1. Introduction
Obesity leads to a wide range of health problems and is associated with an increased risk of metabolic syndrome. Non-alcoholic fatty liver disease (NAFLD), a hepatic manifestation of metabolic syndrome, has been becoming a major public health problem due to the obesity epidemic [
1]. It has been reported that one-third of the urban population in the United States had hepatic steatosis [
2], and the prevalence of NAFLD in Asian countries has been reported to be 16%–18% [
3,
4]. NAFLD covers a spectrum of liver abnormalities ranging from simple steatosis to fibrosis or cirrhosis accompanied by severe inflammation. The mechanisms leading to hepatic lipid accumulation include increased lipid uptake, elevated
de novo lipogenesis, improper fatty acid oxidation, and dysregulation of lipoprotein metabolism. An excessive level of hepatic lipids makes the liver vulnerable to further damage caused by inflammation [
5]. There have been many efforts to discover a pharmacologic target for NAFLD treatment based on the pathological process of NAFLD; however, a strategy for treating NAFLD has not been firmly established [
6]. Thus, lifestyle modifications focusing on improving an individual’s diet and level of physical activity have been proposed as a primary strategy for NAFLD treatment.
Dietary fatty acids play a significant role in modulating hepatic lipid metabolism [
7]. Thus, not only the amount but also the composition of the dietary fat plays an important role in the prevention of NAFLD. It has been reported that saturated fatty acids contribute to the progression of hepatic steatosis by promoting endoplasmic reticulum stress and the apoptosis of hepatocytes [
8], and conjugated linoleic acid (CLA) has also been linked to the development of hepatic steatosis despite its influence on weight reduction [
9]. Conversely, omega-3 polyunsaturated fatty acids (PUFAs) have been known to have beneficial effects on hepatic lipid metabolism. The mechanisms behind these effects include reduced expression and/or activity of hepatic lipogenic genes, activation of hepatic fatty acid β-oxidation, and inhibition of the production of inflammatory mediators [
10], which may result in protection from hepatic lipid accumulation. Recently, oil extracted from the seeds of
Pinus koraiensis, pine nut oil (PNO), has been investigated for its impact on lipid metabolism. PNO contains pinolenic acid (18:3, Δ5, 9, 12), which is an unusual omega-6 PUFA characterized by polymethylene-interrupted double bonds. Asset
et al. [
11,
12] showed that PNO consumption lowered the plasma cholesterol level compared with the consumption of other lipids such as sunflower oil, coconut oil, or lard in mice. Two previous studies have investigated PNO’s effects on weight-loss and NAFLD prevention. In a study by Ferramosca
et al. [
13], the consumption of a PNO-supplemented diet significantly reduced body weight gain and the liver weight relative to the consumption of a maize oil-supplemented diet in mice. In this study, a standard diet (12% of the energy from fat) was supplemented with PNO or maize oil, which made the diets have 30% of the energy from fat. However, a low-fat control group was not included in Ferramosca’s study, and the fat content of the diets was significantly lower than the amount in the commonly used high-fat diet (HFD) for diet-induced obesity. In another study by Ferramosca
et al. [
14], it was shown that the combination of CLA and PNO prevented a CLA-induced fatty liver and improved insulin sensitivity in mice. In this study, a standard diet (12% of the energy from fat) was supplemented with olive oil (control group), CLA and olive oil (CLA group), or CLA and PNO (CLA and PNO group) to make diets contain 30% of the energy from fat. Collectively, these findings suggest that PNO can affect weight gain and has the potential for NAFLD prevention by its modulation of lipid metabolism. However, the information regarding the role of PNO on NAFLD prevention is still limited. There is no study that has examined the effect of PNO in both a control diet and a HFD.
Some studies have demonstrated that PNO has appetite controlling effects [
15,
16], which suggested that PNO can lead to less energy consumption. Sirtuins (SIRTs), class 3 NAD
+-dependent deacetylases, are known to be upregulated in the chronic caloric restriction (CR). It has been well-documented that SIRTs are responsible for the health benefits of CR such as longevity and protection from age-related diseases including cancer, neurodegeneration, and cardiovascular disease [
17,
18]. There are seven mammalian SIRTs, ranging from SIRT1 to SIRT7. Among the SIRTs, SIRT3 is the major mitochondrial form that has relevance in supporting fatty acid oxidation, enhancing the antioxidant defense system, and repairing mitochondrial DNA damage, which implies that SIRT3 can be a new target for protection against lipotoxicity and obesity-induced metabolic complications [
19]. Thus, we examined whether the SIRT3 protein was upregulated in PNO-fed mice. Among the SIRTs, SIRT3 was chosen because of its significance not only in CR and but also in lipid metabolism as well.
In the present study, HFDs with 45% of the energy from fat with 10% from PNO or soybean oil (SBO) and 35% from lard were used to examine whether partial replacement with PNO in the HFD can ameliorate NAFLD in HFD-induced obese mice. The impact of PNO was also examined using control diets with 10% of the energy from fat from PNO or SBO. We measured the hepatic lipid contents and hepatic mRNA levels of genes involved in fatty acid oxidation and lipogenic pathways to evaluate the influence of PNO on hepatic fatty acid metabolism. Expression of the SIRT3 protein was measured in order to determine whether PNO had a similar effect to that of caloric restriction.
