Independent and Synergistic Effects of Knocking out Two ABC Transporter Genes on Resistance to Bacillus thuringiensis Toxins Cry1Ac and Cry1Fa in Diamondback Moth


Abstract
:1. Introduction
2. Results
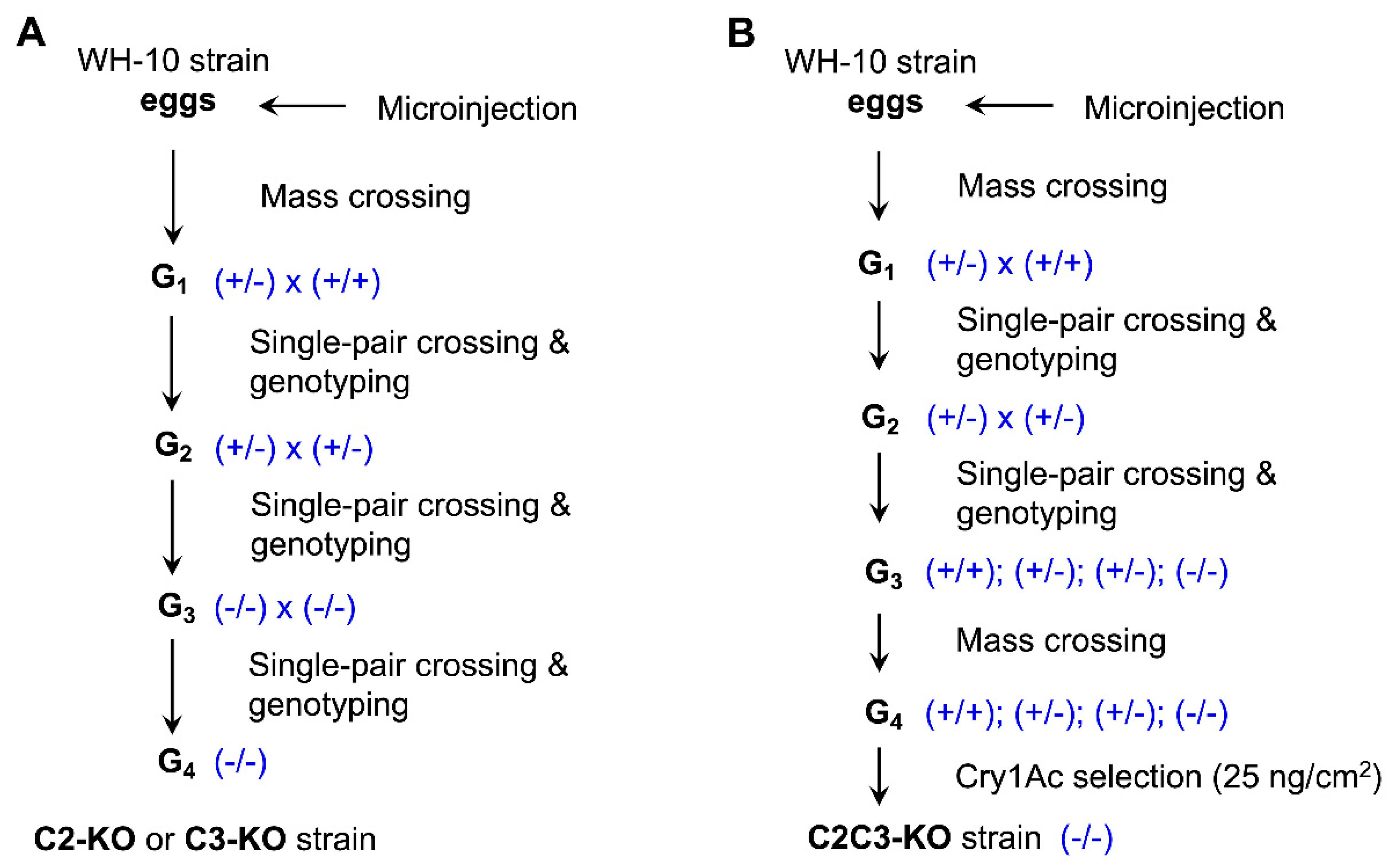
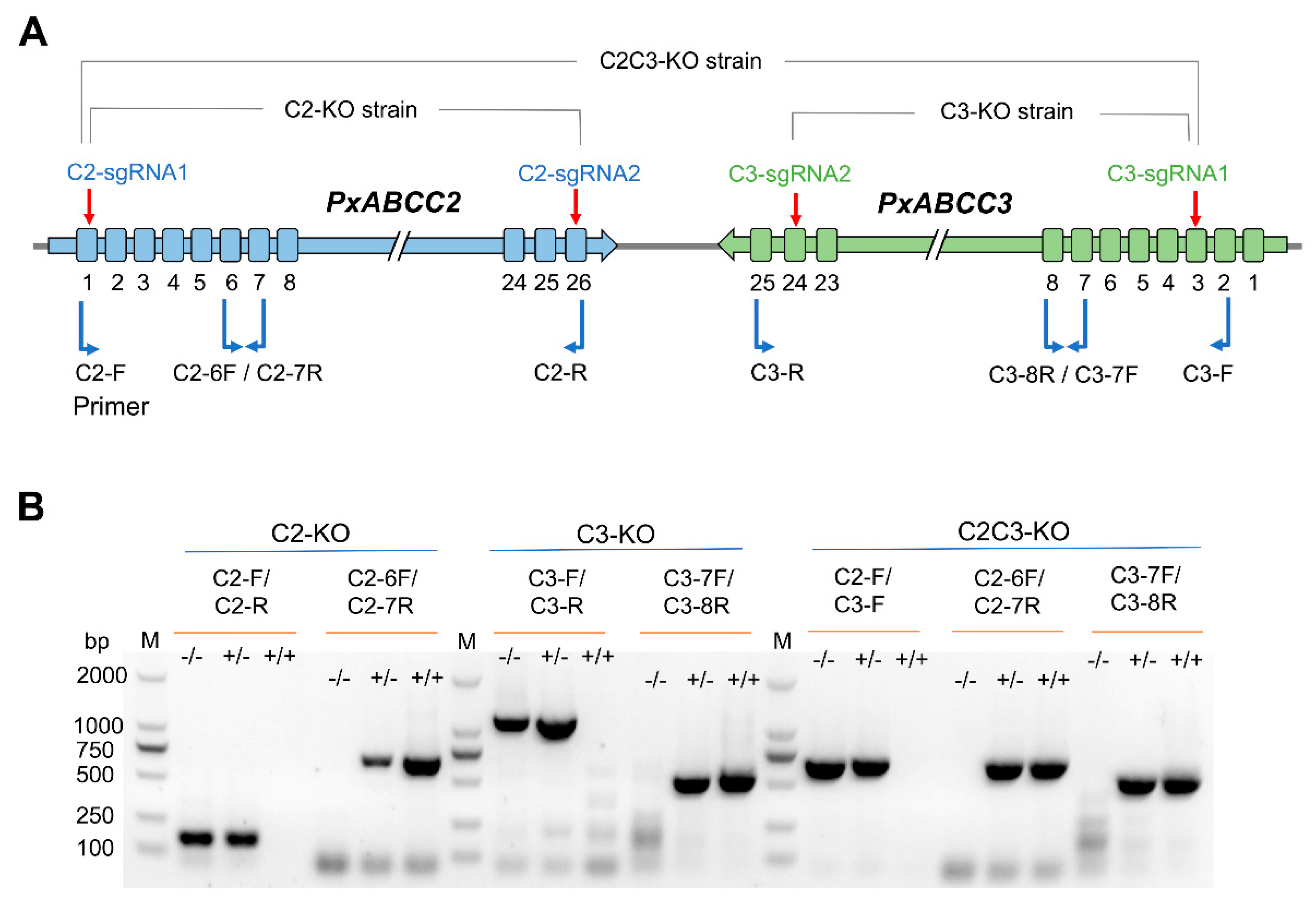
2.1. CRISPR-Mediated Single Knockouts of PxABCC2 or PxABCC3
2.2. CRISPR-Mediated Double Knockout of PxABCC2 and PxABCC3
2.3. Responses of Knockout Strains to Bt Toxins Cry1Ac and Cry1Fa
2.4. Recessive Resistance to Cry1Ac in the Double Knockout Strain C2C3-KO
3. Discussion
4. Materials and Methods
4.1. Insect Strains
4.2. Bt Toxins and Bioassays
4.3. Cas9 Protein and sgRNAs
4.4. Embryo Microinjection
4.5. Genomic DNA Extraction and Mutation Identification
4.6. Inheritance of Cry1Ac Resistance in the Double Knockout Strain
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sanahuja, G.; Banakar, R.; Twyman, R.M.; Capell, T.; Christou, P. Bacillus thuringiensis: A century of research, development and commercial applications. Plant Biotechnol. J. 2011, 9, 283–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ISAAA. Global Status of Commercialized Biotech/GM Crops in 2018; ISAAA Brief no. 54; ISAAA: Ithaca, NY, USA, 2018. [Google Scholar]
- Furlong, M.J.; Wright, D.J.; Dosdall, L.M. Diamondback moth ecology and management: Problems, progress, and prospects. Annu. Rev. Entomol. 2013, 58, 517–541. [Google Scholar] [CrossRef] [PubMed]
- Janmaat, A.; Franklin, M.; Myers, J.H. Resistance of cabbage loopers to Btk in a greenhouse setting: Occurrence, spread and management. In Bt Resistance: Characterization and Strategies for GM Crops Producing Bacillus thuringiensis Toxins; Soberón, M., Gao, Y., Bravo, A., Eds.; CABI: London, UK, 2015; pp. 49–55. [Google Scholar]
- Tabashnik, B.E.; Carrière, Y. Global patterns of resistance to Bt crops highlighting pink bollworm in the United States, China, and India. J. Econ. Entomol. 2019, 112, 2513–2523. [Google Scholar] [CrossRef] [PubMed]
