Molecular Docking and Comparative Inhibitory Efficacy of Naturally Occurring Compounds on Vegetative Growth and Deoxynivalenol Biosynthesis in Fusarium culmorum

 ,
,  ,
,  , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
2. Results
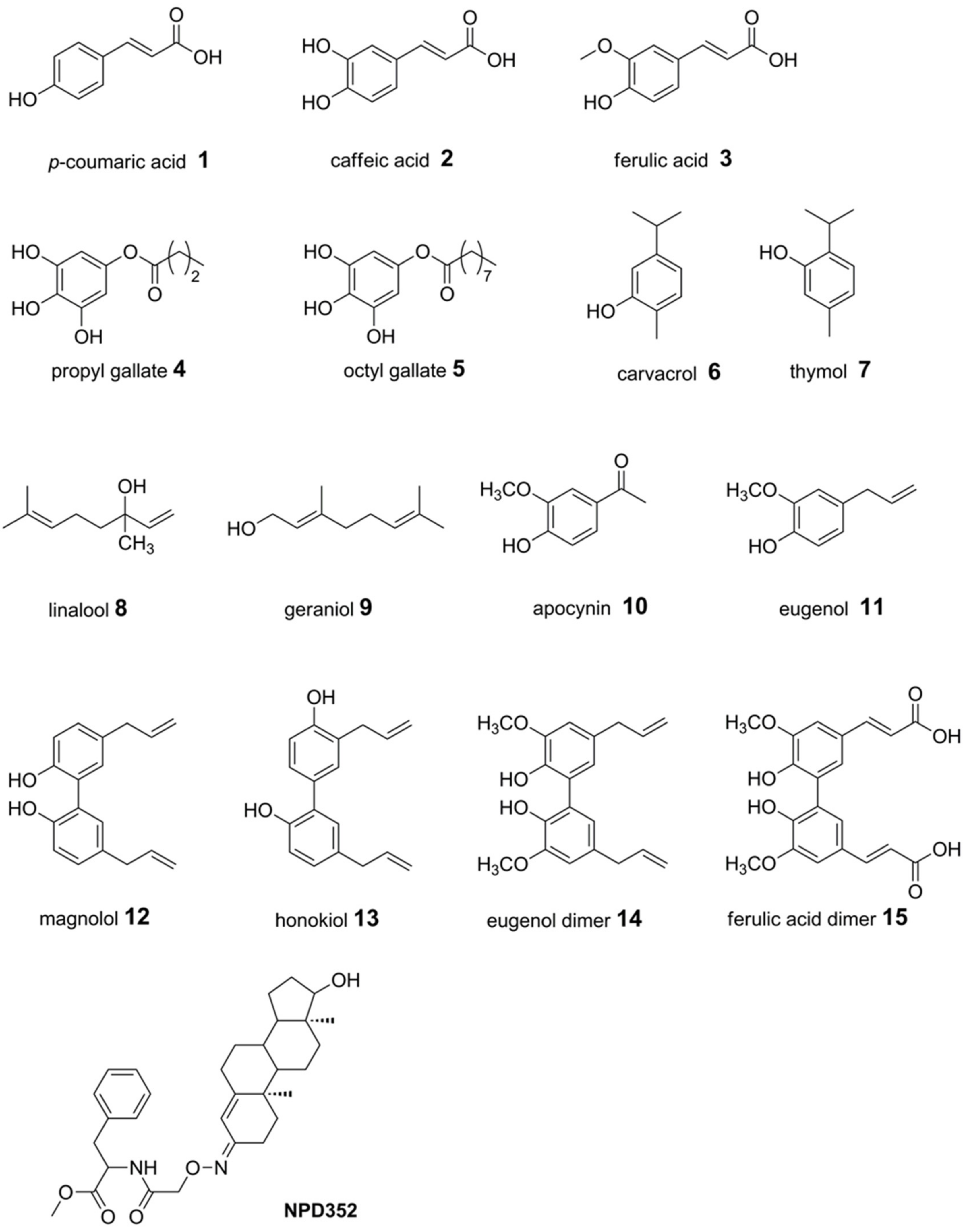
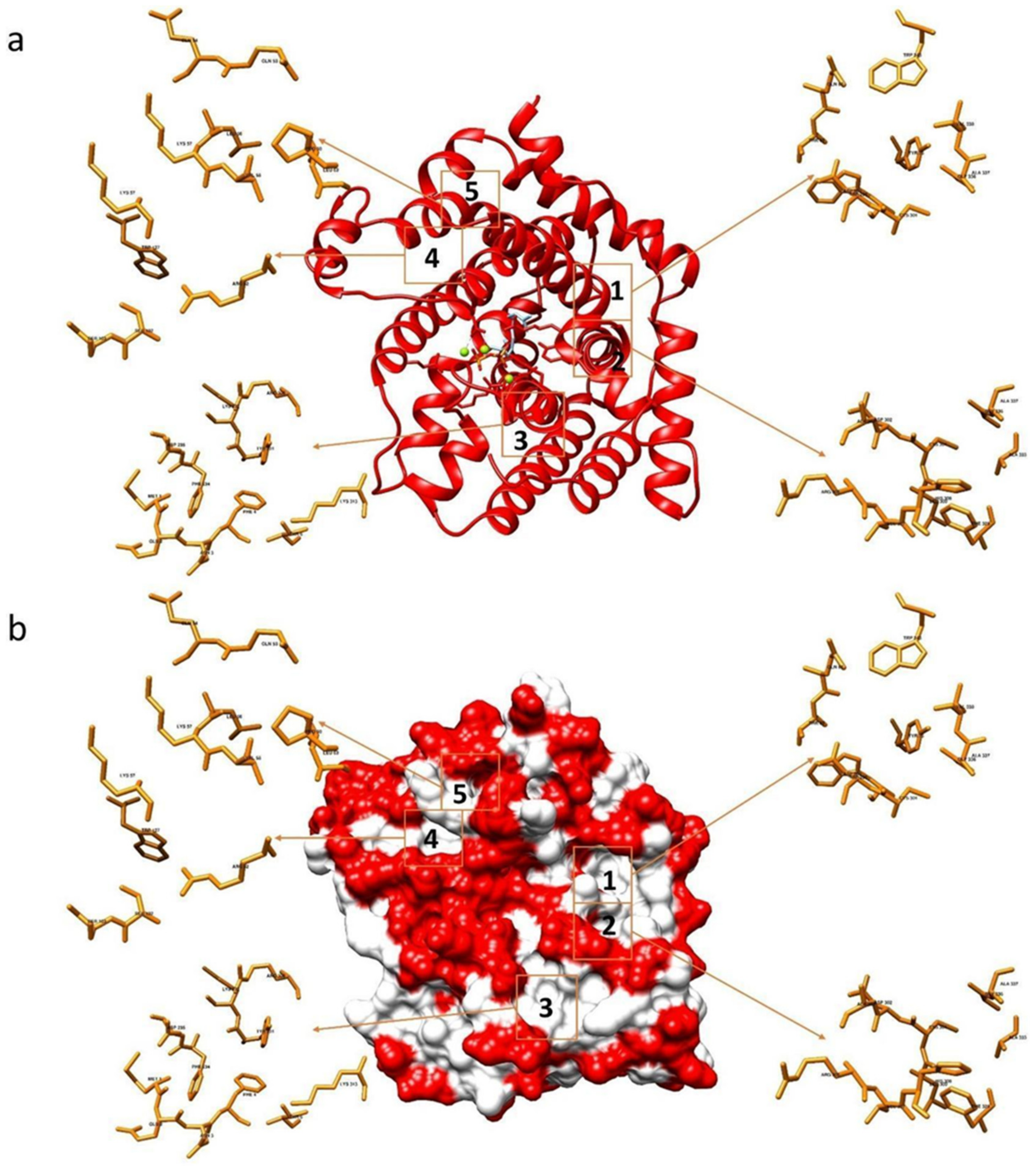
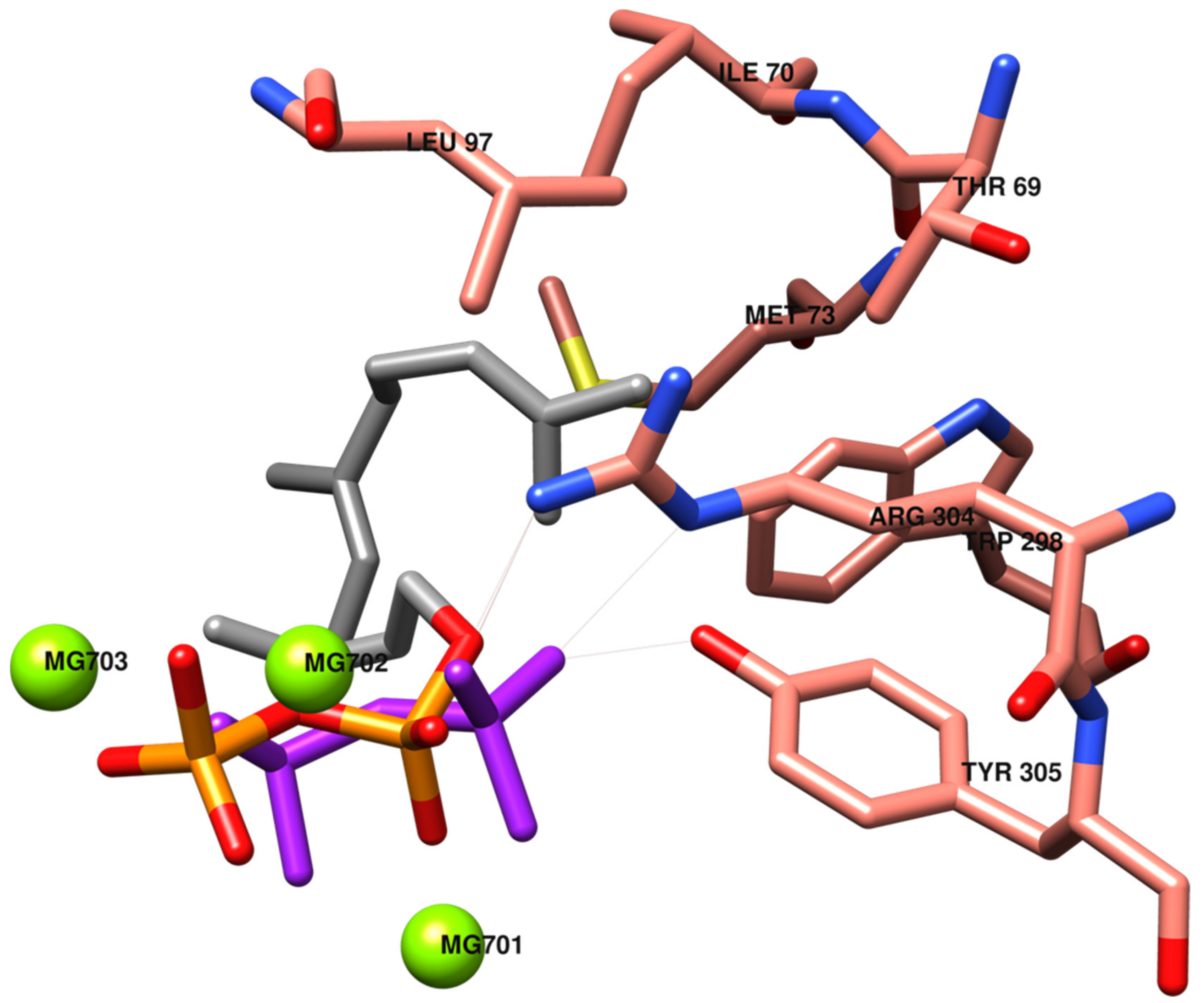
2.1. Docking of Phenols and Hydroxylated Biphenyls to TRI5
2.2. Antifungal Activity and Trichothecene Inhibitory Effect of Tested Phenols against F. culmorum Strain UK99
2.2.1. In Vitro Assay
2.2.2. Greenhouse Assay
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Fungal Strain and Culture Conditions
5.2. Selected Phenolic Compounds
5.3. Amendment with Compounds 1–15 and Sample Preparation for DON Analysis
5.4. Greenhouse Assay
5.5. Analytical Method, Instruments and Equipment for The Detection of Type B Trichothecenes
5.5.1. Sample Preparation
5.5.2. Q-Orbitrap-HRMS Analyses
5.6. Statistical Analysis
5.7. Molecular Docking: In Silico Analysis of Selected Phenols-TRI5 Binding
5.8. Estimation of Selected Physicochemical Descriptors
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Goswami, R.S.; Kistler, H.C. Heading for disaster: Fusarium graminearum on cereal crops. Mol. Plant Pathol. 2004, 5, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Miedaner, T.; Cumagun, C.J.R.; Chakraborty, S. Population genetics of three important head blight pathogens Fusarium graminearum, F. pseudograminearum and F. culmorum. J. Phytopathol. 2008, 156, 129–139. [Google Scholar] [CrossRef]
- Scherm, B.; Balmas, V.; Spanu, F.; Pani, G.; Delogu, G.; Pasquali, M.; Migheli, Q. Fusarium culmorum: Causal agent of foot and root rot and head blight on wheat. Mol. Plant Pathol. 2013, 14, 323–341. [Google Scholar] [CrossRef]
- Haile, J.K.; N’Diaye, A.; Walkowiak, S.; Nilsen, K.T.; Clarke, J.M.; Kutcher, H.R.; Steiner, B.; Buerstmayr, H.; Pozniak, C.J. Fusarium head blight in durum wheat: Recent status, breeding directions, and future research prospects. Phytopathology 2019, 109, 1664–1675. [Google Scholar] [CrossRef]
- Fernando, W.D.; Oghenekaro, A.O.; Tucker, J.R.; Badea, A. Building on a foundation: Advances in epidemiology, resistance breeding, and forecasting research for reducing the impact of Fusarium head blight in wheat and barley. Can. J. Plant Pathol. 2021, 43, 495–526. [Google Scholar] [CrossRef]
- Pasquali, M.; Migheli, Q. Genetic approaches to chemotype determination in type B trichothecene producing Fusaria. Int. J. Food Microbiol. 2014, 189, 164–182. [Google Scholar] [CrossRef]
- Foroud, N.A.; Baines, D.; Gagkaeva, T.Y.; Thakor, N.; Badea, A.; Steiner, B.; Bürstmayr, M.; Bürstmayr, H. Trichothecenes in Cereal grains—An update. Toxins 2019, 11, 634. [Google Scholar] [CrossRef] [Green Version]
- Proctor, R.H.; McCormick, S.P.; Gutiérrez, S. Genetic bases for variation in structure and biological activity of trichothecene toxins produced by diverse fungi. Appl. Microbiol. Biotechnol. 2020, 104, 5185–5199. [Google Scholar] [CrossRef]
- Sudakin, D.L. Trichothecenes in the environment: Relevance to human health. Toxicol. Lett. 2003, 143, 97–107. [Google Scholar] [CrossRef]
- Zhang, J.; You, L.; Wu, W.; Wang, X.; Chrienova, Z.; Nepovimova, E.; Qinghua, W.; Kuca, K. The neurotoxicity of trichothecenes T-2 toxin and deoxynivalenol (DON): Current status and future perspectives. Food Chem. Toxicol. 2020, 145, 111676. [Google Scholar] [CrossRef]
- Wei, C.M.; McLaughlin, C.S. Structure-function relationship in 12, 13-epoxy trichothecenes novel inhibitors of protein-synthesis. Biochem. Biophys. Res. Commun. 1974, 57, 838–844. [Google Scholar] [CrossRef]
- Desmond, O.J.; Manners, J.M.; Stephens, A.E.; Maclean, D.J.; Schenk, P.M.; Gardiner, D.M.; Munn, A.L.; Kazan, K. The Fusarium mycotoxin deoxynivalenol elicits hydrogen peroxide production, programmed cell death and defence responses in wheat. Mol. Plant Pathol. 2008, 9, 435–445. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.H.; Jarvis, B.B.; Chung, Y.J.; Pestka, J.J. Apoptosis induction by the satratoxins and other trichothecene mycotoxins: Relationship to ERK, p8 MAPK, and SAPK/JNK activation. Toxicol. Appl. Pharmacol. 2000, 164, 149–160. [Google Scholar] [CrossRef]
- Spanu, F.; Scherm, B.; Camboni, I.; Balmas, V.; Pani, G.; Oufensou, S.; Macciotta, N.; Pasquali, M.; Migheli, Q. FcRav2, a gene with a ROGDI domain involved in Fusarium head blight and crown rot on durum wheat caused by Fusarium culmorum. Mol. Plant Pathol. 2018, 19, 677–688. [Google Scholar] [CrossRef] [Green Version]