2. Materials and Methods
2.1. Animals and Diets
Five-week-old male C57BL/6 mice (Central Lab, Animal Inc., Seoul, Korea) were randomly divided into four groups after 3 days of acclimation. Mice were fed the experimental diets for 12 weeks
ad libitum. Control diets contained 10% of the energy from fat from PNO (PC,
n = 11) or SBO (SC,
n = 10). HFDs contained 45% of the energy from fat with 35% from lard and 10% from PNO (PHFD,
n = 11) or SBO (SHFD,
n = 11). The PNO was a gift from the Dubio Co., Ltd. (Gyeonggi-do, Korea). The compositions of the experimental diets are shown in
Table 1. The mice were housed individually and maintained in an animal facility with controlled temperature (23 ± 3 °C), controlled humidity (55% ± 10%), and a 12 h light/12 h dark cycle. SBO was chosen as the control oil because its fatty acid composition was similar to that of PNO except for its pinolenic acid content. The fatty acid compositions of the experimental diets are shown in
Table 2. An antioxidant was added to the diets (0.2 μg of t-butylhydroquinone/g oil) to prevent the oxidation of polyunsaturated fatty acids, and fresh food was provided every other day. The body weights of the mice were recorded once a week, and their food intake was measured four times a week. At the end of the experimental period, the mice were fasted for 12 h and euthanized by asphyxiation with CO
2. Blood was obtained by heart puncture, and serum was collected after centrifugation and stored at −80 °C for later analysis. White adipose tissue (epididymal, abdominal subcutaneous, and retroperitoneal-perirenal depots) was collected and stored at −80 °C for later analysis. For liver histology, fresh liver tissue was dissected into pieces in a consistent manner and fixed in 10% buffered neutral formalin. The rest of the liver tissue was stored at −80 °C for later analysis. This study was approved by the Animal Care and Use Committee at Seoul National University (approval No. SNU-101029-1).
Table 1.
Composition of the experimental diets 1.
Table 1.
Composition of the experimental diets 1.
| | Control Diet (g) (10% of the kcal from Fat) | High-Fat Diet (g) (45% of the kcal from Fat) |
|---|
| | 10% Oil | 10% Oil + 35% Lard |
| Casein | 200 | 200 |
| l-Cystine | 3 | 3 |
| Sucrose | 350 | 172.8 |
| Cornstarch | 315 | 72.8 |
| Dyetrose | 35 | 100 |
| PNO 2 or SBO | 45 | 45 |
| Lard | 0 | 157.5 |
| t-Butylhydroquinone | 0.009 | 0.009 |
| Cellulose | 50 | 50 |
| Mineral Mix 3 | 35 | 35 |
| Vitamin Mix 4 | 10 | 10 |
| Choline Bitartrate | 2 | 2 |
| Total | 1045.0 | 848.1 |
| kcal/g diet | 3.69 | 4.64 |
Table 2.
Fatty acid composition of the experimental diets (% of fatty acids) 1.
Table 2.
Fatty acid composition of the experimental diets (% of fatty acids) 1.
| | SBO | PNO |
|---|
| | Control Diet (SC) | High-Fat Diet (SHFD) | Control Diet (PC) | High-Fat Diet (PHFD) |
|---|
| Myristic acid (C14:0) | ND | 0.9 | ND | 0.9 |
| Palmitic acid (C16:0) | 11.9 | 18.9 | 7.0 | 17.8 |
| Stearic acid (C18:0) | 4.8 | 11.1 | 3.6 | 10.7 |
| Total saturated fatty acid | 16.7 | 30.9 | 10.6 | 29.4 |
| Palmitoleic acid (C16:1, Δ9) | ND | 1.4 | ND | 1.4 |
| Oleic acid (C18:1, Δ9) | 21.1 | 34.7 | 27.4 | 36.0 |
| Total monounsaturated fatty acid | 21.1 | 36.1 | 27.4 | 37.4 |
| Linoleic acid (C18:2, Δ9, 12) | 54.9 | 30.3 | 47.2 | 28.6 |
| α-linolenic acid (C18:3, Δ9, 12, 15) | 7.4 | 2.8 | 0.8 | 1.3 |
| Pinolenic acid (C18:3, Δ5, 9, 12) | ND | ND | 14.0 | 3.3 |
| Total polyunsaturated fatty acid | 62.3 | 33.1 | 62.0 | 33.2 |
2.2. Serum Lipid Concentrations
The serum non-esterified fatty acid (NEFA) (SICDIA NEFAZYME; Shin Yang Chemical, Seoul, Korea), triacylglycerol (TG) (Cleantech TG-S; Asan Pharmaceutical, Seoul, Korea), and total cholesterol (T-CHO; Asan Pharmaceutical, Seoul, Korea) concentrations were measured by enzymatic colorimetric methods according to the manufacturer’s instructions.
2.3. Serum Leptin and Fetuin-A Concentrations
Serum leptin (Quantikine® ELISA kit; R & D Systems, Minneapolis, MN, USA) and fetuin-A (mouse Fetuin-A/AHSG Duo set; R & D Systems) levels were determined using enzyme-linked immunosorbent assays (ELISAs).
2.4. Liver Histology
For hematoxylin and eosin staining, liver tissue was embedded in paraffin and stained with hematoxylin and eosin. For oil red O staining, liver tissue was frozen in OCT compound and stained with oil red O. Histological images were captured at 200× magnification using a BX51 light microscope (Olympus, Optical Co. Ltd., Tokyo, Japan).