- Calles-Torrez, V.; Knodel, J.J.; Boetel, M.A.; French, B.W.; Fuller, B.W.; Ransom, J.K. Field-evolved resistance of northern and western corn rootworm (Coleoptera: Chrysomelidae) populations to corn hybrids expressing single and pyramided Cry3Bb1 and Cry34/35Ab1 Bt proteins in North Dakota. J. Econ. Entomol. 2019, 112, 1875–1886. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.L.; Farhan, Y.; Schaafsma, A.W. Practical resistance of Ostrinia nubilalis (Lepidoptera: Crambidae) to Cry1F Bacillus thuringiensis maize discovered in Nova Scotia, Canada. Sci. Rep. 2019, 9, 18247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talekar, N.S.; Shelton, A.M. Biology, ecology, and management of the diamondback moth. Annu. Rev. Entomol. 1993, 38, 275–301. [Google Scholar] [CrossRef]
- Zalucki, M.P.; Shabbir, A.; Silva, R.; Adamson, D.; Liu, S.S.; Furlong, M.J. Estimating the economic cost of one of the world’s major insect pests, Plutella xylostella (Lepidoptera: Plutellidae): Just how long is a piece of string? J. Econ. Entomol. 2012, 105, 1115–1129. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Cushing, N.L.; Finson, N.; Johnson, M.W. Field development of resistance to Bacillus thuringiensis in diamondback moth (Lepidoptera, Plutellidae). J. Econ. Entomol. 1990, 83, 1671–1676. [Google Scholar] [CrossRef]
- Heckel, D.G. Learning the ABCs of Bt: ABC transporters and insect resistance to Bacillus thuringiensis provide clues to a crucial step in toxin mode of action. Pestic. Biochem. Phys. 2012, 104, 103–110. [Google Scholar] [CrossRef]
- Heckel, D.G. Roles of ABC proteins in the mechanism and management of Bt resistance. In Bt Resistance: Characterization and Strategies for GM Crops Producing Bacillus thuringiensis Toxins; Soberón, M., Gao, Y., Bravo, A., Eds.; CABI: London, UK, 2015; pp. 107–118. [Google Scholar]
- Tabashnik, B.E. ABCs of insect resistance to Bt. PLoS Genet. 2015, 11, e1005646. [Google Scholar] [CrossRef] [Green Version]
- Mitsuhashi, W.; Miyamoto, K. Interaction of Bacillus thuringiensis Cry toxins and the insect midgut with a focus on the silkworm (Bombyx mori) midgut. Biocontrol Sci. Tech. 2020, 30, 68–84. [Google Scholar] [CrossRef]
