
Phytosterols and Cardiovascular Risk Evaluated against the Background of Phytosterolemia Cases—A German Expert Panel Statement †
 , , , , , , ,
, , , , , , , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
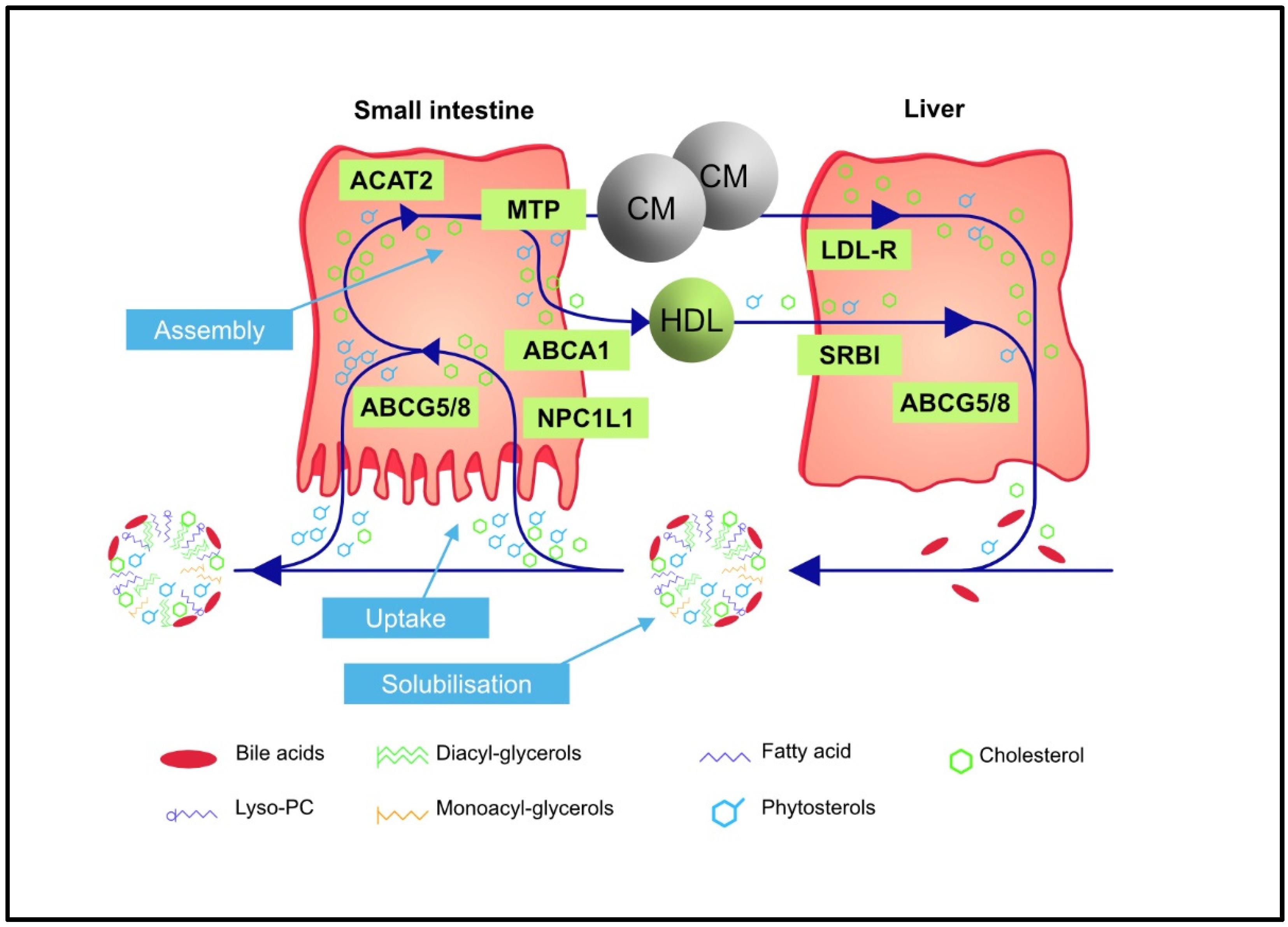
2. Regulation of Intestinal Absorption and Biliary Excretion of Phytosterols
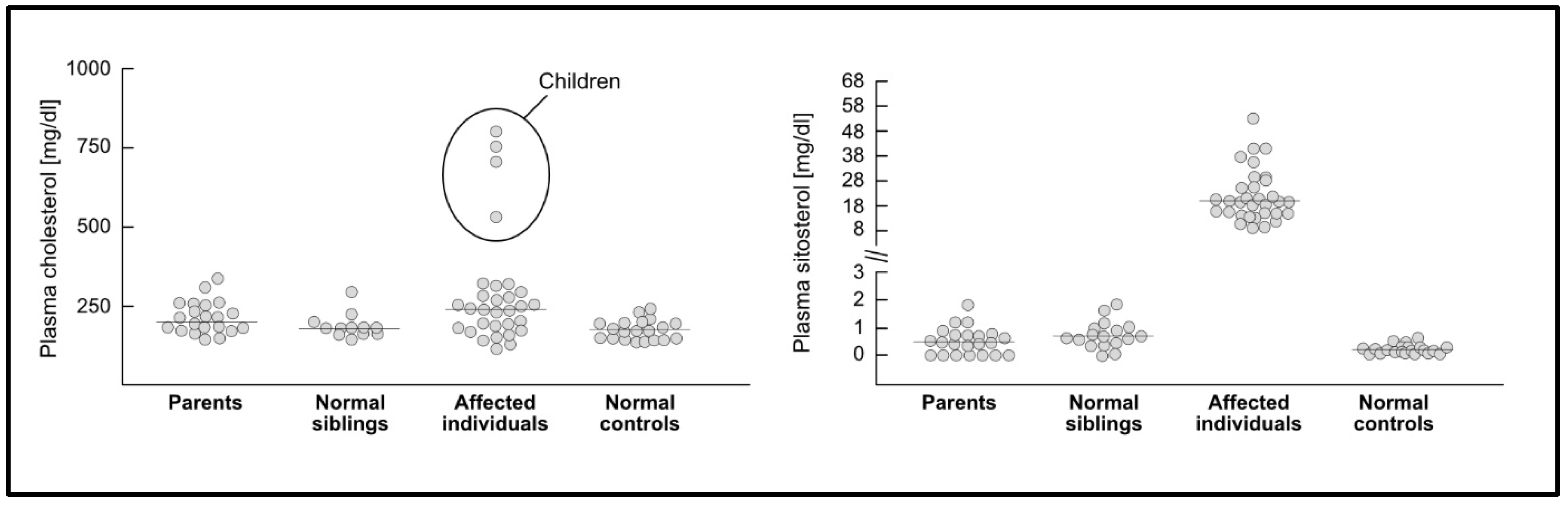
3. Phytosterolemia—An Inherited Disorder of Sterol Transport
3.1. Plasma Sterol Concentrations and Cardiovascular Disease in Phytosterolemia
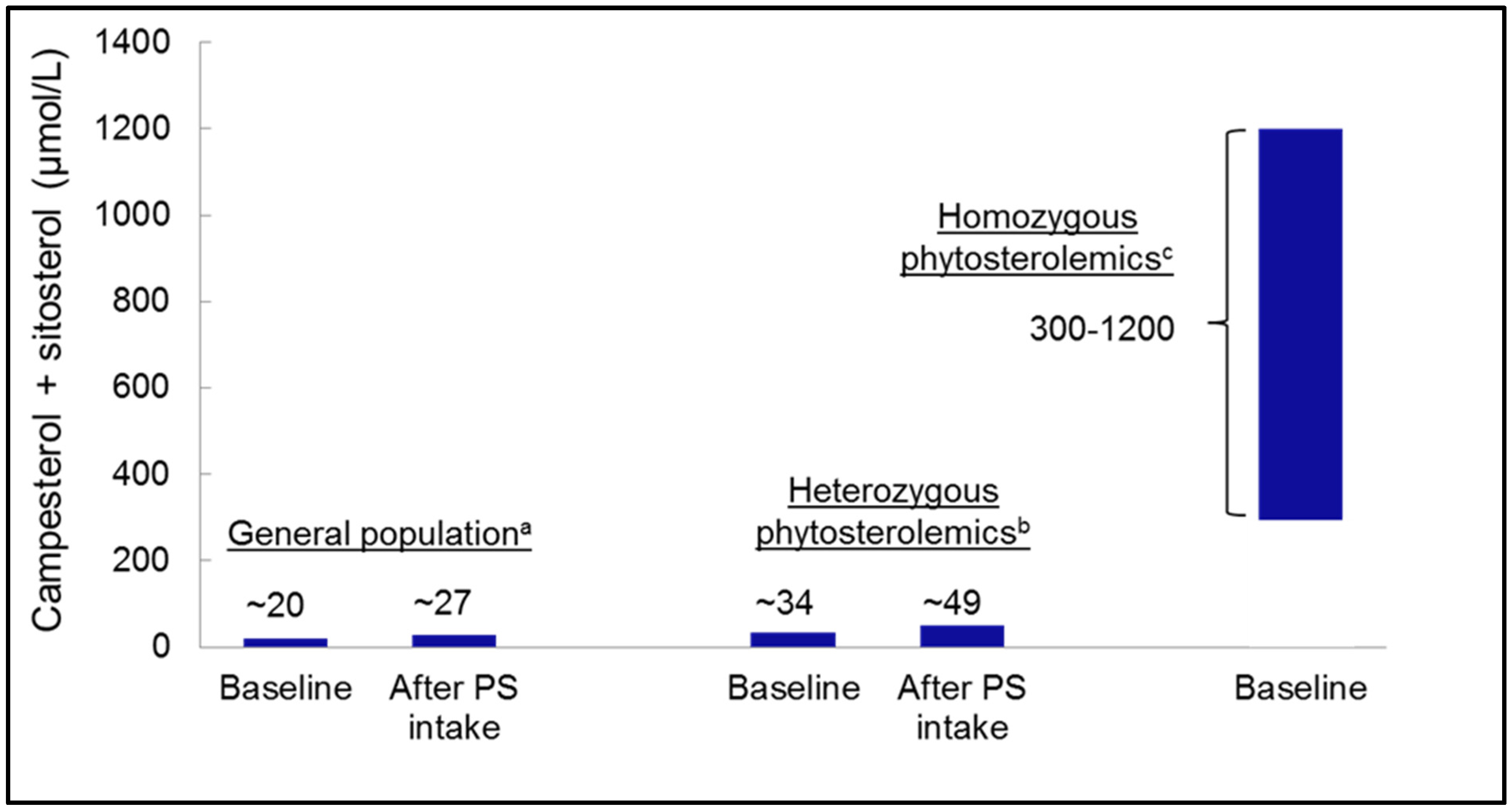
3.2. Heterozygous Phytosterolemia and Plasma Phytosterol Concentrations
3.3. Phytosterol Concentrations in Tissues and Risk of Atherosclerosis
3.4. Phytosterolemia to Evaluate Effects of Dietary Phytosterol Intake
3.5. Can Thrombocytopenia Be Used to Define a Threshold of Phytosterol Toxicity?
4. Phytosterols in Animal Models for Atherosclerosis
5. Are plasma Phytosterol Concentrations Associated with Cardiovascular Risk?
5.1. Evidence from Observational Studies
5.2. Evidence from Genetic Epidemiology
6. Do Phytosterol-Enriched Foods Increase the Risk of Atherosclerosis?
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tada, H.; Nohara, A.; Inazu, A.; Sakuma, N.; Mabuchi, H.; Kawashiri, M.A. Sitosterolaemia, Hypercholesterolemia, and Coronary Artery Disease. J. Atheroscler. Thromb. 2018, 25, 783–789. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.; Segard, A.; Graf, G.A. Sitosterolemia: Twenty Years of Discovery of the Function of ABCG5 ABCG8. Int. J. Mol. Sci. 2021, 22, 2641. [Google Scholar] [CrossRef]
- Gylling, H.; Plat, J.; Turley, S.; Ginsberg, H.N.; Ellegård, L.; Jessup, W.; Jones, P.J.; Lütjohann, D.; Maerz, W.; Masana, L.; et al. Plant sterols and plant stanols in the management of dyslipidaemia and prevention of cardiovascular disease. Atherosclerosis 2014, 232, 346–360. [Google Scholar] [CrossRef]
- Ference, B.A.; Ginsberg, H.N.; Graham, I.; Ray, K.K.; Packard, C.J.; Bruckert, E.; Hegele, R.A.; Krauss, R.M.; Raal, F.J.; Schunkert, H.; et al. Low density lipoproteins cause atherosclerotic cardiovascular disease. Evidence from genetic, epidemiologic, and clinical studies. A consensus statement from the European Atherosclerosis Society Consensus Panel. Eur. Heart J. 2017, 38, 2459–2472. [Google Scholar] [CrossRef] [PubMed]
- Hazard, S.E.; Patel, S.B. Sterolins ABCG5 and ABCG8: Regulators of whole body dietary sterols. Pflügers Arch. 2007, 453, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Salen, G.; Tint, G.S.; Shefer, S.; Shore, V.; Nguyen, L. Increased sitosterol absorption is offset by rapid elimination to prevent accumulation in heterozygotes with sitosterolemia. Arterioscler. Thromb. 1992, 12, 563–568. [Google Scholar] [CrossRef] [PubMed]
- Calpe-Berdiel, L.; Escolà-Gil, J.C.; Blanco-Vaca, F. New insights into the molecular actions of plant sterols and stanols in cholesterol metabolism. Atherosclerosis 2009, 203, 18–31. [Google Scholar] [CrossRef]
- Davis, H.R., Jr.; Zhu, L.J.; Hoos, L.M.; Tetzloff, G.; Maguire, M.; Liu, J.; Yao, X.; Iyer, S.P.; Lam, M.H.; Lund, E.G.; et al. Niemann-Pick C1 Like 1 (NPC1L1) is the intestinal phytosterol and cholesterol transporter and a key modulator of whole-body cholesterol homeostasis. J. Biol. Chem. 2004, 279, 33586–33592. [Google Scholar] [CrossRef]
- Yamanashi, Y.; Takada, T.; Suzuki, H. Niemann-Pick C1-like 1 overexpression facilitates ezetimibe-sensitive cholesterol and β-sitosterol uptake in CaCo-2 cells. J. Pharmacol. Exp. Ther. 2007, 320, 559–564. [Google Scholar] [CrossRef]
- Lütjohann, D.; Bjorkhem, I.; Beil, U.F.; von Bergmann, K. Sterol absorption and sterol balance in phytosterolemia evaluated by deuterium-labeled sterols: Effect of sitostanol treatment. J. Lipid Res. 1995, 36, 1763–1773. [Google Scholar] [CrossRef]
- Temel, R.E.; Gebre, A.K.; Parks, J.S.; Rudel, L.L. Compared with acyl- CoA:cholesterol O-acyltransferase (ACAT) 1 and lecithin:cholesterol acyltransferase, ACAT2 displays the greatest capacity to differentiate cholesterol from sitosterol. J. Biol. Chem. 2003, 278, 47594–47601. [Google Scholar] [CrossRef]
- Nguyen, T.M.; Sawyer, J.K.; Kelley, K.L.; Davis, M.A.; Kent, C.R.; Rudel, L.L. ACAT2 and ABCG5/G8 are both required for efficient cholesterol absorption in mice: Evidence from thoracic lymph duct cannulation. J. Lipid Res. 2012, 53, 1598–1609. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; von Bergmann, K.; Lütjohann, D.; Hobbs, H.H.; Cohen, J.C. Selective sterol accumulation in ABCG5/ABCG8-deficient mice. J. Lipid Res. 2004, 45, 301–307. [Google Scholar] [CrossRef]