2.5. Hepatic Lipid Contents
The total lipids were extracted by the Folch method [
20] from the liver tissue. The lipids were extracted in chloroform, dried in nitrogen gas and redissolved in isopropanol. The hepatic TG and cholesterol levels were determined by the same enzymatic colorimetric methods used for the measurement of the serum lipids. The hepatic TG and cholesterol contents are presented as mg/g tissue.
2.6. Real-Time PCR Analysis of the Genes in Liver Tissue
The mRNA levels of fetuin-A and genes involved in fatty acid oxidation and lipogenic pathways in the liver were determined. The oligonucleotide sequences of primers used in this study are presented in
Table 3. The total RNA was isolated from the liver tissue using TRIzol reagent (Life Technologies Co., Carlsbad, CA, USA). The RNA concentration was measured by a spectrophotometer (Beckman Instruments, Fullerton, CA, USA), and the quality of the RNA was confirmed by agarose gel electrophoresis. The total RNA (2 μg) was used to synthesize single-stranded cDNA using a cDNA synthesis kit (PrimeScript™ II 1st strand cDNA synthesis kit; Takara Bio Inc., Shiga, Japan). The relative mRNA level was determined by the real-time quantitative PCR analysis using the SYBR green assay (SYBR
® Premix Ex TaqTM; Takara Bio Inc., Shiga, Japan), and the analysis was performed with a Step-One-Plus RT-PCR system (Life Technologies, Carlsbad, CA, USA). All of the samples were analyzed in duplicate with an endogenous control gene being analyzed at the same time. The relative expression levels of each gene were calculated by the 2
−ΔΔCT method, with the
Gapdh gene being used as the endogenous control.
Table 3.
Primer sequences used in real-time PCR quantitative analysis.
Table 3.
Primer sequences used in real-time PCR quantitative analysis.
| Gene | Function | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|
| Ahsg/fetuin-A | fatty liver indicator | TTGCTCAGCTCTGGGGCT | GGCAAGTGGTCTCCAGTGTG |
| Ppara | transcription factor promoting fatty acid oxidation | GCAGTGGAAGAATCGGACCT | CAACCCGCCTTTTGTCATAC |
| Cpt1a | mitochondrial β-oxidation | GATGTTCTTCGTCTGGCTTGA | CTTATCGTGGTGGTGGGTGT |
| Acadl | mitochondrial β-oxidation | TCGCAATATAGGGCATGACA | ACTTGGGAAGAGCAAGCGTA |
| Hadha | mitochondrial β-oxidation | CCCTTTGAACACTTGCTGCT | GCCCAGGTCTCTGTGGATAA |
| Acox1 | peroxisomal β-oxidation | GTCAAAGGCATCCACCAAAG | GAGGGGAACATCATCACAGG |
| Cyp4a10 | microsomal ω-oxidation | CAGAAAGGAGGGAAGATGGAG | CATGGTCTCCAAAATCCAAGG |
| Sod2 | anti-oxidative defense | TTAGAGCAGGCAGCAATCTGT | GCGTGACTTTGGGTCTTTTG |
| Ucp2 | anti-oxidative defense | CAGGTCACTGTGCCCTTACCA | CACTACGTTCCAGGATCCCAA |
| Pparg | transcription factor promoting adipogenesis | CAGCAGGTTGTCTTGGATGTC | AGCCCTTTGGTGACTTTATGG |
| Srebf1 | de novo lipogenesis | GTCTCCACCACTTCGGGTTT | CGACTACATCCGCTTCTTGC |
| Fasn | de novo lipogenesis | GCGGTGTGAAAACGAACTTT | CTGTCTGGGCATAACGGTCT |
| Slc25a1 | de novo lipogenesis | TTCCCTTTAGCCCTTGTTCC | TGACCAGACTTCCTCCAACC |
| Fabp1 | fatty acid transport | GAACTCATTGCGGACCACTT | CATCCAGAAAGGGAAGGACAT |
| Cd36 | fatty acid transport | CCAAGCTATTGCGACATGATT | TCTCAATGTCCGAGACTTTTCA |
| Gapdh | endogenous control | GGAGAAACCTGCCAAGTA | AAGAGTGGGAGTTGCTGTTG |
2.7. Western Blot Analysis of SIRT3 in White Adipose Tissue
The epididymal fat pad was homogenized in an ice-cold protein lysis buffer containing 50 mM Tris-Cl (pH 7.4), 150 mM NaCl, 1 mM EDTA, 1 mM PMSF, 1 mM Na3VO4, 1 mM NaF, 1 mM Na4P2O7, 1 mM β-glycerophosphate, 1% NP-40, 0.25% sodium deoxycholate, 10% glycerol, and protease inhibitor cocktail (Complete Mini Protease Inhibitor Cocktail; Roche Diagnostics GmbH, Penzberg, Germany). The supernatant was obtained by centrifugation at 10,000× g for 30 min at 4 °C. A Bradford assay (Bio-Rad, Hercules, CA, USA) was used to determine the total protein concentration. Fifty micrograms of the protein sample was separated in 10% SDS-PAGE and transferred onto a PVDF membrane (Bio-Rad, Hercules, CA, USA). The membrane was blocked with 3% bovine serum albumin (BSA) in Tris-buffered saline containing 0.1% Tween-20 (pH 7.6, TBST) for 1 h. The membrane was incubated with rabbit anti-mouse SIRT3 antibody (1:1000; Cell Signaling Technology, Beverly, MA, USA) in 3% BSA/TBST overnight at 4 °C. The membrane was washed with TBST and then incubated with secondary anti-rabbit IgG HRP-linked antibody (Cell Signaling Technology, Beverly, MA, USA) in 3% BSA/TBST for 1 h at room temperature. The signal was visualized by a chemiluminescent detection system and quantified using Quantity One analysis software (Bio-Rad, Hercules, CA, USA).
