Genomic and Metabolomic Analyses of Natural Products in Nodularia spumigena Isolated from a Shrimp Culture Pond

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Biosynthetic Potential
2.2. Metabolomics
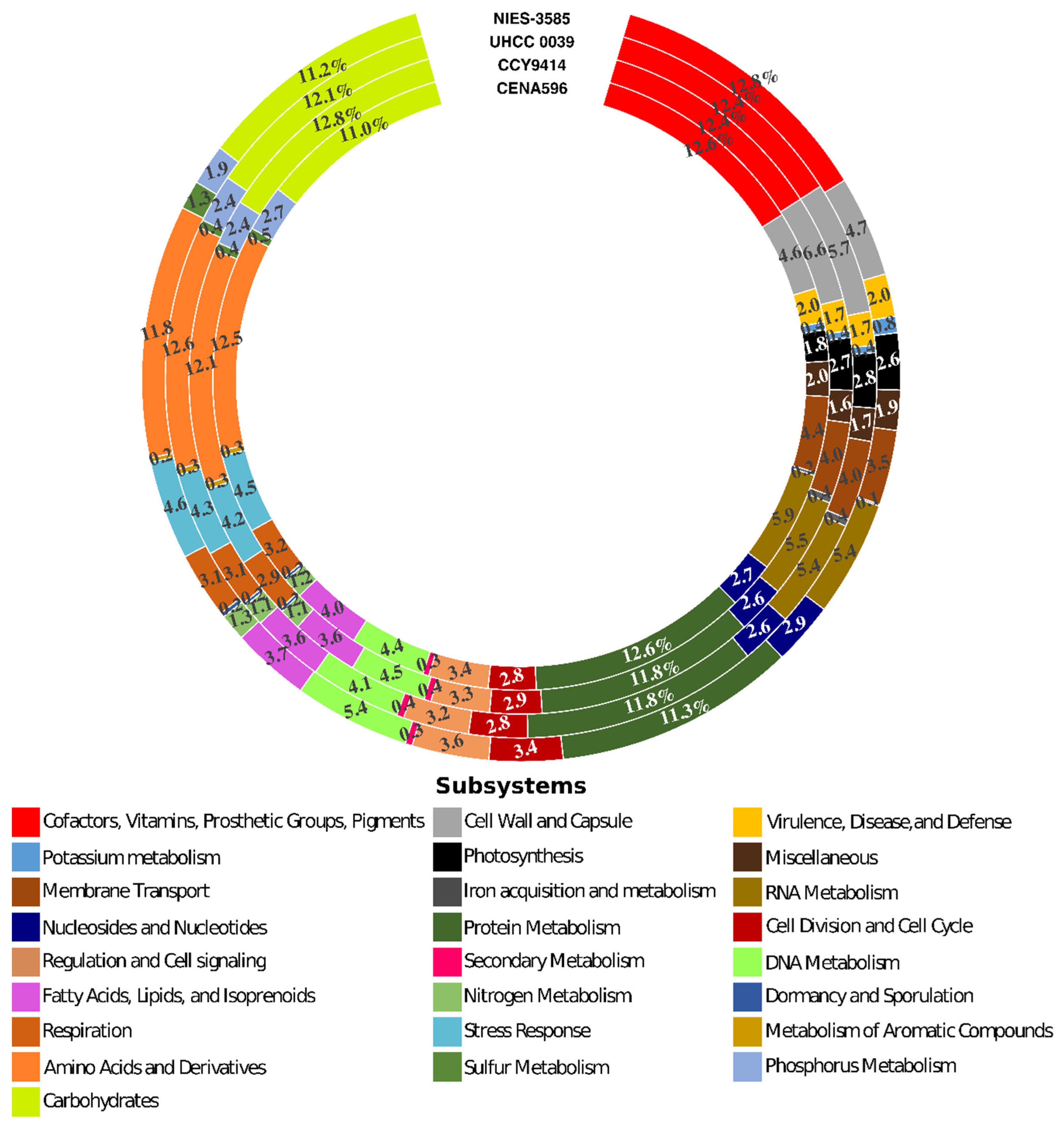
2.3. General Features of Nodularia Genomes
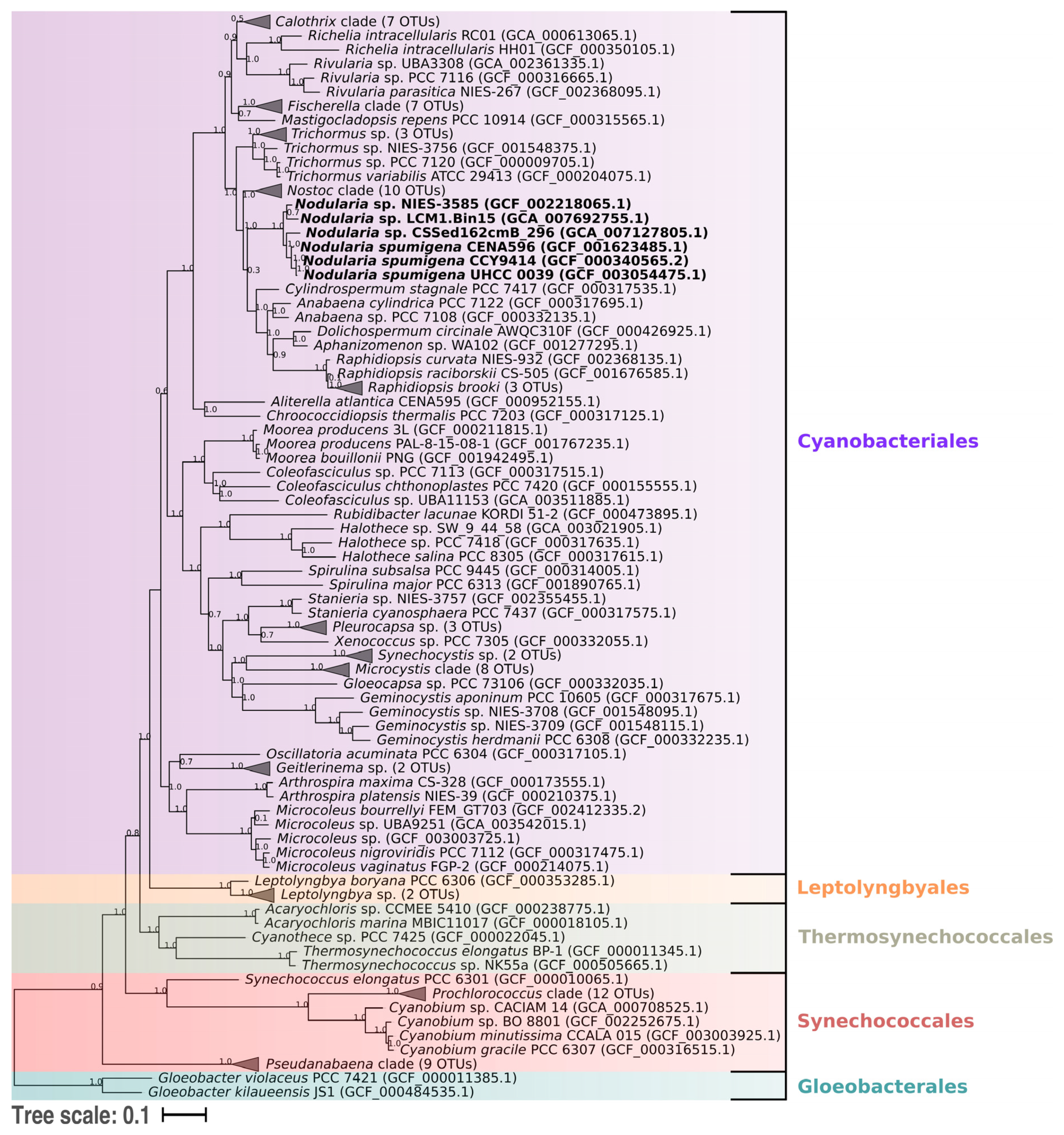
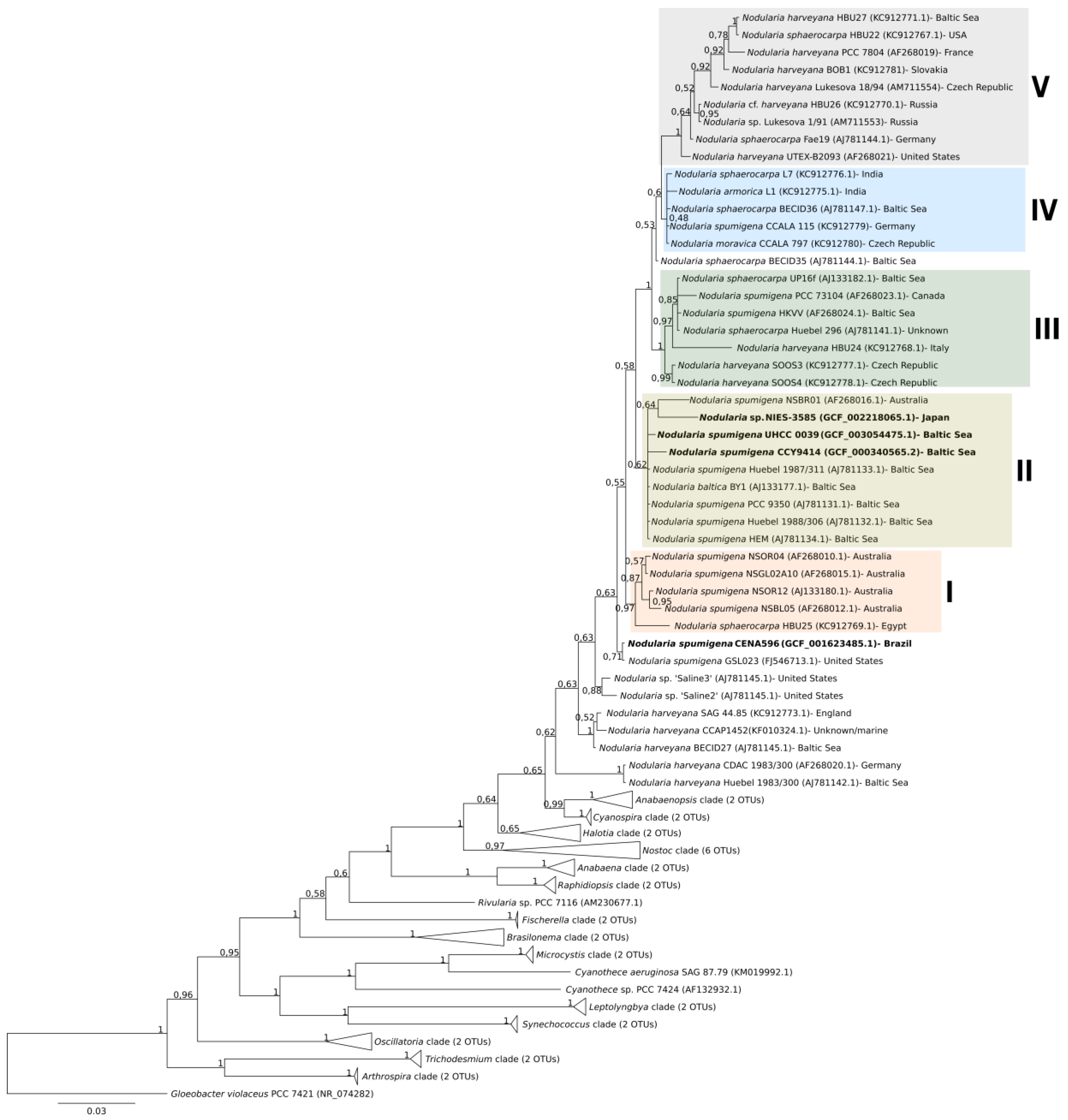
2.4. Phylogenomic Analysis
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Cyanobacterial Genomes
5.2. Natural Product Pathways in N. spumigena CENA596
5.3. Targeted Metabolomics
5.4. Comparative Analyses
5.5. Phylogenomic Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2018—Meeting the Sustainable Development Goals; FAO: Rome, Italy, 2018; ISBN 978-92-5-130562-1. [Google Scholar]
- Ferreira, N.C.; Bonetti, C.; Seiffert, W.Q. Hydrological and Water Quality Indices as management tools in marine shrimp culture. Aquaculture 2011, 318, 425–433. [Google Scholar] [CrossRef]
- Ma, Z.; Song, X.; Wan, R.; Gao, L. A modified water quality index for intensive shrimp ponds of Litopenaeus vannamei. Ecol. Indic. 2013, 24, 287–293. [Google Scholar] [CrossRef]
- Jescovitch, L.N.; Ullman, C.; Rhodes, M.; Davis, D.A. Effects of different feed management treatments on water quality for Pacific white shrimp Litopenaeus vannamei. Aquac. Res. 2018, 49, 526–531. [Google Scholar] [CrossRef]
- Barraza-Guardado, R.H.; Arreola-Lizárraga, J.A.; López-Torres, M.A.; Casillas-Hernández, R.; Miranda-Baeza, A.; Magallón-Barrajas, F.; Ibarra-Gámez, C. Effluents of Shrimp Farms and Its Influence on the Coastal Ecosystems of Bahía de Kino, Mexico. Sci. World J. 2013, 2013, 1–8. [Google Scholar] [CrossRef]
- Alonso-Rodríguez, R.; Páez-Osuna, F. Nutrients, phytoplankton and harmful algal blooms in shrimp ponds: A review with special reference to the situation in the Gulf of California. Aquaculture 2003, 219, 317–336. [Google Scholar]
- Am, A.; Silvester, R.; Alexander, D.; M, N.; Abdulla, M.H. Characterization of blooming algae and bloom-associated changes in the water quality parameters of traditional pokkali cum prawn fields along the South West coast of India. Environ. Monit. Assess. 2016, 188. [Google Scholar] [CrossRef]
- Gao, J.; Zuo, H.; Yang, L.; He, J.-H.; Niu, S.; Weng, S.; He, J.; Xu, X. Long-term influence of cyanobacterial bloom on the immune system of Litopenaeus vannamei. Fish Shellfish Immunol. 2017, 61, 79–85. [Google Scholar] [CrossRef]
- Yang, P.; Lai, D.Y.F.; Jin, B.; Bastviken, D.; Tan, L.; Tong, C. Dynamics of dissolved nutrients in the aquaculture shrimp ponds of the Min River estuary, China: Concentrations, fluxes and environmental loads. Sci. Total Environ. 2017, 603–604, 256–267. [Google Scholar] [CrossRef] [Green Version]
- Main, D.C.; Berry, P.H.; Peet, R.L.; Robertson, J.P. Sheep mortalities associated with the blue green alga Nodularia spumigena. Aust. Vet. J. 1977, 53, 578–581. [Google Scholar] [CrossRef]
- Carmichael, W.W.; Eschedor, J.T.; Patterson, G.M.; Moore, R.E. Toxicity and partial structure of a hepatotoxic peptide produced by the cyanobacterium Nodularia spumigena Mertens emend. L575 from New Zealand. Appl. Environ. Microbiol. 1988, 54, 2257–2263. [Google Scholar] [CrossRef] [Green Version]
- Sivonen, K.; Kononen, K.; Carmichael, W.W.; Dahlem, A.M.; Rinehart, K.L.; Kiviranta, J.; Niemela, S.I. Occurrence of the hepatotoxic cyanobacterium Nodularia spumigena in the Baltic Sea and structure of the toxin. Appl. Environ. Microbiol. 1989, 55, 1990–1995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seaman, M.T.; Ashton, P.J.; Williams, W.D. Inland salt waters of southern Africa. Hydrobiologia 1991, 210, 75–91. [Google Scholar] [CrossRef]
- Nehring, S. Mortality of dogs associated with a mass development of Nodularia spumigena (Cyanophyceae) in a brackish lake at the German North Sea coast. J. Plankton Res. 1993, 15, 867–872. [Google Scholar] [CrossRef]
- Jones, G.; Blackburn, S.; Parker, N. A toxic bloom of Nodularia spumigena Mertens in Orielton Lagoon, Tasmania. Mar. Freshw. Res. 1994, 45, 787–800. [Google Scholar] [CrossRef]
- Harding, W.R.; Rowe, N.; Wessels, J.C.; Beattie, K.A.; Codd, G.A. Death of a dog attributed to the cyanobacterial (blue-green algal) hepatotoxin nodularin in South Africa. J. S. Afr. Vet. Assoc. 1995, 66, 256–259. [Google Scholar]