- Rauwane, M.E.; Ogugua, U.V.; Kalu, C.M.; Ledwaba, L.K.; Woldesemayat, A.A.; Ntushelo, K. Pathogenicity and virulence factors of Fusarium graminearum including factors discovered using next generation sequencing technologies and proteomics. Microorganisms 2020, 8, 305. [Google Scholar] [CrossRef] [Green Version]
- Hohn, T.M.; Beremand, P. Isolation and nucleotide sequence of a sesquiterpene cyclase gene from the trichothecene-producing fungus Fusarium sporotrichioides. Gene 1989, 79, 131–138. [Google Scholar] [CrossRef]
- Vedula, L.S.; Zhao, Y.; Coates, R.M.; Koyama, T.; Cane, D.; Christianson, D.W. Exploring biosynthetic diversity with trichodiene synthase. Arch. Biochem. Biophys. 2007, 466, 260–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vedula, L.S.; Jiang, J.; Zakharian, T.; Cane, D.E.; Christianson, D.W. Structural and mechanistic analysis of trichodiene synthase using site-directed mutagenesis: Probing the catalytic function of tyrosine-295 and the asparagine-225/serine-229/glutamate-233-Mg2+ B motif. Arch. Biochem. Biophys. 2008, 469, 184–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christianson, D.W. Structural biology and chemistry of the terpenoid cyclases. Chem. Rev. 2006, 106, 3412–3442. [Google Scholar] [CrossRef] [PubMed]
- Rynkiewicz, M.J.; Cane, D.E.; Christianson, D.W. X-ray crystal structures of D100E trichodiene synthase and its pyrophosphate complex reveal the basis for terpene product diversity. Biochemistry 2002, 41, 1732–1741. [Google Scholar] [CrossRef] [PubMed]
- Pani, G.; Scherm, B.; Azara, E.; Balmas, V.; Jahanshiri, Z.; Carta, P.; Fabbri, D.; Dettori, M.A.; Fadda, A.; Dessì, A.; et al. Natural and natural-like phenolic inhibitors of type B trichothecene in vitro production by the wheat (Triticum sp.) pathogen Fusarium culmorum. J. Agric. Food Chem. 2014, 62, 4969–4978. [Google Scholar] [CrossRef] [PubMed]
- Chala, A.; Weinert, J.; Wolf, G.A. An integrated approach to the evaluation of the efficacy of fungicides against Fusarium culmorum, the cause of head blight of wheat. J. Phytopathol. 2003, 151, 673–678. [Google Scholar] [CrossRef]
- Haidukowski, M.; Pascale, M.; Perrone, G.; Pancaldi, D.; Campagna, C.; Visconti, A. Effect of fungicides on the development of Fusarium head blight, yield and deoxynivalenol accumulation in wheat inoculated under field conditions with Fusarium graminearum and Fusarium culmorum. J. Sci. Food Agric. 2005, 85, 191–198. [Google Scholar] [CrossRef]
- Simpson, D.R.; Weston, G.E.; Turner, J.A.; Jennings, P.; Nicholson, P. Differential control of head blight pathogens of wheat by fungicides and consequences for mycotoxin contamination of grain. Eur. J. Plant Pathol. 2001, 107, 421–431. [Google Scholar] [CrossRef]
- Picot, A.; Atanasova-Pénichon, V.; Pons, S.; Marchegay, G.; Barreau, C.; Pinson-Gadais, L.; Roucolle, J.; Daveau, F.; Caron, D.; Richard-Forget, F. Maize kernel antioxidants and their potential involvement in Fusarium Ear Rot resistance. J. Agric. Food Chem. 2013, 61, 3389–3395. [Google Scholar] [CrossRef] [PubMed]
- Barral, B.; Chillet, M.; Minier, J.; Léchaudel, M.; Schorr-Galindo, S. Evaluating the response to Fusarium ananatum inoculation and antifungal activity of phenolic acids in pineapple. Fungal Biol. 2017, 121, 1045–1053. [Google Scholar] [CrossRef]
- Zhou, X.; Jia, H.; Ge, X.; Fengzhi, W. Effects of vanillin on the community structures and abundances of Fusarium and Trichoderma spp. in cucumber seedling rhizosphere. J. Plant Interact. 2018, 13, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Pizzolitto, R.P.; Jacquat, A.G.; Usseglio, V.L.; Achimón, F.; Cuello, A.E.; Zygadlo, J.A.; Dambolena, J.S. Quantitative-structure-activity relationship study to predict the antifungal activity of essential oils against Fusarium verticillioides. Food Control 2020, 108, 106836. [Google Scholar] [CrossRef]
- Bakan, B.; Bily, A.C.; Melcion, D.; Cahagnier, B.; Regnault-Roger, C.; Philogène, B.J.; Richard-Molard, D. Possible role of plant phenolics in the production of trichothecenes by Fusarium graminearum strains on different fractions of maize kernels. J. Agric. Food Chem. 2003, 51, 2826–2831. [Google Scholar] [CrossRef]
- Boutigny, A.L.; Atanasova-Pénichon, V.; Benet, M.; Barreau, C.; Richard-Forget, F. Natural phenolic acids from wheat bran inhibit Fusarium culmorum trichothecene biosynthesis in vitro by repressing Tri gene expression. Eur. J. Plant Pathol. 2010, 127, 275–286. [Google Scholar] [CrossRef]
- Pagnussatt, F.A.; Del Ponte, E.M.; Garda-Buffon, J.; Badiale-Furlong, E. Inhibition of Fusarium graminearum growth and mycotoxin production by phenolic extract from Spirulina sp. Pestic. Biochem. Physiol. 2014, 108, 21–26. [Google Scholar] [CrossRef]