2.8. Statistical Analysis
Statistical analysis was performed using PASW Statistics 18 (SPSS Inc., Chicago, IL, USA). Two-way ANOVA was used to evaluate the overall effects of the fat amount, oil type, and their interaction. When the effects of the fat amount and/or oil type were significant, a LSD multiple-comparison post-hoc test was performed for the individual group comparisons. Student’s t test was used for the comparison between the PC and SC groups or the PHFD and SHFD groups if an interaction was significant. Pearson’s correlation was used to determine the relationship between the two variables. The results from all of the comparisons were considered significant at p < 0.05. Data are reported as the means ± SEM.
4. Discussion
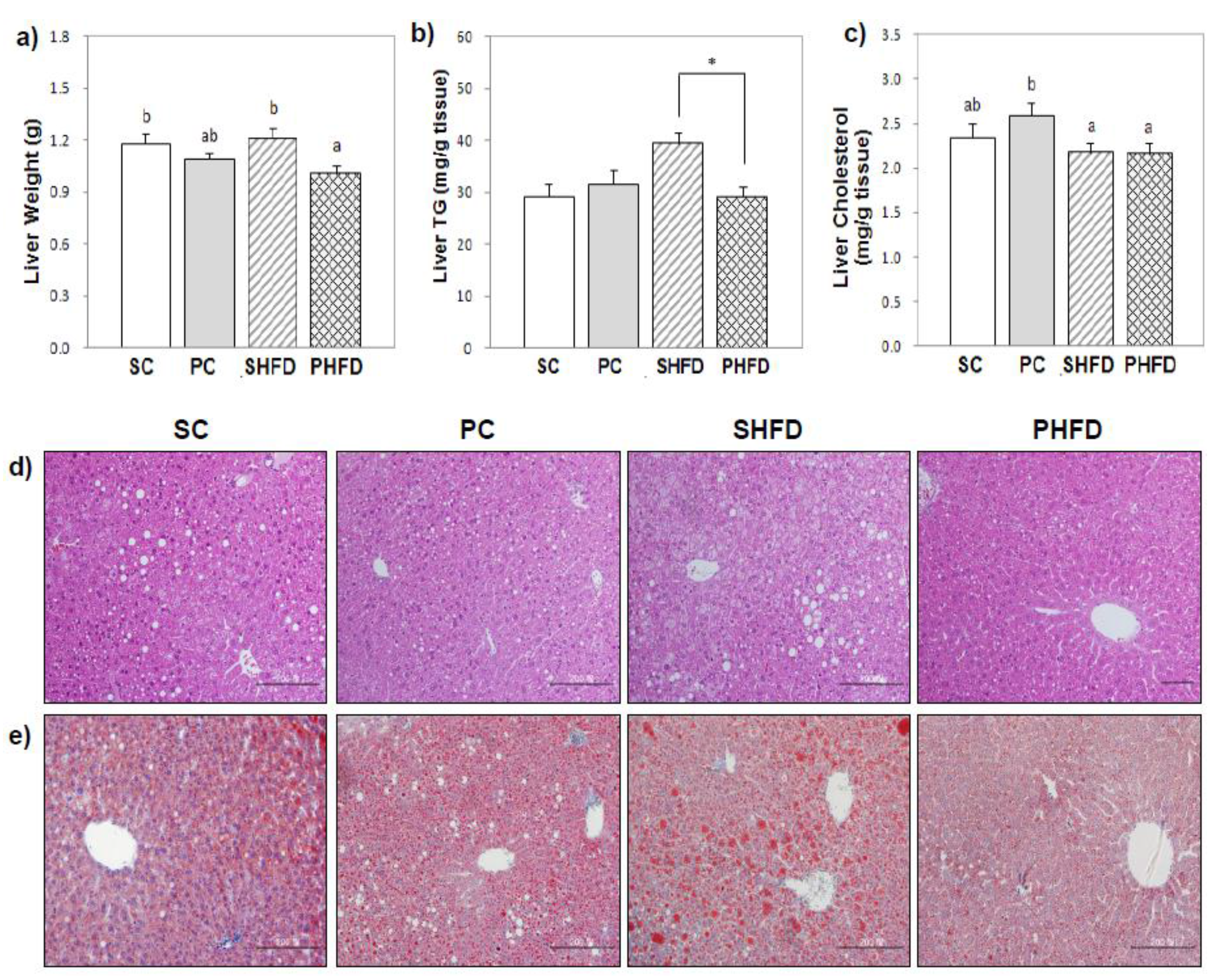
In this study, we showed that hepatic TG accumulation was attenuated by PNO replacement in HFD-fed mice. Mice in the SHFD group showed 30% higher hepatic TG concentrations compared to the control group, but mice in the PHFD group maintained hepatic TG concentrations at the level of lean mice. NAFLD induced by feeding the mice a HFD (45% of the energy from fat) for 12 weeks in this study seemed to remain in the initial stage of a simple steatotic condition. A hepatic TG content over 5% of the liver weight, which is the criteria for a fatty liver [
21], was not observed overall. On pathological observation, inflammation and hepatocyte degeneration were mild (
Table A1) even in the SHFD group. The expression of
Pparg mRNA was also not higher in the HFD-fed mice compared with the control diet-fed mice. Meanwhile, the hepatic cholesterol level was lower in HFD-fed mice, and PNO had no effect on the hepatic cholesterol level. In the study by de Vogel-van den Bosch
et al. [
22], feeding a cholesterol-free high-fat diet lowered fractional cholesterol absorption, which resulted in cholesterol synthesis and down-regulation of cholesterol efflux to spare intracellular cholesterol for chylomicron formation. Desmarchelier
et al. [
23] also reported that mice fed with the high-fat western diet containing 0.03% (
w/
w) cholesterol showed reduced intestinal and hepatic cholesterol concentrations despite the hypercholesterolemia and increased hepatic TG levels. The HFD used in this study provided 0.02% (
w/
w) cholesterol from lard. It seems that providing this amount of cholesterol is not sufficient to meet the increased demand for chylomicron packaging due to lipid overload with high-fat diet feeding. The duration of high-fat feeding was not very long in this study (12 weeks) and two studies by others (8 weeks in the study by de Vogel-van den Bosch
et al. [
22] and 12 weeks in the study by Desmarchelier
et al. [
23]). Taken together, lower levels of hepatic cholesterol in HFD fed mice in this study might be interpreted as an adaptive response to deal with large quantities of fat in the diet.