- Blackburn, S.I.; McCausland, M.A.; Bolch, C.J.S.; Newman, S.J.; Jones, G.J. Effect of salinity on growth and toxin production in cultures of the bloom-forming cyanobacterium Nodularia spumigena from Australian waters. Phycologia 1996, 35, 511–522. [Google Scholar] [CrossRef]
- Mazur-Marzec, H.; Meriluoto, J.; Pliński, M.; Szafranek, J. Characterization of nodularin variants in Nodularia spumigena from the Baltic Sea using liquid chromatography/mass spectrometry/mass spectrometry. Rapid Commun. Mass Spectrom. 2006, 20, 2023–2032. [Google Scholar] [CrossRef]
- Akcaalan, R.; Mazur-Marzec, H.; Zalewska, A.; Albay, M. Phenotypic and toxicological characterization of toxic Nodularia spumigena from a freshwater lake in Turkey. Harmful Algae 2009, 8, 273–278. [Google Scholar] [CrossRef]
- Galat, D.L.; Verdin, J.P.; Sims, L.L. Large-scale patterns of Nodularia spumigena blooms in Pyramid Lake, Nevada, determined from Landsat imagery: 1972–1986. Hydrobiologia 1990, 197, 147–164. [Google Scholar] [CrossRef]
- Beutel, M.W.; Horne, A.J.; Roth, J.C.; Barratt, N.J. Limnological effects of anthropogenic desiccation of a large, saline lake, Walker Lake, Nevada. Hydrobiologia 2001, 466, 91–105. [Google Scholar] [CrossRef]
- Falcón, L.I.; Escobar-Briones, E.; Romero, D. Nitrogen fixation patterns displayed by cyanobacterial consortia in Alchichica crater-lake, Mexico. Hydrobiologia 2002, 467, 71–78. [Google Scholar] [CrossRef]
- Oliva, M.G.; Lugo, J.; Alcocer, L.; Peralta, L.; Oseguera, L. Planktonic bloom-forming Nodularia in the saline lake Alchichica, Mexico. Nat. Resour. Environ. Issues 2009, 15, 121–126. [Google Scholar]
- Bazán, G.I.; Alvarez, S.B. Cianoficeas continentals de la provincia de la pampa (Argentina)-I. Rev. Fac. Agron. Luz. 1994, 7, 43–62. [Google Scholar]
- Pérez Baliero, M.; Del, C.; Bonilla, S.; De León, L.; Šmarda, J.; Komárek, J. A bloom of Nodularia baltica-spumigena group (Cyanobacteria) in a shallow coastal lagoon of Uruguay, South America. Algol. Stud. 1999, 93, 91–101. [Google Scholar]
- Laamanen, M.J.; Gugger, M.F.; Lehtimaki, J.M.; Haukka, K.; Sivonen, K. Diversity of Toxic and Nontoxic Nodularia Isolates (Cyanobacteria) and Filaments from the Baltic Sea. Appl. Environ. Microbiol. 2001, 67, 4638–4647. [Google Scholar] [CrossRef] [Green Version]
- Mazur-Marzec, H.; Kaczkowska, M.; Blaszczyk, A.; Akcaalan, R.; Spoof, L.; Meriluoto, J. Diversity of Peptides Produced by Nodularia spumigena from Various Geographical Regions. Mar. Drugs 2012, 11, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Francis, G. Poisonous Australian Lake. Nature 1878, 18, 11–12. [Google Scholar] [CrossRef] [Green Version]
- Edler, L.; Fernö, S.; Lind, M.G.; Lundberg, R.; Nilsson, P.O. Mortality of dogs associated with a bloom of the cyanobacterium Nodularia spumigena in the Baltic Sea. Ophelia 1985, 24, 103–109. [Google Scholar] [CrossRef]
- Van Halderen, A.; Harding, W.R.; Wessels, J.C.; Schneider, D.J.; Heine, E.W.; Van der Merwe, J.; Fourie, J.M. Cyanobacterial (blue-green algae) poisoning of livestock in the western Cape Province of South Africa. J. S. Afr. Vet. Assoc. 1995, 66, 260–264. [Google Scholar]
- Simola, O.; Wiberg, M.; Jokela, J.; Wahlsten, M.; Sivonen, K.; Syrja, P. Pathologic Findings and Toxin Identification in Cyanobacterial (Nodularia spumigena) Intoxication in a Dog. Vet. Pathol. 2012, 49, 755–759. [Google Scholar] [CrossRef] [Green Version]
- Pacheco, L.; Kunrath, N. Identification of the Toxic Pentapeptide Nodularin in a Cyanobacterial Bloom in a Shrimp Farm in South American Atlantic Coast. Pharm. Anal. Acta 2016, 7, 479. [Google Scholar]
- Da Silveira, S.B.; Wasielesky, W.; Andreote, A.P.D.; Fiore, M.F.; Odebrecht, C. Morphology, phylogeny, growth rate and nodularin production of Nodularia spumigena from Brazil. Mar. Biol. Res. 2017, 13, 1095–1107. [Google Scholar] [CrossRef]
- Rinehart, K.L.; Harada, K.; Namikoshi, M.; Chen, C.; Harvis, C.A.; Munro, M.H.G.; Blunt, J.W.; Mulligan, P.E.; Beasley, V.R.; Dahlem, A.M.; et al. Nodularin, microcystin, and the configuration of Adda. J. Am. Chem. Soc. 1988, 110, 8557–8558. [Google Scholar] [CrossRef]
- Yoshizawa, S.; Matsushima, R.; Watanabe, M.F.; Harada, K.; Ichihara, A.; Carmichael, W.W.; Fujiki, H. Inhibition of protein phosphatases by microcystins and nodularin associated with hepatotoxicity. J. Cancer Res. Clin. Oncol. 1990, 116, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Honkanen, R.E.; Dukelow, M.; Zwiller, J.; Moore, R.E.; Khatra, B.S.; Boynton, A.L. Cyanobacterial nodularin is a potent inhibitor of type 1 and type 2A protein phosphatases. Mol. Pharmacol. 1991, 40, 577–583. [Google Scholar] [PubMed]
