Enzymatically-Processed Wheat Bran Enhances Macrophage Activity and Has in Vivo Anti-Inflammatory Effects in Mice


Abstract
:1. Introduction
2. Experimental Methods
2.1. Preparation of Enzyme-Treated Wheat Bran
2.2. Determination of Dietary Fiber Content
2.3. Determination of Water Soluble Arabinoxylan
2.4. Analysis of Sugar Composition
2.5. Animals
2.6. In Vivo Experiment 1
2.7. iExperiment 2
2.8. Flow Cytometry
2.9. Cell Culture Ex Vivo
2.10. Phagocytosis Assay
2.11. Nitric Oxide Determination
2.12. Cytokine Analysis
2.13. Statistical Analysis
3. Results
3.1. Analysis of Dietary Fiber, Arabinoxylan, and Monosaccharides
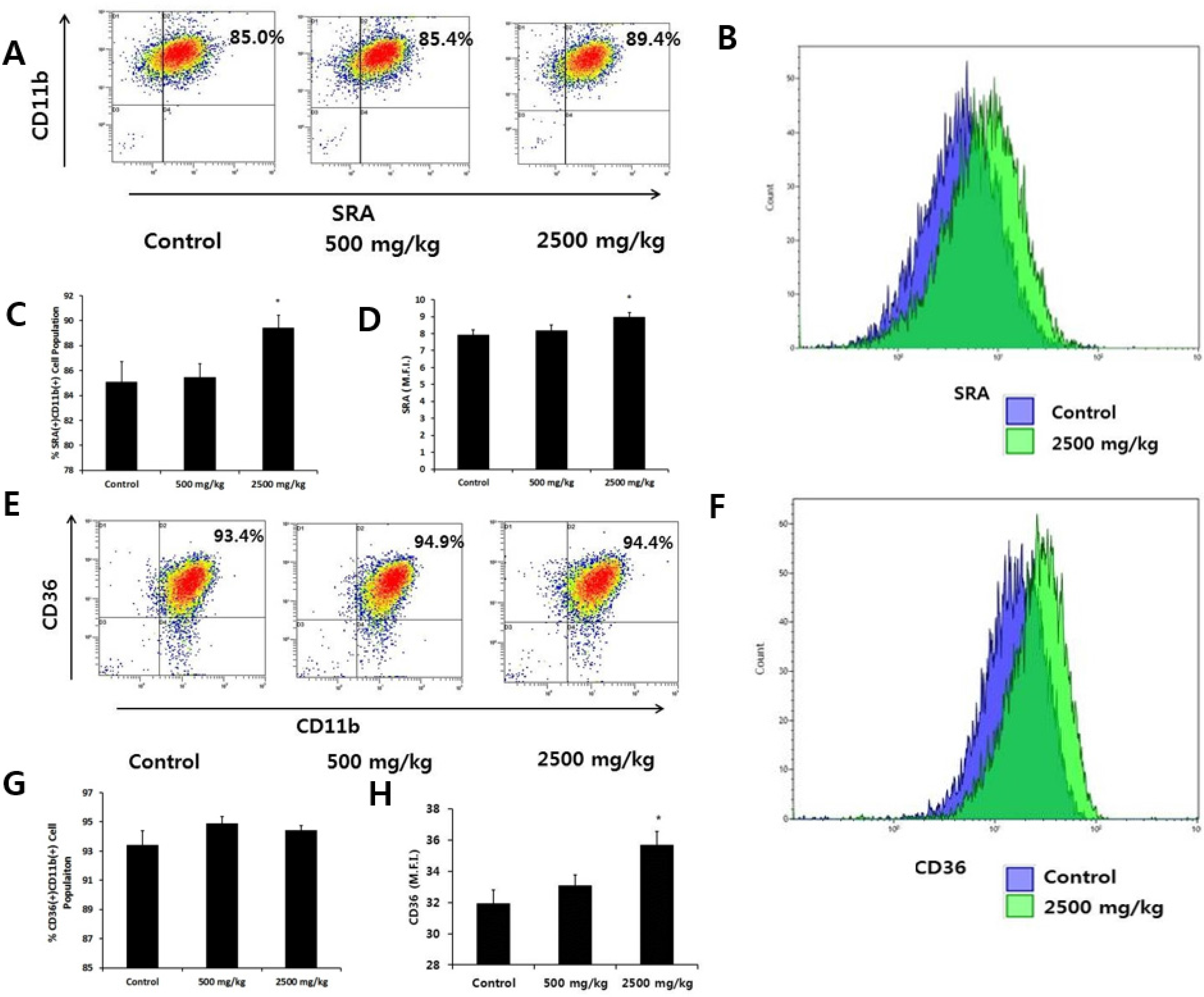
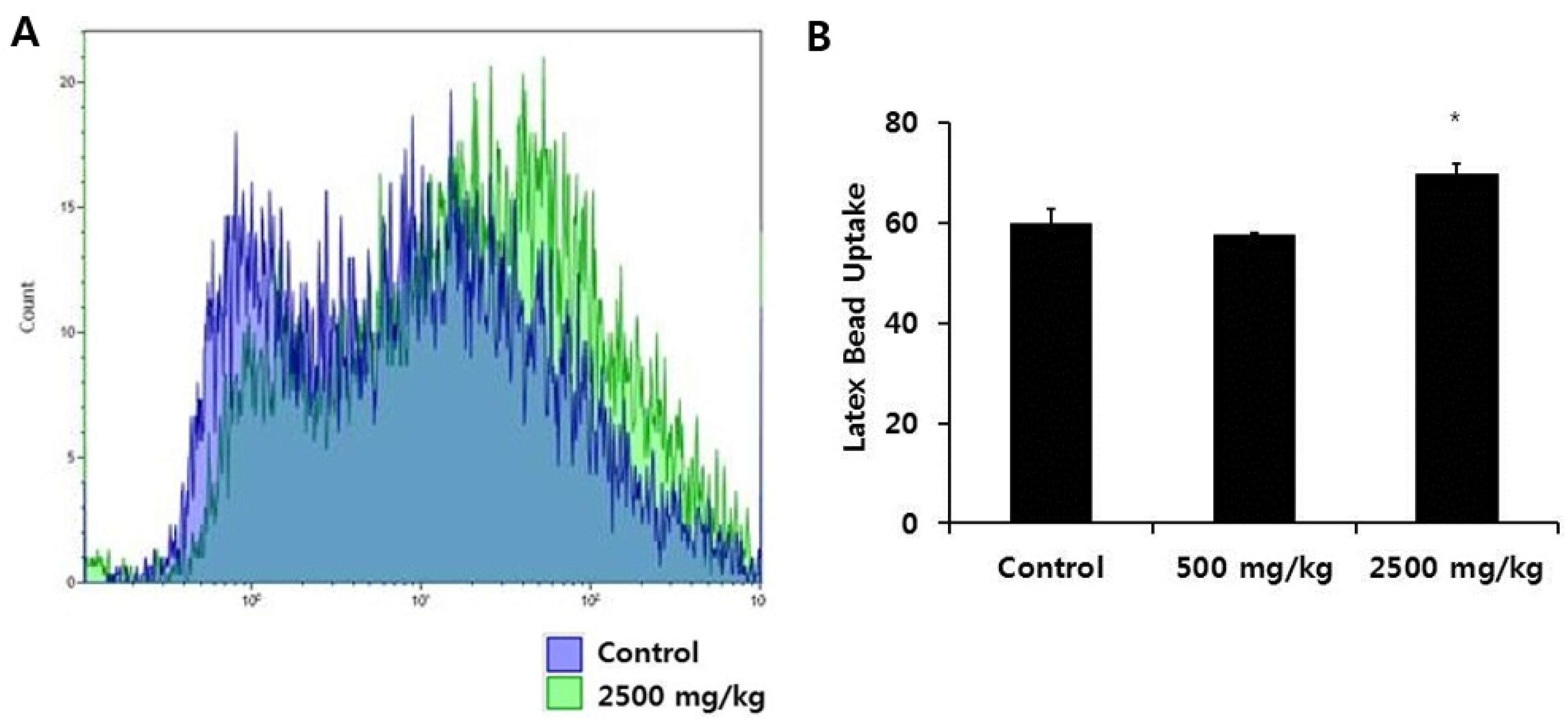
3.2. Effect of Enzymatically-Processed Wheat Bran on Scavenger Receptor Expression and Phagocytic Activity in Macrophages
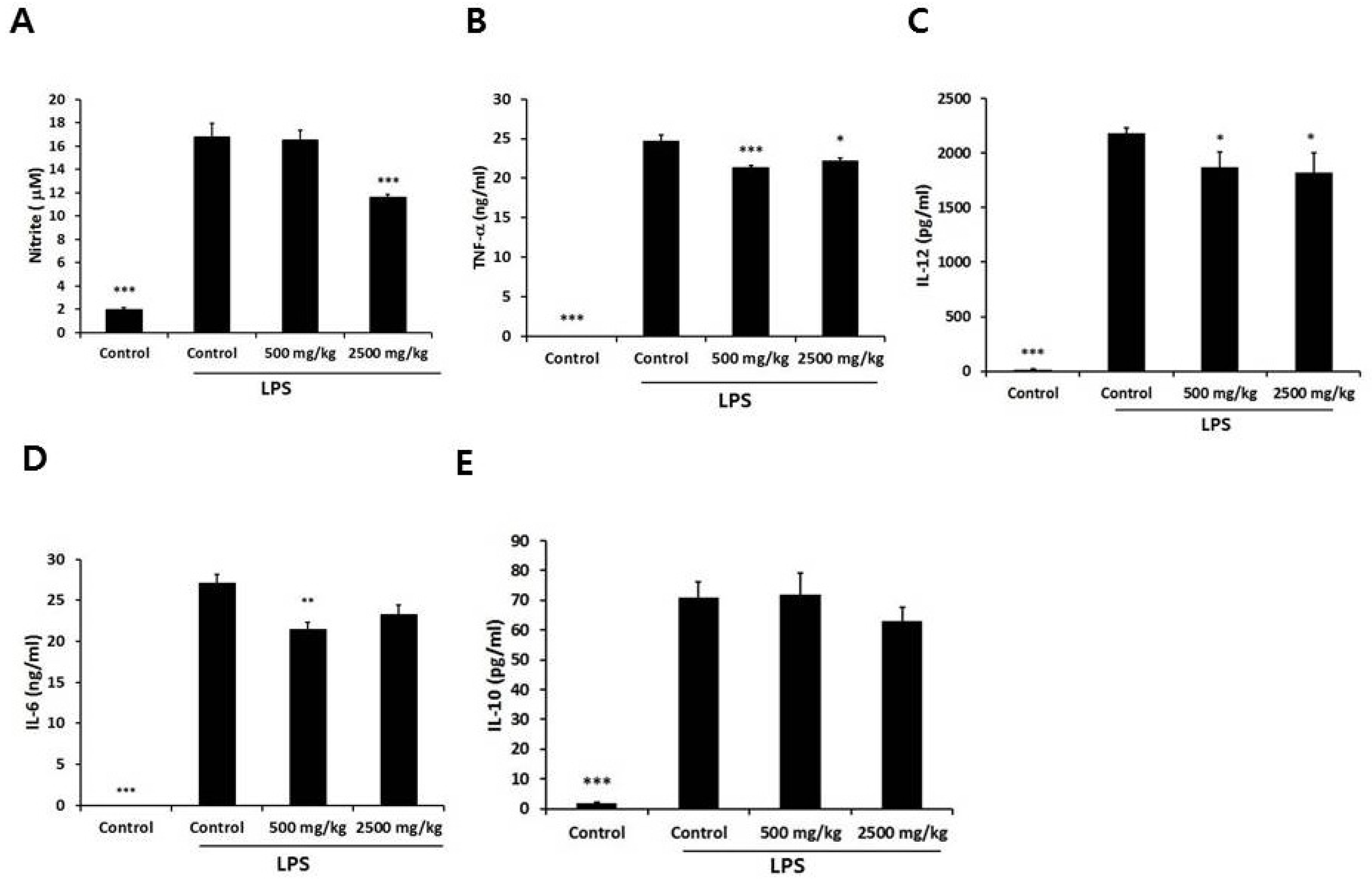
3.3. Effect of Enzymatically-Processed Wheat Bran on NO and Cytokines in LPS-Stimulated Macrophages
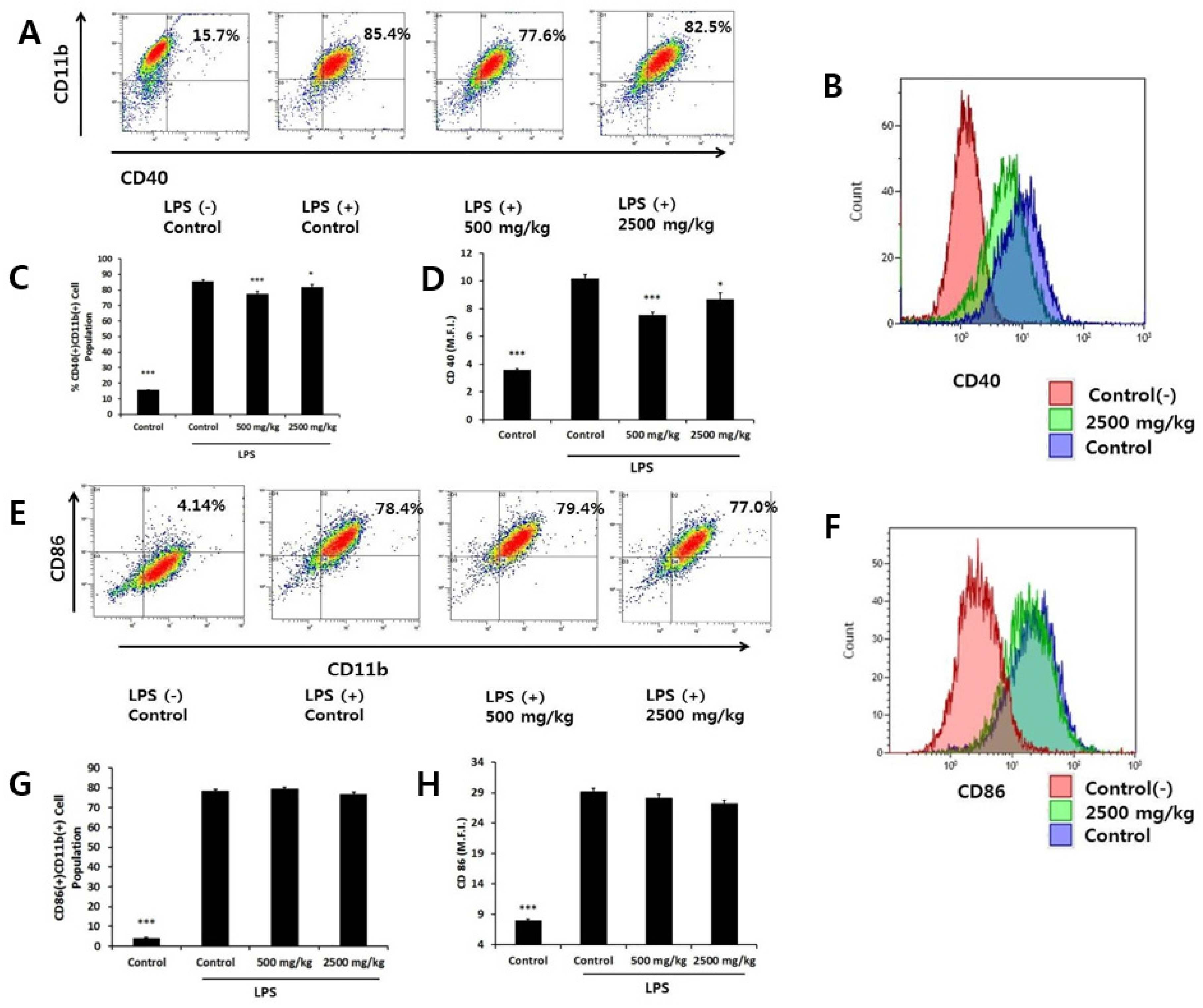
3.4. Effect of Oral Administration of Enzymatically-Processed Wheat Bran on Contact Dependent Inflammatory Molecules in LPS-Activated Macrophages
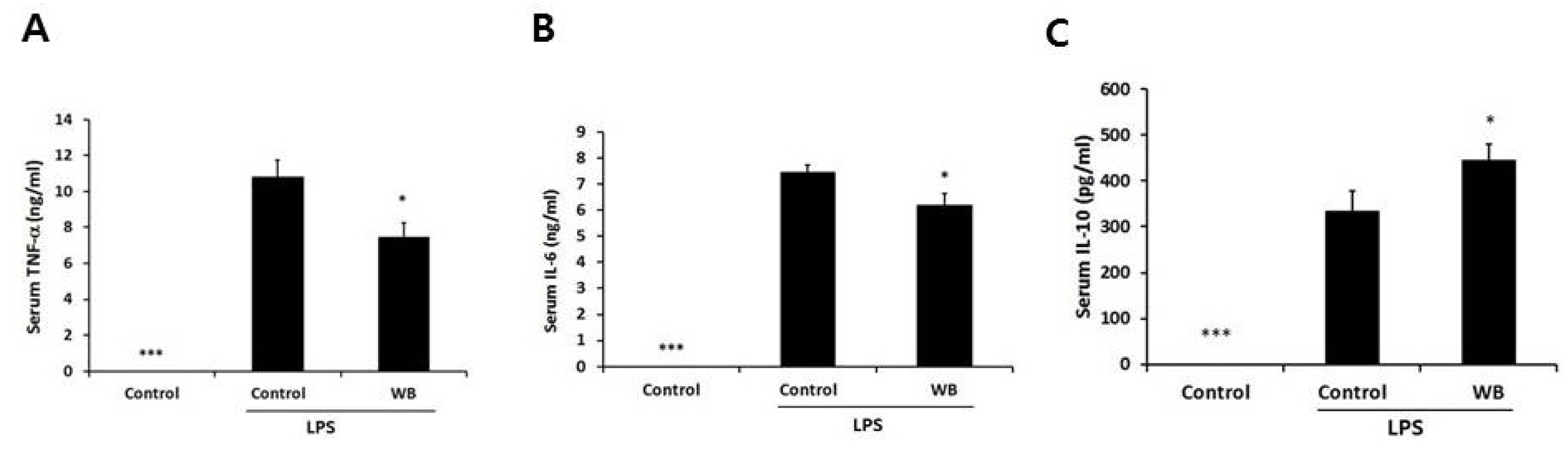
3.5. Effect of Enzymatically-Processed Wheat Bran on Serum Cytokines in Response to Intraperitoneal Injection of LPS
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Maes, C.; Delcour, J.A. Structural characterisation of water-extractable and water-unextractable arabinoxylans in wheat bran. J. Cereal Sci. 2002, 35, 315–326. [Google Scholar] [CrossRef]