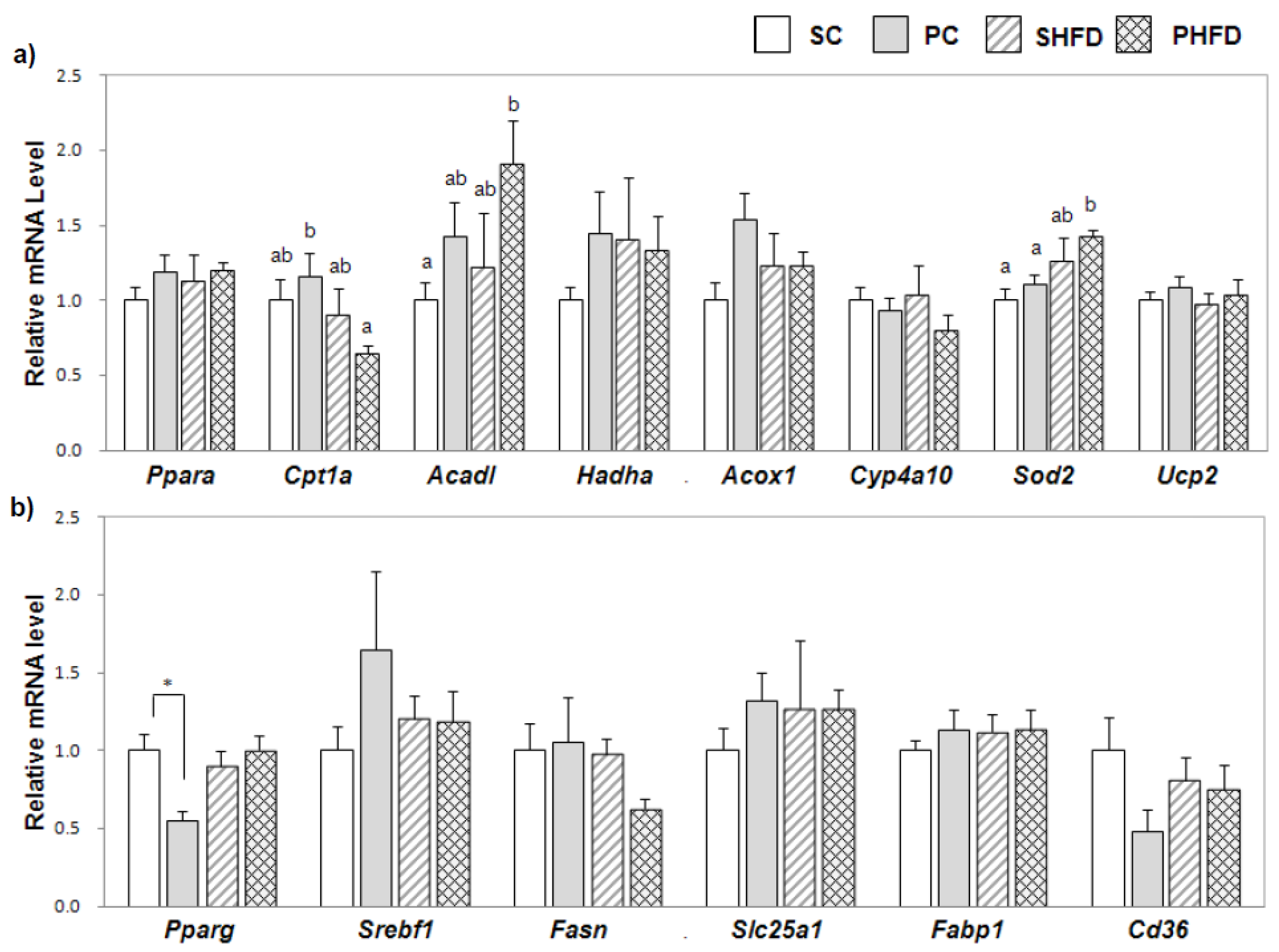
The mRNA level of long-chain acyl-CoA dehydrogenase
(Acadl) in PNO-fed mice was significantly higher, and the PHFD group expressed the highest level of
Acadl among the four groups. The up-regulation of
Acadl mRNA might contribute to the enhancement of fatty acid β-oxidation, which could partially explain the smaller hepatic TG accumulation in PNO-fed mice. The role of fatty acid oxidation in NAFLD is controversial. Studies by Schrauwen
et al. [
24] and Sunny
et al. [
25] reported that fatty acid oxidation was higher in steatotic livers, which could lead to oxidative stress and liver damage. However, the increase in fatty acid oxidation in both studies was attributed to the excessive amount of TG stored in the liver tissue, whereas PNO-fed mice in this study showed little TG accumulation. Additionally, the PHFD group had the highest level of superoxide dismutase 2 (
Sod2) mRNA among the four groups, which might confer a protection from the reactive oxygen species produced by increased fatty acid oxidation. Because
Sod2 can also be induced by reactive oxygen species [
26], this result is consistent with the fact that the PHFD group showed increased expression of
Acadl. However, the PFHD group had the lowest mRNA expression level of carnitine palmitoyltransferase 1a
(Cpt1a) among the four groups. Depletion of unesterified coenzyme A, a cofactor required for fatty acid oxidation in the mitochondria, would further inhibit the entry of fatty acids into the mitochondria [
27,
28]. As increased mitochondrial β-oxidation could lead to a shortage of available cofactors, down-regulation of
Cpt1a mRNA in the PHFD group might be the result of an intramitochondrial control mechanism to prevent a further influx of fatty acids in order to maintain mitochondrial homeostasis. The results from a recent study by Le
et al. [
29], in which PNO increased both
Cpt1b and
Acadl mRNA levels in the muscle tissue of HFD-fed mice, suggested a tissue-specific regulation. In fact, liver and muscle tissues express different isoforms of