- Ohta, T.; Sueoka, E.; Iida, N.; Komori, A.; Suganuma, M.; Nishiwaki, R.; Tatematsu, M.; Kim, S.J.; Carmichael, W.W.; Fujiki, H. Nodularin, a potent inhibitor of protein phosphatases 1 and 2A, is a new environmental carcinogen in male F344 rat liver. Cancer Res. 1994, 54, 6402–6406. [Google Scholar]
- Kankaanpää, H.; Turunen, A.-K.; Karlsson, K.; Bylund, G.; Meriluoto, J.; Sipiä, V. Heterogeneity of nodularin bioaccumulation in northern Baltic Sea flounders in 2002. Chemosphere 2005, 59, 1091–1097. [Google Scholar] [CrossRef]
- Sipiä, V.; Kankaanpää, H.; Peltonen, H.; Vinni, M.; Meriluoto, J. Transfer of nodularin to three-spined stickleback (Gasterosteus aculeatus L.), herring (Clupea harengus L.), and salmon (Salmo salar L.) in the northern Baltic Sea. Ecotoxicol. Environ. Saf. 2007, 66, 421–425. [Google Scholar] [CrossRef]
- Barda, I.; Kankaanpää, H.; Purina, I.; Balode, M.; Sjövall, O.; Meriluoto, J. Bioaccumulation of hepatotoxins—A considerable risk in the Latvian environment. Environ. Pollut. 2015, 196, 313–320. [Google Scholar] [CrossRef]
- Drobac, D.; Tokodi, N.; Lujić, J.; Marinović, Z.; Subakov-Simić, G.; Dulić, T.; Važić, T.; Nybom, S.; Meriluoto, J.; Codd, G.A.; et al. Cyanobacteria and cyanotoxins in fishponds and their effects on fish tissue. Harmful Algae 2016, 55, 66–76. [Google Scholar] [CrossRef]
- Fewer, D.P.; Jokela, J.; Rouhiainen, L.; Wahlsten, M.; Koskenniemi, K.; Stal, L.J.; Sivonen, K. The non-ribosomal assembly and frequent occurrence of the protease inhibitors spumigins in the bloom-forming cyanobacterium Nodularia spumigena. Mol. Microbiol. 2009, 73, 924–937. [Google Scholar] [CrossRef] [PubMed]
- Rouhiainen, L.; Jokela, J.; Fewer, D.P.; Urmann, M.; Sivonen, K. Two Alternative Starter Modules for the Non-Ribosomal Biosynthesis of Specific Anabaenopeptin Variants in Anabaena (Cyanobacteria). Chem. Biol. 2010, 17, 265–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fewer, D.P.; Jokela, J.; Paukku, E.; Österholm, J.; Wahlsten, M.; Permi, P.; Aitio, O.; Rouhiainen, L.; Gomez-Saez, G.V.; Sivonen, K. New Structural Variants of Aeruginosin Produced by the Toxic Bloom Forming Cyanobacterium Nodularia spumigena. PLoS ONE 2013, 8, e73618. [Google Scholar] [CrossRef] [Green Version]
- Suurnäkki, S.; Gomez-Saez, G.V.; Rantala-Ylinen, A.; Jokela, J.; Fewer, D.P.; Sivonen, K. Identification of geosmin and 2-methylisoborneol in cyanobacteria and molecular detection methods for the producers of these compounds. Water Res. 2015, 68, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Voß, B.; Bolhuis, H.; Fewer, D.P.; Kopf, M.; Möke, F.; Haas, F.; El-Shehawy, R.; Hayes, P.; Bergman, B.; Sivonen, K.; et al. Insights into the Physiology and Ecology of the Brackish-Water-Adapted Cyanobacterium Nodularia spumigena CCY9414 Based on a Genome-Transcriptome Analysis. PLoS ONE 2013, 8, e60224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popin, R.V.; Rigonato, J.; Abreu, V.A.C.; Andreote, A.P.D.; Silveira, S.B.; Odebrecht, C.; Fiore, M.F. Draft Genome Assembly of the Bloom-Forming Cyanobacterium Nodularia spumigena Strain CENA596 in Shrimp Production Ponds. Genome Announc. 2016, 4, e00466-16. [Google Scholar] [CrossRef] [Green Version]
- Teikari, J.E.; Fewer, D.P.; Shrestha, R.; Hou, S.; Leikoski, N.; Mäkelä, M.; Simojoki, A.; Hess, W.R.; Sivonen, K. Strains of the toxic and bloom-forming Nodularia spumigena (cyanobacteria) can degrade methylphosphonate and release methane. ISME J. 2018, 12, 1619–1630. [Google Scholar] [CrossRef] [Green Version]
- Teikari, J.E.; Hou, S.; Wahlsten, M.; Hess, W.R.; Sivonen, K. Comparative Genomics of the Baltic Sea Toxic Cyanobacteria Nodularia spumigena UHCC 0039 and Its Response to Varying Salinity. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Mazur-Marzec, H.; Bertos-Fortis, M.; Toruńska-Sitarz, A.; Fidor, A.; Legrand, C. Chemical and Genetic Diversity of Nodularia spumigena from the Baltic Sea. Mar. Drugs 2016, 14, 209. [Google Scholar] [CrossRef] [Green Version]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid Annotations using Subsystems Technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Overbeek, R. The Subsystems Approach to Genome Annotation and its Use in the Project to Annotate 1000 Genomes. Nucleic Acids Res. 2005, 33, 5691–5702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck, C.; Knoop, H.; Axmann, I.M.; Steuer, R. The diversity of cyanobacterial metabolism: Genome analysis of multiple phototrophic microorganisms. BMC Genom. 2012, 13, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crab, R.; Defoirdt, T.; Bossier, P.; Verstraete, W. Biofloc technology in aquaculture: Beneficial effects and future challenges. Aquaculture 2012, 356, 351–356. [Google Scholar] [CrossRef]
