Whole Genome Analysis Revealed the Genes Responsible for Citreoviridin Biosynthesis in Penicillium citreonigrum

Abstract
:1. Introduction
2. Results
2.1. Whole Genome Sequence
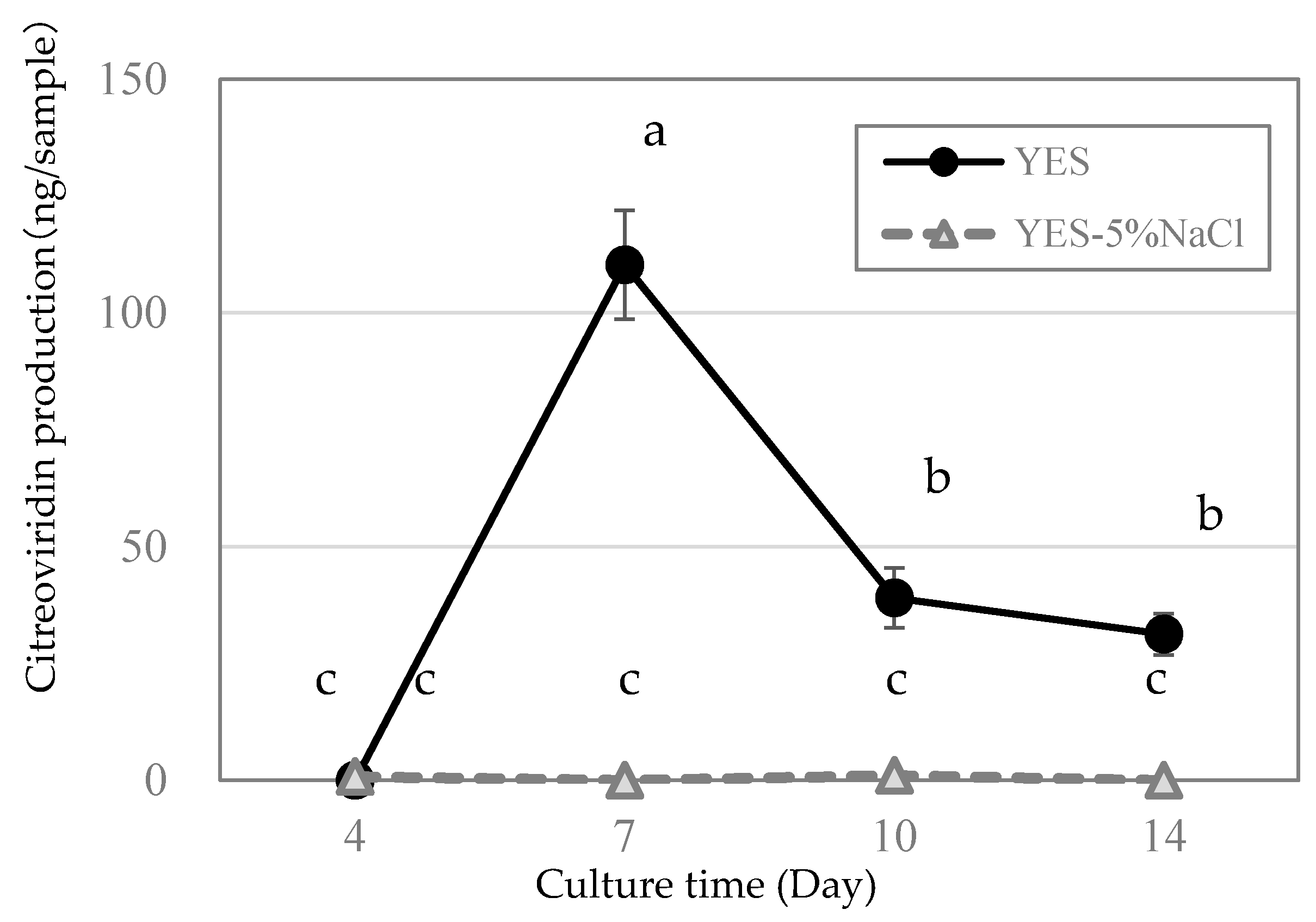
2.2. Citreoviridin Production Profile
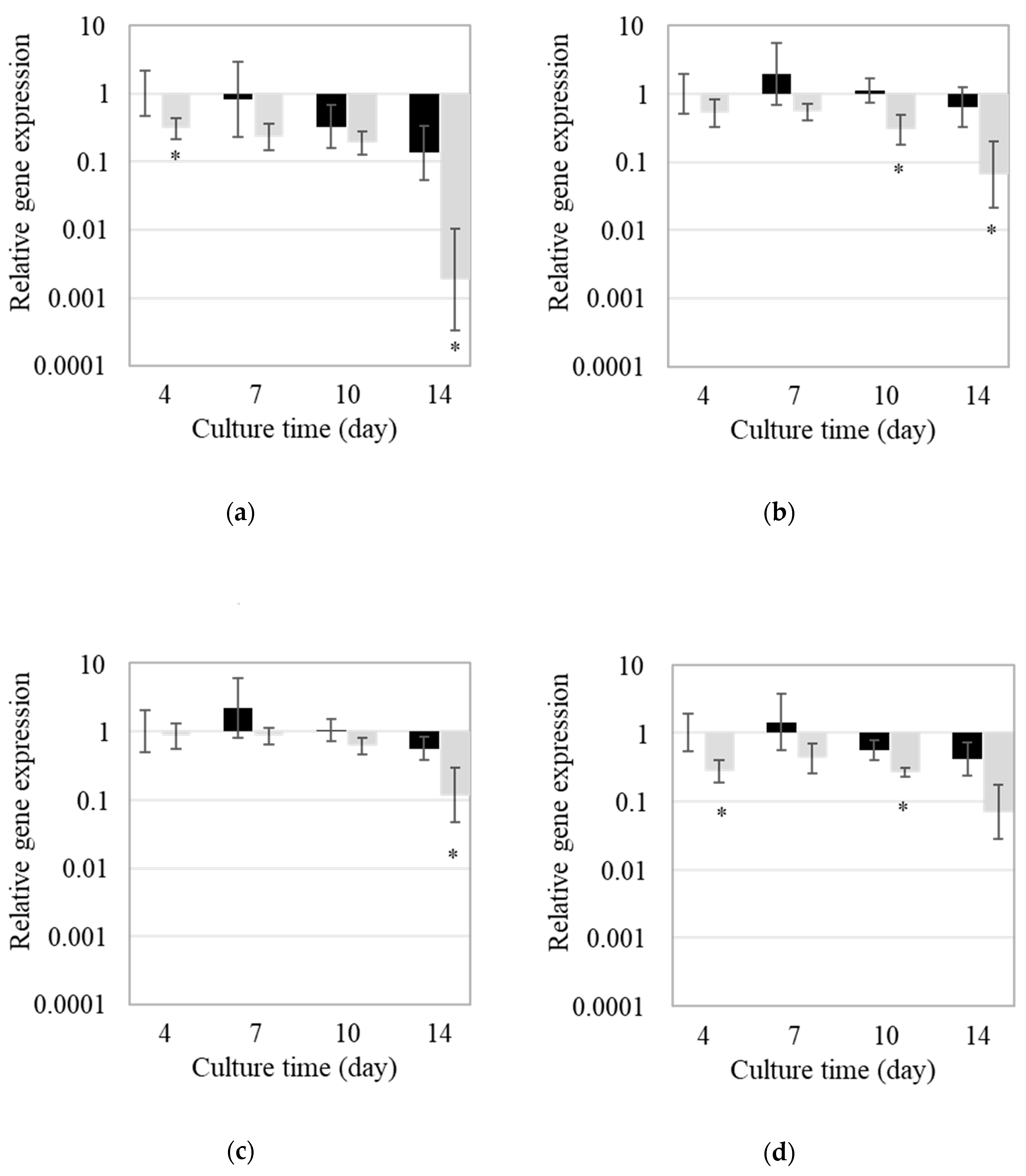
2.3. Gene Expression
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Strain
5.2. Whole Genome Sequencing
5.3. Citreoviridin Production
5.4. LC–MS/MS Conditions
5.5. Gene Expression
5.6. Statistical Analyses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shiratori, N.; Kobayashi, N.; Tulayakul, P.; Sugiura, Y.; Takino, M.; Endo, O.; Sugita-Konishi, Y. Occurrence of Penicillium brocae and Penicillium citreonigrum, which produce a mutagenic metabolite and a mycotoxin citreoviridin, respectively, in selected commercially available rice grains in Thailand. Toxins 2017, 9, 194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, H.C.A.V.; Porto, E.A.S.; Marins, J.R.P.; Alves, R.M.; Machado, R.R.; Braga, K.N.L.; de Paiva, F.B.; Carmo, G.M.I.; Faria Silva e Santelli, A.C.; Sobel, J. Outbreak of beriberi in the state of Maranhão, Brazil: Revisiting the mycotoxin aetiologic hypothesis. Trop. Doct. 2010, 40, 95–97. [Google Scholar] [CrossRef] [PubMed]
- Wicklow, D.T.; Cole, R.J. Citreoviridin in standing corn infested by Eupenicillium ochrosalmoneum. Mycologia 1984, 76, 959–961. [Google Scholar] [CrossRef]
- Morrissey, R.E.; Vesonder, R.F. Teratogenic potential of the mycotoxin, citreoviridin, in rats. Food Chem. Toxicol. 1986, 24, 1315–1320. [Google Scholar] [CrossRef]
- Datta, S.C.; Ghosh, J.J. Effect of citreoviridin, a mycotoxin from Penicillium citreoviride, on kinetic constants of acetylcholinesterase and ATPase in synaptosomes and microsomes from rat brain. Toxicon 1981, 19, 555–562. [Google Scholar] [CrossRef]
- Ueno, Y. Temperature-dependent production of citreoviridin, a neurotoxin of Penicillium citreo-viride Biourge. Jpn. J. Exp. Med. 1972, 42, 107–114. [Google Scholar] [PubMed]
- Kushiro, M. Historical review of researches on yellow rice and mycotoxigenic fungi adherent to rice in Japan. JSM Mycotoxins 2015, 65, 19–23. [Google Scholar] [CrossRef] [Green Version]
