Pharmaceutical Assessment Suggests Locomotion Hyperactivity in Zebrafish Triggered by Arecoline Might Be Associated with Multiple Muscarinic Acetylcholine Receptors Activation

 , ,
, , 
Abstract
:1. Introduction
2. Results
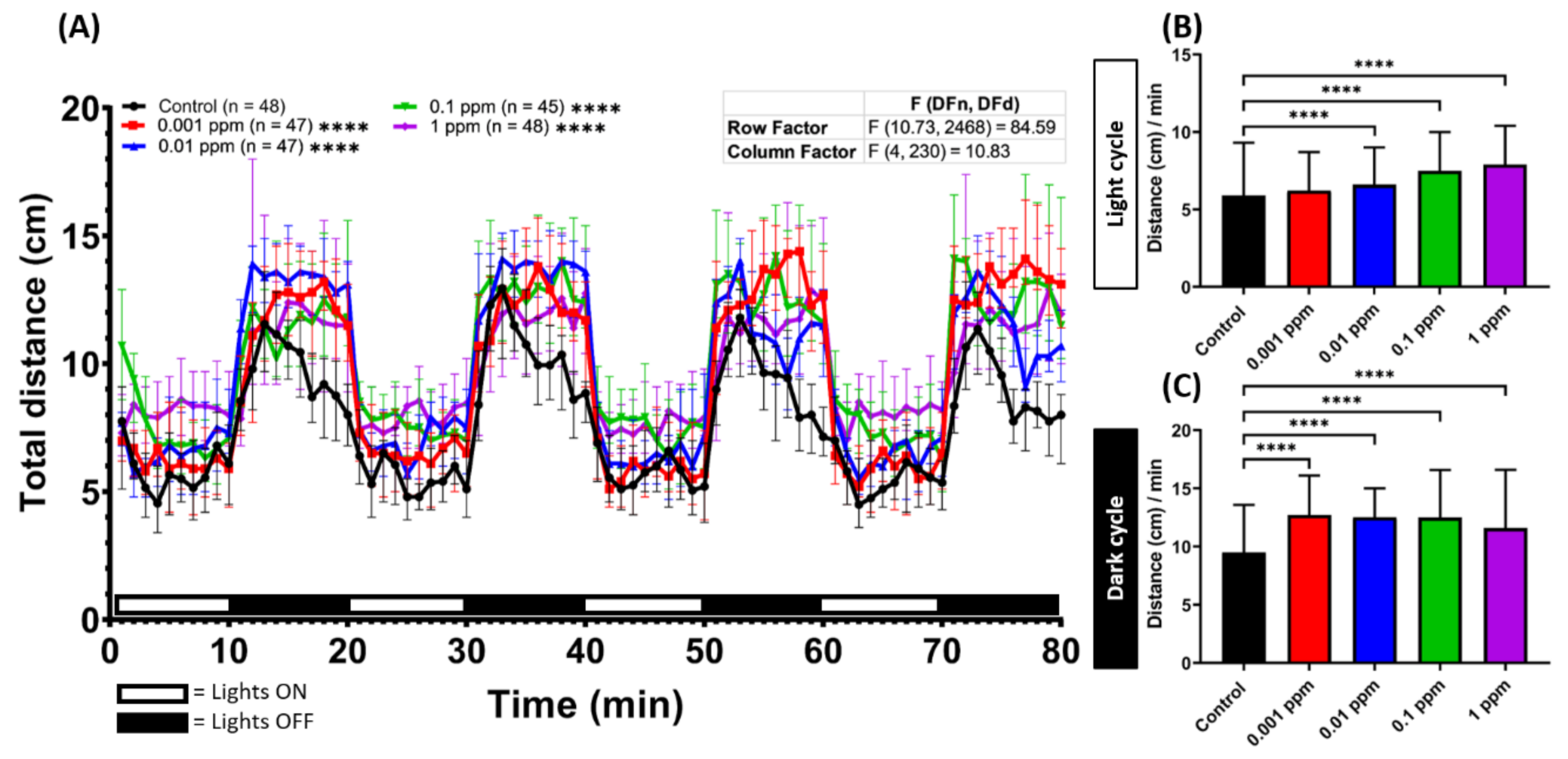
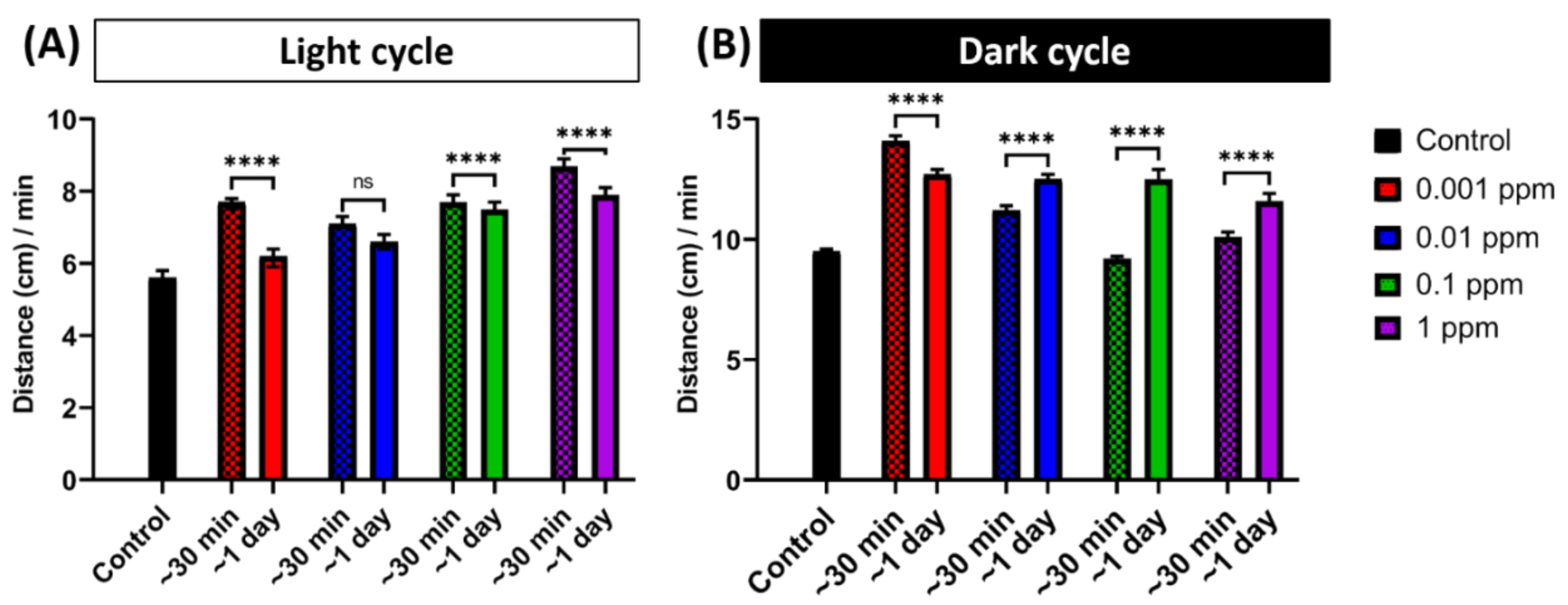
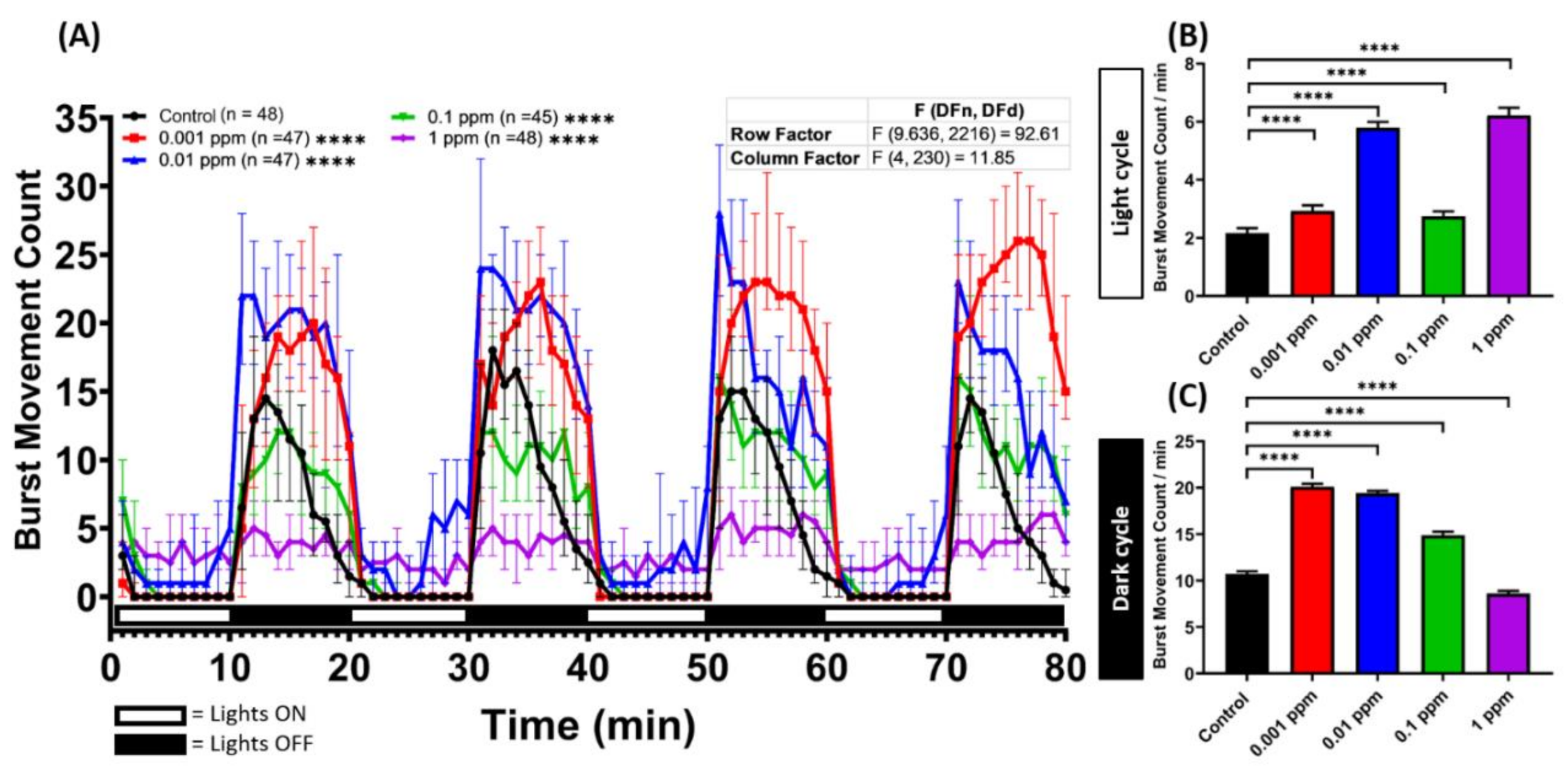
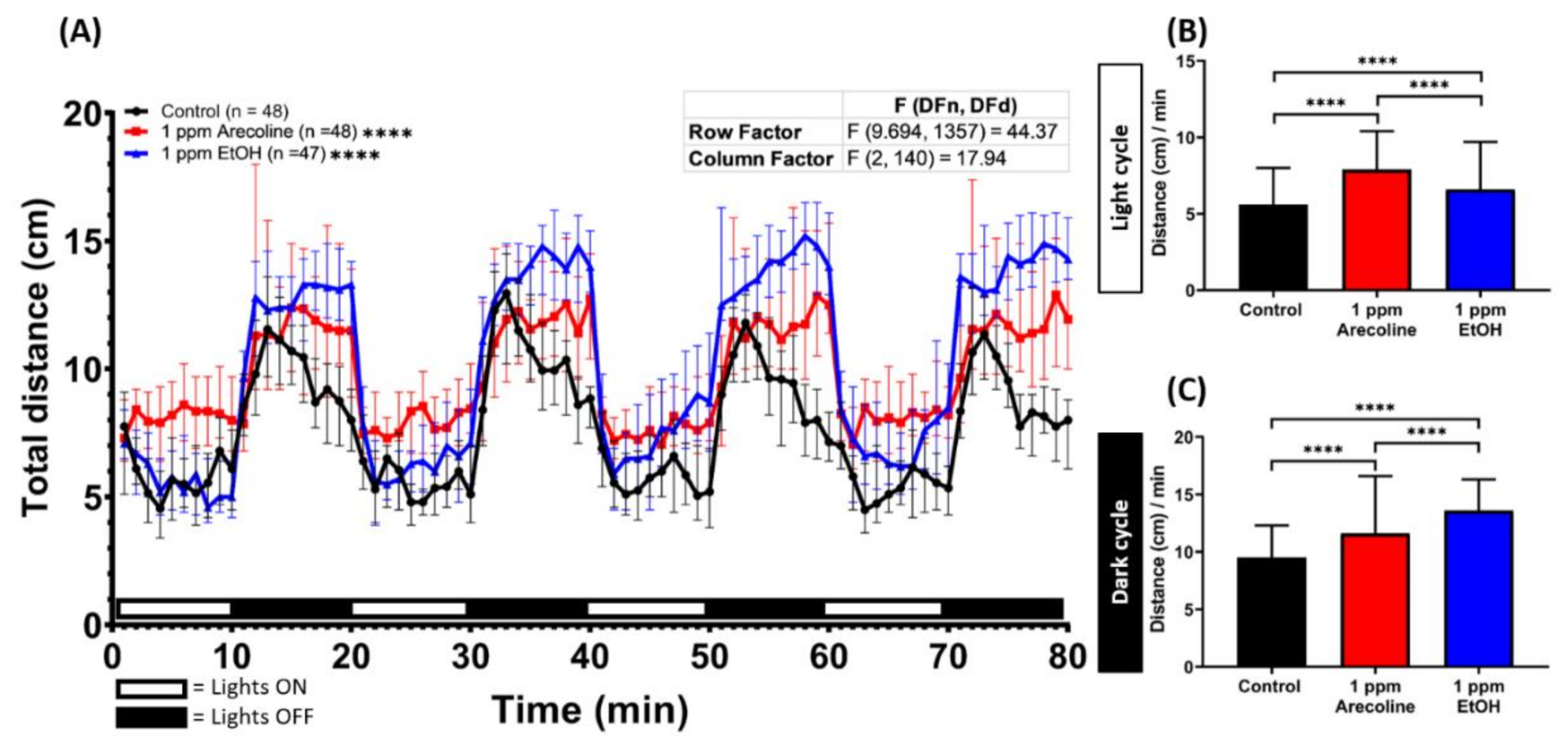
2.1. Low Dose Arecoline Treatment Elevates Locomotor Activity in Zebrafish Larvae
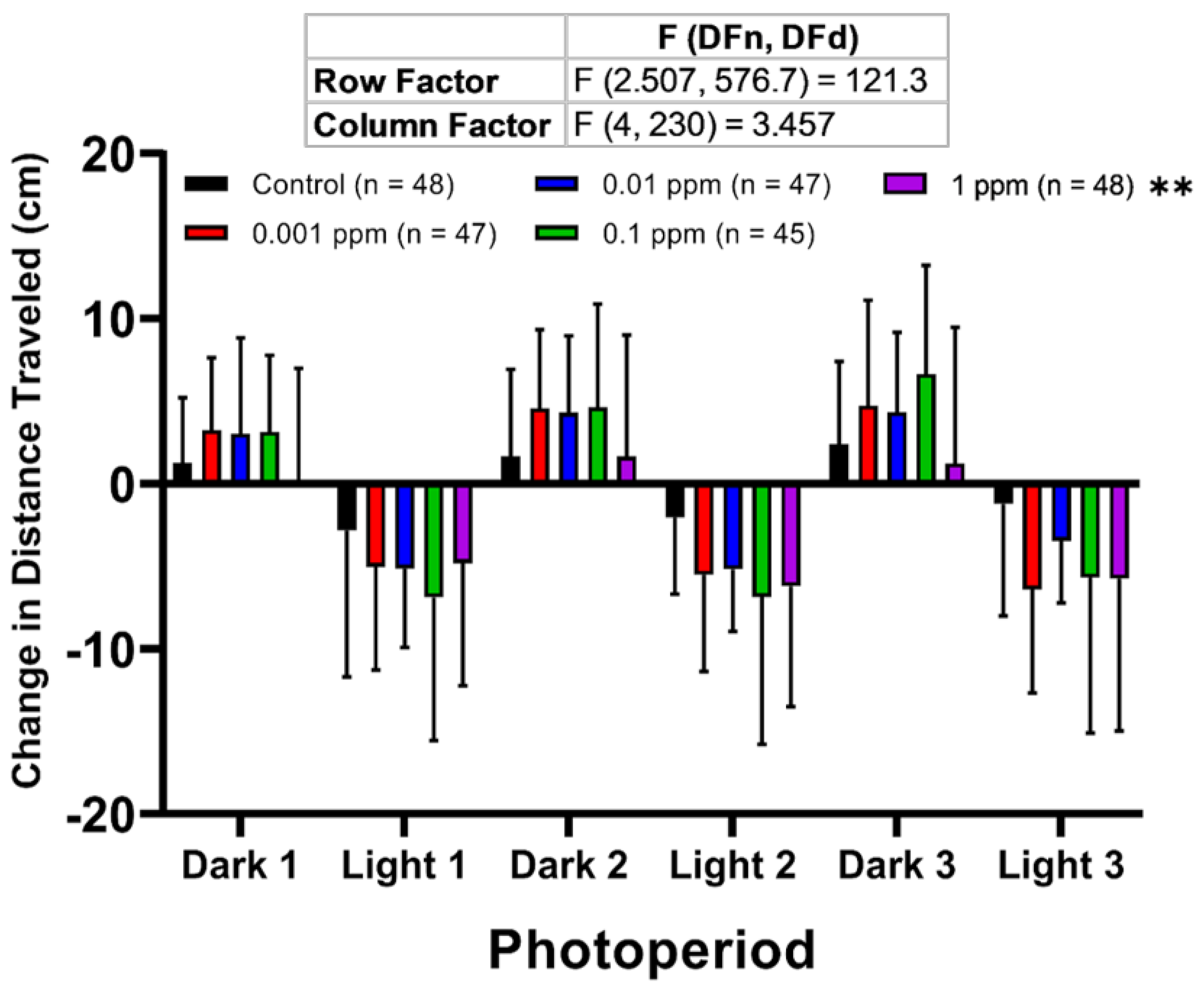
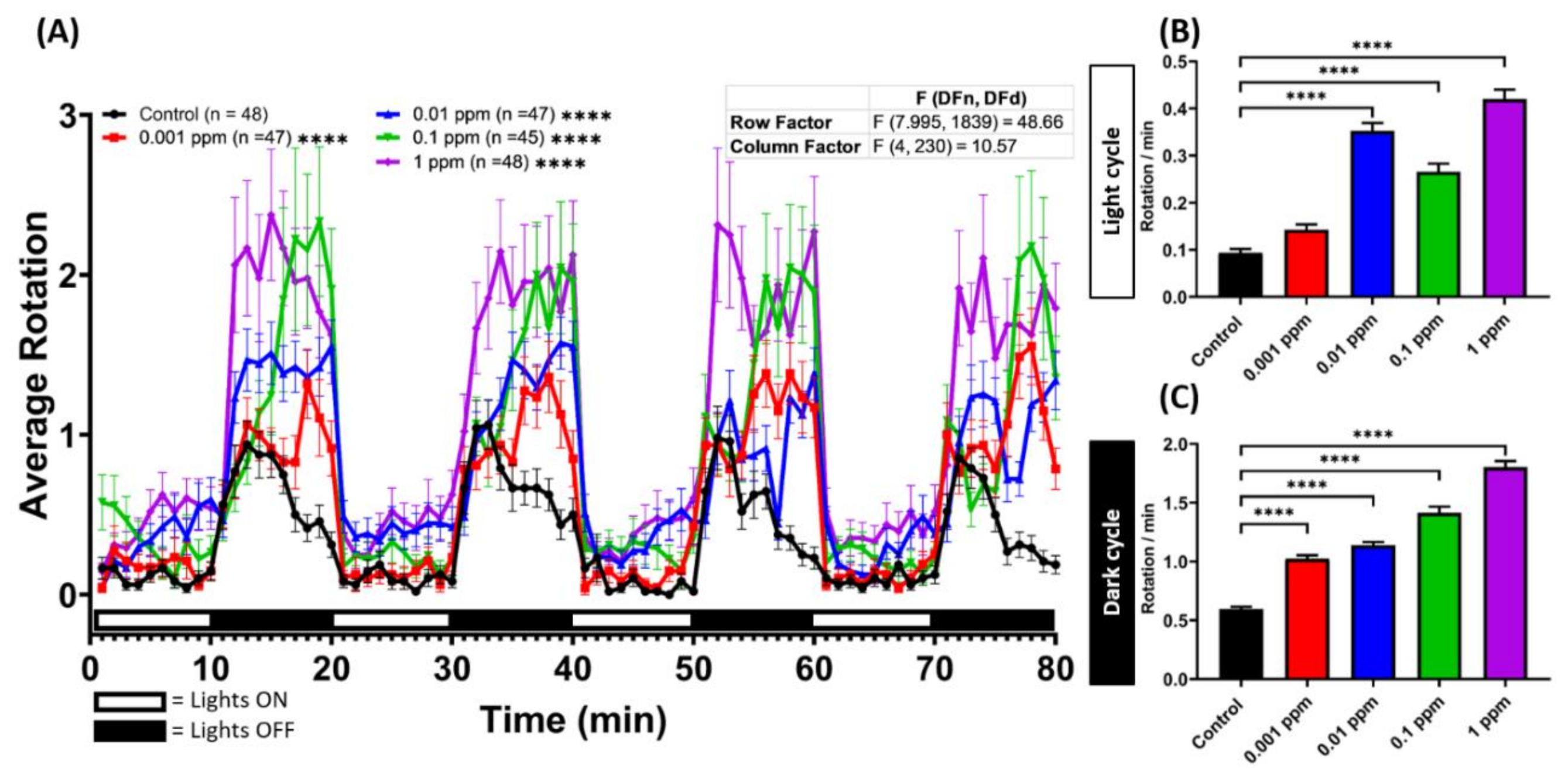