- Pani, G.; Dessì, A.; Dallocchio, R.; Scherm, B.; Azara, E.; Delogu, G.; Migheli, Q. Natural phenolic inhibitors of trichothecene biosynthesis by the wheat fungal pathogen Fusarium culmorum: A computational insight into the structure-activity relationship. PLoS ONE 2016, 11, e0157316. [Google Scholar] [CrossRef] [PubMed]
- Ferruz, E.; Loran, S.; Herrera, M.; Gimenez, I.; Bervis, N.; Barcena, C.; Carramiñana, J.J.; Juan, T.; Herrera, A.; Ariño, A. Inhibition of Fusarium growth and mycotoxin production in culture medium and in maize kernels by natural phenolic acids. J. Food Prot. 2016, 79, 1753–1758. [Google Scholar] [CrossRef] [PubMed]
- Schöneberg, T.; Martin, C.; Wettstein, F.E.; Bucheli, T.D.; Mascher, F.; Bertossa, M.; Musa, T.; Keller, B.; Vogelgsang, S. Fusarium and mycotoxin spectra in Swiss barley are affected by various cropping techniques. Food Addit. Contam. 2016, 33, 1608–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulik, T.; Stuper-Szablewska, K.; Bilska, K.; Buśko, M.; Ostrowska-Kołodziejczak, A.; Załuski, D.; Perkowski, J. Trans-cinnamic and chlorogenic acids affect the secondary metabolic profiles and ergosterol biosynthesis by Fusarium culmorum and F. graminearum sensu stricto. Toxins 2017, 9, 198. [Google Scholar] [CrossRef]
- Kulik, T.; Stuper-Szablewska, K.; Bilska, K.; Buśko, M.; Ostrowska-Kołodziejczak, A.; Załuski, D.; Perkowski, J. Sinapic acid affects phenolic and trichothecene profiles of F. culmorum and F. graminearum sensu stricto. Toxins 2017, 9, 264. [Google Scholar] [CrossRef] [Green Version]
- Scaglioni, P.T.; Scarpino, V.; Marinaccio, F.; Vanara, F.; Badiale-Furlong, E.; Blandino, M. Impact of microalgal phenolic extracts on the control of Fusarium graminearum and deoxynivalenol contamination in wheat. World Mycotoxin J. 2019, 12, 367–378. [Google Scholar] [CrossRef]
- Oufensou, S.; Scherm, B.; Pani, G.; Balmas, V.; Fabbri, D.; Dettori, M.A.; Carta, P.; Malbrán, I.; Migheli, Q.; Delogu, G. Honokiol, magnolol and its monoacetyl derivative show strong antifungal effect on Fusarium isolates of clinical relevance. PLoS ONE 2019, 14, e0221249. [Google Scholar] [CrossRef]
- Oufensou, S.; Balmas, V.; Azara, E.; Fabbri, D.; Dettori, M.A.; Schüller, C.; Zehetbauer, F.; Strauss, J.; Delogu, G.; Migheli, Q. Naturally occurring phenols modulate vegetative growth and deoxynivalenol biosynthesis in Fusarium graminearum. ACS Omega 2020, 5, 29407–29415. [Google Scholar] [CrossRef]
- Malbrán, I.; Mourelos, C.A.; Pardi, M.; Oufensou, S.; Balmas, V.; Delogu, G.; Migheli, Q.; Lori, G.A.; Juárez, M.P.; Girotti, J.A. Commercially available natural inhibitors of trichothecene production in Fusarium graminearum: A strategy to manage Fusarium head blight of wheat. Crop. Prot. 2020, 138, 105313. [Google Scholar] [CrossRef]
- Montibus, M.; Vitrac, X.; Coma, V.; Loron, A.; Pinson-Gadais, L.; Ferrer, N.; Verdal-Bonnin, M.N.; Gabaston, J.; Waffo-Téguo, P.; Richard-Forget, F.; et al. Screening of wood/forest and vine by-products as sources of new drugs for sustainable strategies to control Fusarium graminearum and the production of mycotoxins. Molecules 2021, 26, 405. [Google Scholar] [CrossRef] [PubMed]
- Boutigny, A.L.; Barreau, C.; Atanasova-Pénichon, V.; Verdal-Bonnin, M.N.; Pinson-Gadais, L.; Richard-Forget, F. Ferulic acid, an efficient inhibitor of type B trichothecene biosynthesis and Tri gene expression in Fusarium liquid cultures. Mycol. Res. 2009, 113, 746–753. [Google Scholar] [CrossRef] [PubMed]
- Dambolena, J.S.; López, A.G.; Meriles, J.M.; Rubinstein, H.R.; Zygadlo, J.A. Inhibitory effect of 10 natural phenolic compounds on Fusarium verticillioides. A structure–property–activity relationship study. Food Control 2012, 28, 163–170. [Google Scholar] [CrossRef]
- Chen, C.; Long, L.; Zhang, F.; Chen, Q.; Chen, C.; Yu, X.; Liu, Q.; Bao, J.; Long, Z. Antifungal activity, main active components and mechanism of Curcuma longa extract against Fusarium graminearum. PLoS ONE 2018, 13, e0194284. [Google Scholar]
- Zhang, M.; Ge, J.; Yu, X. Transcriptome analysis reveals the mechanism of fungicidal of thymol against Fusarium oxysporum f. sp. niveum. Curr. Microbiol. 2018, 75, 410–419. [Google Scholar] [CrossRef]
- Boddu, J.; Cho, S.; Muehlbauer, G.J. Transcriptome analysis of trichothecene-induced gene expression in Barley. Mol. Plant-Microbe Interact. 2007, 20, 1364–1375. [Google Scholar] [CrossRef] [Green Version]
- Boutigny, A.L.; Richard-Forget, F.; Barreau, C. Natural mechanisms for cereal resistance to the accumulation of Fusarium trichothecenes. Eur. J. Plant Pathol. 2008, 121, 411–423. [Google Scholar] [CrossRef]