- Othman, R.A.; Myrie, S.B.; Jones, P.J. Non-cholesterol sterols and cholesterol metabolism in sitosterolemia. Atherosclerosis 2013, 231, 291–299. [Google Scholar] [CrossRef]
- Bosner, M.S.; Lange, L.G.; Stenson, W.F.; Ostlund, R.E., Jr. Percent cholesterol absorption in normal women and men quantified with dual stable isotopic tracers and negative ion mass spectrometry. J. Lipid Res. 1999, 40, 302–308. [Google Scholar] [CrossRef]
- Chan, Y.M.; Varady, K.A.; Lin, Y.; Trautwein, E.; Mensink, R.P.; Plat, J.; Jones, P.J. Plasma concentrations of plant sterols: Physiology and relationship with coronary heart disease. Nutr. Rev. 2006, 64, 385–402. [Google Scholar] [CrossRef]
- Temel, R.E.; Tang, W.; Ma, Y.; Rudel, L.L.; Willingham, M.C.; Ioannou, Y.A.; Davies, J.P.; Nilsson, L.M.; Yu, L. Hepatic Niemann-Pick C1-like 1 regulates biliary cholesterol concentration and is a target of ezetimibe. J. Clin. Investig. 2007, 117, 1968–1978. [Google Scholar] [CrossRef] [PubMed]
- Myrie, S.B.; Mymin, D.; Triggs-Raine, B.; Jones, P.J. Serum lipids, plant sterols, and cholesterol kinetic responses to plant sterol supplementation in phytosterolemia heterozygotes and control individuals. Am. J. Clin. Nutr. 2012, 95, 837–844. [Google Scholar] [CrossRef]
- Escolà-Gil, J.C.; Quesada, H.; Julve, J.; Martín-Campos, J.M.; Cedó, L.; Blanco-Vaca, F. Sitosterolemia: Diagnosis, investigation, and management. Curr. Atheroscler. Rep. 2014, 16, 424. [Google Scholar] [CrossRef] [PubMed]
- Sudhop, T.; von Bergmann, K. Sitosterolemia—A rare disease. Are Elev. Plant Sterols Addit. Risk Factor? Z. Kardiol. 2004, 93, 921–928. [Google Scholar] [CrossRef]
- Jessup, W.; Herman, W.; Chapman, M.J. Phytosterols in cardiovascular disease: Innocuous dietary components, or accelerators of atherosclerosis? Future Lipidol. 2008, 3, 301–310. [Google Scholar] [CrossRef]
- Bhattacharyya, A.K.; Connor, W.E. Beta-sitosterolemia and xanthomatosis. A newly described lipid storage disease in two sisters. J. Clin. Investig. 1974, 53, 1033–1043. [Google Scholar] [CrossRef]
- Simonen, P.P.; Gylling, H.; Miettinen, T.A. The distribution of squalene and non-cholesterol sterols in lipoproteins in type 2 diabetes. Atherosclerosis 2007, 194, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Gylling, H.; Simonen, P. Phytosterols, phytostanols, and lipoprotein metabolism. Nutrients 2015, 7, 7965–7977. [Google Scholar] [CrossRef] [PubMed]
- Robins, S.J.; Fasulo, J.M. High density lipoproteins, but not other lipoproteins, provide a vehicle for sterol transport to bile. J. Clin. Investig. 1997, 99, 380–384. [Google Scholar] [CrossRef]
- Silbernagel, G.; Baumgartner, I.; Wanner, C.; März, W. Toward individualized cholesterol-lowering treatment in end-stage renal disease. J. Ren. Nutr. 2014, 24, 65–71. [Google Scholar] [CrossRef]
- Patel, S.B.; Honda, A.; Salen, G. Sitosterolemia: Exclusion of genes involved in reduced cholesterol biosynthesis. J. Lipid Res. 1998, 39, 1055–1056. [Google Scholar] [CrossRef]
- Patel, S.B.; Salen, G.; Hidaka, H.; Kwiterovich, P.O.; Stalenhoef, A.F.; Miettinen, T.A.; Grundy, S.M.; Lee, M.H.; Rubenstein, J.S.; Polymeropoulos, M.H.; et al. Mapping a gene involved in regulating dietary cholesterol absorption. The sitosterolemia locus is found at chromosome 2p21. J. Clin. Investig. 1998, 102, 1041–1044. [Google Scholar] [CrossRef] [PubMed]
- Berge, K.E.; Tian, H.; Graf, G.A.; Yu, L.; Grishin, N.V.; Schultz, J.; Kwiterovich, P.; Shan, B.; Barnes, R.; Hobbs, H.H. Accumulation of dietary cholesterol in sitosterolemia caused by mutations in adjacent ABC transporters. Science 2000, 290, 1771–1775. [Google Scholar] [CrossRef]
- Lee, M.H.; Gordon, D.; Ott, J.; Lu, K.; Ose, L.; Miettinen, T.; Gylling, H.; Stalenhoef, A.F.; Pandya, A.; Hidaka, H.; et al. Fine mapping of a gene responsible for regulating dietary cholesterol absorption; founder effects underlie cases of phytosterolaemia in multiple communities. Eur. J. Hum. Genet. 2001, 9, 375–384. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.H.; Lu, K.; Hazard, S.; Yu, H.; Shulenin, S.; Hidaka, H.; Kojima, H.; Allikmets, R.; Sakuma, N.; Pegoraro, R.; et al. Identification of a gene, ABCG5, important in the regulation of dietary cholesterol absorption. Nat. Genet. 2001, 27, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.; Lee, M.H.; Hazard, S.; Brooks-Wilson, A.; Hidaka, H.; Kojima, H.; Ose, L.; Stalenhoef, A.F.; Mietinnen, T.; Bjorkhem, I.; et al. Two genes that map to the STSL locus cause sitosterolemia: Genomic structure and spectrum of mutations involving sterolin-1 and sterolin-2, encoded by ABCG5 and ABCG8, respectively. Am. J. Hum. Genet. 2001, 69, 278–290. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.B.; Salen, G. Sitosterolemia: Xenophobia for the Body. In Evidence-Based Management of Lipid Disorders; Vissers, M.N., Kastelein, J.J.P., Stroes, E.S., Eds.; TFM Publishing: Harley, UK, 2010; pp. 217–230. [Google Scholar]