2.2. Low Dose Arecoline Treatment Altered Larvae Photomotor Response (LPMR) in Zebrafish Larvae
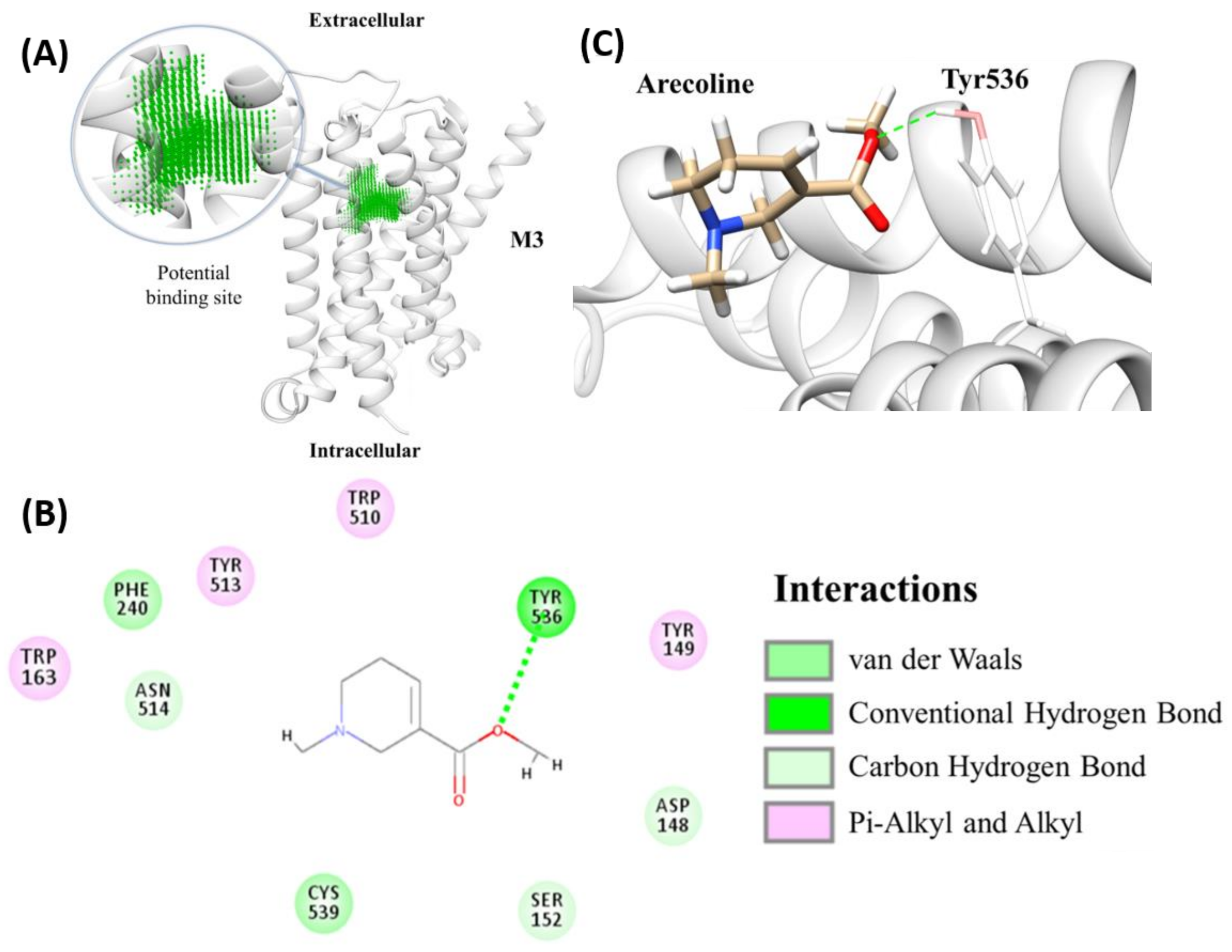
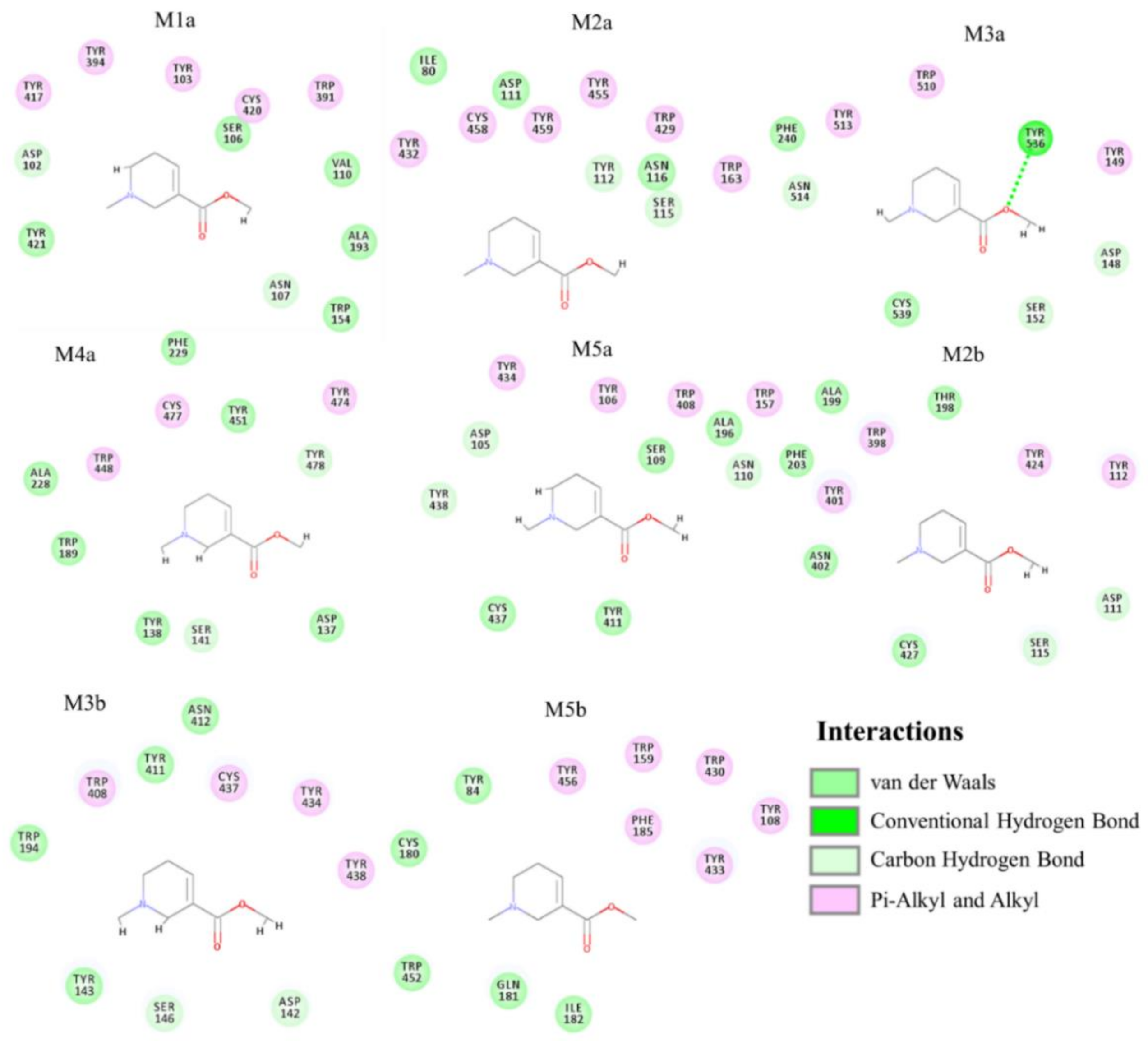
2.3. Molecular Docking for Arecoline and Muscarinic Acetylcholine Receptor
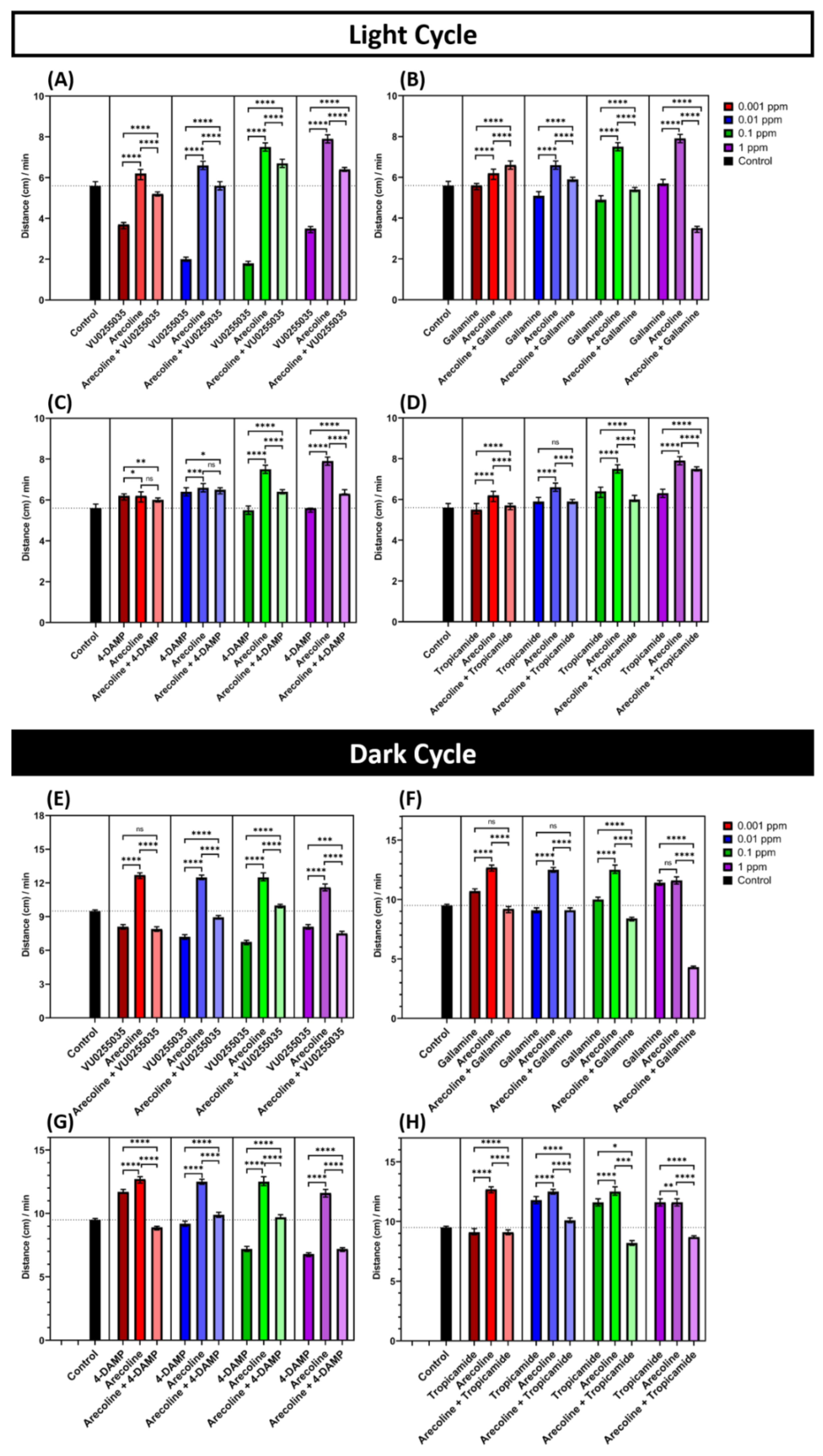
2.4. Muscarinic Receptor Antagonist Suppressed Locomotion Hyperactivity Induced by Arecoline
3. Discussion
4. Materials and Methods
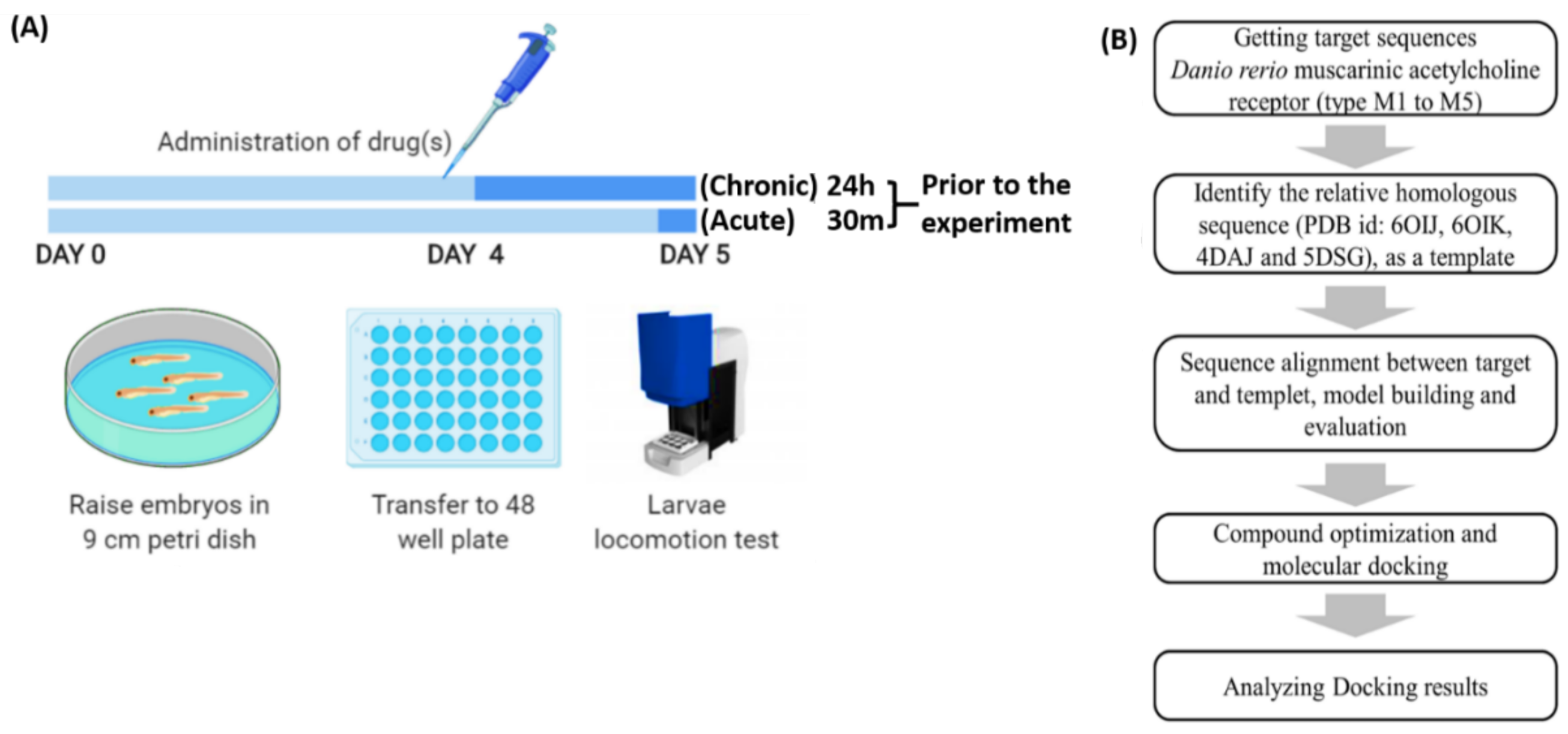
4.1. Zebrafish Locomotion Assay to Evaluate Arecoline Bioactivity Workflow Overview