- Guo, Z.J.; Sun, D.; Kang, S.; Zhou, J.L.; Gong, L.J.; Qin, J.Y.; Guo, L.; Zhu, L.H.; Bai, Y.; Luo, L.; et al. CRISPR/Cas9-mediated knockout of both the PxABCC2 and PxABCC3 genes confers high-level resistance to Bacillus thuringiensis Cry1Ac toxin in the diamondback moth, Plutella xylostella (L.). Insect Biochem. Mol. Biol. 2019, 107, 31–38. [Google Scholar] [CrossRef]
- Liu, Z.X.; Fu, S.; Ma, X.L.; Baxter, S.W.; Vasseur, L.; Xiong, L.; Huang, Y.P.; Yang, G.; You, S.J.; You, M.S. Resistance to Bacillus thuringiensis Cry1Ac toxin requires mutations in two Plutella xylostella ATP-binding cassette transporter paralogs. PLoS Path. 2020, 16, e1008697. [Google Scholar] [CrossRef] [PubMed]
- Baxter, S.W.; Badenes-Perez, F.R.; Morrison, A.; Vogel, H.; Crickmore, N.; Kain, W.; Wang, P.; Heckel, D.G.; Jiggins, C.D. Parallel evolution of Bacillus thuringiensis toxin resistance in Lepidoptera. Genetics 2011, 189, 675–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ocelotl, J.; Sánchez, J.; Gómez, I.; Tabashnik, B.E.; Bravo, A.; Soberón, M. ABCC2 is associated with Bacillus thuringiensis Cry1Ac toxin oligomerization and membrane insertion in diamondback moth. Sci. Rep. 2017, 7, 2386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Cabrera, J.; Herrero, S.; Sayyed, A.H.; Escriche, B.; Liu, Y.B.; Meyer, S.K.; Wright, D.J.; Tabashnik, B.E.; Ferré, J. Variation in susceptibility to Bacillus thuringiensis toxins among unselected strains of Plutella xylostella. Appl. Environ. Microb. 2001, 67, 4610–4613. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.; Kang, S.; Chen, D.; Wu, Q.; Wang, S.; Xie, W.; Zhu, X.; Baxter, S.W.; Zhou, X.; Jurat-Fuentes, J.L.; et al. MAPK signaling pathway alters expression of midgut ALP and ABCC genes and causes resistance to Bacillus thuringiensis Cry1Ac toxin in diamondback moth. PLoS Genet. 2015, 11, e1005124. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.L.; Xu, Y.J.; Zuo, Y.Y.; Yang, Y.H.; Tabashnik, B.E.; Wu, Y.D. Evaluation of five candidate receptors for three Bt toxins in the beet armyworm using CRISPR-mediated gene knockouts. Insect Biochem. Mol. Biol. 2020, 121, 103361. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Finson, N.; Johnson, K.W.; Heckel, D.G. Cross-resistance to Bacillus thuringiensis toxin Cry1F in the diamondback moth (Plutella xylostella). Appl. Environ. Microbiol. 1994, 60, 4627–4629. [Google Scholar] [CrossRef] [Green Version]
- Tabashnik, B.E.; Johnson, K.W.; Engleman, J.T.; Baum, J.A. Cross-resistance to Bacillus thuringiensis toxin Cry1Ja in a strain of diamondback moth adapted to artificial diet. J. Inveterbr. Pathol. 2000, 76, 81–83. [Google Scholar] [CrossRef]
- Wang, J.; Ma, H.H.; Zhao, S.; Huang, J.L.; Yang, Y.H.; Tabashnik, B.E.; Wu, Y.D. Functional redundancy of two ABC transporter proteins in mediating toxicity of Bacillus thuringiensis to cotton bollworm. PLoS Path. 2020, 16, e1008427. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.L.; Xu, Y.J.; Huang, J.L.; Jin, W.Z.; Yang, Y.H.; Wu, Y.D. CRISPR-mediated knockout of the ABCC2 gene in Ostrinia furnacalis confers high-level resistance to the Bacillus thuringiensis Cry1Fa toxin. Toxins 2020, 12, 246. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.H.; Tao, J.H.; Li, Q.; Cheng, Y.; Sun, X.X.; Wu, K.M.; Xiao, Y.T. Genome editing of the SfABCC2 gene confers resistance to Cry1F toxin from Bacillus thuringiensis in Spodoptera frugiperda. J. Integr. Agric. 2020. [Google Scholar] [CrossRef]
- Wang, S.H.; Kain, W.; Wang, P. Bacillus thuringiensis Cry1A toxins exert toxicity by multiple pathways in insects. Insect Biochem. Mol. Biol. 2018, 102, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, R.; Hasler, J.; Meagher, R.; Nagoshi, R.; Hietala, L.; Huang, F.N.; Narva, K.; Jurat-Fuentes, J.L. Mechanism and DNA-based detection of field-evolved resistance to transgenic Bt corn in fall armyworm (Spodoptera frugiperda). Sci. Rep. 2017, 7, 10877. [Google Scholar] [CrossRef] [Green Version]