- Krummenauer, D.; Peixoto, S.; Cavalli, R.O.; Poersch, L.H.; Wasielesky, W. Superintensive Culture of White Shrimp, Litopenaeus vannamei, in a Biofloc Technology System in Southern Brazil at Different Stocking Densities. J. World Aquac. Soc. 2011, 42, 726–733. [Google Scholar] [CrossRef]
- Sunda, W.G.; Graneli, E.; Gobler, C.J. Positive feedback and the development and persistence of ecosystem disruptive algal blooms. J. Phycol. 2006, 42, 963–974. [Google Scholar] [CrossRef]
- Rosas, C.; López, N.; Mercado, P.; Martínez, E. Effect of Salinity Acclimation on Oxygen Consumption of Juveniles of the White Shrimp Litopenaeus vannamei. J. Crustacean Biol. 2001, 21, 912–922. [Google Scholar] [CrossRef] [Green Version]
- Roy, L.A.; Davis, D.A.; Saoud, I.P.; Boyd, C.A.; Pine, H.J.; Boyd, C.E. Shrimp culture in inland low salinity waters: Shrimp culture in inland low salinity waters. Rev. Aquac. 2010, 2, 191–208. [Google Scholar] [CrossRef]
- Silveira, S.B.; Odebrecht, C. Effects of Salinity and Temperature on the Growth, Toxin Production, and Akinete Germination of the Cyanobacterium Nodularia spumigena. Front. Mar. Sci. 2019, 6, 339. [Google Scholar] [CrossRef]
- Yusoff, F.M.; Zubaidah, M.S.; Matias, H.B.; Kwan, T.S. Phytoplankton succession in intensive marine shrimp culture ponds treated with a commercial bacterial product. Aquac. Res. 2002, 33, 269–278. [Google Scholar] [CrossRef]
- Hayes, P.K.; Barker, G.L.A. Genetic diversity within Baltic sea populations of Nodularia (cyanobacteria). J. Phycol. 1997, 33, 919–923. [Google Scholar] [CrossRef]
- Walsh, C.T.; Chen, H.; Keating, T.A.; Hubbard, B.K.; Losey, H.C.; Luo, L.; Marshall, C.G.; Miller, D.A.; Patel, H.M. Tailoring enzymes that modify nonribosomal peptides during and after chain elongation on NRPS assembly lines. Curr. Opin. Chem. Biol. 2001, 5, 525–534. [Google Scholar] [CrossRef]
- Welker, M.; Von Döhren, H. Cyanobacterial peptides—Nature’s own combinatorial biosynthesis. FEMS Microbiol. Rev. 2006, 30, 530–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, A.C.; Gerwick, L.; Gonzalez, D.; Dorrestein, P.C.; Gerwick, W.H. Transcriptional analysis of the jamaicamide gene cluster from the marine cyanobacterium Lyngbya majuscula and identification of possible regulatory proteins. BMC Microbiol. 2009, 9, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearson, L.A.; Dittmann, E.; Mazmouz, R.; Ongley, S.E.; D’Agostino, P.M.; Neilan, B.A. The genetics, biosynthesis and regulation of toxic specialized metabolites of cyanobacteria. Harmful Algae 2016, 54, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Pearson, L.; Mihali, T.; Moffitt, M.; Kellmann, R.; Neilan, B. On the Chemistry, Toxicology and Genetics of the Cyanobacterial Toxins, Microcystin, Nodularin, Saxitoxin and Cylindrospermopsin. Mar. Drugs 2010, 8, 1650–1680. [Google Scholar] [CrossRef] [Green Version]
- Mazur-Marzec, H.; Tymińska, A.; Szafranek, J.; Pliński, M. Accumulation of nodularin in sediments, mussels, and fish from the Gulf of Gdańsk, southern Baltic Sea. Environ. Toxicol. 2007, 22, 101–111. [Google Scholar] [CrossRef]
- Persson, K.-J.; Legrand, C.; Olsson, T. Detection of nodularin in European flounder (Platichthys flesus) in the west coast of Sweden: Evidence of nodularin mediated oxidative stress. Harmful Algae 2009, 8, 832–838. [Google Scholar] [CrossRef]
- Moffitt, M.C.; Neilan, B.A. Characterization of the Nodularin Synthetase Gene Cluster and Proposed Theory of the Evolution of Cyanobacterial Hepatotoxins. Appl. Environ. Microbiol. 2004, 70, 6353–6362. [Google Scholar] [CrossRef] [Green Version]
- Jonasson, S.; Vintila, S.; Sivonen, K.; El-Shehawy, R. Expression of the nodularin synthetase genes in the Baltic Sea bloom-former cyanobacterium Nodularia spumigena strain AV1: Expression of nda genes in Nodularia spumigena. FEMS Microbiol. Ecol. 2008, 65, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Brutemark, A.; Engström-Öst, J. Does the presence of zooplankton influence growth and toxin production of Nodularia spumigena? Int. Rev. Hydrobiol. 2013, 98, 225–234. [Google Scholar]