- Bjorkhem, I.; Danielsson, H. Biosynthesis and metabolism of bile acids in man. Prog. Liver Dis. 1976, 5, 215–231. [Google Scholar] [PubMed]
- Bjorkhem, I. Mechanism of degradation of the steroid side chain in the formation of bile acids. J. Lipid Res. 1992, 33, 455–471. [Google Scholar] [CrossRef]
- Calandra, S.; Tarugi, P.; Speedy, H.E.; Dean, A.F.; Bertolini, S.; Shoulders, C.C. Mechanisms and genetic determinants regulating sterol absorption, circulating LDL levels, and sterol elimination: Implications for classification and disease risk. J. Lipid Res. 2011, 52, 1885–1926. [Google Scholar] [CrossRef]
- Drozdowski, L.A.; Clandinin, T.; Thomson, A.B. Ontogeny, growth and development of the small intestine: Understanding pediatric gastroenterology. World J. Gastroenterol. 2010, 16, 787–799. [Google Scholar]
- Rios, J.; Stein, E.; Shendure, J.; Hobbs, H.H.; Cohen, J.C. Identification by whole-genome resequencing of gene defect responsible for severe hypercholesterolemia. Hum. Mol. Genet. 2010, 19, 4313–4318. [Google Scholar] [CrossRef]
- Belamarich, P.F.; Deckelbaum, R.J.; Starc, T.J.; Dobrin, B.E.; Tint, G.S.; Salen, G. Response to diet and cholestyramine in a patient with sitosterolemia. Pediatrics 1990, 86, 977–981. [Google Scholar] [CrossRef]
- Yoshida, A.; Naito, M.; Miyazaki, K. Japanese sisters associated with pseudohomozygous familial hypercholesterolemia and sitosterolemia. J. Atheroscler. Thromb. 2000, 7, 33–38. [Google Scholar] [CrossRef]
- Rajaratnam, R.A.; Gylling, H.; Miettinen, T.A. Independent association of serum squalene and noncholesterol sterols with coronary artery disease in postmenopausal women. J. Am. Coll. Cardiol. 2000, 35, 1185–1191. [Google Scholar] [CrossRef]
- Tsubakio-Yamamoto, K.; Nishida, M.; Nakagawa-Toyama, Y.; Masuda, D.; Ohama, T.; Yamashita, S. Current therapy for patients with sitosterolemia--effect of ezetimibe on plant sterol metabolism. J. Atheroscler. Thromb. 2010, 17, 891–900. [Google Scholar] [CrossRef] [Green Version]
- Shulman, R.S.; Bhattacharyya, A.K.; Connor, W.E.; Fredrickson, D.S. Beta-sitosterolemia and xanthomatosis. N. Engl. J. Med. 1976, 294, 482–483. [Google Scholar] [CrossRef] [PubMed]
- Khachadurian, A.K.; Clancy, K.F. Familial phytosterolemia (beta-sitosterolemia): Report of five cases and studies in cultured skin fibroblasts. Clin. Res. 1978, 26, 379A. [Google Scholar]
- Miettinen, T.A. Phytosterolaemia, xanthomatosis and premature atherosclerotic arterial disease: A case with high plant sterol absorption, impaired sterol elimination and low cholesterol synthesis. Eur. J. Clin. Investig. 1980, 10, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Lin, H.J.; Chan, T.K.; Salen, G.; Chan, W.C.; Tse, T.F. A unique patient with coexisting cerebrotendinous xanthomatosis and beta-sitosterolemia. Am. J. Med. 1981, 71, 313–319. [Google Scholar] [CrossRef]
- Salen, G.; Horak, I.; Rothkopf, M.; Cohen, J.L.; Speck, J.; Tint, G.S.; Shore, V.; Dayal, B.; Chen, T.; Shefer, S. Lethal atherosclerosis associated with abnormal plasma and tissue sterol composition in sitosterolemia with xanthomatosis. J. Lipid Res. 1985, 26, 1126–1133. [Google Scholar] [CrossRef] [PubMed]
- Berger, G.M.; Deppe, W.M.; Marais, A.D.; Biggs, M. Phytosterolaemia in three unrelated South African families. Postgrad. Med. J. 1994, 70, 631–637. [Google Scholar] [CrossRef]
- Rees, D.C.; Iolascon, A.; Carella, M.; O’marcaigh, A.S.; Kendra, J.R.; Jowitt, S.N.; Wales, J.K.; Vora, A.; Makris, M.; Manning, N.; et al. Stomatocytic haemolysis and macrothrombocytopenia (Mediterranean stomatocytosis/macrothrombocytopenia) is the haematological presentation of phytosterolaemia. Br. J. Haematol. 2005, 130, 297–309. [Google Scholar] [CrossRef]
- Kruit, J.K.; Drayer, A.L.; Bloks, V.W.; Blom, N.; Olthof, S.G.; Sauer, P.J.; de Haan, G.; Kema, I.P.; Vellenga, E.; Kuipers, F. Plant sterols cause macrothrombocytopenia in a mouse model of sitosterolemia. J. Biol. Chem. 2008, 283, 6281–6287. [Google Scholar] [CrossRef]
- Neff, A.T. Sitosterolemia’s stomatocytosis and macrothrombocytopenia. Blood 2012, 120, 4283. [Google Scholar] [CrossRef]
- Kanaji, T.; Kanaji, S.; Montgomery, R.R.; Patel, S.B.; Newman, P.J. Platelet hyperreactivity explains the bleeding abnormality and macrothrombocytopenia in a murine model of sitosterolemia. Blood 2013, 122, 2732–2742. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Cao, L.; Su, Y.; Wang, G.; Wang, R.; Yu, Z.; Bai, X.; Ruan, C. Specific macrothrombocytopenia/hemolytic anemia associated with sitosterolemia. Am. J. Hematol. 2014, 89, 320–324. [Google Scholar] [CrossRef]