- Padilha, E.M.; Fujimori, E.; Borges, A.L.V.; Sato, A.P.S.; Gomes, M.N.; dos Remédios Freitas Carvalho Branco, M.; dos Santos, H.J.; Junior, N.L. Perfil epidemiológico do beribéri notificado de 2006 a 2008 no Estado do Maranhão, Brasil. Cad. Saude Publica 2011, 27, 449–459. [Google Scholar] [CrossRef]
- Da Rocha, M.W.; Resck, I.S.; Caldas, E.D. Purification and full characterisation of citreoviridin produced by Penicillium citreonigrum in yeast extract sucrose (YES) medium. Food Addit. Contam. Part A Chem. Anal. Control. Exp. Risk Assess. 2015, 32, 584–595. [Google Scholar] [CrossRef]
- Sun, S. Chronic exposure to cereal mycotoxin likely citreoviridin may be a trigger for Keshan disease mainly through oxidative stress mechanism. Med. Hypotheses 2010, 74, 841–842. [Google Scholar] [CrossRef]
- Hou, H.; Zhou, R.; Jia, Q.; Li, Q.; Kang, L.; Jiao, P.; Li, D.; Jiang, B. Citreoviridin enhances tumor necrosis factor-α-induced adhesion of human umbilical vein endothelial cells. Toxicol. Ind. Health 2015, 31, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Sakabe, N.; Goto, T.; Hirata, Y. The structure of citreoviridin, a toxic compound produced by P. citreoviride molded on rice. Tetrahedron Lett. 1964, 5, 1825–1830. [Google Scholar] [CrossRef]
- Linnett, P.E.; Mitchell, A.D.; Osselton, M.D.; Mulheirn, L.J.; Beechey, R.B. Citreoviridin, a specific inhibitor of the mitochondrial adenosine triphosphatase. Biochem. J. 1978, 170, 503–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayood, S.F.; Suh, H.; Wilcox, C.S.; Schuster, S.M. Effect of citreoviridin and isocitreoviridin on beef heart mitochondrial ATPase. Arch. Biochem. Biophys. 1989, 270, 714–721. [Google Scholar] [CrossRef]
- Cataldi de Flombaum, M.A.; Stoppani, A.O.M. Influence of efrapeptin, aurovertin and citreoviridin on the mitochondrial adenosine triphosphatase from Trypanosoma cruzi. Mol. Biochem. Parasitol. 1981, 3, 143–155. [Google Scholar] [CrossRef]
- Lin, T.S.; Chiang, Y.M.; Wang, C.C.C. Biosynthetic pathway of the reduced polyketide product citreoviridin in Aspergillus terreus var. aureus revealed by heterologous expression in Aspergillus nidulans. Org. Lett. 2016, 18, 1366–1369. [Google Scholar] [CrossRef]
- Tang, X.; Li, J.; Millán-Aguiñaga, N.; Zhang, J.J.; O’Neill, E.C.; Ugalde, J.A.; Jensen, P.R.; Mantovani, S.M.; Moore, B.S. Identification of thiotetronic acid antibiotic biosynthetic pathways by target-directed genome mining. ACS Chem. Biol. 2015, 10, 2841–2849. [Google Scholar] [CrossRef]