- Atanasova-Pénichon, V.; Barreauand, C.; Richard-Forget, F. Antioxidant secondary metabolites in cereals: Potential involvement in resistance to Fusarium and mycotoxin accumulation. Front. Microbiol. 2016, 7, 566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadjout, S.; Chéreau, S.; Atanasova-Pénichon, V.; Marchegay, G.; Mekliche, L.; Boureghda, H.; Barreau, C.; Touati-Hattab, S.; Bouznad, Z.; Richard-Forget, F. Phenotypic and biochemical characterization of new advanced durum wheat breeding lines from Algeria that show resistance to Fusarium head blight and to mycotoxin accumulation. J. Plant Pathol. 2017, 99, 671–680. [Google Scholar]
- Kimura, M.; Tokai, T.; Takahashi-Ando, N.; Ohsato, S.; Fujimura, M. Molecular and genetic studies of Fusarium trichothecene biosynthesis: Pathways, genes and evolution. Biosci. Biotechnol. Biochem. 2007, 71, 2105–2123. [Google Scholar] [CrossRef] [Green Version]
- Rynkiewicz, M.J.; Cane, D.E.; Christianson, D.W. Structure of trichodiene synthase from Fusarium sporotrichioides provides mechanistic inferences on the terpene cyclization cascade. Proc. Natl. Acad. Sci. USA 2001, 98, 13543–13548. [Google Scholar] [CrossRef] [Green Version]
- Maeda, K.; Nakajima, Y.; Motoyama, T.; Kondoh, Y.; Kawamura, T.; Kanamaru, K.; Ohsato, S.; Nishiuchi, T.; Yoshida, M.; Osada, H.; et al. Identification of a trichothecene production inhibitor by chemical array and library screening using trichodiene synthase as a target protein. Pestic. Biochem. Physiol. 2017, 138, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Wolf, V.G.; Bonacorsi, C.; Raddi, M.S.G.; da Fonseca, L.M.; Ximenes, V.F. Octyl gallate, a food additive with potential beneficial properties to treat Helicobacter pylori infection. Food Funct. 2017, 8, 2500–2511. [Google Scholar] [CrossRef]
- Singh, S.; Fatima, Z.; Hameed, S. Octyl gallate reduces ABC multidrug transporter CaCdr1p expression and leads to its mislocalisation in azole-resistant clinical isolates of Candida albicans. J. Glob. Antimicrob. Resist. 2020, 22, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Chan, K.L.; Cheng, L.W. Octyl gallate as an intervention catalyst to augment antifungal efficacy of Caspofungin. Multidiscip. Sci. J. 2018, 1, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.H.; Lu, M.H.; Guo, D.S.; Zhai, Y.Y.; Miao, D.; Yue, J.Y.; Yuan, C.H.; Zhao, M.M.; An, D.R. Antifungal effect of magnolol and honokiol from Magnolia officinalis on Alternaria alternata causing tobacco brown spot. Molecules 2019, 24, 2140. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Liu, Z.Q. Comparison of antioxidant abilities of magnolol and honokiol to scavenge radicals and to protect DNA. Biochimie 2011, 93, 1755–1760. [Google Scholar] [CrossRef]
- Amorati, R.; Zotova, J.; Baschieri, A.; Valgimigli, L. Antioxidant activity of magnolol and honokiol: Kinetic and mechanistic investigations of their reaction with peroxyl radicals. J. Org. Chem. 2015, 60, 10651–10659. [Google Scholar] [CrossRef]
- Rachitha, P.; Krupashree, K.; Jayashree, G.V.; Gopalan, N.; Khanum, F. Growth Inhibition and morphological alteration of Fusarium sporotrichioides by Mentha piperita essential oil. Pharmacogn. Res. 2017, 9, 74–79. [Google Scholar]
- Liu, D.; Pan, Y.; Li, K.; Li, D.; Li, P.; Gao, Z. Proteomics reveals the mechanism underlying the inhibition of Phytophthora sojae by propyl gallate. J. Agric. Food Chem. 2020, 68, 8151–8162. [Google Scholar] [CrossRef]
- Takai, E.; Hirano, A.; Shiraki, K. Effects of alkyl chain length of gallate on self-association and membrane binding. J. Biochem. 2011, 150, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Hyldgaard, M.; Mygind, T.; Meyer, R.L. Essential oils in food preservation: Mode of action, synergies, and interactions with food matrix components. Front. Microbiol. 2012, 3, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, T.; Zhou, H.; Zhou, W.; Hu, L.; Chen, J.; Shi, Z. The fungicidal activity of thymol against Fusarium graminearum via inducing lipid peroxidation and disrupting ergosterol biosynthesis. Molecules 2016, 21, 770. [Google Scholar] [CrossRef] [Green Version]
- Divband, K.; Shokri, H.; Khosravi, A.R. Down-regulatory effect of Thymus vulgaris L. on growth and Tri4 gene expression in Fusarium oxysporum strains. Microb. Pathog. 2017, 104, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Fernández, J.; García-Armesto, M.R.; Álvarez-Alonso, R.; del Valle, P.; de Arriaga, D.; Rúa, J. Antimicrobial activity of binary combinations of natural and synthetic phenolic antioxidants against Enterococcus faecalis. J. Dairy Sci. 2013, 96, 4912–4920. [Google Scholar] [CrossRef] [Green Version]
