Effects of Cassava Juice (Manihot esculenta Crantz) on Renal and Hepatic Function and Motor Impairments in Male Rats

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Body Weight
2.2. Behavioral Tests
2.2.1. Open Field Test
Number of Crossings
Time Spent Rearing
Time Spent Grooming
2.2.2. Rotarod
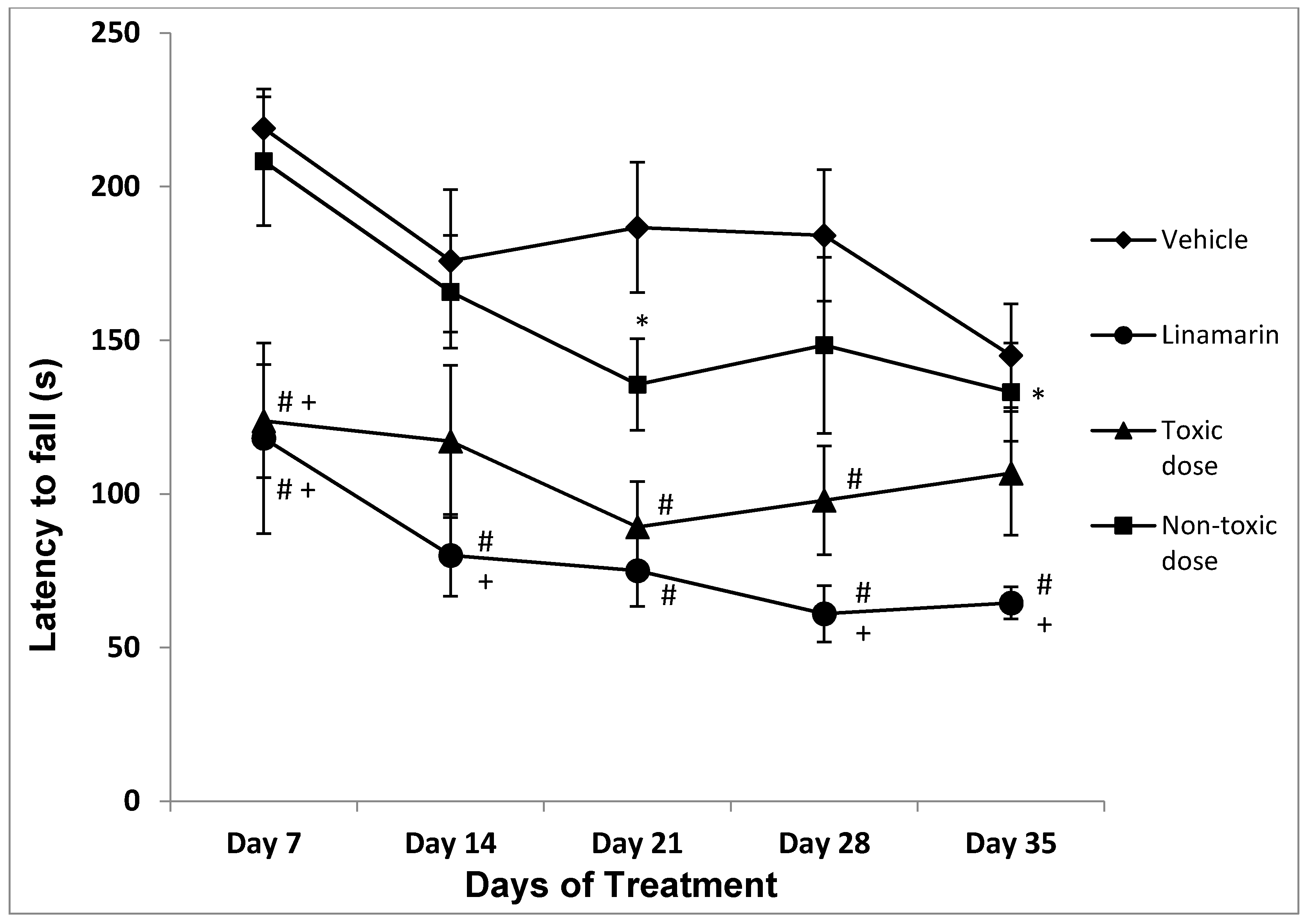
Latency to Fall
2.2.3. Swim Test
Spin Behavior
2.3. Biochemical Tests
2.3.1. Renal Function
2.3.2. Liver Function
2.4. Correlation between Biochemical Measures and Principal Behavioral Variables
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Animals
5.2. Ethical Approval
5.3. Biological Material
5.4. Cassava Juice
5.5. Dose Selection
5.6. Experimental Groups
5.7. Behavioral Tests
5.7.1. Open Field Test
5.7.2. Rotarod
5.7.3. Swim Test
5.8. Blood Samples
5.9. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vladimir-Zaninovic, M.D. Posible asociación de algunas enfermedades neurológicas con el consumo excesivo de la yuca mal procesada y de otros vegetales neurotóxicos. Colomb. Med. 2010, 34, 82–91. [Google Scholar]
- Suarez-Guerra, L.; Mederos-Vega, V.R. Apuntes sobre el cultivo de la yuca (Manihot esculenta Crantz): Tendencias actuales. Cultiv. Trop. 2011, 32, 27–35. [Google Scholar]
- Ceballos, H.; Iglesias, C.A.; Perez, J.C.; Dixon, A.G.O. Cassava breeding: Opportunities and challenges. Plant Mol. Biol. 2004, 56, 503–516. [Google Scholar] [CrossRef]
- Jorgensen, K.; Morant, A.V.; Morant, M.; Jensen, N.B.; Olsen, C.E.; Kannangara, R.; Motawia, M.S.; Moller, B.L.; Bak, S. Biosynthesis of the cyanogenic glucosides linamarin and lotaustralin in cassava: Isolation, biochemical characterization, and expression pattern of CYP71E7, the oxime-metabolizing cytochrome P450 enzyme. Plant Physiol. 2010, 155, 282–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duarte, F.; Sandoval-Castro, C. Efecto de la metionina en una dieta integral de yuca para ovinos en la desintoxicación de ácido cianhídrico. Rev. Biomed. 2002, 13, 116–119. [Google Scholar] [CrossRef]
- Nzwalo, H.; Cliff, J. Konzo: From poverty, cassava, and cyanogen intake to toxico-nutritional neurological disease. PLoS Negl. Trop. Dis. 2011, 5, e1051. [Google Scholar] [CrossRef] [Green Version]
- Tshala-Katumbay, D.; Banea-Mayambu, J.; Kazadi-Kayembe, T.; Nunga, M.; Bikangi, N.; Eeg-Olofsson, K.E.; Tylleskär, T. Neuroepidemiology of konzo aspastic para-tetraparesis of acute onset in a new area of the Democratic Republic of Congo. Afr. J. Neurol. Sci. 2001, 20, 8–12. [Google Scholar]
- Ernesto, M.; Cardoso, A.P.; Nicala, D.; Mirione, E.; Massaza, F.; Cliff, J.; Haque, M.R.; Bradbury, J.H. Persistent konzo and cyanogens toxicity from cassava in northen Mozambique. Acta Trop. 2002, 82, 357–362. [Google Scholar] [CrossRef]
