aA | 30.73 ± 2.64 bcdA |
| HCD-B | 48.19 ± 3.75 aA | 44.76 ± 6.50 aA | 40.08 ± 5.31 aB | 42.32 ± 3.73 aA | 37.87 ± 2.69 aBC | 37.74 ± 3.00 aBC | 34.83 ± 1.69 aBC | 31.40 ± 1.75 cdC |
| HCD-mix | 42.78 ± 10.03 aA | 42.70 ± 4.62 aA | 37.62 ± 4.56 aAB | 40.04 ± 3.33 aA | 38.13 ± 1.92 aAB | 37.90 ± 2.06 aAB | 35.56 ± 2.41 aAB | 29.02 ± 1.86 abdB |
| Groups. | Parameters | |||
|---|---|---|---|---|
| Abdominal Fat (g) | Abdominal Fat Index (mg/g Body Weight) | Liver Weight (g) | Liver Index (mg/g Body Weight) | |
| Control | 9.90 ± 0.56 a | 22.65 ± 1.53 a | 10.81 ± 0.72 a | 24.71 ± 1.60 a |
| Probiotics | 9.93 ± 0.92 a | 22.21 ± 2.87 a | 11.07 ± 0.97 a | 24.62 ± 1.46 a |
| HCD | 19.06 ± 2.30 b | 35.08 ± 4.89 b | 18.22 ± 1.01 b | 33.54 ± 2.92 b |
| HCD-L | 13.57 ± 1.92 c | 28.01 ± 4.66 ab | 14.31 ± 3.13 c | 29.45 ± 6.42 ab |
| HCD-E | 13.99 ± 1.91 c | 28.47 ± 3.59 ab | 14.35 ± 1.18 c | 29.25 ± 2.70 ab |
| HCD-B | 13.99 ± 1.24 c | 28.10 ± 2.31 ab | 14.40 ± 0.63 c | 28.95 ± 1.44 ab |
| HCD-mix | 11.89 ± 1.74 ac | 25.28 ± 3.79 a | 11.38 ± 1.85 ac | 24.14 ± 3.62 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puttarat, N.; Kasorn, A.; Vitheejongjaroen, P.; Chantarangkul, C.; Tangwattanachuleeporn, M.; Taweechotipatr, M. Beneficial Effects of Indigenous Probiotics in High-Cholesterol Diet-Induced Hypercholesterolemic Rats. Nutrients 2023, 15, 2710. https://0-doi-org.brum.beds.ac.uk/10.3390/nu15122710
Puttarat N, Kasorn A, Vitheejongjaroen P, Chantarangkul C, Tangwattanachuleeporn M, Taweechotipatr M. Beneficial Effects of Indigenous Probiotics in High-Cholesterol Diet-Induced Hypercholesterolemic Rats. Nutrients. 2023; 15(12):2710. https://0-doi-org.brum.beds.ac.uk/10.3390/nu15122710
Chicago/Turabian StylePuttarat, Narathip, Anongnard Kasorn, Porntipha Vitheejongjaroen, Chantanapa Chantarangkul, Marut Tangwattanachuleeporn, and Malai Taweechotipatr. 2023. "Beneficial Effects of Indigenous Probiotics in High-Cholesterol Diet-Induced Hypercholesterolemic Rats" Nutrients 15, no. 12: 2710. https://0-doi-org.brum.beds.ac.uk/10.3390/nu15122710