- Wadsack, D.; Sattler, H.; Kleber, M.E.; Eirich, K.; Scharnagl, H.; Fauler, G.; März, W. Grammer TBRecurrent tendosynovitis as a rare manifestation of a lipid disorder. Clin. Lipidol. 2019, 13, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Kwiterovich, P.O.; Bachorik, P.S., Jr.; Smith, H.H.; McKusick, V.A.; Connor, W.E.; Teng, B.; Sniderman, A.D. Hyperapobetalipoproteinaemia in two families with xanthomas and phytosterolaemia. Lancet 1981, 1, 466–469. [Google Scholar] [CrossRef] [PubMed]
- Hansel, B.; Carrié, A.; Brun-Druc, N.; Leclert, G.; Chantepie, S.; Coiffard, A.S.; Kahn, J.F.; Chapman, M.J.; Bruckert, E. Premature atherosclerosis is not systematic in phytosterolemic patients: Severe hypercholesterolemia as a confounding factor in five subjects. Atherosclerosis 2014, 234, 162–168. [Google Scholar] [CrossRef]
- Morganroth, J.; Levy, R.I.; McMahon, A.E.; Gotto, A.M., Jr. Pseudohomozygous type II hyperlipoproteinemia. J. Pediatr. 1974, 85, 639–643. [Google Scholar] [CrossRef]
- Park, J.H.; Chung, I.H.; Kim, D.H.; Choi, M.H.; Garg, A.; Yoo, E.G. Sitosterolemia presenting with severe hypercholesterolemia and intertriginous xanthomas in a breastfed infant: Case report and brief review. J. Clin. Endocrinol. Metab. 2014, 99, 1512–1518. [Google Scholar] [CrossRef]
- Niu, D.M.; Chong, K.W.; Hsu, J.H.; Wu, T.J.; Yu, H.C.; Huang, C.H.; Lo, M.Y.; Kwok, C.F.; Kratz, L.E.; Ho, L.T. Clinical observations, molecular genetic analysis, and treatment of sitosterolemia in infants and children. J. Inherit. Metab. Dis. 2010, 33, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.D.; Thompson, P.D. Phytosterols and vascular disease. Atherosclerosis 2006, 186, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Kwiterovich, P.O.; Chen, S.C., Jr.; Virgil, D.G.; Schweitzer, A.; Arnold, D.R.; Kratz, L.E. Response of obligate heterozygotes for phytosterolemia to a low-fat diet and to a plant sterol ester dietary challenge. J. Lipid Res. 2003, 44, 1143–1155. [Google Scholar] [CrossRef]
- Stalenhoef, A.F.; Hectors, M.; Demacker, P.N. Effect of plant sterol-enriched margarine on plasma lipids and sterols in subjects heterozygous for phytosterolaemia. J. Intern. Med. 2001, 249, 163–166. [Google Scholar] [CrossRef]
- Zhao, H.L.; Houweling, A.H.; Vanstone, C.A.; Jew, S.; Trautwein, E.A.; Duchateau, G.S.; Jones, P.J. Genetic variation in ABC G5/G8 and NPC1L1 impact cholesterol response to plant sterols in hypercholesterolemic men. Lipids 2008, 43, 1155–1164. [Google Scholar] [CrossRef]
- Fuentes, F.; Lopez-Miranda, J.; Garcia, A.; Perez-Martinez, P.; Moreno, J.; Cofan, M.; Caballero, J.; Paniagua, J.A.; Ros, E.; Perez-Jimenez, F. Basal plasma concentrations of plant sterols can predict LDL-C response to sitosterol in patients with familial hypercholesterolemia. Eur. J. Clin. Nutr. 2008, 62, 495–501. [Google Scholar] [CrossRef]
- Dumolt, J.H.; Rideout, T.C. The Lipid-Lowering Effects and Associated Mechanisms of Dietary Phytosterol Supplementation. Curr. Pharm. Des. 2017, 23, 5077–5085. [Google Scholar] [CrossRef]
- Nomura, A.; Emdin, C.A.; Won, H.H.; Peloso, G.M.; Natarajan, P.; Ardissino, D.; Danesh, J.; Schunkert, H.; Correa, A.; Bown, M.J.; et al. Heterozygous ABCG5 Gene Deficiency and Risk of Coronary Artery Disease. Circ. Genom. Precis. Med. 2020, 13, 417–423. [Google Scholar] [CrossRef]
- Helske, S.; Miettinen, T.; Gylling, H.; Mäyränpää, M.; Lommi, J.; Turto, H.; Werkkala, K.; Kupari, M.; Kovanen, P.T. Accumulation of cholesterol precursors and plant sterols in human stenotic aortic valves. J. Lipid Res. 2008, 49, 1511–1518. [Google Scholar] [CrossRef] [PubMed]
- Vanmierlo, T.; Weingärtner, O.; van der Pol, S.; Husche, C.; Kerksiek, A.; Friedrichs, S.; Sijbrands, E.; Steinbusch, H.; Grimm, M.; Hartmann, T.; et al. Dietary intake of plants sterols stably increases plant sterols in the murine brain. J. Lipid Res. 2012, 53, 726–735. [Google Scholar] [CrossRef]
- Miettinen, T.A.; Nissinen, M.; Lepäntalo, M.; Albäck, A.; Railo, M.; Vikatmaa, P.; Kaste, M.; Mustanoja, S.; Gylling, H. Non-cholesterol sterols in serum and endarterectomized carotid arteries after a short-term plant stanol and sterol ester challenge. Nutr. Metab. Cardiovasc. Dis. 2011, 21, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Horenstein, R.B.; Braxton, D.M.; Post, W.S.; Lütjohann, D.; von Bergmann, K.; Ryan, K.A.; Terrin, M.; Shuldiner, A.R.; Steinle, N.I. The ABCG8 G574R Variant, Serum Plant Sterol Levels, and Cardiovascular Disease Risk in the Old Order Amish. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 413–419. [Google Scholar] [CrossRef]