- Broekaert, W.F.; Courtin, C.M.; Verbeke, K.; van de Wiele, T.; Verstraete, V.; Delcour, J.A. Prebiotic and other health-related effects of cereal-derived arabinoxylans, arabinoxylan-oligosaccharides, and xylooligosaccharides. Crit. Rev. Food Sci. Nutr. 2011, 51, 178–194. [Google Scholar] [CrossRef] [PubMed]
- Mudgil, D.; Barak, S. Composition, properties and health benefits of indigestible carbohydrate polymers as dietary fiber: A review. Int. J. Biol. Macromol. 2013, 61, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Zoran, D.L.; Turner, N.D.; Taddeo, S.S.; Chapkin, R.S.; Lupton, J.R. Wheat bran diet reduces tumor incidence in a rat model of colon cancer independent of effects on distal luminal butyrate concentrations. J. Nutr. 1997, 127, 2217–2225. [Google Scholar] [PubMed]
- Kritchevsky, D. Protective role of wheat bran fiber: Preclinical data. Am. J. Med. 1999, 106, 28S–31S. [Google Scholar] [CrossRef]
- Neyrinck, A.M.; de Backer, F.; Cani, P.D.; Bindels, L.B.; Stroobants, A.; Portetelle, D.; Delzenne, N.M. Immunomodulatory properties of two wheat bran fractions—Aleurone-enriched and crude fractions—In obese mice fed a high fat diet. Int. Immunopharmacol. 2008, 8, 1423–1432. [Google Scholar] [CrossRef] [PubMed]