- de Castro, R.D.; de Souza, T.M.; Bezerra, L.M.; Ferreira, G.L.; Costa, E.M.; Cavalcanti, A.L. Antifungal activity and mode of action of thymol and its synergism with nystatin against Candida species involved with infections in the oral cavity: An in vitro study. BMC Complement. Altern. Med. 2015, 15, 417. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Liao, K.; Hang, C.; Wang, D. Honokiol induces reactive oxygen species-mediated apoptosis in Candida albicans through mitochondrial dysfunction. PLoS ONE 2017, 12, e0172228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Liao, K.; Wang, D. Effect of magnolol and honokiol on adhesion, yeast-hyphal transition, and formation of biofilm by Candida albicans. PLoS ONE 2015, 10, e0117695. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z. Plant-derived antifungal compounds trigger a common transcriptional response. Inf. Genet. Evol. 2017, 54, 1–3. [Google Scholar] [CrossRef]
- Wang, Z.; Shen, Y. Antifungal compound honokiol triggers oxidative stress responsive signalling pathway and modulates central carbon metabolism. Mycology 2016, 7, 124–133. [Google Scholar] [CrossRef] [Green Version]
- Lowe, R.G.T.; Allwood, J.W.; Galster, A.; Urban, M.; Daudi, A.; Canning, C.; Ward, J.L.; Beale, M.H.; Hammond-Kosack, K.E. A combined HNMR and ESI-MS analysis to understand the basal metabolism of plant pathogenic Fusarium species. Mol. Plant-Microbe Interact. 2010, 23, 1605–1618. [Google Scholar] [CrossRef] [Green Version]
- De Farias Dias, A. An improved high yield synthesis of dehydrodieugenol. Phytochemistry 1988, 27, 3008–3009. [Google Scholar] [CrossRef]
- Russell, W.R.; Scobbie, L.; Chesson, A. Structural modification of phenylpropanoid-derived compounds and the effects on their participation in redox processes. Bioorganic Med. Chem. 2005, 13, 2537–2546. [Google Scholar] [CrossRef] [PubMed]
- Vogel, H.J. A convenient growth medium for Neurospora (Med N). Microbiol. Genet. Bull. 1956, 13, 42–43. [Google Scholar]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple ligand-protein interaction diagrams for drug discovery. J. Chem. Inf. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| PPi | |||||
|---|---|---|---|---|---|
| Tested Ligands | % | Sites | E.F.E.B. a | E.I.C., Ki b | Interaction with Amino Acids |
| p-Coumaric acid 1 | 75 | c.d. c | −13.23 | 199.19 pM | Asp100 Glu164 Pro178 Leu181 Arg182 Asn185 Asp239 Ile241 Ser242 Leu243 Asn246 PPi700 Mg702 Mg703 |
| 1 | c.d. | −7.73 | 2.17 uM | Ile70 Met73 Tyr93 Leu97 Asp100 Phe157 Arg182 Asn185 Leu187 PPi700 Mg703 | |
| 4 | 3 | −5.59 | 79.22 uM | Met1 Glu2 Asn3 Phe4 Thr6 Phe234 Asp235 Lys313 | |
| 11 | 3 | −5.56 | 84.50 uM | Phe4 Thr6 Tyr231 Phe234 Asp235 Arg306 Lys313 | |
| Caffeic acid 2 | 39 | c.d. | −12.06 | 1.45 nM | Asp100 Glu164 Leu181 Arg182 Asn185 Asp239 Ile241 Ser242 Leu243 Asn246 PPi700 Mg702 Mg703 |
| 16 | 4 | −5.81 | 55.08 uM | Met55 Leu56 Lys57 Arg62 Val98 Ser102 Ser103 Pro126 Trp127 | |
| 11 | 3 | −5.45 | 101.75 uM | Met1 Glu2 Asn3 Phe4 Thr6 Thr231 Phe234 Asp235 Lys313 | |
| Ferulic acid 3 [32] | 2 | c.d. | −9.62 | 89.12 nM | Asp100 Glu164 Leu181 Arg182 Asn185 Asp226 Glu233 Arg238 Asp239 Ser242 Leu243 Asn246 PPi700 Mg703 |
| 6 | c.d. | −7.78 | 1.99 uM | Ile70 Met73 Tyr93 Thr96 Asp100 Phe157 Arg182 Asn185 Leu187 PPi700 Mg703 | |
| 13 | 3 | −5.58 | 80.65 uM | Met1 Glu2 Asn3 Phe4 Tyr231 Lys232 Phe234 Asp235 Arg306 | |
| 8 | 4 | −5.51 | 92.16 uM | Met55 Leu56 Lys57 Arg62 Val98 Ser102 Ser103 Pro126 Trp127 | |
| 14 | 3 | −5.48 | 96.45 uM | Phe4 Thr6 Tyr231 Phe234 Asp235 Arg306 Lys313 | |
| 19 | 5 | −5.19 | 157.79 uM | Gln53 Gln54 Leu56 Lys57 Val58 Pro60 Leu63 | |
| Propyl gallate 4 [32] | 40 | 1 | −5.86 | 50.71 uM | Ala65 Gln68 Thr69 Tyr76 Trp298 Cys301 Asp302 Ala303 Gly336 Val338 Pro340 Trp343 |
| 13 | 2 | −5.28 | 133.84 uM | Leu300 Cys301 Asp302 Ala303 Arg306 Leu307 His308 Phe329 Ala333 Ala337 | |
| Octyl gallate 5 | 12 | 2-1 | −6.42 | 19.85 uM | Gly72 Tyr76 Leu300 Cys301 Asp302 Ala303 Arg306 Leu307 His308 Phe329 Gly336 Ala337 Val338 |
| 9 | 1 | −6.30 | 23.91 uM | Ala65 Ser66 Gln68 Thr69 Tyr76 Cys301 Asp302 Ala303 Arg304 Gly336 Val338 Trp343 | |
| 16 | 1-2 | −6.28 | 24.94 uM | Gln68 Thr69 Gly72 Cys301 Asp302 Ala303 Leu307 His308 Phe329 Gly336 Ala337 Val338 Trp343 | |
| 5 | 1-2 | −5.83 | 53.08 uM | Leu36 Gly72 Tyr76 Leu300 Cys301 Asp302 Ala303 Arg306 Leu307 His308 Gly336 Val338 Trp343 | |
| Carvacrol 6 | 48 | 1 | −5.31 | 127.65 uM | Leu36 Gln68 Thr69 Gly72 Tyr76 Cys301 Asp302 Gly336 Ala337 Val338 Trp343 |
| 21 | 2 | −5.29 | 132.10 uM | Leu300 Cys301 Asp302 Ala303 Arg306 Leu307 His308 Phe329 Ala333 | |
| 11 | c.d. | −5.27 | 136.69 uM | Met73 Tyr93 Thr96 Leu97 Asp100 Phe157 Leu187 Val191 Met221 | |
| 16 | 4 | −5.18 | 160.32 uM | Met55 Lys57 Arg62 Val98 Leu99 Ser102 Asp104 Pro126 Trp127 | |