- Lyra, C.; Laamanen, M.; Lehtimäki, J.M.; Surakka, A.; Sivonen, K. Benthic cyanobacteria of the genus Nodularia are non-toxic, without gas vacuoles, able to glide and genetically more diverse than planktonic Nodularia. Int. J. Syst. Evol. Microbiol. 2005, 55, 555–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, S.T.; Alvarenga, D.O.; Etchegaray, A.; Fewer, D.P.; Jokela, J.; Varani, A.M.; Sanz, M.; Dörr, F.A.; Pinto, E.; Sivonen, K.; et al. Genetic Organization of Anabaenopeptin and Spumigin Biosynthetic Gene Clusters in the Cyanobacterium Sphaerospermopsis torques-reginae ITEP-024. ACS Chem. Biol. 2017, 12, 769–778. [Google Scholar] [CrossRef]
- Turk, B. Targeting proteases: Successes, failures and future prospects. Nat. Rev. Drug Discov. 2006, 5, 785–799. [Google Scholar] [CrossRef] [PubMed]
- Sanz, M.; Salinas, R.K.; Pinto, E. Namalides B and C and Spumigins K–N from the Cultured Freshwater Cyanobacterium Sphaerospermopsis torques-reginae. J. Nat. Prod. 2017, 80, 2492–2501. [Google Scholar] [CrossRef] [PubMed]
- Shishido, T.K.; Jokela, J.; Fewer, D.P.; Wahlsten, M.; Fiore, M.F.; Sivonen, K. Simultaneous Production of Anabaenopeptins and Namalides by the Cyanobacterium Nostoc sp. CENA543. ACS Chem. Biol. 2017, 12, 2746–2755. [Google Scholar] [CrossRef] [PubMed]
- Cheruku, P.; Plaza, A.; Lauro, G.; Keffer, J.; Lloyd, J.R.; Bifulco, G.; Bewley, C.A. Discovery and Synthesis of Namalide Reveals a New Anabaenopeptin Scaffold and Peptidase Inhibitor. J. Med. Chem. 2012, 55, 735–742. [Google Scholar] [CrossRef] [Green Version]
- Tillier, E.R.M.; Collins, R.A. Genome rearrangement by replication-directed translocation. Nat. Genet. 2000, 26, 195–197. [Google Scholar] [CrossRef]
- Kummerfeld, S.K.; Teichmann, S.A. Relative rates of gene fusion and fission in multi-domain proteins. Trends Genet. 2005, 21, 25–30. [Google Scholar] [CrossRef]
- Ishida, K.; Welker, M.; Christiansen, G.; Cadel-Six, S.; Bouchier, C.; Dittmann, E.; Hertweck, C.; Tandeau de Marsac, N. Plasticity and Evolution of Aeruginosin Biosynthesis in Cyanobacteria. Appl. Environ. Microbiol. 2009, 75, 2017–2026. [Google Scholar] [CrossRef] [Green Version]
- Dittmann, E.; Gugger, M.; Sivonen, K.; Fewer, D.P. Natural Product Biosynthetic Diversity and Comparative Genomics of the Cyanobacteria. Trends Microbiol. 2015, 23, 642–652. [Google Scholar] [CrossRef]
- Murakami, M.; Okita, Y.; Matsuda, H.; Okino, T.; Yamaguchi, K. Aeruginosin 298-A, a thrombin and trypsin inhibitor from the blue-green alga Microcystis aeruginosa (NIES-298). Tetrahedron Lett. 1994, 35, 3129–3132. [Google Scholar] [CrossRef]
- Ersmark, K.; Del Valle, J.R.; Hanessian, S. Chemistry and Biology of the Aeruginosin Family of Serine Protease Inhibitors. Angew. Chem. Int. Ed. 2008, 47, 1202–1223. [Google Scholar] [CrossRef] [PubMed]
- Kohler, E.; Grundler, V.; Häussinger, D.; Kurmayer, R.; Gademann, K.; Pernthaler, J.; Blom, J.F. The toxicity and enzyme activity of a chlorine and sulfate containing aeruginosin isolated from a non-microcystin-producing Planktothrix strain. Harmful Algae 2014, 39, 154–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholz, B.; Liebezeit, G. Biochemical composition, biological activities and toxicological effects of two non-nodularin producing strains of Nodularia spumigena Mertens in Jürgens. J. Appl. Phycol. 2013, 25, 643–660. [Google Scholar] [CrossRef]
- Jüttner, F.; Watson, S.B. Biochemical and Ecological Control of Geosmin and 2-Methylisoborneol in Source Waters. Appl. Environ. Microbiol. 2007, 73, 4395–4406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behr, M.; Serchi, T.; Cocco, E.; Guignard, C.; Sergeant, K.; Renaut, J.; Evers, D. Description of the mechanisms underlying geosmin production in Penicillium expansum using proteomics. J. Proteom. 2014, 96, 13–28. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.P.; Häder, D.-P. UV-protectants in cyanobacteria. Plant. Sci. 2008, 174, 278–289. [Google Scholar] [CrossRef]
- Matsui, K.; Nazifi, E.; Hirai, Y.; Wada, N.; Matsugo, S.; Sakamoto, T. The cyanobacterial UV-absorbing pigment scytonemin displays radical-scavenging activity. J. Gen. Appl. Microbiol. 2012, 58, 137–144. [Google Scholar] [CrossRef] [Green Version]
- D’Agostino, P.M.; Javalkote, V.S.; Mazmouz, R.; Pickford, R.; Puranik, P.R.; Neilan, B.A. Comparative Profiling and Discovery of Novel Glycosylated Mycosporine-Like Amino Acids in Two Strains of the Cyanobacterium Scytonema cf. crispum. Appl. Environ. Microbiol. 2016, 82, 5951–5959. [Google Scholar] [CrossRef] [Green Version]