- Howlett, W.P.; Brubaker, G.R.; Mlingi, N.; Rosling, H. Konzo, epidemic upper motor neuron disease studied in Tanzania. Brain 1990, 113, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Rivadeneyra-Domínguez, E.; Rodríguez-Landa, J.F.; Salas-Montero, D.R. Neuropatía atáxica tropical y konzo asociadas al consumo excesivo de yuca. Arch. Neurocienc. 2012, 17, 45–48. [Google Scholar]
- Kittirachra, R. Effects of cassava on thyroid gland in rats. Thai J. Pharm. Sci. 2006, 20, 57–62. [Google Scholar]
- Adam-Shama, I.Y.; Ahmed-Wasma, A.A. Evaluation of the toxicity of Manihot esculenta of Wistar rats after traditional Sudanese processing. J. Pharmacol. Toxicol. 2011, 6, 418–426. [Google Scholar] [CrossRef] [Green Version]
- Mathangi, D.C.; Mohan, N.A. Effect of cassava on motor co-ordination and neurotransmitter level in the albino rat. Food Chem. Toxicol. 1999, 37, 57–60. [Google Scholar] [CrossRef]
- Spencer, P.S.; Palmer, V.S. Interrelationships of undernutrition and neurotoxicity: Food for thought and research attention. Neurotoxicology 2012, 33, 605–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivadeneyra-Dominguez, E.; Vázquez-Luna, A.; Rodríguez-Landa, J.F.; Díaz-Sobac, R. Neurotoxic effect of linamarin in rats associated with cassava (Manihot esculenta Crantz) consumption. Food Chem. Toxicol. 2013, 59, 230–235. [Google Scholar] [CrossRef]
- Rivadeneyra-Domínguez, E.; Rodríguez-Landa, J.F. Preclinical and clinical research on the toxic and neurological effects of cassava (Manihot esculenta Crantz) consumption. Metab. Brain Dis. 2019, 35, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Gerhart, J.M.; Hong, J.S.; Uphouse, L.L.; Tilson, H.A. Chlordecone-induced tremor: Quantification and pharmacological analysis. Toxicol. Appl. Pharmacol. 1982, 66, 234–243. [Google Scholar] [CrossRef]
- Kulig, B.M.; Vanwersch, R.A.P.; Wolthuis, O.L. The automated analysis of coordinated hindlimb movement in rats during acute and prolonged exposure to toxic agents. Toxicol. Appl. Pharmacol. 1985, 80, 1–10. [Google Scholar] [CrossRef]
- Fowler, S.C.; Liao, R.M.; Skjoldager, P. A new rodent model for neuroleptic-induced pseudo-parkinsonism: Low doses of haloperidol increase forelimb tremor in the rat. Behav. Neurosci. 1990, 104, 449–456. [Google Scholar] [CrossRef]
- Kulig, B.; Alleva, E.; Bignami, G.; Cohn, J.; Cory-Slechta, D.; Landa, V.; O’Donoghue, J.; Peakall, D. Animal behavioral methods in neurotoxicity assessment: SGOMSEC Joint Report. Environ. Health Perspect. 1996, 104, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Wieland, S.; Lucki, I. Antidepressant-like activity of 5-HT1A agonists measured with the forced swim test. Psychopharmacology 1990, 101, 497–504. [Google Scholar] [CrossRef]