Cpt1, and the two isoforms have different kinetic characteristics [
28]. However, direct mechanistic evidence to explain the contradictory expression pattern of the genes involved in mitochondrial β-oxidation (higher
Acadl and lower
Cpt1) shown in this study is lacking. A direct comparison of the mitochondrial fatty acid oxidation rates would help confirm the effect of PNO on fatty acid oxidation in the liver.
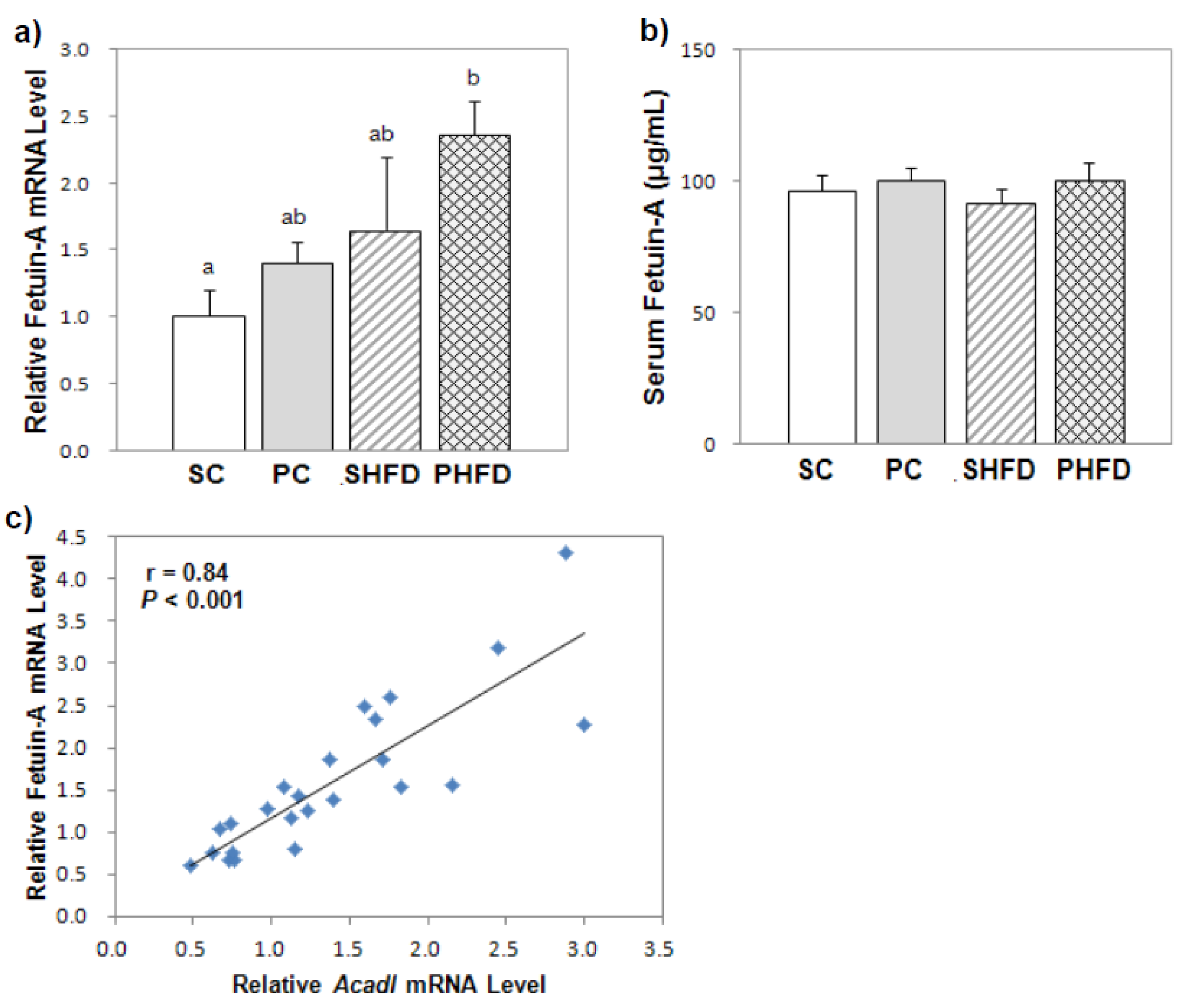
Fetuin-A has been linked to weight gain, insulin resistance, and NAFLD [
30,
31,
32]. Fetuin-A knockout mice were insulin-sensitive and resistant to weight gain when fed a HFD [
33]. Stefan
et al. [
34] reported that the plasma fetuin-A level positively correlated with liver fat accumulation in humans, and the expression of fetuin-A mRNA was greater in mice with fatty livers. In this study, the serum fetuin-A concentration was not higher in obese mice despite the upregulated hepatic fetuin-A mRNA level in obese mice. Because fetuin-A is present in systemic circulation at a high concentration, the magnitude of the elevated fetuin-A expression in the liver might have been insufficient to cause a significant change in the serum fetuin-A concentration. However, the hepatic expression of fetuin-A mRNA was greater in PNO-fed mice without evidence of hepatic steatosis, and it positively correlated with the genes involved in fatty acid oxidation. Haukeland
et al. [
35] reported that hepatic fetuin-A mRNA levels positively correlated with
Pepck1 and
Srebp1c mRNA levels in human subjects. A positive correlation between fetuin-A and
Cpt1 mRNA levels was also observed, although it was less consistent. These results suggest that fetuin-A mRNA expression could be co-regulated with glucose and lipid metabolism. Based on our results, the expression of fetuin-A mRNA seemed to be closely related to fatty acid oxidative metabolism, and the up-regulation of fetuin-A mRNA in PNO-fed mice could be the result of increased fatty acid oxidation. Further studies are needed to elucidate the relationship between fetuin-A and fatty acid metabolism.