- Rastogi, R.P.; Sonani, R.R.; Madamwar, D. Cyanobacterial Sunscreen Scytonemin: Role in Photoprotection and Biomedical Research. Appl. Biochem. Biotechnol. 2015, 176, 1551–1563. [Google Scholar] [CrossRef]
- Komárek, J.; Mareš, J. An update to modern taxonomy (2011) of freshwater planktic heterocytous cyanobacteria. Hydrobiologia 2012, 698, 327–351. [Google Scholar] [CrossRef]
- Wang, H.; Sivonen, K.; Rouhiainen, L.; Fewer, D.P.; Lyra, C.; Rantala-Ylinen, A.; Vestola, J.; Jokela, J.; Rantasärkkä, K.; Li, Z.; et al. Genome-derived insights into the biology of the hepatotoxic bloom-forming cyanobacterium Anabaena sp. strain 90. BMC Genom. 2012, 13, 613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes, V.R.; Antunes, A.; Welker, M.; Martins, R.F.; Vasconcelos, V.M. Morphological, toxicological and molecular characterization of a benthic Nodularia isolated from Atlantic estuarine environments. Res. Microbiol. 2010, 161, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Beattie, K.A.; Kaya, K.; Codd, G.A. The cyanobacterium Nodularia PCC 7804, of freshwater origin, produces [L-Har2]nodularin. Phytochemistry 2000, 54, 57–61. [Google Scholar] [CrossRef]
- Moffitt, M.C.; Neilan, B.A. On the presence of peptide synthetase and polyketide synthase genes in the cyanobacterial genus Nodularia. FEMS Microbiol. Lett. 2001, 196, 207–214. [Google Scholar] [CrossRef]
- Tatusov, R.L.; Koonin, E.V.; Lipman, D.J. A genomic perspective on protein families. Science 1997, 278, 631–637. [Google Scholar] [CrossRef] [Green Version]
- Gabaldón, T.; Koonin, E.V. Functional and evolutionary implications of gene orthology. Nat. Rev. Genet. 2013, 14, 360–366. [Google Scholar] [CrossRef] [Green Version]
- Řeháková, K.; Mareš, J.; Lukešová, A.; Zapomělová, E.; Bernardová, K.; Hrouzek, P. Nodularia (Cyanobacteria, Nostocaceae): A phylogenetically uniform genus with variable phenotypes. Phytotaxa 2014, 172, 235. [Google Scholar] [CrossRef] [Green Version]
- Lehtimäki, J.; Lyra, C.; Suomalainen, S.; Sundman, P.; Rouhiainen, L.; Paulin, L.; Salkinoja-Salonen, M.; Sivonen, K. Characterization of Nodularia strains, cyanobacteria from brackish waters, by genotypic and phenotypic methods. Int. J. Syst. Evol. Microbiol. 2000, 50, 1043–1053. [Google Scholar] [CrossRef] [Green Version]
- Bolch, C.J.S.; Orr, P.T.; Jones, G.J.; Blackburn, S.I. Genetic, morphological, and toxicological variation among globally distributed strains of Nodularia (cyanobacteria). J. Phycol. 1999, 35, 339–355. [Google Scholar] [CrossRef]
- Komárek, J. Recent changes (2008) in cyanobacteria taxonomy based on a combination of molecular background with phenotype and ecological consequences (genus and species concept). Hydrobiologia 2010, 639, 245–259. [Google Scholar] [CrossRef]
- Janda, J.M.; Abbott, S.L. 16S rRNA Gene Sequencing for Bacterial Identification in the Diagnostic Laboratory: Pluses, Perils, and Pitfalls. J. Clin. Microbiol. 2007, 45, 2761–2764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz, F.; Eloe-Fadrosh, E.A.; Bowers, R.M.; Jarett, J.; Nielsen, T.; Ivanova, N.N.; Kyrpides, N.C.; Woyke, T. Towards a balanced view of the bacterial tree of life. Microbiome 2017, 5, 140. [Google Scholar] [CrossRef] [PubMed]
- Větrovský, T.; Baldrian, P. The Variability of the 16S rRNA Gene in Bacterial Genomes and Its Consequences for Bacterial Community Analyses. PLoS ONE 2013, 8, e57923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, J.M.; Darling, A.E.; Eisen, J.A. Phylogeny of Bacterial and Archaeal Genomes Using Conserved Genes: Supertrees and Supermatrices. PLoS ONE 2013, 8, e62510. [Google Scholar] [CrossRef] [Green Version]
- Chun, J.; Rainey, F.A. Integrating genomics into the taxonomy and systematics of the Bacteria and Archaea. Int. J. Syst. Evol. Microbiol. 2014, 64, 316–324. [Google Scholar] [CrossRef]
- Shih, P.M.; Wu, D.; Latifi, A.; Axen, S.D.; Fewer, D.P.; Talla, E.; Calteau, A.; Cai, F.; Tandeau de Marsac, N.; Rippka, R.; et al. Improving the coverage of the cyanobacterial phylum using diversity-driven genome sequencing. Proc. Natl. Acad. Sci. USA 2013, 110, 1053–1058. [Google Scholar] [CrossRef] [Green Version]