- Miettinen, T.A.; Klett, E.L.; Gylling, H.; Isoniemi, H.; Patel, S.B. Liver transplantation in a patient with sitosterolemia and cirrhosis. Gastroenterology 2006, 130, 542–547. [Google Scholar] [CrossRef] [PubMed]
- Kidambi, S.; Patel, S.B. Sitosterolaemia: Pathophysiology, clinical presentation and laboratory diagnosis. J. Clin. Pathol. 2008, 61, 588–594. [Google Scholar] [CrossRef] [PubMed]
- Hidaka, H.; Nakamura, T.; Aoki, T.; Kojima, H.; Nakajima, Y.; Kosugi, K.; Hatanaka, I.; Harada, M.; Kobayashi, M.; Tamura, A.; et al. Increased plasma plant sterol levels in heterozygotes with sitosterolemia and xanthomatosis. J. Lipid Res. 1990, 31, 881–888. [Google Scholar] [CrossRef]
- Hatanaka, I.; Yasuda, H.; Hidaka, H.; Harada, N.; Kobayashi, M.; Okabe, H.; Matsumoto, K.; Hukuda, S.; Shigeta, Y. Spinal cord compression with paraplegia in xanthomatosis due to normocholesterolemic sitosterolemia. Ann. Neurol. 1990, 28, 390–393. [Google Scholar] [CrossRef]
- Lütjohann, D.; von Bergmann, K. Phytosterolaemia: Diagnosis, characterization and therapeutical approaches. Ann. Med. 1997, 29, 181–184. [Google Scholar] [CrossRef]
- Ratnayake, W.M.; L’Abbe, M.R.; Mueller, R.; Hayward, S.; Plouffe, L.; Hollywood, R.; Trick, K. Vegetable oils high in phytosterols make erythrocytes less deformable and shorten the life span of stroke-prone spontaneously hypertensive rats. J. Nutr. 2000, 130, 1166–1178. [Google Scholar] [CrossRef]
- Yu, L.; Hammer, R.E.; Li-Hawkins, J.; Von Bergmann, K.; Lütjohann, D.; Cohen, J.C.; Hobbs, H.H. Disruption of Abcg5 and Abcg8 in mice reveals their crucial role in biliary cholesterol secretion. Proc. Natl. Acad. Sci. USA 2002, 99, 16237–16242. [Google Scholar] [CrossRef]
- Klett, E.L.; Lu, K.; Kosters, A.; Vink, E.; Lee, M.H.; Altenburg, M.; Shefer, S.; Batta, A.K.; Yu, H.; Chen, J.; et al. A mouse model of sitosterolemia: Absence of Abcg8/sterolin-2 results in failure to secrete biliary cholesterol. BMC Med. 2004, 2, 5. [Google Scholar] [CrossRef]
- Plösch, T.; Bloks, V.W.; Terasawa, Y.; Berdy, S.; Siegler, K.; Van Der Sluijs, F.; Kema, I.P.; Groen, A.K.; Shan, B.; Kuipers, F.; et al. Sitosterolemia in ABC-transporter G5-deficient mice is aggravated on activation of the liver-X receptor. Gastroenterology 2004, 126, 290–300. [Google Scholar] [CrossRef]
- Su, Y.; Wang, Z.; Yang, H.; Cao, L.; Liu, F.; Bai, X.; Ruan, C. Clinical and molecular genetic analysis of a family with sitosterolemia and co-existing erythrocyte and platelet abnormalities. Haematologica 2006, 91, 1392–1395. [Google Scholar] [PubMed]
- Solca, C.; Tint, G.S.; Patel, S.B. Dietary xenosterols lead to infertility and loss of abdominal adipose tissue in sterolin-deficient mice. J. Lipid Res. 2013, 54, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.B. Recent advances in understanding the STSL locus and ABCG5/ABCG8 biology. Curr. Opin. Lipidol. 2014, 25, 169–175. [Google Scholar] [CrossRef]
- McDaniel, A.L.; Alger, H.M.; Sawyer, J.K.; Kelley, K.L.; Kock, N.D.; Brown, J.M.; Temel, R.E.; Rudel, L.L. Phytosterol feeding causes toxicity in ABCG5/G8 knockout mice. Am. J. Pathol. 2013, 182, 1131–1138. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Mitsche, M.A.; Lütjohann, D.; Cohen, J.C.; Xie, X.S.; Hobbs, H.H. Relative roles of ABCG5/ABCG8 in liver and intestine. J. Lipid Res. 2015, 56, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Othman, R.A.; Myrie, S.B.; Mymin, D.; Merkens, L.S.; Roullet, J.B.; Steiner, R.D.; Jones, P.J. Ezetimibe reduces plant sterol accumulation and favorably increases platelet count in sitosterolemia. J. Pediatr. 2015, 166, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Salen, G.; Starc, T.; Sisk, C.M.; Patel, S.B. Intestinal cholesterol absorption inhibitor ezetimibe added to cholestyramine for sitosterolemia and xanthomatosis. Gastroenterology 2006, 130, 1853–1857. [Google Scholar] [CrossRef]
- Hung, C.N.; Lee, C.Y. A case of sitosterolaemia with stomatocyticanaemia and thrombocytopenia treated with Ezetimibe with good response. Int. J. Pediatr. Endocrinol. 2013, 2013 (Suppl. S1), 174. [Google Scholar] [CrossRef]
- de Jong, A.; Plat, J.; Mensink, R.P. Plant sterol or stanol consumption does not affect erythrocyte osmotic fragility in patients on statin treatment. Eur. J. Clin. Nutr. 2006, 60, 985–990. [Google Scholar] [CrossRef] [PubMed]
- Kritchevsky, D.; Chen, S.C. Phytosterols-health benefits and potential concerns: A review. J. Nutr. Res. 2005, 25, 413–428. [Google Scholar] [CrossRef]