4.2. Zebrafish Maintenance
4.3. Arecoline and Muscarinic Acetylcholine Receptor Antagonist Treatment
4.4. Larvae Locomotion Tracking
4.5. Structure-Based Molecular Simulation for Arecoline and Muscarinic Acetylcholine Receptor Binding
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A





References
- Chu, N.-S. Effects of betel chewing on the central and autonomic nervous systems. J. Biomed. Sci. 2001, 8, 229–236. [Google Scholar] [CrossRef]
- Williams, S.; Malik, A.; Chowdhury, S.; Chauhan, S. Sociocultural aspects of areca nut use. Addict. Biol. 2002, 7, 147–154. [Google Scholar] [CrossRef]
- Peng, W.-H.; Lee, Y.-C.; Chau, Y.-P.; Lu, K.-S.; Kung, H.-N. Short-term exposure of zebrafish embryos to arecoline leads to retarded growth, motor impairment, and somite muscle fiber changes. Zebrafish 2015, 12, 58–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lord, G.; Lim, C.; Warnakulasuriya, S.; Peters, T. Chemical and analytical aspects of areca nut. Addict. Biol. 2002, 7, 99–102. [Google Scholar] [CrossRef]
- Arjungi, K. Areca nut: A review. Arzneim. Forsch. 1976, 26, 951–956. [Google Scholar]
- Tsopelas, N.D.; Marin, D.B. Cholinergic Treatments of Alzheimer’s Disease. In Functional Neurobiology of Aging; Elsevier: Amsterdam, The Netherlands, 2001; pp. 475–486. [Google Scholar]
- Euler, U.V.; Domeij, B. Nicotine-like actions of arecoline. Acta Pharmacol. Toxicol. 1945, 1, 263–269. [Google Scholar] [CrossRef]
- Paul, K.; Moitra, P.; Mukherjee, I.; Maity, C.; Ghosal, S. Teratogenicity of arecoline hydrobromide on developing chick embryos: A preliminary report. Bull. Environ. Contam. Toxicol. 1999, 62, 356–362. [Google Scholar] [CrossRef]