- Calteau, A.; Fewer, D.P.; Latifi, A.; Coursin, T.; Laurent, T.; Jokela, J.; Kerfeld, C.A.; Sivonen, K.; Piel, J.; Gugger, M. Phylum-wide comparative genomics unravel the diversity of secondary metabolism in Cyanobacteria. BMC Genom. 2014, 15, 977. [Google Scholar] [CrossRef] [Green Version]
- Komárek, J.; Kaštovský, J.; Mareš, J.; Johansen, J. Taxonomic classification of cyanoprokaryotes (cyanobacterial genera) 2014, using a polyphasic approach. Preslia 2014, 86, 295–335. [Google Scholar]
- Parks, D.H.; Chuvochina, M.; Waite, D.W.; Rinke, C.; Skarshewski, A.; Chaumeil, P.-A.; Hugenholtz, P. A standardized bacterial taxonomy based on genome phylogeny substantially revises the tree of life. Nat. Biotechnol. 2018, 36, 996–1004. [Google Scholar] [CrossRef]
- Weber, T.; Blin, K.; Duddela, S.; Krug, D.; Kim, H.U.; Bruccoleri, R.; Lee, S.Y.; Fischbach, M.A.; Müller, R.; Wohlleben, W.; et al. antiSMASH 3.0—A comprehensive resource for the genome mining of biosynthetic gene clusters. Nucleic Acids Res. 2015, 43, W237–W243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutherford, K.; Parkhill, J.; Crook, J.; Horsnell, T.; Rice, P.; Rajandream, M.A.; Barrell, B. Artemis: Sequence visualization and annotation. Bioinformatics 2000, 16, 944–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rottig, M.; Medema, M.H.; Blin, K.; Weber, T.; Rausch, C.; Kohlbacher, O. NRPSpredictor2--a web server for predicting NRPS adenylation domain specificity. Nucleic Acids Res. 2011, 39, W362–W367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillard, R.R.L. Culture of Phytoplankton for Feeding Marine Invertebrates. In Culture of Marine Invertebrate Animals; Smith, W.L., Chanley, M.H., Eds.; Springer: Boston, MA, USA, 1975; pp. 29–60. ISBN 978-1-4615-8716-3. [Google Scholar]
- Bradnam, K.R.; Fass, J.N.; Alexandrov, A.; Baranay, P.; Bechner, M.; Birol, I.; Boisvert, S.; Chapman, J.A.; Chapuis, G.; Chikhi, R.; et al. Assemblathon 2: Evaluating de novo methods of genome assembly in three vertebrate species. GigaScience 2013, 2. [Google Scholar] [CrossRef] [PubMed]
- Petkau, A.; Stuart-Edwards, M.; Stothard, P.; Van Domselaar, G. Interactive microbial genome visualization with GView. Bioinformatics 2010, 26, 3125–3126. [Google Scholar] [CrossRef]
- Wang, Y.; Coleman-Derr, D.; Chen, G.; Gu, Y.Q. OrthoVenn: A web server for genome wide comparison and annotation of orthologous clusters across multiple species. Nucleic Acids Res. 2015, 43, W78–W84. [Google Scholar] [CrossRef]
- Meyer, F.; Paarmann, D.; D’Souza, M.; Olson, R.; Glass, E.; Kubal, M.; Paczian, T.; Rodriguez, A.; Stevens, R.; Wilke, A.; et al. The metagenomics RAST server—A public resource for the automatic phylogenetic and functional analysis of metagenomes. BMC Bioinformatics 2008, 9, 386. [Google Scholar] [CrossRef] [Green Version]
- Contreras-Moreira, B.; Vinuesa, P. GET_HOMOLOGUES, a Versatile Software Package for Scalable and Robust Microbial Pangenome Analysis. Appl. Environ. Microbiol. 2013, 79, 7696–7701. [Google Scholar] [CrossRef] [Green Version]
- Vinuesa, P.; Contreras-Moreira, B. Robust Identification of Orthologues and Paralogues for Microbial Pan-Genomics Using GET_HOMOLOGUES: A Case Study of pIncA/C Plasmids. In Bacterial Pangenomics; Mengoni, A., Galardini, M., Fondi, M., Eds.; Springer New York: New York, NY, USA, 2015; Volume 1231, pp. 203–232. ISBN 978-1-4939-1719-8. [Google Scholar]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Posada, D. jModelTest: Phylogenetic Model Averaging. Mol. Biol. Evolut. 2008, 25, 1253–1256. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Natural Product | Structural Subunits | Strain | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Nodularin | MW | 1 | 2 | 3 | 4 | 5 | 6 | CENA596 | CCY9414 | UHCC0039 |
| NOD | 824 | MeAsp | Arg | Adda | Glu | MeDhb | * | + | + | + |
| [D-Asp1] NOD | 810 | Asp | Arg | Adda | Glu | MeDhb | * | + | + | + |
| Spumigin | MW | 1 | 2 | 3 | 4 | 5 | 6 | CENA596 | CCY9414 | UHCC0039 |
| A | 612 | Hpla | Hty | mPro | Argol | * | * | - | + | + |
| B1 | 626 | Hpla | Hty | mPro | Arg | * | * | - | + | + |
| B2 | 626 | Hpla | Hty | mPro | Arg | * | * | - | + | + |
| D | 598 | Hpla | Hty | Pro | Argol | * | * | + | + | + |
| E | 610 | Hpla | Hty | mPro | Argal | * | * | - | + | + |
| F | 596 | Hpla | Hty | Pro | Argal | * | * | + | + | + |
| G | 596 | Hpla | Hph | mPro | Argal | * | * | - | + | + |
| H | 580 | Hpla | Hph | Pro | Argal | * | * | - | + | + |
| 582b | 582 | Hpla | Hty | mPro | Agm | * | * | - | - | + |
| Anabaenopeptin | MW | 1 | 2 | 3 | 4 | 5 | 6 | CENA596 | CCY9414 | UHCC0039 |