- Wilund, K.R.; Yu, L.; Xu, F.; Vega, G.L.; Grundy, S.M.; Cohen, J.C.; Hobbs, H.H. No association between plasma levels of plant sterols and atherosclerosis in mice and men. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 2326–2332. [Google Scholar] [CrossRef] [PubMed]
- Plat, J.; Beugels, I.; Gijbels, M.J.; de Winther, M.P.; Mensink, R.P. Plant sterol or stanol esters retard lesion formation in LDL receptor-deficient mice independent of changes in serum plant sterols. J. Lipid Res. 2006, 47, 2762–2771. [Google Scholar] [CrossRef]
- Weingärtner, O.; Lütjohann, D.; Ji, S.; Weisshoff, N.; List, F.; Sudhop, T.; von Bergmann, K.; Gertz, K.; König, J.; Schäfers, H.J.; et al. Vascular effects of diet supplementation with plant sterols. J. Am. Coll. Cardiol. 2008, 51, 1553–1561. [Google Scholar] [CrossRef]
- Bombo, R.P.; Afonso, M.S.; Machado, R.M.; Lavrador, M.S.; Nunes, V.S.; Quintão, E.R.; Koike, M.; Catanozi, S.; Lin, C.J.; Nakandakare, E.R.; et al. Dietary phytosterol does not accumulate in the arterial wall and prevents atherosclerosis of LDLr-KO mice. Atherosclerosis 2013, 231, 442–447. [Google Scholar] [CrossRef]
- Cannon, C.P.; Blazing, M.A.; Giugliano, R.P.; McCagg, A.; White, J.A.; Theroux, P.; Darius, H.; Lewis, B.S.; Ophuis, T.O.; Jukema, J.W.; et al. Ezetimibe added to statin therapy after acute coronary syndromes. N. Engl. J. Med. 2015, 372, 2387–2397. [Google Scholar] [CrossRef] [PubMed]
- Hibi, K.; Sonoda, S.; Kawasaki, M.; Otsuji, Y.; Murohara, T.; Ishii, H.; Sato, K.; Koshida, R.; Ozaki, Y.; Sata, M.; et al. Effects of Ezetimibe-Statin Combination Therapy on Coronary Atherosclerosis in Acute Coronary Syndrome. Circ. J. 2017, 82, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Getz, G.S.; Reardon, C.A. Animal models of atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 1104–1115. [Google Scholar] [CrossRef] [PubMed]
- Wouters, K.; Shiri-Sverdlov, R.; van Gorp, P.J.; van Bilsen, M.; Hofker, M.H. Understanding hyperlipidemia and atherosclerosis: Lessons from genetically modified apoe and ldlr mice. Clin. Chem. Lab. Med. 2005, 43, 470–479. [Google Scholar] [CrossRef] [PubMed]
- Assmann, G.; Cullen, P.; Erbey, J.; Ramey, D.R.; Kannenberg, F.; Schulte, H. Plasma sitosterol elevations are associated with an increased incidence of coronary events in men: Results of a nested case-control analysis of the Prospective Cardiovascular Münster (PROCAM) study. Nutr. Metab. Cardiovasc. Dis. 2006, 16, 13–21. [Google Scholar] [CrossRef]
- Fassbender, K.; Lütjohann, D.; Dik, M.G.; Bremmer, M.; König, J.; Walter, S.; Liu, Y.; Letièmbre, M.; von Bergmann, K.; Jonker, C. Moderately elevated plant sterol levels are associated with reduced cardiovascular risk--the LASA study. Atherosclerosis 2008, 196, 283–288. [Google Scholar] [CrossRef]
- Windler, E.; Zyriax, B.C.; Kuipers, F.; Linseisen, J.; Boeing, H. Association of plasma phytosterol concentrations with incident coronary heart disease. Data from the CORA study, a case-control study of coronary artery disease in women. Atherosclerosis 2009, 203, 284–290. [Google Scholar] [CrossRef]
- Strandberg, T.E.; Gylling, H.; Tilvis, R.S.; Miettinen, T.A. Serum plant and other noncholesterol sterols, cholesterol metabolism and 22-year mortality among middle-aged men. Atherosclerosis 2010, 210, 282–287. [Google Scholar] [CrossRef]
- Escurriol, V.; Cofán, M.; Moreno-Iribas, C.; Larrañaga, N.; Martínez, C.; Navarro, C.; Rodríguez, L.; González, C.A.; Corella, D.; Ros, E. Phytosterol plasma concentrations and coronary heart disease in the prospective Spanish EPIC cohort. J. Lipid Res. 2010, 51, 618–624. [Google Scholar] [CrossRef] [PubMed]
- Weingärtner, O.; Pinsdorf, T.; Rogacev, K.S.; Blömer, L.; Grenner, Y.; Gräber, S.; Ulrich, C.; Girndt, M.; Böhm, M.; Fliser, D.; et al. The relationships of markers of cholesterol homeostasis with carotid intima-media thickness. PLoS ONE 2010, 5, e13467. [Google Scholar] [CrossRef]
- Silbernagel, G.; Fauler, G.; Renner, W.; Landl, E.M.; Hoffmann, M.M.; Winkelmann, B.R.; Boehm, B.O.; März, W. The relationships of cholesterol metabolism and plasma plant sterols with the severity of coronary artery disease. J. Lipid Res. 2009, 50, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Silbernagel, G.; Fauler, G.; Hoffmann, M.M.; Lütjohann, D.; Winkelmann, B.R.; Boehm, B.O.; März, W. The associations of cholesterol metabolism and plasma plant sterols with all-cause and cardiovascular mortality. J. Lipid Res. 2010, 51, 2384–2393. [Google Scholar] [CrossRef]