Although the overall effect of PNO on the mRNA expression of fatty acid synthase
(Fasn), a multi-enzyme protein involved in
de novo lipogenesis, was not significant, it was significantly lower in the PHFD group than in the SHFD group (Student’s
t-test,
p < 0.05). Ferramosca
et al. [
14] reported that the activities of hepatic lipogenic enzymes including acetyl-CoA carboxylase and fatty acid synthase were significantly decreased in PNO-fed mice after 16 weeks of feeding, and PNO-fed mice were protected from liver fat accumulation when CLA was fed, which partially supports our results. In the study by Donnelly
et al. [
36],
de novo lipogenesis was elevated in the fasting state and failed to increase in the postprandial state in NAFLD patients, which suggested dysregulated
de novo lipogenesis in NAFLD patients. Our results indicated that the attenuation of the lipogenic process under the condition of HFD consumption occurred in mice in the PHFD group, while this was not observed in mice in the SHFD group. Collectively, these data suggested that PNO replacement contributes to the maintenance of the homeostatic regulation of hepatic fatty acid metabolism in HFD-induced obesity by increasing fatty acid β-oxidation and alleviating lipogenesis.
The changes in the expression of hepatic genes related to fatty acid oxidation and lipogenesis by PNO replacement have been discussed so far. In this study, PNO-fed mice showed reduced weight gain and body fat accumulation along with the less hepatic TG accumulation in the HFD group. It is well established that an excess flow of free fatty acids arising from adipose tissue is the major contributor to hepatic TG in obese patients [
36], which is mainly explained by the failed suppression of lipolysis caused by peripheral insulin resistance. Thus, there is a possibility that less hepatic TG accumulation in the PHFD group in this study could be an indirect consequence of less body fat accumulation followed by an overall improvement in energy metabolism.