- Schepetkin, I.A.; Quinn, M.T. Botanical polysaccharides: Macrophage immunomodulation and therapeutic potential. Int. Immunopharmacol. 2006, 6, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Moretti, J.; Blander, J.M. Insights into phagocytosis-coupled activation of pattern recognition receptors and inflammasomes. Curr. Opin. Immunol. 2014, 26, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, M.; O’Garra, A. The regulation of IL-10 production by immune cells. Nat. Rev. Immunol. 2010, 10, 170–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramberg, J.E.; Nelson, E.D.; Sinnott, R.A. Immunomodulatory dietary polysaccharides: A systematic review of the literature. Nutr. J. 2010, 9, 2031–2132. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Liu, X.; Guo, Y.; Wang, Q.; Peng, D.; Cao, L. Comparison of the immunological activities of arabinoxylans from wheat bran with alkali and xylanse-aided extraction. Carbohydr. Polym. 2010, 81, 784–789. [Google Scholar] [CrossRef]
- AOAC International. Total, solule, and insoluble dietary fibre in foods. In Official Methods of Analysis of AOAC International, 17th ed.; AOAC International: Gaithersburg, MD, USA, 2000; Volume 1. [Google Scholar]
- Douglas, S.G. A rapid method for the determination of pentosans in wheat flour. Food Chem. 1981, 7, 139–145. [Google Scholar] [CrossRef]
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D.; Crocke, D.H.B. Determination of Structural Carbohydrates and Lignin in Biomass; Laboratory Analytical Procedure (LAP): NREL/TP-510-42618; National Renewable Energy Laboratory: Golden, CO, USA, 2011. [Google Scholar]