- Regueira, T.B.; Kildegaard, K.R.; Hansen, B.G.; Mortensen, U.H.; Hertweck, C.; Nielsen, J. Molecular basis for mycophenolic acid biosynthesis in Penicillium brevicompactum. Appl. Environ. Microbiol. 2011, 77, 3035–3043. [Google Scholar] [CrossRef] [Green Version]
- Osbourn, A. Secondary metabolic gene clusters: Evolutionary toolkits for chemical innovation. Trends Genet. 2010, 26, 449–457. [Google Scholar] [CrossRef]
- Keller, N.P.; Turner, G.; Bennett, J.W. Fungal secondary metabolism—From biochemistry to genomics. Nat. Rev. Microbiol. 2005, 3, 937–947. [Google Scholar] [CrossRef]
- Yu, J.; Chang, P.K.; Ehrlich, K.C.; Cary, J.W.; Bhatnagar, D.; Cleveland, T.E.; Payne, G.A.; Linz, J.E.; Woloshuk, C.P.; Bennett, J.W. Clustered pathway genes in aflatoxin biosynthesis. Appl. Environ. Microbiol. 2004, 70, 1253–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payne, G.A.; Nierman, W.C.; Wortman, J.R.; Pritchard, B.L.; Brown, D.; Dean, R.A.; Bhatnagar, D.; Cleveland, T.E.; Machida, M.; Yu, J. Whole genome comparison of Aspergillus flavus and A. oryzae. Med. Mycol. 2006, 44, S9–S11. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Jurick, W.M.; Cao, H.; Yin, Y.; Gaskins, V.L.; Losada, L.; Zafar, N.; Kim, M.; Bennett, J.W.; Nierman, W.C. Draft Genome Sequence of Penicillium expansum Strain R19, Which Causes Postharvest Decay of Apple Fruit. Genome Announc. 2014, 2, e00635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Wu, G.; Jurick, W.M.; Gaskins, V.L.; Yin, Y.; Yin, G.; Bennett, J.W.; Shelton, D.R. Genome sequence of Penicillium solitum RS1, which causes postharvest apple decay. Genome Announc. 2016, 4, e00363. [Google Scholar] [CrossRef] [Green Version]
- Van Den Berg, M.A.; Albang, R.; Albermann, K.; Badger, J.H.; Daran, J.M.; Driessen, A.J.; Garcia-Estrada, C.; Fedorova, N.D.; Harris, D.M.; Heijne, W.H.M.; et al. Genome sequencing and analysis of the filamentous fungus Penicillium chrysogenum. Nat. Biotechnol. 2008, 26, 1161–1168. [Google Scholar] [CrossRef] [Green Version]