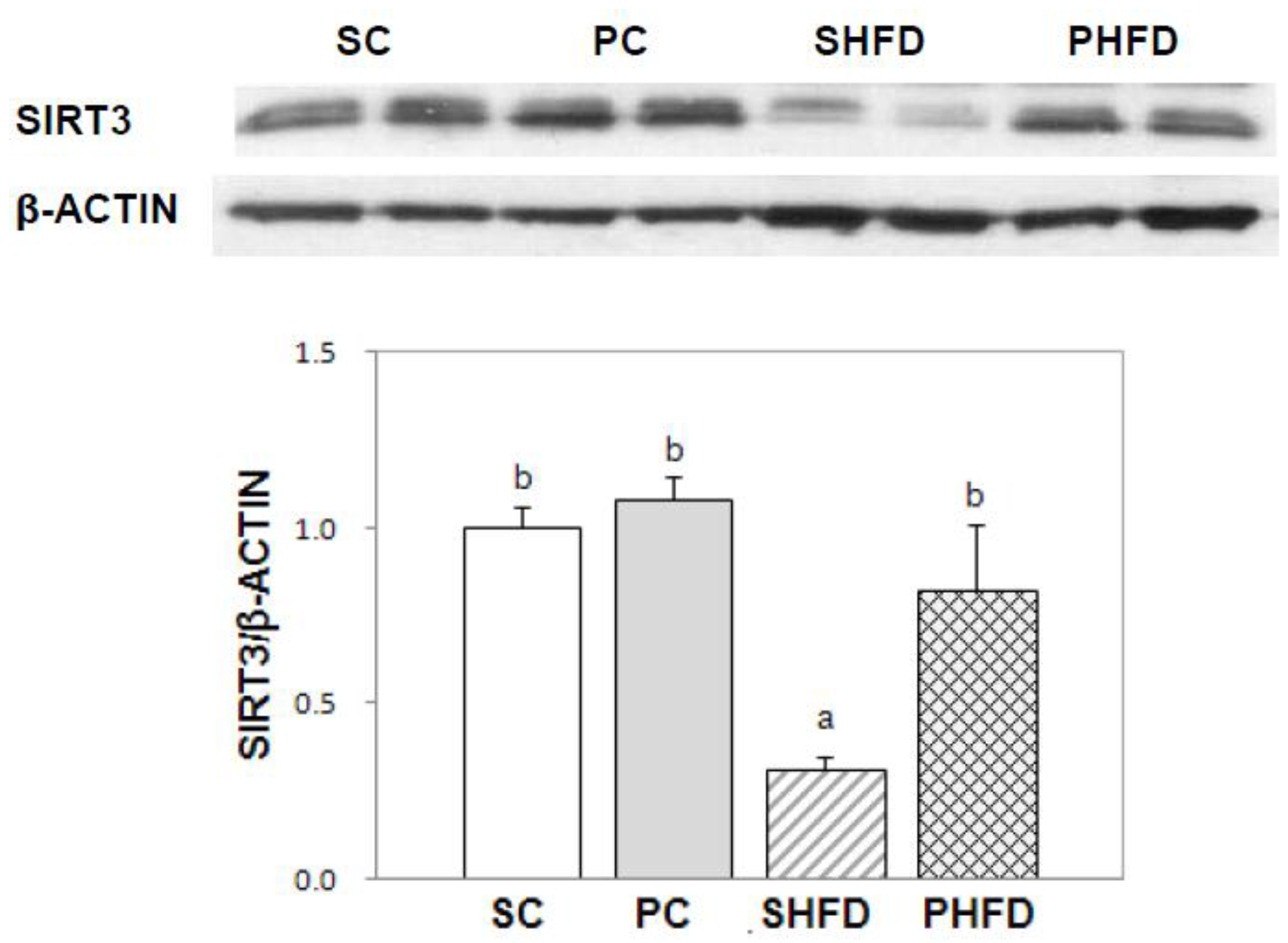
Among the SIRTs (SIRT1-7), SIRT3 plays a pivotal role in energy metabolism and mitochondrial function by regulating the activity of mitochondrial enzymes related to fatty acid metabolism including ACADL [
37] and SOD2 [
38]. We observed that SIRT3 protein expression was significantly lower in the white adipose tissue of the SHFD group compared with that of mice fed control diets, which seemed to be the result of excess energy storage in the white adipose tissue of mice in the SHFD group. Conversely, SIRT3 protein expression in the white adipose tissue from the PHFD group was maintained at the same level as that of the control diet-fed mice and was not reduced despite the HFD feeding. These results suggested that partial PNO replacement prevented the down-regulation of SIRT3 caused by high-fat feeding and that PNO could alleviate mitochondrial dysregulation by maintaining SIRT3 protein expression in HFD-induced obesity.
Collectively, it seemed that PNO-fed mice showed mild CR-mimetic responses in both the control diet and HFD groups, although the actual energy intake was not significantly different from the corresponding SBO-containing diet groups. First, mice in the PHFD group showed less hepatic lipid accumulation accompanied with less whole-body fat accumulation compared with the mice in the SHFD group. Hepatic lipid reduction with whole-body weight loss is commonly observed when obese animals [
39] or human subjects [
40,
41] were placed on CR. Higher SIRT3 protein expression in white adipose tissue from the PHFD group than that from the SHFD group further supports this idea. Secondly, mice in the PC group also showed the least body weight gain among the four groups and also showed markedly reduced
Pparg mRNA expression compared to the other groups. It was reported that the hepatic
Pparg mRNA level was higher in NAFLD patients [
42], and CR reduced the expression of hepatic
Pparg mRNA in mice [
43]. Thirdly, the serum NEFA levels of mice in the PC group were higher than those of mice in the SC group (Student’s
t-test,
p < 0.05) even though mice in the PC group had less white adipose tissue than those in the SC group. Elevation of fasting serum NEFA is a metabolic feature of long-term CR [
44], which is a result of enhanced fat mobilization as an adaptive response to CR.
The pinolenic acid (18:3, Δ5, 9, 12) in PNO (14% of the fatty acid composition) is a strong candidate for the cause of the PNO-mediated effect observed in this study. In the study by Le
et al. [
29], pinolenic acid elicited ligand activity for PPARα and PPARδ, and pinolenic acid treatment upregulated the downstream target genes of PPARα and PPARδ involved in fatty acid oxidation, including
Pgc1a,
Ucp3,
Cpt1b,
Acadm, and
Acadl in the C2C12 myotubes cell line. The ligand activity of pinolenic acid for PPARα and PPARδ could be a reason for the lower body fat accumulation in PNO-fed mice considering the importance of PPAR in the lipid metabolism of adipose tissue. However, Le
et al. mainly focused on skeletal muscle and brown adipose tissue. Thus, mechanistic studies that examine the expression of downstream genes of PPARα and PPARδ using liver and white adipose tissue cell lines could be helpful for achieving further understanding.


{kind=link}
{kind=link}
{kind=link}
{kind=link}