- Contreras, C.M.; Martínez-Mota, L.; Saavedra, M. Desipramine restricts estral cycle oscillations in swimming. Prog. Neuropsychopharmacol. Biol. Psychiatry 1998, 22, 1121–1128. [Google Scholar] [CrossRef]
- Whishaw, I.Q.; Berg dall, V.; Kolb, B. Analysis of behavior in laboratory rats. In The Laboratory Rat; Krinke, G., Ed.; Academic Press: London, UK, 2000; pp. 359–483. [Google Scholar]
- Wu, Y.; Qiu, A.; Yang, Z.; Wu, J.; Li, X.; Bao, K.; Wang, M.; Wu, B. Malva sylvestris extract alleviates the astrogliosis and inflammatory stress in LPS-induced depression mice. J. Neuroinmun. 2019, 336, 577029. [Google Scholar] [CrossRef]
- Zheng, M.; Liu, C.; Fan, Y.; Shi, D.; Jian, W. Total glucosides of paeony (TGP) extracted from Radix Paeoniae Alba exerts neuroprotective effects in MPTP-induced experimental parkinsonism by regulating the cAMP/PKA/CREB signaling pathway. J. Ethnopharmacol. 2019, 245, 112182. [Google Scholar] [CrossRef] [PubMed]
- Song, S.H.; Jee, Y.S.; Ko, I.G.; Lee, S.W.; Sim, Y.J.; Kim, D.Y.; Lee, S.J.; Cho, Y.S. Treadmill exercise and wheel exercise improve motor function by suppressing apoptotic neuronal cell death in brain inflammation rats. J. Exerc. Rehabil. 2018, 14, 911–919. [Google Scholar] [CrossRef]
- Angelova, V.T.; Rangelov, M.; Todorova, N.; Dangalov, M.; Andreeva-Gateva, P.; Kondeva-Burdina, M.; Karabeliov, V.; Schivachev, B.; Tchekalarova, J. Discovery of novel indole-based arylhydrazones as anticonvulsants: Pharmacophore-based design. Bioorg. Chem. 2019, 90, 103028. [Google Scholar] [CrossRef]
- Cao, W.; Dong, Y.; Zhao, W.; Lu, X.; Sun, L. Mulberrin attenuates 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-induced Parkinson’s disease by promoting Wnt/β-catenin signaling pathway. J. Chem. Neuroanat. 2019, 98, 63–70. [Google Scholar] [CrossRef]
- Coimbra, D.T.; de Oliveira, J.R.G.; Medeiros, M.A.M.B.; e Silva, M.G.; de Andrade, T.R.B.; dos Passos, M.P.; de Sousa, B.M.H.; Abrahao, F.L.; de Souza, A.A.A.; Russo, S.M.; et al. Anticonvulsant, sedative, anxiolytic and antidepressant activities of the essential oil of Annona vepretorum in mice: Involvement of GABAergic and serotonergic systems. Biomed. Pharmacother. 2019, 111, 1074–1087. [Google Scholar] [CrossRef]
- Oliveira-Macêdo, L.A.R.; Pacheco, A.G.M.; Lima-Saraiva, S.R.G.; Silva, J.C.; Oliveira-Júnior, R.G.; Souza, G.R.; Lavor, E.M.; Silva, M.G.; Ferro, J.N.S.; Barreto, E.; et al. Fractions of Selaginella convoluta (Arn.) Spring (Selaginellaceae) attenuate the nociceptive behavior events in mice. Braz. J. Biol. 2020, 80, 57–65. [Google Scholar] [CrossRef]