- Abramson, S.B.; Amin, A.R.; Clancy, R.M.; Attur, M. The role of nitric oxide in tissue destruction, best practice & research. Clin. Rheumatol. 2001, 15, 831–845. [Google Scholar]
- Purswani, M.U.; Eckert, S.J.; Arora, H.K.; Noel, G.J. Effect of ciprofloxacin on lethal and sublethal challenge with endotoxin and on early cytokine responses in a murine in vivo model. J. Antimicrob. Chemother. 2002, 50, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Baublis, A.; Decker, E.A.; Clydesdale, F.M. Antioxidant effect of aqueous extracts from wheat based ready-to-eat breakfast cereals. Food Chem. 2000, 68, 1–6. [Google Scholar] [CrossRef]
- Kim, K.H.; Tsao, R.; Yang, R.; Cui, S.W. Phenolic acid profiles and antioxidant activities of wheat bran extracts and the effect of hydrolysis conditions. Food Chem. 2006, 95, 466–473. [Google Scholar] [CrossRef]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef] [PubMed]
- Canton, J.; Neculai, D.; Grinstein, S. Scavenger receptors in homeostasis and immunity. Nat. Rev. Immunol. 2013, 13, 621–634. [Google Scholar] [CrossRef] [PubMed]
- Freeman, S.A.; Grinstein, S. Phagocytosis: Receptors, signal integration, and the cytoskeleton. Immunol. Rev. 2014, 262, 193–215. [Google Scholar] [CrossRef] [PubMed]
- Bogdan, C. Nitric oxide synthase in innate and adaptive immunity: An update. Trends Immunol. 2015, 36, 161–178. [Google Scholar] [CrossRef] [PubMed]
- Kaiko, G.E.; Horvat, J.C.; Beagley, K.W.; Hansbro, P.M. Immunological decision-making: How does the immune system decide to mount a helper T-cell response? Immunology 2008, 123, 326–338. [Google Scholar] [CrossRef] [PubMed]