| Thymol 7 | 5 | c.d. | −5.63 | 74.28 uM | Glu164 Pro178 Leu181 Arg182 Asn185 Asp226 Glu233 Ile241 Ser242 Leu243 Asn246 PPi700 |
| 20 | 4 | −5.50 | 92.72 uM | Met55 Leu56 Lys57 Val58 Val98 Leu99 Asp104 His125 Pro126 Trp127 | |
| 58 | 1 | −5.34 | 121.84 uM | Tyr76 Trp298 Cys301 Asp302 Gly336 Val338 Pro340 Trp343 | |
| 10 | c.d. | −5.17 | 161.91 uM | Ile70 Met73 Tyr93 Thr96 Leu97 Asp100 Phe157 Leu187 Val191 PPi700 | |
| R-Linalool 8 | 36 | 1 | −5.32 | 126.02 uM | Leu36 Gln68 Thr69 Gly72 Tyr76 Cys301 Asp302 Gly336 Ala337 Val338 Trp343 |
| 21 | c.d. | −5.31 | 128.71 uM | Ile70 Met73 Trp78 Tyr93 Thr96 Leu97 Phe157 Leu187 Val191 Met221 Tyr295 | |
| 12 | 1 | −5.24 | 145.29 uM | Leu36 Ala65 Gln68 Thr69 Gly72 Tyr76 Asp302 Ala303 Trp343 | |
| 10 | 4 | −5.07 | 193.14 uM | Met55 Leu56 Lys57 Val98 His125 Pro126 Trp127 | |
| S-Linalool 8 | 11 | 4 | −5.26 | 140.56 uM | Met55 Leu56 Lys57 Arg62 Val98 His125 Pro126 Trp127 |
| 36 | 1 | −5.22 | 148.58 uM | Gln68 Thr69 Tyr76 Cys301 Asp302 Gly336 Val338 Pro340 Trp343 | |
| 27 | c.d. | −5.18 | 160.63 uM | Ile70 Met73 Trp78 Tyr93 Thr96 Leu97 Val191 Met221 Tyr295 | |
| 17 | 1 | −5.14 | 171.53 uM | Ala65 Gln68 Thr69 Gly72 Tyr76 Cys301 Asp302 Ala303 Trp343 | |
| Geraniol 9 | 2 | c.d. | −6.60 | 14.53 uM | Pro178 Leu181 Arg182 Asn185 Glu233 Arg238 Asp239 Ile241 Ser242 Leu243 Asn246 PPi700 Mg702 MGg703 |
| 12 | 2 | −5.42 | 106.54 uM | Leu300 Cys301 Asp302 Ala303 Tyr305 Arg306 Leu307 His308 Glu309 Phe329 | |
| 47 | 1 | −5.30 | 131.30 uM | Gln68 Thr69 Gly72 Tyr76 Cys301 Asp302 Gly336 Ala337 Val338 Pro340 Trp343 | |
| 16 | c.d. | −5.04 | 200.93 uM | Ile70 Met73 Tyr93 Thr96 Leu97 Phe157 Arg182 Leu187 Met221 Asn225 Tyr295 Trp298 Tyr305 PPi700 | |
| Apocynin 10 [32] | 4 | c.d. | −6.86 | 9.29 uM | Asp100 Glu164 Arg182 Asn185 Asp226 Glu233 Arg238 Asp239 Ser242 Leu243 PPi700 Mg703 |
| 60 | 1 | −5.35 | 119.64 uM | Gln68 Tyr76 Cys301 Asp302 Gly336 Ala337 Val338 Trp343 | |
| 11 | 1 | −5.25 | 134.28 uM | Gln68 Tyr69 Gly72 Tyr76 Trp298 Cys301 Asp302 Gly336 Ala337 Val338 | |
| Eugenol 11 [32] | 3 | c.d. | −5.65 | 72.36 uM | Asp100 Glu164 Leu181 Arg182 Asn185 Asp226 Glu233 Arg238 Asp239 Ile241 Ser242 Leu243 Asn246 PPi700 |
| 31 | 2 | −5.12 | 177.48 uM | His299 Leu300 Cys301 Asp302 Ala303 Arg306 Leu307 His308 Glu309 Phe329 | |
| 19 | 2 | −5.07 | 193.10 uM | His299 Leu300 Cys301 Asp302 Ala303 Arg306 Leu307 His308 Phe329 Ala333 Ala337 | |
| 22 | 1 | −4.98 | 224.72 uM | Gln68 Thr69 Tyr76 Trp298 Cys301 Asp302 Gly336 Ala337 Val338 Trp343 | |
| Magnolol 12 [32] | 29 | 1 | −6.91 | 8.56 uM | Gln68 Thr69 Gly72 Tyr76 Leu300 Cys301 Asp302 Phe329 Gly336 Ala337 Val338 Pro340 |
| 11 | 1-2 | −6.49 | 17.43 uM | Leu36 Gly72 Tyr76 Trp298 Leu300 Cys301 Asp302 Ala303 Phe329 Ala333 Ala337 Val338 Pro340 Trp343 | |
| 14 | 4 | −5.91 | 46.77 uM | Met55 Leu56 Lys57 Val58 Arg62 Val98 Ser102 Ser103 Pro126 Trp127 | |
| Honokiol 13 | 25 | 1-2 | −7.25 | 4.81 uM | Gln68 Thr69 Gly72 Trp298 Leu300 Cys301 Asp302 Phe329 Ala333 Ala337 Val338 Trp343 |
| 6 | 2 | −7.15 | 5.72 uM | Leu300 Cys301 Asp302 Ala303 Tyr305 Arg306 Leu307 His308 Tyr311 Phe329 Glu330 Ala333 Ala337 | |
| 24 | 1-2 | −7.04 | 6.91 uM | Ala65 Gln68 Thr69 Gly72 Tyr76:Trp298 Cys301 Asp302 Ala303 Gly336 Ala337 Val338 Pro340 | |
| 16 | 1-2 | −6.92 | 8.44 uM | Gln68 Thr69 Gly72 Tyr76 Trp298 Cys301 Asp302 His308 Ala337 Val338 | |
| Eugenol dimer 14 [32] | 14 | 1-2 | −6.69 | 12.41 uM | Gln68 Gly72 Tyr76 Trp298 Cys301 Asp302 Ala303 His308 Ala337 Val338 |
| 28 | 1-2 | −6.67 | 12.98 uM | Thr69 Gly72 Tyr76 Trp298 Leu300 Cys301 Asp302 Ala303 Phe329 Ala333 Ala337 Val338 Ala339 Pro340 Trp343 | |
| 10 | 1-2 | −6.27 | 25.41 uM | Leu36 Gln68 Thr69 Tyr76 Leu300 Cys301 Asp3Ala303 Phe329 Ala333 Gly336 Ala337 Val338 Trp343 | |
| 14 | 4 | −5.41 | 108.76 uM | Lys57 Val58 Arg62 Val98 Leu99 Ser102 Ser103 Asp104 Pro126 Trp127 | |
| Ferulic acid dimer 15 | 4 | 1-2 | −6.48 | 17.71 uM | Leu36 Ala65 Gln68 Thr69 Gly72 Tyr76 Cys301 Asp302 Ala303 Arg304 Ala337 Pro340 Trp343 |
| 26 | 2-1 | −5.55 | 85.48 uM | Gln68 Thr69 Tyr76 Trp298 Cys301 Asp302 Ala303 Gly336 Ala337 Val338 Ala339 Pro340 | |
| NPD352 | 6 | 1-2 | −9.52 | 104.59 nM | Tyr76 Cys301 Asp302 Ala303 His308 Tyr311 Phe329 Glu330 Ala333 Ala337 Val338 Pro340 Trp343 |
| 22 | 1-2 | −8.05 | 1.25 uM | Gly72 Tyr76 Cys301 Asp302 Ala303 His308 Phe329 Ala333 Ala337 Val338 Ala339 Trp343 | |
| FPP | |||||
|---|---|---|---|---|---|
| Tested Ligands | % | Sites | E.F.E.B. a | E.I.C., Ki b | Interaction with Amino Acids |
| p-Coumaric acid 1 | 27 | 3 | −5.50 | 92.97 uM | Glu2 Phe4 Thr6 Phe234 Asp235 Lys313 |
| 45 | 3 | −5.47 | 98.12 uM | Phe4 Thr6 Phe234 Asp235 Arg306 Lys313 | |
| 22 | 5 | −5.21 | 139.08 uM | Gln54 Leu56 Lys57 Val58 Pro60 Leu63 | |