- Matthan, N.R.; Pencina, M.; LaRocque, J.M.; Jacques, P.F.; D’Agostino, R.B.; Schaefer, E.J.; Lichtenstein, A.H. Alterations in cholesterol absorption/synthesis markers characterize Framingham offspring study participants with CHD. J. Lipid Res. 2009, 50, 1927–1935. [Google Scholar] [CrossRef]
- Genser, B.; Silbernagel, G.; De Backer, G.; Bruckert, E.; Carmena, R.; Chapman, M.J.; Deanfield, J.; Descamps, O.S.; Rietzschel, E.R.; Dias, K.C.; et al. Plant sterols and cardiovascular disease: A systematic review and meta-analysis. Eur. Heart J. 2012, 33, 444–451. [Google Scholar] [CrossRef]
- Jaceldo-Siegl, K.; Lütjohann, D.; Sirirat, R.; Mashchak, A.; Fraser, G.E.; Haddad, E. Variations in dietary intake and plasma concentrations of plant sterols across plant-based diets among North American adults. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef]
- Miettinen, T.A.; Gylling, H.; Lindbohm, N.; Miettinen, T.E.; Rajaratnam, R.A.; Relas, H. Finnish Treat-to-Target Study Investigators. Serum noncholesterol sterols during inhibition of cholesterol synthesis by statins. J. Lab. Clin. Med. 2003, 141, 131–137. [Google Scholar] [PubMed]
- Nunes, V.S.; de Campos, E.V.S.; Baracat, J.; França, V.; Gomes, É.I.L.; Coelho, R.P.; Nakandakare, E.R.; Zago, V.H.S.; de Faria, E.C.; Quintão, E.C.R. Plasma Campesterol Is Positively Associated with Carotid Plaques in Asymptomatic Subjects. Int. J. Mol. Sci. 2022, 23, 11997. [Google Scholar] [CrossRef] [PubMed]
- Teupser, D.; Baber, R.; Ceglarek, U.; Scholz, M.; Illig, T.; Gieger, C.; Holdt, L.M.; Leichtle, A.; Greiser, K.H.; Huster, D.; et al. Genetic regulation of serum phytosterol levels and risk of coronary artery disease. Circ. Cardiovasc. Genet. 2010, 3, 331–339. [Google Scholar] [CrossRef]
- Silbernagel, G.; Chapman, M.J.; Genser, B.; Kleber, M.E.; Fauler, G.; Scharnagl, H.; Grammer, T.B.; Boehm, B.O.; Mäkelä, K.M.; Kähönen, M.; et al. High intestinal cholesterol absorption is associated with cardiovascular disease and risk alleles in ABCG8 and ABO: Evidence from the LURIC and YFS cohorts and from a meta-analysis. J. Am. Coll. Cardiol. 2013, 62, 291–299. [Google Scholar] [CrossRef]
- CARDIoGRAMplusC4D Consortium; Deloukas, P.; Kanoni, S.; Willenborg, C.; Farrall, M.; Assimes, T.L.; Thompson, J.R.; Ingelsson, E.; Saleheen, D.; Erdmann, J.; et al. Large-scale association analysis identifies new risk loci for coronary artery disease. Nat. Genet. 2013, 45, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Weingärtner, O.; Teupser, D.; Patel, S.B. The atherogenicity of plant sterols: The evidence from genetics to clinical trials. J. AOAC Int. 2015, 98, 742–749. [Google Scholar] [CrossRef] [PubMed]
- Helgadottir, A.; Thorleifsson, G.; Alexandersson, K.F.; Tragante, V.; Thorsteinsdottir, M.; Eiriksson, F.F.; Gretarsdottir, S.; Björnsson, E.; Magnusson, O.; Sveinbjornsson, G.; et al. Genetic variability in the absorption of dietary sterols affects the risk of coronary artery disease. Eur. Heart J. 2020, 41, 2618–2628. [Google Scholar] [CrossRef]
- Helgadottir, A.; Thorleifsson, G.; Stefansson, K. Increased absorption of phytosterols is the simplest and most plausible explanation for coronary artery disease risk not accounted for by non-HDL cholesterol in high cholesterol absorbers. Eur. Heart J. 2021, 42, 283–284. [Google Scholar] [CrossRef] [PubMed]
- Plat, J.; Strandberg, T.E.; Gylling, H. Intestinal cholesterol and phytosterol absorption and the risk of coronary artery disease. Eur. Heart J. 2021, 42, 281–282. [Google Scholar] [CrossRef]
- Scholz, M.; Horn, K.; Pott, J.; Gross, A.; Kleber, M.E.; Delgado, G.E.; Mishra, P.P.; Kirsten, H.; Gieger, C.; Müller-Nurasyid, M.; et al. Genome-wide meta-analysis of phytosterols reveals five novel loci and a detrimental effect on coronary atherosclerosis. Nat. Commun. 2022, 13, 1122. [Google Scholar] [CrossRef]
- Stender, S.; Frikke-Schmidt, R.; Nordestgaard, B.G.; Tybjaerg-Hansen, A. The ABCG5/8 cholesterol transporter and myocardial infarction versus gallstone disease. J. Am. Coll. Cardiol. 2014, 63, 2121–2128. [Google Scholar] [CrossRef]
- Silbernagel, G.; Duarte, K.; Sadiku, S.; Fauler, G.; März, W.; Schmieder, R.E.; Jardine, A.G.; Massy, Z.A.; Girerd, N.; Fellström, B.; et al. High cholesterol absorption is associated with increased cardiovascular risk in haemodialysis patients: Insights from the AURORA study. Eur. J. Prev. Cardiol. 2022, 29, 1731–1739. [Google Scholar] [CrossRef]
- Otto, S.; Lütjohann, D.; Kerksiek, A.; Friedrichs, S.; Christian Schulze, P.; Möbius-Winkler, S.; Pörner, T.C.; Weingärtner, O. Increased cholesterol absorption is associated with In-stent-restenosis after stent implantation for stable coronary artery disease. Steroids 2022, 187, 109079. [Google Scholar] [CrossRef]