- Rivadeneyra-Domínguez, E.; Rodríguez-Landa, J.F. Motor impairments induced by microinjection of linamarin in the dorsal hippocampus of Wistar rats. Neurología 2016, 31, 516–522. [Google Scholar] [CrossRef]
- Rivadeneyra-Domínguez, E.; Vázquez-Luna, A.; Díaz-Sobac, R.; Briones-Céspedes, E.E.; Rodríguez-Landa, J.F. Participación del área CA1 del hipocampo en la incoordinación motora inducida por acetonacianohidrina en la rata. Neurología 2017, 32, 230–235. [Google Scholar] [CrossRef]
- Saavedra, M.; Rivadeneyra-Dominguez, E.; Rodriguez-Landa, J.F. Alteraciones motoras inducidas por la microinyección intrahipocampal de metilazoximetanol en ratas macho forzadas a nadar. Arch. Neurocienc. 2011, 4, 186–192. [Google Scholar]
- Rivadeneyra-Domínguez, E.; Rodríguez-Landa, J.F. Las cícadas y su relación con algunas enfermedades neurodegenerativas. Neurología 2013, 29, 517–522. [Google Scholar] [CrossRef]
- Rivadeneyra-Domínguez, E.; Saavedra, M.; Rodríguez-Landa, J.F. El tratamiento con progesterona previene las alteraciones motoras inducidas por la intoxicación con semillas de cícada (Dioon spinulosum) en la rata macho. Rev. Toxicol. 2009, 26, 117–121. [Google Scholar]
- Tellez-Nagel, I.; Korthals, J.K.; Vlassara, H.V.; Cerami, A. An ultrastructural study of chronic sodium cyanate-induced neuropthy. J. Neuropathol. Exp. Neurol. 1977, 36, 352–363. [Google Scholar] [CrossRef]
- Tellez, I.; Johnson, D.; Nagel, R.L.; Cerami, A. Neurotoxicity of sodium cyanate: New pathological and ultrastructural observations in Macaca nemestrina. Acta Neuropathol. 1979, 47, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Sobac, R.; Vázquez-Luna, A.; Rivadeneyra-Domínguez, E.; Rodríguez-Landa, J.F.; Guerrero, T.; Durand-Niconoff, J.S. New paths of cyanogenesis from enzymatic-promoted cleavage of β-cyanoglucosides are suggested by a mixed DFT/QTAIM approach. J. Mol. Model. 2019, 25, 295. [Google Scholar] [CrossRef]
- Tejada-Cifuentes, F. Hepatotoxicidad por Fármacos. Rev. Clin. Med. Fam. 2010, 3, 177–191. [Google Scholar] [CrossRef] [Green Version]
- Martín-Abreu, L.; Martín-Armendáriz, L.G. Fundamentos del Diagnóstico; Méndez Editors: Mexico City, Mexico, 2008; pp. 924–1049. [Google Scholar]
- Baynes, J.Q.; Dominiczak, M.H. Bioquímica Médica; Elsevier: Barcelona, Spain, 2006; pp. 323–417. [Google Scholar]
- Sousa, A.B.; Soto-Blanco, B.; Guerra, J.L.; Kimura, E.T.; Górniak, S.L. Does prolonged oral exposure to cyanide promote hepatotoxicity and nephrotoxicity? Toxicology 2002, 174, 87–95. [Google Scholar] [CrossRef]
- Rivadeneyra-Domínguez, E.; Rosas-Jarquín, C.J.; Vázquez-Luna, A.; Díaz-Sobac, R.; Rodríguez-Landa, J.F. Efecto de la acetonacianohidrina, un derivado de la yuca, sobre la actividad motora y la función renal y hepática en ratas Wistar. Neurología 2019, 34, 300–308. [Google Scholar] [CrossRef]