| Caffeic acid 2 | 20 | 4 | −5.62 | 75.82 uM | Met55 Leu56 Lys57 Arg62 Val98 Ser103 Pro126 Trp127 |
| 15 | 5 | −5.48 | 96.25 uM | Gln54 Leu56 Lys57 Val58 Pro60 Leu63 | |
| 25 | 3 | −5.35 | 119.73 uM | Met1 Glu2 Asn3 Phe4 Thr6 Tyr231 Phe234 Asp235 Lys313 | |
| Ferulic acid 3 | 16 | 3 | −5.72 | 64.31 uM | Met1 Glu2 Phe4 Thr6 Tyr231 Phe234 Asp235 Lys313 |
| 9 | 4 | −5.38 | 113.66 uM | Met55 Leu56 Lys57 Arg62 Val98 Ser102 Ser103 Pro126 Trp127 | |
| 23 | 5 | −5.14 | 170.90 uM | Gln53 Gln54 Leu56 Lys57 Val58 Pro60 Leu63 | |
| 8 | 3 | −5.13 | 173.60 uM | Met1 Glu2 Asn3 Phe4 Tyr231 Lys232 Phe234 Asp235 Arg306 | |
| 19 | 3 | −4.78 | 313.10 uM | Phe4 Thr6 Tyr231 Phe234 Asp235 Arg306 Lys313 | |
| 9 | 2 | −4.51 | 491.13 uM | Leu300 Cys301 Asp302 Ala303 Tyr305 Arg306 Leu307 His308 Phe329 Ala333 Ala337 | |
| Propyl gallate 4 | 2 | 1 | −5.72 | 64.62 uM | Gln68 Thr69 Gly72 Tyr76 Cys301 Asp302 Gly336 Val338 Pro340 Trp343 |
| 39 | 1 | −5.55 | 85.47 uM | Ala65 Gln68 Thr69 Tyr76 Cys301 Asp302 Ala303 Gly336 Val338 Trp343 | |
| 16 | 2 | −5.29 | 131.61 uM | Cys301 Asp302 Ala303 Arg306 Leu307 His308 Phe329 Ala333 Ala337 | |
| 7 | 1-2 | −5.04 | 201.42 uM | Gly72 Tyr76 Cys301 Asp302 Ala333 Gly336 Ala337 Val338 | |
| 12 | 4 | −4.36 | 632.98 uM | Lys57 Val58 Asp59 Arg62 Ser103 | |
| Octyl gallate 5 | 12 | 2-1 | −6.53 | 16.47 uM | Tyr76 Leu300 Cys301 Asp302 Ala303 Tyr305 Arg306 Leu307 His308 Phe329 Ala333 Gly336 Ala337 Val338 |
| 15 | 1-2 | −6.25 | 26.31 uM | Gly72 Tyr76 Leu300 Cys301 Asp302 Arg306 Leu307 His308 Phe329 Gly336 Ala337 Val338 | |
| 10 | 1 | −6.24 | 26.46 uM | Ala65 Ser66 Gln68 Thr69 Tyr76 Cys301 Asp302 Ala303 Arg304 Gly336 Val338 Trp343 | |
| 9 | 2-1 | −5.65 | 72.30 uM | Tyr76 Leu300 Cys301 Asp302 Ala303 Tyr305 Arg306 Leu307 His308 Phe329 | |
| Carvacrol 6 | 62 | 1 | −5.31 | 127.77 uM | Leu36 Gln68 Thr69 Gly72 Tyr76 Asp302 Gly336 Ala337 Val338 Trp343 |
| 24 | 2 | −5.29 | 132.22 uM | Cys301 Asp302 Ala303 Arg306 Leu307 His308 Phe329 Ala333 | |
| 9 | 4 | −5.18 | 158.87 uM | Met55 Lys57 Arg62 Val98 Leu99 Asp104 Pro126 Trp127 | |
| Thymol 7 | 18 | 4 | −5.51 | 91.27 uM | Met55 Leu56 Lys57 Val58 Val98 Leu99 Asp104 His125 Pro126 Trp127 |
| 72 | 1 | −5.34 | 120.99 uM | Tyr76 Trp298 Cys301 Asp302 Gly336 Val338 Pro340 Trp343 | |
| 6 | 1 | −5.14 | 171.69 uM | Leu36 Gly72 Tyr76 Cys301 Asp302 Ala303 Gly336 Val338 Trp343 | |
| R-Linalool 8 | 11 | 1 | −5.22 | 148.63 uM | Leu36 Ala65 Gln68 Thr69 Gly72 Tyr76 Asp302 Ala303 Trp343 |
| 60 | 1 | −5.13 | 172.51 uM | Gln68 Thr69 Tyr76 Gly336 Ala337 Val338 Pro340 Trp343 | |
| 9 | 4 | −5.06 | 196.45 uM | Met55 Leu56 Lys57 Val98 His125 Pro126 Trp127 | |
| 9 | 1 | −4.92 | 248.45 uM | Leu36 Gly72 Tyr76 Cys301 Asp302 Gly336 Ala337 Val338 Trp343 | |
| S-Linalool 8 | 8 | 4 | −5.30 | 131.37 uM | Met55 Leu56 Lys57 Arg62 Val98 His125 Pro126 Trp127 |
| 55 | 1 | −5.25 | 142.97 uM | Gln68 Thr69 Tyr76 Cys301 Asp302 Gly336 Ala337 Val338 Pro340 Trp343 | |
| 25 | 1 | −5.22 | 148.04 uM | Leu36 Ala65 Gln68 Thr69 Gly72 Tyr76 Cys301 Asp302 Ala303 Trp343 | |
| Geraniol 9 | 13 | 2 | −5.40 | 110.42 uM | Leu300 Cys301 Asp302 Ala303 Tyr305 Arg306 Leu307 His308 Glu309 Phe329 |
| 55 | 1 | −5.31 | 128.06 uM | Gln68 Thr69 Gly72 Tyr76 Cys301 Asp302 Gly336 Ala337 Val338 Pro340 Trp343 | |
| 7 | 1 | −4.99 | 218.54 uM | Gln68 Gly72 Tyr76 Cys301 Asp302 Ala303 Gly336 Trp343 | |
| 12 | 4 | −4.98 | 225.19 uM | Met55 Leu56 Lys57 Arg62 Val98 Ser103 His125 Pro126 Trp127 | |
| Apocynin 10 | 67 | 1 | −5.39 | 111.78 uM | Gln68 Tyr76 Asp302 Gly336 Ala337 Val338 Trp343 |
| 12 | 1 | −5.32 | 125.59 uM | Gln68 Thr69 Gly72 Tyr76 Cys301 Asp302 Gly336 Ala337 Val338 | |
| 11 | 4 | −5.15 | 168.53 uM | Met55 Leu56 Lys57 Arg62 Val98 Ser103 Asp104 Trp127 | |
| Eugenol 11 | 27 | 2 | −5.20 | 153.84 uM | His299 Leu300 Cys301 Asp302 Ala303 Tyr305 Arg306 Leu307 His308 Phe329 Ala333 |
| 23 | 2 | −5.16 | 165.71 uM | His299 Leu300 Cys301 Asp302 Ala303 Arg306 Leu307 His308 Glu309 Phe329 | |
| 32 | 1 | −5.02 | 208.24 uM | Gln68 Thr69 Tyr76 Trp298 Cys301 Asp302 Gly336 Ala337 Val338 Trp343 | |
| 12 | 4 | −4.81 | 296.34 uM | Met55 Leu56 Lys57 Arg62 Val98 Ser102 Asp104 His125 Pro126 Trp127 | |
| Magnolol 12 | 38 | 1-2 | −6.93 | 8.36 uM | Gln68 Thr69 Tyr76 Leu300 Cys301 Asp302 Ala303 Phe329 Ala333 Gly336 Ala337 Val338 Pro340 Trp343 |
| 14 | 1-2 | −6.79 | 10.59 uM | Gln68 Thr69 Gly72 Tyr76 Cys301 Asp302 Ala303 Gly336 Ala337 Val338 Trp343 | |
| 10 | 1-2 | −6.46 | 18.40 uM | Gly72 Tyr76 Trp298 Leu300 Cys301 Asp302 His308 Phe329 Gly336 Ala337 Val338 Pro340 | |
| 17 | 4 | −5.87 | 50.12 uM | Met55 Leu56 Lys57 Arg62 Val98 Ser102 Ser103 His125 Pro126 Trp127 | |