- Jin, L.; Wang, J.; Guan, F.; Zhang, J.P.; Yu, S.; Liu, S.Y.; Xue, Y.Y.; Li, L.L.; Wu, S.W.; Wang, X.L.; et al. Dominant point mutation in a tetraspanin gene associated with field-evolved resistance of cotton bollworm to transgenic Bt cotton. Proc. Natl. Acad. Sci. USA 2018, 115, 11760–11765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathew, L.G.; Ponnuraj, J.; Mallappa, B.; Chowdary, L.R.; Zhang, J.; Tay, W.T.; Walsh, T.K.; Gordon, K.H.J.; Heckel, D.G.; Downes, S.; et al. ABC transporter mis-splicing associated with resistance to Bt toxin Cry2Ab in laboratory- and field-selected pink bollworm. Sci. Rep. 2018, 8, 13531. [Google Scholar] [CrossRef]
- Boaventura, D.; Ulrich, J.; Lueke, B.; Bolzan, A.; Okuma, D.; Gutbrod, O.; Geibel, S.; Zeng, Q.; Dourado, P.M.; Martinelli, S.; et al. Molecular characterization of Cry1F resistance in fall armyworm, Spodoptera frugiperda from Brazil. Insect Biochem. Mol. Biol. 2019, 116, 103280. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, H.N.; Wang, H.D.; Zhao, S.; Zuo, Y.Y.; Yang, Y.H.; Wu, Y.D. Functional validation of cadherin as a receptor of Bt toxin Cry1Ac in Helicoverpa armigera utilizing the CRISPR/Cas9 system. Insect Biochem. Mol. Biol. 2016, 76, 11–17. [Google Scholar] [CrossRef]
- Wang, J.; Wang, H.D.; Liu, S.Y.; Liu, L.P.; Tay, W.T.; Walsh, T.K.; Yang, Y.H.; Wu, Y.D. CRISPR/Cas9 mediated genome editing of Helicoverpa armigera with mutations of an ABC transporter gene HaABCA2 confers resistance to Bacillus thuringiensis Cry2A toxins. Insect Biochem. Mol. Biol. 2017, 87, 147–153. [Google Scholar] [CrossRef]
- Liu, Y.B.; Tabashnik, B.E. Inheritance of resistance to the Bacillus thuringiensis toxin Cry1C in the diamondback moth. Appl. Environ. Microbiol. 1997, 63, 2218–2223. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Toxin | Strain | Slope ± SE | LC50 (95% FL 1) (ng/cm2) | RR 2 |
|---|---|---|---|---|
| Cry1Ac | WH-10 | 2.92 ± 0.38 | 3.10 (2.57–3.67) | |
| C2-KO | 1.97 ± 0.30 | 8.92 (6.74–13.40) | 2.9 | |
| C3-KO | 3.19 ± 0.55 | 6.59 (4.88–8.25) | 2.1 | |
| C2C3-KO | >32,000 3 | >10,320 | ||