- Pinzón, I.M.C.; Contreras, H.C.M.; Uribe, R.M. Envenenamiento por cianuro. Rev. Colomb. Psiquiatr. 2002, 31, 330–334. [Google Scholar]
- Ruíz-Reyes, G.; Ruíz-Argüelles, A. Fundamentos de Interpretación Clínica de los Exámenes de Laboratorio; Panamericana: Madrid, Spain, 2010. [Google Scholar]
- Peran-Mesa, S. Introducción a la Bioquímica Clínica; Imagraf: Malaga, Spain, 2007. [Google Scholar]
- Papadakis, M.A.; McPhee, A.J.; Rabow, M.W. Diagnóstico Clínico y Tratamiento; McGraw-Hill: New York, NY, USA, 2015. [Google Scholar]
- Canal, C.; Pellicer, R.; Facundo, C.; Gracia-Garcia, M.B.R.; Ruiz-Garcia, C.; Furlano, M.; Da Silva, I.K.; Ballarin, J.A.; Bover, J. Tablas para la estimación del filtrado glomerular mediante la nueva ecuación CKD-EPI a partir de la concentración de creatinina sérica. Nefrología 2013, 34, 223–229. [Google Scholar] [CrossRef]
- Gutiérrez-Vázquez, I.R. La Fisiopatología como Base Fundamental del Diagnóstico Clínico; Panamericana: Madrid, Spain, 2012; pp. 816–817. [Google Scholar]
- Castaño-Bilbao, I.; Slon-Roblero, M.F.; García-Fernández, N. Estudios de función renal: Función glomerular y tubular—Análisis de orina. Nefrología 2009, 2, 17–30. [Google Scholar]
- Haley, T.J.; Berndt, W.O. Handbook of Toxicology; Hemisphere: Washington, DC, USA, 1987; pp. 251–258. [Google Scholar]
- Rivadeneyra-Domínguez, E.; Vázquez-Luna, A.; Rodríguez-Landa, J.F.; Díaz-Sobac, R. A standardized extract of Ginkgo biloba prevents locomotion impairment induced by cassava juice in Wistar rats. Front. Pharmacol. 2014, 5, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosas-Jarquín, C.J.; Rivadeneyra-Domínguez, E.; León-Chávez, B.A.; Nadela, R.; Sánchez-García, A.C.; Remboa-Bojórquez, D.; Rodríguez-Landa, J.F.; Baltazar-Hernández, D. Chronic consumption of casaba juice induces cellular stress in rat substantia nigra. Iran J. Basic Med. Sci. 2020, 23, 93–101. [Google Scholar] [CrossRef]
- National Research Council. Guide for the Care and Use of Laboratory Animals, 8th ed.; National Academy Press: Washington, DC, USA, 2011. [Google Scholar]
- NOM-062-ZOO. Especificaciones Técnicas para la Producción, Cuidado y Uso de los Animales de Laboratorio; Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación: Mexico City, Mexico, 1999. [Google Scholar]
- Russell, W.M.S.; Burch, R.L.; Hume, C.W. Principles of Humane Experimental Technique; Johns Hopkins Bloomberg School of Public Health: Baltimore, MD, USA, 2019; Available online: http://altweb.jhsph.edu/pubs/books/humane_exp/het-toc (accessed on 30 July 2019).