| Honokiol 13 | 23 | 1-2 | −7.25 | 4.89 uM | Gln68 Thr69 Gly72 Trp298 Leu300 Cys301 Asp302 Phe329 Ala337 Val338 Pro340 Trp343 |
| 13 | 2 | −7.14 | 5.87 uM | Leu300 Cys301 Asp302 Ala303 Tyr305 Arg306 Leu307 His308 Tyr311 Phe329 Glu330 Ala333 Ala337 | |
| 23 | 1 | −7.01 | 7.31 uM | Leu36 Ala65 Gln68 Thr69 Gly72 Tyr76 Cys301 Asp302 Ala303 Gly336 Ala337 Val338 Trp343 | |
| 15 | 1-2 | −6.93 | 8.36 uM | Gln68 Thr69 Gly72 Tyr76 Trp298 Cys301 Asp302 His308 Phe329 Ala337 Val338 | |
| Eugenol dimer 14 | 35 | 1-2 | −6.99 | 7.58 uM | Gln68 Gly72 Tyr76 Trp298 Cys301 Asp302 Ala303 His308 Ala337 Val338 Trp343 |
| 19 | 1-2 | −6.85 | 9.55 uM | Thr69 Gly72 Tyr76 Trp298 Leu300 Cys301 Asp302 Ala303 Phe329 Ala333 Ala337 Val338 Ala339 Pro340 Trp343 | |
| 11 | 4 | −5.37 | 116.28 uM | Met55 Leu56 Lys57 Val58 Arg62 Val98 Ser102 Ser103 Pro126 Trp127 | |
| Ferulic acid dimer 15 | 8 | 1-2 | −5.90 | 47.65 uM | Leu36 Ala65 Gln68 Thr69 Gly72 Tyr76 Cys301 Asp302 Ala303 Arg306 His308 Ala337 Val338 Trp343 |
| 25 | 1-2 | −5.37 | 115.39 uM | Gln68 Thr69 Tyr76 Trp298 Leu300 Cys301 Asp302 Ala303 Phe329 Ala333 Gly336 Ala337 Val338 Ala339 Pro340 | |
| 10 | 4-5 | −4.98 | 221.88 uM | Gln53 Gln54 Leu56 Lys57 Val58 Asp59 Arg62 Leu63 | |
| NPD352 | 36 | 1-2 | −8.85 | 327.93 nM | Gln68 Gly72 Tyr76 Leu300 Cys301 Asp302 Ala303 His308 Phe329 Ala333 Ala337 Val338 Trp343 |
| Treatment | a Dry Fungal Biomass (Yield (mg) ± SE) | b DON (Yield (ng/mL) ± SE) | |
|---|---|---|---|
| DON | 3-ADON | ||
| Control | 50.36 ± 1.00 | 19.96 ± 2.30 | 80.26 ± 9.80 |
| p-Coumaric acid 1 | 50.14 ± 0.64 | 53.52 ± 9.94 * | 285.11 ± 32.79 * |
| Caffeic acid 2 | 48.56 ± 1.98 | 21.58 ± 1.32 | 66.85 ± 8.09 |
| Ferulic acid 3 | 44.20 ± 2.08 | 0.73 ± 0.31 | 1.35 ± 0.56 * |
| Propyl gallate 4 | 20.60 ± 1.07 * | N.D. | 86.20 ± 4.89 |
| Octyl gallate 5 | 0 * | N.D. | N.D. |
| Carvacrol 6 | 52.08 ± 2.13 | 0.34 ± 0.22 | 2.11 ± 0.86 * |
| Thymol 7 | 25.80 ± 4.50 * | N.D. | N.D. |
| Linalool 8 | 45.52 ± 0.82 | 33.95 ± 1.77 | 81.68 ± 5.68 |
| Geraniol 9 | 44.82 ± 0.95 | 18.45 ± 2.86 | 56.26 ± 9.89 |
| Apocynin 10 | 0 * | N.D. | N.D. |
| Eugenol 11 | 40.98 ± 1.91 * | N.D. | 2.58 ± 1.68 * |
| Magnolol 12 | 0 * | N.D. | N.D. |
| Honokiol 13 | 0 * | N.D. | N.D. |
| Eugenol dimer 14 | 44.12 ± 1.44 | 119.87 ± 12.51 * | 583.15 ± 51.33 * |
| Ferulic acid dimer 15 | 49.74 ± 1.06 | 14.33 ± 1.22 | 39.94 ± 3.14 |
| Propyl gallate 4 + thymol 7 | 28.20 ± 0.96 * | N.D. | N.D. |
| Magnolol 12 + thymol 7 | 0 * | N.D. | N.D. |
| Honokiol 13 + thymol 7 | 0 * | N.D. | N.D. |
| Treatment | a Disease Index | b Grain Yield (g) | c DON (Relative Yield ± SE) | |
|---|---|---|---|---|
| DON | 3-ADON | |||
| Control | 63.86 ± 9.50 a | 12.54 ± 1.46 a | 4528.47 ± 612.45 a | 79.67 ± 13.35 a |
| Propyl gallate 4 | 37.76 ± 10.49 a,b | 14.41 ± 1.55 a | 1308.49 ± 207.75 c,d | 16.72 ± 1.97 b,c |
| Octyl gallate 5 | 13.34 ± 4.51 b | 17.56 ± 0.94 a | 53.985 ± 37.46 d | N.D. |
| Thymol 7 | 30.0 ± 10.46 a,b | 15.40 ± 1.09 a | 3079.80 ± 546.96 a,b | 51.65 ± 19.63 a,b |
| Magnolol 12 | 19.98 ± 8.68 a,b | 15.64 ± 0.44 a | 1914.40 ± 181.32 b,c | 15.94 ± 5.46 b,c |
| Honokiol 13 | 21.10 ± 6.72 a,b | 17.32 ± 0.67 a | 207.34 ± 51.72 d | 4.01 ± 1.31 c |
| Propyl gallate 4 + thymol 7 | 32.20 ± 9.85 a,b | 15.74 ± 1.42 a | 19.14 ± 9.74 d | 4.10 ± 1.98 c |
| Magnolol 12 + thymol 7 | 36.64 ± 13.89 a,b | 15.74 ± 0.97 a | 3680.76 ± 536.69 a | 60.16 ± 10.93 a,b |
| Tebuconazole | 1.10 ± 1.10 | 17.08 ± 0.48 | 10.06 ± 1.33 | N.D. |
| Analyte | LOD (ng mL−1) | LOQ (ng mL−1) |
|---|---|---|
| DON | 0.05 | 0.7 |
| 3-ADON | 0.05 | 1.5 |
| 15-ADON | 0.05 | 1.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oufensou, S.; Dessì, A.; Dallocchio, R.; Balmas, V.; Azara, E.; Carta, P.; Migheli, Q.; Delogu, G. Molecular Docking and Comparative Inhibitory Efficacy of Naturally Occurring Compounds on Vegetative Growth and Deoxynivalenol Biosynthesis in Fusarium culmorum. Toxins 2021, 13, 759. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13110759
Oufensou S, Dessì A, Dallocchio R, Balmas V, Azara E, Carta P, Migheli Q, Delogu G. Molecular Docking and Comparative Inhibitory Efficacy of Naturally Occurring Compounds on Vegetative Growth and Deoxynivalenol Biosynthesis in Fusarium culmorum. Toxins. 2021; 13(11):759. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13110759
Chicago/Turabian StyleOufensou, Safa, Alessandro Dessì, Roberto Dallocchio, Virgilio Balmas, Emanuela Azara, Paola Carta, Quirico Migheli, and Giovanna Delogu. 2021. "Molecular Docking and Comparative Inhibitory Efficacy of Naturally Occurring Compounds on Vegetative Growth and Deoxynivalenol Biosynthesis in Fusarium culmorum" Toxins 13, no. 11: 759. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13110759