| NP933 | 933 | Phe | Lys | Val | Hty | MeHty | MetO | - | + | + |
| NPA | 929 | Ile | Lys | MetO2 | Hph | MeHty | AcSer | - | + | + |
| NP915a | 915 | Ile | Lys | MetO | Hph | MeHty | Met | - | + | + |
| NPB | 913 | Ile | Lys | MetO | Hph | MeHty | AcSer | - | + | + |
| NP 839 | 839 | Ile | Lys | Met | Hph | MeHph | Ser | - | + | + |
| NP 849 | 849 | Ile | Lys | Val | Hph | MeHph | AcSer | - | - | + |
| NP 855a | 855 | Ile | Lys | Met | Hph | MeHty | Ser | - | + | + |
| NP 855b | 855 | Ile | Lys | MetO | Hph | MeHph | Ser | - | + | - |
| NP 881a | 881 | Ile | Lys | Met | Hph | MeHph | AcSer | - | + | + |
| NP 881b | 881 | Ile | Lys | Ile | Hph | MeHty | Met | - | + | + |
| NP 879 | 879 | Ile | Lys | Ile | Hph | MeHty | AcSer | - | + | + |
| [Ser6] NPB | 871 | Ile | Lys | MetO | Hph | MeHty | Ser | - | + | + |
| NP 867 | 867 | Ile | Lys | Val | Hph | MeHty | Met | - | - | + |
| NP 865 | 865 | Ile | Lys | Val | Hph | MeHty | AcSer | - | - | + |
| NP 863 | 863 | Ile | Lys | Ile | Hph | MeHph | Ser | - | - | + |
| NP 883a | 883 | Ile | Lys | Met | Hph | MeHph | Met | - | + | + |
| [Met6] NPC | 899 | Ile | Lys | Met | Hph | MeHty | Met | - | + | + |
| NPC | 897 | Ile | Lys | Met | Hph | MeHty | AcSer | - | + | + |
| [MeHph5] NPB | 897 | Ile | Lys | MetO | Hph | MeHph | AcSer | - | + | + |
| Namalide | MW | 1 | 2 | 3 | 4 | 5 | 6 | CENA596 | CCY9414 | UHCC0039 |
| B | 575 | Ile | Lys | Ile/Leu | Hty | * | * | + | – | – |
| C | 561 | Ile | Lys | Val | Hty | * | * | + | – | – |
| Aeruginosin | MW | 1 | 2 | 3 | 4 | 5 | 6 | CENA596 | CCY9414 | UHCC0039 |
| NAL1 | 558 | Bu | Tyr | Choi | Argal | * | * | - | + | + |
| NAL2 | 586 | Hex | Tyr | Choi | Argal | * | * | + | + | + |
| NAL3 | 614 | Oct | Tyr | Choi | Argal | * | * | + | + | + |
| NOL1 | 532 | Ac | Tyr | Choi | Argol | * | * | - | + | + |
| NOL2 | 560 | Bu | Tyr | Choi | Argol | * | * | - | + | + |
| NOL3 | 588 | Hex | Tyr | Choi | Argol | * | * | + | + | + |
| NOL4 | 616 | Oct | Tyr | Choi | Argol | * | * | - | + | + |
| Terpenes | MW | 1 | 2 | 3 | 4 | 5 | 6 | CENA596 | CCY9414 | UHCC0039 |
| Geosmin | * | * | * | * | * | * | * | + | - | - |
| Mycosporine-like amino acids | MW | 1 | 2 | 3 | 4 | 5 | 6 | CENA596 | CCY9414 | UHCC0039 |
| Shinorine | 332 | Maa | Gly | Ser | * | * | * | + | - | - |
| Porphyra 334 | 346 | Maa | Gly | Thr | * | * | * | + | - | - |
| Genome Statistics | Nodularia spumigena | Nodularia sp. | ||
|---|---|---|---|---|
| CENA596 | CCY9414 | UHCC 0039 | NIES-3585 | |
| Number of contigs | 291 | 76 | 2 | 20 |
| Number of scaffolds | - | 1 | - | 4 |
| Number of plasmids | - | - | 1 | - |
| Total size (bp) | 5,189,679 | 5,462,271 | 5,386,612 | 5,773,538 |
| Max scaffolds length (bp) | 109,819 | 5,462,271 | 5,294,286 | 5,482,519 |
| Min scaffolds length (bp) | 526 | 5,462,271 | 92,326 | 58,866 |
| Mean scaffold size (bp) | 17,834 | 5,462,271 | 2,693,306 | 1,443,384 |
| Median scaffold size (bp) | 12,498 | 5,462,271 | 2,693,306 | 156,947 |
| GC content (%) | 41.2 | 41.19 | 41.2 | 41.2 |
| N50 (bp) | 32,474 | 5,462,271 | 5,294,286 | 1,022,420 |
| Subsystem annotation statistics | - | - | - | - |
| Number of subsystems | 370 | 378 | 383 | 387 |
| Number of coding sequences | 4907 | 5277 | 5207 | 5546 |
| Coding sequences in subsystems | 1595 (33%) | 1704 (33%) | 1727 (34%) | 1700 (31%) |
| Coding sequences not in subsystems | 3312 (67%) | 3573 (67%) | 3480 (66%) | 3846 (69%) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popin, R.V.; Delbaje, E.; Abreu, V.A.C.d.; Rigonato, J.; Dörr, F.A.; Pinto, E.; Sivonen, K.; Fiore, M.F. Genomic and Metabolomic Analyses of Natural Products in Nodularia spumigena Isolated from a Shrimp Culture Pond. Toxins 2020, 12, 141. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12030141
Popin RV, Delbaje E, Abreu VACd, Rigonato J, Dörr FA, Pinto E, Sivonen K, Fiore MF. Genomic and Metabolomic Analyses of Natural Products in Nodularia spumigena Isolated from a Shrimp Culture Pond. Toxins. 2020; 12(3):141. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12030141
Chicago/Turabian StylePopin, Rafael Vicentini, Endrews Delbaje, Vinicius Augusto Carvalho de Abreu, Janaina Rigonato, Felipe Augusto Dörr, Ernani Pinto, Kaarina Sivonen, and Marli Fatima Fiore. 2020. "Genomic and Metabolomic Analyses of Natural Products in Nodularia spumigena Isolated from a Shrimp Culture Pond" Toxins 12, no. 3: 141. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12030141