- Gaestel, M.; Kotlyarov, A.; Kracht, M. Targeting innate immunity protein kinase signalling in inflammation. Nat. Rev. Drug Discov. 2009, 8, 480–499. [Google Scholar] [CrossRef] [PubMed]
- Suttles, J.; Stout, R.D. Macrophage CD40 signaling: A pivotal regulator of disease protection and pathogenesis. Semin. Immunol. 2009, 21, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Chandel, H.S.; Pandey, S.P.; Shukla, D.; Lalsare, K.; Selvaraj, S.K.; Jha, M.K.; Saha, B. Toll-like receptors and CD40 modulate each other’s expression affecting Leishmania major infection. Clin. Exp. Immunol. 2014, 176, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Edwards, J.P.; Zhang, X.; Frauwirth, K.A.; Mosser, D.M. Biochemical and functional characterization of three activated macrophage populations. J. Leukoc. Biol. 2006, 80, 1298–1307. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.; Taylor, P.R. Monocyte and macrophage heterogeneity. Nat. Rev. Immunol. 2005, 5, 953–964. [Google Scholar] [CrossRef] [PubMed]
- Cloetens, L.; Broekaert, W.F.; Delaedt, Y.; Ollevier, F.; Courtin, C.M.; Delcour, J.A.; Rutgeerts, P.; Verbeke, K.L. Tolerance of arabinoxylan-oligosaccharides and their prebiotic activity in healthy subjects: A randomised, placebo-controlled cross-over study. Br. J. Nutr. 2010, 103, 703–713. [Google Scholar] [CrossRef] [PubMed]