- Mardones, W.; Di Genova, A.; Cortés, M.P.; Travisany, D.; Maass, A.; Eyzaguirre, J. The genome sequence of the soft-rot fungus Penicillium purpurogenum reveals a high gene dosage for lignocellulolytic enzymes. Mycology 2018, 9, 59–69. [Google Scholar] [CrossRef] [Green Version]
- Carbone, I.; Ramirez-Prado, J.H.; Jakobek, J.L.; Horn, B.W. Gene duplication, modularity and adaptation in the evolution of the aflatoxin gene cluster. BMC Evol. Biol. 2007, 7, 111. [Google Scholar] [CrossRef] [Green Version]
- Brown, D.W.; Yu, J.H.; Kelkar, H.S.; Fernandes, M.; Nesbitt, T.C.; Keller, N.P.; Adams, T.H.; Leonard, T.J. Twenty-five coregulated transcripts define a sterigmatocystin gene cluster in Aspergillus nidulans. Proc. Natl. Acad. Sci. USA 1996, 93, 1418–1422. [Google Scholar] [CrossRef] [Green Version]
- Quistgaard, E.M.; Löw, C.; Guettou, F.; Nordlund, P. Understanding transport by the major facilitator superfamily (MFS): Structures pave the way. Nat. Rev. Mol. Cell Biol. 2016, 17, 123–132. [Google Scholar] [CrossRef]
- Wuchiyama, J.; Kimura, M.; Yamaguchi, I. A Trichothecene Efflux Pump Encoded by Tri102 in the Biosynthetic Gene Cluster of Fusarium graminearum. J. Antibiot. 2000, 53, 196–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, N.J.; McCormick, S.P.; Hohn, T.M. TRI12, a trichothecene efflux pump from Fusarium sporotrichioides: Gene isolation and expression in yeast. Mol. Gen. Genet. 1999, 261, 977–984. [Google Scholar] [CrossRef] [PubMed]
- Kajitani, R.; Toshimoto, K.; Noguchi, H.; Toyoda, A.; Ogura, Y.; Okuno, M.; Yabana, M.; Harada, M.; Nagayasu, E.; Maruyama, H.; et al. Efficient de novo assembly of highly heterozygous genomes from whole-genome shotgun short reads. Genome Res. 2014, 24, 1384–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanke, M.; Morgenstern, B. AUGUSTUS: A web server for gene prediction in eukaryotes that allows user-defined constraints. Nucleic Acids Res. 2005, 33, W465–W467. [Google Scholar] [CrossRef] [Green Version]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Yin, H.B.; Chen, C.H.; Kollanoor-Johny, A.; Darre, M.J.; Venkitanarayanan, K. Controlling Aspergillus flavus and Aspergillus parasiticus growth and aflatoxin production in poultry feed using carvacrol and trans-cinnamaldehyde. Poult. Sci. 2015, 94, 2183–2190. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Number of Scaffolds | Minimum Length of Scaffold | Maximum Length of Scaffold | N50 | Sum of Length |