- Sotocinal, S.G.; Sorge, R.E.; Zaloum, A.; Tuttle, A.H.; Martin, L.J.; Wieskopf, J.S.; Mapplebeck, J.C.S.; Wei, P.; Zhan, S.; Zhang, S.; et al. The rat grimace scale: A partially automated method for quantifying pain in the laboratory rat via facial expressions. Mol. Pain 2011, 7, 1744–8069. [Google Scholar] [CrossRef] [Green Version]
- García-González, M.; Díaz, C.; Villalobos, R. Estudio toxicológico y farmacológico de los extractos hidroalcohólicos de algunas especies de Smilax de Centroamérica. Rev. Fitoter 2008, 8, 49–57. [Google Scholar]
- Donovan, J.; Brown, P. Blood collection. In Current Protocols in Immunology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2006; pp. 1–7. [Google Scholar]
- Suckow, M.A.; Weisbroth, S.H.; Franklin, C.L. The Laboratory Rat, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2006. [Google Scholar]
- Sharp, P.E.; Villano, J.S. The Laboratory Rat, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2013; pp. 385–400. [Google Scholar]


{kind=link}
| Treatments | |||||
|---|---|---|---|---|---|
| Treatment Days | Control | Linamarin | Toxic Doses | Non-Toxic Doses | Treatment Days Factor |
| 7 | 316 ± 4 | 316 ± 9 | 314 ± 6 | 322 ± 6 | 317 ± 3 |
| 14 | 335 ± 4 | 327 ± 11 | 333 ± 6 | 339 ± 7 | 334 ± 3 * |
| 21 | 344 ± 5 | 329 ± 18 | 348 ± 5 | 351 ± 6 | 344 ± 4 * |
| 28 | 357 ± 5 | 353 ± 10 | 361 ± 6 | 363 ± 7 | 359 ± 3 *,# |
| 35 | 372 ± 5 | 371 ± 11 | 377 ± 5 | 377 ± 7 | 374 ± 3 *,#,$ |
| Treatment factor | 345 ± 4 | 339 ± 6 | 346 ± 4 | 350 ± 4 | |
| Treatments | ||||||
|---|---|---|---|---|---|---|
| Test | Treatment Days | Control | Linamarin | Toxic Doses | Non-Toxic Doses | Total |
| Crossing (n) | 7 | 48.6 ± 4.7 | 57.8 ± 4.0 | 54.4 ± 6.4 | 60.1 ± 5.2 | 55.3 ± 2.6 |
| 14 | 41.6 ± 5.1 | 38.6 ± 6.4 | 52.9 ± 3.4 | 45.1 ± 7.4 | 45.1 ± 2.9 * | |
| 21 | 33.5 ± 6.8 | 32.4 ± 3.9 | 29.4 ± 5.1 | 31.2 ± 5.9 | 31.5 ± 2.7 * | |
| 28 | 25.4 ± 2.3 | 60.2 ± 10.2 | 30.7 ± 5.9 | 36.6 ± 7.1 | 37.1 ± 3.9 * | |
| 35 | 27.0 ± 5.0 | 40.7 ± 9.3 | 27.6 ± 2.9 | 31.6 ± 5.9 | 31.3 ± 2.9 * | |
| Total | 35.2 ± 2.6 | 45.9 ± 3.6 | 39.0 ± 2.8 | 40.9 ± 3.2 | ||
| Rearing (s) | 7 | 65.3 ± 8.6 | 63.7 ± 9.7 | 65.8 ± 6.8 | 63.4 ± 14.8 | 64.6 ± 5.0 |
| 14 | 40.4 ± 11.1 | 39.2± 7.2 | 59.6 ± 5.8 | 38.1 ± 7.0 | 44.8 ± 4.1 * | |
| 21 | 21.8 ± 7.0 | 41.5 ± 7.4 | 38.5 ± 4.0 | 33.1 ± 4.6 | 33.6 ± 3.0 * | |