- Francois, I.E.; Lescroart, O.; Veraverbeke, W.S.; Marzorati, M.; Possemiers, S.; Evenepoel, P.; Hamer, H.; Houben, E.; Windey, K.; Welling, G.W.; et al. Effects of a wheat bran extract containing arabinoxylan oligosaccharides on gastrointestinal health parameters in healthy adult human volunteers: A double-blind, randomised, placebo-controlled, cross-over trial. Br. J. Nutr. 2012, 108, 2229–2242. [Google Scholar] [CrossRef] [PubMed]
- Maki, K.C.; Gibson, G.R.; Dickmann, R.S.; Kendall, C.W.; Chen, C.Y.; Costabile, A.; Comelli, E.M.; McKay, D.L.; Almeida, N.G.; Jenkins, D.; et al. Digestive and physiologic effects of a wheat bran extract, arabino-xylan-oligosaccharide, in breakfast cereal. Nutrition 2012, 28, 1115–1121. [Google Scholar] [CrossRef] [PubMed]
- Francois, I.E.; Lescroart, O.; Veraverbeke, W.S.; Marzorati, M.; Possemiers, S.; Hamer, H.; Windey, K.; Welling, G.W.; Delcour, J.A.; Courtin, C.M.; et al. Effects of wheat bran extract containing arabinoxylan oligosaccharides on gastrointestinal parameters in healthy preadolescent children. J. Pediatr. Gastroenterol. Nutr. 2014, 58, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Windey, K.; de Preter, V.; Huys, G.; Broekaert, W.F.; Delcour, J.A.; Louat, T.; Herman, J.; Verbeke, K. Wheat bran extract alters colonic fermentation and microbial composition, but does not affect faecal water toxicity: A randomised controlled trial in healthy subjects. Br. J. Nutr. 2014, 12, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Taipale, T.; Pienihakkinen, K.; Isolauri, E.; Larsen, C.; Brockmann, E.; Alanen, P.; Jokela, J.; Soderling, E. Bifidobacterium animalis subsp. lactis BB-12 in reducing the risk of infections in infancy. Br. J. Nutr. 2011, 105, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Guglielmetti, S.; Mora, D.; Gschwender, M.; Popp, K. Randomised clinical trial: Bifidobacterium bifidum MIMBb75 significantly alleviates irritable bowel syndrome and improves quality of life—A double-blind, placebo-controlled study. Aliment. Pharmacol. Ther. 2011, 33, 1123–1132. [Google Scholar] [CrossRef] [PubMed]
- Hooper, L.V.; Macpherson, A.J. Immune adaptations that maintain homeostasis with the intestinal microbiota. Nat. Rev. Immunol. 2010, 10, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Pekkinen, J.; Rosa, N.N.; Savolainen, O.I.; Keski-Rahkonen, P.; Mykkanen, H.; Poutanen, K.; Micard, V.; Hanhineva, K. Disintegration of wheat aleurone structure has an impact on the bioavailability of phenolic compounds and other phytochemicals as evidenced by altered urinary metabolite profile of diet-induced obese mice. Nutr. Metab. 2014, 11. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.O.; Min, K.J.; Kwon, T.K.; Um, B.H.; Moreau, R.A.; Choi, S.W. Anti-inflammatory activity of hydroxycinnamic acid derivatives isolated from corn bran in lipopolysaccharide-stimulated Raw 264.7 macrophages. Food Chem. Toxicol. 2012, 50, 1309–1316. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Total Dietary Fiber (%) | Soluble Dietary Fiber (%) | Insoluble Dietary Fiber (%) | Water Soluble Arabinoxylan (%) |