- Plat, J.; Baumgartner, S.; Vanmierlo, T.; Lütjohann, D.; Calkins, K.L.; Burrin, D.G.; Guthrie, G.; Thijs, C.; Te Velde, A.A.; Vreugdenhil, A.C.E.; et al. Plant-based sterols and stanols in health & disease: Consequences of human development in a plant-based environment? Prog. Lipid Res. 2019, 74, 87–102. [Google Scholar] [CrossRef] [PubMed]
- Eli Lilly Package Insert. Package Insert M100 Suspension Cytellin (Beta and Di-hydro beta Sitosterols)—A Drug Product Indicated for the Reduction of Hypercholesterolemia; Eli Lilly Package Insert: Indianapolis, IN, USA, 1954. [Google Scholar]
- Lees, R.S.; Lees, A.M. Effects of Sitosterol on Plasma Lipid and Lipoprotein Concentration. In Lipoprotein Metabolism; Springer: Berlin/Heidelberg, Germany, 1976; pp. 119–130. [Google Scholar]
- Oster, P.; Schlierf, G.; Heuck, C.C.; Greten, H.; Gundert-Remy, U.; Haase, W.; Klose, G.; Nothelfer, A.; Raetzer, H.; Schellenberg, B.; et al. Sitosterin bei familiärer Hyperlipoproteinämie Typ II. Eine randomisierte gekreuzte Doppelblindstudie. Dtsch. Med. Wochenschr. 1976, 101, 1308–1311. [Google Scholar] [CrossRef] [PubMed]
- Musa-Veloso, K.; Poon, T.H.; Elliot, J.A.; Chung, C. A comparison of the LDL-cholesterol lowering efficacy of plant stanols and plant sterols over a continuous dose range: Results of a meta-analysis of randomized, placebo-controlled trials. Prostaglandins Leukot. Essent. Fat. Acids 2011, 85, 9–28. [Google Scholar] [CrossRef] [PubMed]
- Ras, R.T.; Geleijnse, J.M.; Trautwein, E.A. LDL-cholesterol-lowering effect of plant sterols and stanols across different dose ranges: A meta-analysis of randomised controlled studies. Br. J. Nutr. 2014, 112, 214–219. [Google Scholar] [CrossRef]
- Ras, R.T.; Hiemstra, H.; Lin, Y.; Vermeer, M.A.; Duchateau, G.S.; Trautwein, E.A. Consumption of plant sterol-enriched foods and effects on plasma plant sterol concentrations--a meta-analysis of randomized controlled studies. Atherosclerosis 2013, 230, 336–346. [Google Scholar] [CrossRef]
- Kratz, M.; Kannenberg, F.; Gramenz, E.; Berning, B.; Trautwein, E.; Assmann, G.; Rust, S. Similar serum plant sterol responses of human subjects heterozygous for a mutation causing sitosterolemia and controls to diets enriched in plant sterols or stanols. Eur. J. Clin. Nutr. 2007, 61, 896–905. [Google Scholar] [CrossRef]
- Racette, S.B.; Lin, X.; Ma LOstlund, R.E.J.R. Natural dietary phytosterols. J. AOAC Int. 2015, 98, 679–684. [Google Scholar] [CrossRef]
- Hagiwara, N.; Kawada-Watanabe, E.; Koyanagi, R.; Arashi, H.; Yamaguchi, J.; Nakao, K.; Tobaru, T.; Tanaka, H.; Oka, T.; Endoh, Y.; et al. Low-density lipoprotein cholesterol targeting with pitavastatin + ezetimibe for patients with acute coronary syndrome and dyslipidaemia: The HIJ-PROPER study, a prospective, open-label, randomized trial. Eur. Heart J. 2017, 38, 2264–2276. [Google Scholar] [CrossRef]
- Päivä, H.; Thelen, K.; Van Coster, R.; Smet, J.; De Paepe, B.; Mattila, K.M.; Laakso, J.; Lehtimäki, T.; von Bergmann, K.; Lütjohann, D.; et al. High-dose statins and skeletal muscle metabolism in humans: A randomized, controlled trial. Clin. Pharm. Ther. 2005, 78, 60–68. [Google Scholar] [CrossRef]
- Van Himbergen, T.M.; Matthan, N.R.; Resteghini, N.A.; Otokozawa, S.; Ai, M.; Stein, E.A.; Jones, P.H.; Schaefer, E.J. Comparison of the effects of maximal dose atorvastatin and rosuvastatin therapy on cholesterol synthesis and absorption markers. J. Lipid Res. 2009, 50, 730–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Windler, E.; Beil, F.-U.; Berthold, H.K.; Gouni-Berthold, I.; Kassner, U.; Klose, G.; Lorkowski, S.; März, W.; Parhofer, K.G.; Plat, J.; et al. Phytosterols and Cardiovascular Risk Evaluated against the Background of Phytosterolemia Cases—A German Expert Panel Statement. Nutrients 2023, 15, 828. https://0-doi-org.brum.beds.ac.uk/10.3390/nu15040828
Windler E, Beil F-U, Berthold HK, Gouni-Berthold I, Kassner U, Klose G, Lorkowski S, März W, Parhofer KG, Plat J, et al. Phytosterols and Cardiovascular Risk Evaluated against the Background of Phytosterolemia Cases—A German Expert Panel Statement. Nutrients. 2023; 15(4):828. https://0-doi-org.brum.beds.ac.uk/10.3390/nu15040828
Chicago/Turabian StyleWindler, Eberhard, Frank-Ulrich Beil, Heiner K. Berthold, Ioanna Gouni-Berthold, Ursula Kassner, Gerald Klose, Stefan Lorkowski, Winfried März, Klaus G. Parhofer, Jogchum Plat, and et al. 2023. "Phytosterols and Cardiovascular Risk Evaluated against the Background of Phytosterolemia Cases—A German Expert Panel Statement" Nutrients 15, no. 4: 828. https://0-doi-org.brum.beds.ac.uk/10.3390/nu15040828