|---|---|---|---|---|
| 79 | 503 bp | 2,963,310 bp | 1,448,320 bp | 27,997,905 bp |
| Query Gene 1 | Hit Open Reading Frame (ORF) | E-value | Identity (%) | Coverage (%) | |||||
|---|---|---|---|---|---|---|---|---|---|
| Name | Size (aa) | Number | Size (aa) | Scaffold | Direction | Position in Scaffold | |||
| ctvA | 2436 | g1460 | 2513 | 16 | + | 519,757..527,298 | 0 | 70.4 | 99.9 |
| ctvB | 228 | g1459 | 233 | 16 | - | 518,431..519,132 | 3E-143 | 79.9 | 100 |
| ctvC | 473 | g1456 | 472 | 16 | - | 512,588..514,173 | 0 | 77.4 | 97.9 |
| ctvD | 354 | g1458 | 341 | 16 | - | 517,137..518,229 | 8E-146 | 59.9 | 99.7 |
| ctvE | 468 | g2666 | 519 | 19 | + | 939,425..941,478 | 0 | 79.4 | 95.9 |
| Gene | Primer Name | Sequence (5′ to 3′) | Reference |
|---|---|---|---|
| ctvA | ctvA_1F | AGCGTGGCATGATTACACCAAACC | this study |
| ctvA_1R | CAACGTCGGCCATTGAAGAACCTC | ||
| ctvB | ctvB_1F | TGTCTGAGAAAGGCTGCCAATCGTG | this study |
| ctvB_1R | CAGCACGTACATCAGCGAGATGGA | ||
| ctvC | ctvC_1F | CCAATACCGCCATGGAAGCAG | this study |
| ctvC_1R | GCAAGCGCTCGTTCAATCGTATC | ||
| ctvD | ctvD_1F | GGCGAATCTCTTGGCAGACATC | this study |
| ctvD_1R | CCACAGCAAGAAACCACTCATCC | ||
| ctvE | ctvE_1F | GTGACTCCAAGGTGTCTCTG | this study |
| ctvE_1R | CAATGAAGAGCAGCACATCC | ||
| β-tubulin | β-tublinF | CGTGTCGGCGACCAGTTC | [37] |
| β-tublinR | CCTCACCAGTGTACCAATGCA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okano, T.; Kobayashi, N.; Izawa, K.; Yoshinari, T.; Sugita-Konishi, Y. Whole Genome Analysis Revealed the Genes Responsible for Citreoviridin Biosynthesis in Penicillium citreonigrum. Toxins 2020, 12, 125. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12020125
Okano T, Kobayashi N, Izawa K, Yoshinari T, Sugita-Konishi Y. Whole Genome Analysis Revealed the Genes Responsible for Citreoviridin Biosynthesis in Penicillium citreonigrum. Toxins. 2020; 12(2):125. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12020125
Chicago/Turabian StyleOkano, Takumi, Naoki Kobayashi, Kazuki Izawa, Tomoya Yoshinari, and Yoshiko Sugita-Konishi. 2020. "Whole Genome Analysis Revealed the Genes Responsible for Citreoviridin Biosynthesis in Penicillium citreonigrum" Toxins 12, no. 2: 125. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12020125