|---|---|---|---|---|
| WB | 43.75 ± 0.15 b | 4.62 ± 0.21 a | 39.44 ± 0.13 c | 0.32 ± 0.14 a |
| XWB | 46.49 ± 0.19 c | 4.98 ± 0.16 c | 38.65 ± 0.12 b | 0.71 ± 0.17 b |
| EXWB | 38.65 ± 0.11 a | 4.85 ± 0.14 b | 30.84 ± 0.15 a | 7.14 ± 40.18 c |
| Group | Arabinose (mg/g) | Galactose (mg/g) | Glucose (mg/g) | Xylose (mg/g) |
|---|---|---|---|---|
| WB | 0.04 ± 0.002 a | 0.01 ± 0.002 a | 0.23 ± 0.001 a | 0.08 ± 0.001 a |
| XWB | 0.06 ± 0.004 b | 0.01 ± 0.001 a | 0.30 ± 0.004 b | 0.10 ± 0.004 b |
| EXWB | 0.09 ± 0.004 c | 0.02 ± 0.003 b | 0.34 ± 0.001 c | 0.16 ± 0.002 c |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, H.; Lee, M.-G.; Lee, J.-K.; Choi, Y.-H.; Choi, Y.-S. Enzymatically-Processed Wheat Bran Enhances Macrophage Activity and Has in Vivo Anti-Inflammatory Effects in Mice. Nutrients 2016, 8, 188. https://0-doi-org.brum.beds.ac.uk/10.3390/nu8040188
Kang H, Lee M-G, Lee J-K, Choi Y-H, Choi Y-S. Enzymatically-Processed Wheat Bran Enhances Macrophage Activity and Has in Vivo Anti-Inflammatory Effects in Mice. Nutrients. 2016; 8(4):188. https://0-doi-org.brum.beds.ac.uk/10.3390/nu8040188
Chicago/Turabian StyleKang, Hee, Mi-Gi Lee, Jae-Kang Lee, Yong-Hyun Choi, and Yong-Seok Choi. 2016. "Enzymatically-Processed Wheat Bran Enhances Macrophage Activity and Has in Vivo Anti-Inflammatory Effects in Mice" Nutrients 8, no. 4: 188. https://0-doi-org.brum.beds.ac.uk/10.3390/nu8040188