| 28 | 20.8 ± 4.4 | 22.9 ± 8.8 * | 31.6 ± 6.1 | 30.6 ± 4.3 | 26.9 ± 2.9 * | |
| 35 | 35.8 ± 4.7 | 28.4± 6.9 * | 26.4 ± 6.1 | 26.6 ± 4.2 | 29.1 ± 2.3 * | |
| Total | 36.8 ± 4.2 | 39.2 ± 4.2 | 44.4 ± 3.4 | 38.4 ± 3.9 | ||
| Self-grooming (s) | 7 | 32.8 ± 7.1 | 72.1 ± 3.8 | 56.1 ± 13.8 | 71.4 ± 13.4 | 58.0 ± 6.1 |
| 14 | 44.0 ± 19.5 | 68.4 ± 19.1 | 45.6 ± 12.8 | 43.7 ± 12.9 | 49.4 ± 7.7 | |
| 21 | 22.7 ± 12.8 | 51.2 ± 15.4 | 61.7 ± 16.0 | 60.2 ± 12.0 | 49.7 ± 7.3 | |
| 28 | 13.1 ± 4.7 | 28.2 ± 7.6 | 30.8 ± 15.0 | 36.2 ± 10.7 | 27.5 ± 5.4 * | |
| 35 | 13.5 ± 4.8 | 26.7 ± 12.0 | 30.5 ± 14.2 | 27.5 ± 9.4 | 24.7 ± 5.3 * | |
| Total | 25.2 ± 5.1 | 49.3 ± 6.4 + | 44.9 ± 6.4 + | 47.8 ± 5.6 + | ||
| Treatments | |||||
|---|---|---|---|---|---|
| Test | Treatment Days | Control | Non-Toxic Doses | Linamarin | Toxic Doses |
| Spin behavior (n) | 7 | 0 | 0 | 2.8 | 3.4 |
| 14 | 0 | 0 | 3.0 | 6.2 | |
| 21 | 0 | 0 | 5.0 | 6.5 | |
| 28 | 0 | 0 | 5.6 | 6.3 | |
| 35 | 0 | 0 | 9.2 | 8.5 | |
| Treatment | ||||
|---|---|---|---|---|
| Analyte | Control | Linamarin | Toxic Dose | Non-Toxic Dose |
| Glucose (mmol/L) | 6.0 ± 0.2 | 7.6 ± 0.1 * | 9.4 ± 0.1 * | 6.2 ± 0.1 |
| Urea (mmol/L) | 16.8 ± 1.1 | 21.7 ± 0.6 * | 29.8 ± 1.2 * | 18.0 ± 0.2 |
| Creatinine (µmol/L) | 10.1 ± 0.5 | 58.8 ± 0.8 * | 66.3 ± 1.0 * | 13.2 ± 0.2 * |
| Blood Urea Nitrogen (mmol/L) | 6.3 ± 0.1 | 8.1 ± 0.2 * | 8.6 ± 0.1 * | 6.2 ± 0.1 |
| Treatment | ||||
|---|---|---|---|---|
| Analyte | Control | Linamarin | Toxic Dose | Non-Toxic Dose |
| Aspartate Amino Transferase (UI/L) | 93.80 ± 1.49 | 158.25 ± 10.51 * | 188.26 ± 1.93 * | 79.81 ± 2.26 * |
| Alanine Amino Transferase (UI/L) | 49.21 ± 2.39 | 67.08 ± 1.42 * | 67.47 ± 1.04 * | 43.17 ± 3.38 |
| Alkaline Phosphatase (UI/L) | 242.85 ± 10.27 | 381.66 ± 2.69 * | 422.62 ± 33.15 * | 234.00 ± 10.69 |
| Total Bilirubin (mg/dL) | 0.39 ± 0.01 | 0.67 ± 0.01 * | 0.67 ± 0.01 * | 0.39 ± 0.01 |
| Indirect Bilirubin (mg/dL) | 0.02 ± 0.01 | 0.23 ± 0.01 * | 0.19 ± 0.02 * | 0.01 ± 0.01 |
| Direct Bilirubin (mg/dL) | 0.01 ± 0.01 | 0.14 ± 0.01 * | 0.25 ± 0.03 * | 0.05 ± 0.02 |
| Total Proteins (g/dL) | 5.38 ± 0.04 | 3.66 ± 0.39 * | 4.33 ± 0.10 * | 5.32 ± 0.08 |
| Albumin (g/dL) | 4.15 ± 0.06 | 2.96 ± 0.29 * | 3.10 ± 0.16 * | 4.16 ± 0.05 |
| γ-Glutamyl-Transferase | 23.28 ± 0.86 | 83.50 ± 1.25 * | 95.25 ± 1.19 * | 23.62 ± 0.37 |
| Biochemical Measures | Behavioral Variables | ||
|---|---|---|---|