| Cry1Fa | WH-10 | 8.72 ± 1.17 | 10.48 (9.77–11.14) | |
| C2-KO | 5.99 ± 0.96 | 10.60 (9.05–11.97) | 1.0 | |
| C3-KO | 7.20 ± 1.09 | 7.47 (6.89–8.10) | 0.7 | |
| C2C3-KO | 3.91 ± 0.59 | 3971 (3314–4821) | 380 |
| Strain/Cross | N 1 | Survival (%) | Dominance Value (h) 2 |
|---|---|---|---|
| WH-10 | 48 | 0 | |
| C2C3-KO | 48 | 100 | |
| C2C3-KO♀×WH-10♂ (F1a) | 96 | 3 | 0.03 |
| C2C3-KO♂×WH-10♀ (F1b) | 96 | 0 | 0 |
| Name | Primer Sequences (5′ > 3′) | PCR Cycling Condition | Product Size (bp) |
|---|---|---|---|
| C2-F C2-R | GAGCCCGGAAAGAGTCGGAAGA TGTTGTCTCCGGTCTCCTC | 95 °C 30 s 59.5 °C 30 s 72 °C 30 s | ~170 |
| C2-6F C2-7R | GGGAGATCCCCTTCCAGAAG AACTCCTGAAGTCTTTCCAATGAG | 95 °C 30 s 57 °C 30 s 72 °C 60 s | ~660 |
| C3-F C3-R | GAGCCGTCGTACCCCAAGGTGTTAT TGCTTTGAAAATACGCTTCCT | 95 °C 30 s 57.5 °C 30 s 72 °C 90 s | ~1130 |
| C3-7F C3-8R | GAACATCACGCTGATCCTGC GACTGATAGGACAAGGGCCG | 95 °C 30 s 58.5 °C 30 s 72 °C 40 s | ~530 |
| C2-F C3-F | GAGCCCGGAAAGAGTCGGAAGA GAGCCGTCGTACCCCAAGGTGTTAT | 95 °C 30 s 62 °C 30 s 72 °C 60 s | ~670 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, S.; Jiang, D.; Wang, F.; Yang, Y.; Tabashnik, B.E.; Wu, Y. Independent and Synergistic Effects of Knocking out Two ABC Transporter Genes on Resistance to Bacillus thuringiensis Toxins Cry1Ac and Cry1Fa in Diamondback Moth. Toxins 2021, 13, 9. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13010009
Zhao S, Jiang D, Wang F, Yang Y, Tabashnik BE, Wu Y. Independent and Synergistic Effects of Knocking out Two ABC Transporter Genes on Resistance to Bacillus thuringiensis Toxins Cry1Ac and Cry1Fa in Diamondback Moth. Toxins. 2021; 13(1):9. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13010009
Chicago/Turabian StyleZhao, Shan, Dong Jiang, Falong Wang, Yihua Yang, Bruce E. Tabashnik, and Yidong Wu. 2021. "Independent and Synergistic Effects of Knocking out Two ABC Transporter Genes on Resistance to Bacillus thuringiensis Toxins Cry1Ac and Cry1Fa in Diamondback Moth" Toxins 13, no. 1: 9. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13010009