| Crossing (n) | Latency to Fall (s) | Spin Behavior (n) | |
| Renal function | |||
| Glucose (mmol/L) | r −0.001 | r −0.304 | r 0.673 * |
| Urea (mmol/L) | r −0.033 | r −0.299 | r 0.594 * |
| Creatinine (µmol/L) | r 0.072 | r −0.012 * | r 0.793 * |
| Blood Urea Nitrogen (mmol/L) | r 0.020 | r −0.036 * | r 0.687 * |
| Liver function | |||
| Aspartate Amino Transferase (UI/L) | r 0.024 | r −0.384 * | r 0.078 * |
| Alanine Amino Transferase (UI/L) | r 0.038 | r −0.372 * | r 0.633 * |
| Alkaline Phosphatase (UI/L) | r 0.092 | r −0.415 * | r 0.644 * |
| Total Bilirubin (mg/dL) | r 0.118 | r −0.398 * | r 0.738 * |
| Indirect Bilirubin (mg/dL) | r 0.170 | r −0.434 * | r 0.792 * |
| Direct Bilirubin (mg/dL) | r −0.211 | r 0.354 | r −0.568 * |
| Total proteins(g/dL) | r −0.211 | r 0.354 | r −0.568 * |
| Albumin (g/dL) | r −0.268 | r 0.312 | r −0.633 * |
| γ-Glutamyl-Transferase | r 0.083 | r −0.445 * | r 0.804 * |
| Analyte | Reference Intervals |
|---|---|
| Glucose | 6–10 mmol/L |
| Creatinine | 11–28 µmol/L |
| Urea | 10.7–20 mmol/L |
| Blood Urea Nitrogen | 3–7 mmol/L |
| Total proteins | 5–7 g/dL |
| Albumin | 4–5 g/dL |
| Total bilirubin | 0.18–0.54 mg/dL |
| Direct bilirubin | 0.03–0.06 mg/dL |
| Indirect bilirubin | 0–0.1 mg/dL |
| Alkaline Phosphatase | 36–312 UI/L |
| γ-Glutamil-Transferase | 8.8–24 UI/L |
| Aspartate Amino Transferase | 63–157 UI/L |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivadeneyra-Domínguez, E.; Pérez-Pérez, J.E.; Vázquez-Luna, A.; Díaz-Sobac, R.; Rodríguez-Landa, J.F. Effects of Cassava Juice (Manihot esculenta Crantz) on Renal and Hepatic Function and Motor Impairments in Male Rats. Toxins 2020, 12, 708. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12110708
Rivadeneyra-Domínguez E, Pérez-Pérez JE, Vázquez-Luna A, Díaz-Sobac R, Rodríguez-Landa JF. Effects of Cassava Juice (Manihot esculenta Crantz) on Renal and Hepatic Function and Motor Impairments in Male Rats. Toxins. 2020; 12(11):708. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12110708
Chicago/Turabian StyleRivadeneyra-Domínguez, Eduardo, José Eduardo Pérez-Pérez, Alma Vázquez-Luna, Rafael Díaz-Sobac, and Juan Francisco Rodríguez-Landa. 2020. "Effects of Cassava Juice (Manihot esculenta Crantz) on Renal and Hepatic Function and Motor Impairments in Male Rats" Toxins 12, no. 11: 708. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12110708