- Sinha, A.; Rao, A.R. Embryotoxicity of betel nuts in mice. Toxicology 1985, 37, 315–326. [Google Scholar] [CrossRef]
- Chang, B.E.; Liao, M.H.; Kuo, M.Y.P.; Chen, C.H. Developmental toxicity of arecoline, the major alkaloid in betel nuts, in zebrafish embryos. Birth Defects Res. Part A Clin. Mol. Teratol. 2004, 70, 28–36. [Google Scholar] [CrossRef]
- Asthana, S.; Raffaele, K.C.; Greig, N.H.; Berardi, A.; Morris, P.P.; Schapiro, M.B.; Rapoport, S.I.; Blackman, M.R.; Soncrant, T.T. Neuroendocrine responses to intravenous infusion of arecoline in patients with Alzheimer’s disease. Psychoneuroendocrinology 1995, 20, 623–636. [Google Scholar] [CrossRef]
- Dasgupta, R.; Chatterji, U.; Nag, T.; Chaudhuri-Sengupta, S.; Nag, D.; Maiti, B. Ultrastructural and hormonal modulations of the thyroid gland following arecoline treatment in albino mice. Mol. Cell. Endocrinol. 2010, 319, 1–7. [Google Scholar] [CrossRef]
- Eddins, D.; Cerutti, D.; Williams, P.; Linney, E.; Levin, E.D. Zebrafish provide a sensitive model of persisting neurobehavioral effects of developmental chlorpyrifos exposure: Comparison with nicotine and pilocarpine effects and relationship to dopamine deficits. Neurotoxicol. Teratol. 2010, 32, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Selderslaghs, I.W.; Hooyberghs, J.; De Coen, W.; Witters, H.E. Locomotor activity in zebrafish embryos: A new method to assess developmental neurotoxicity. Neurotoxicol. Teratol. 2010, 32, 460–471. [Google Scholar] [CrossRef] [PubMed]
- Echevarria, D.J.; Hammack, C.M.; Pratt, D.W.; Hosemann, J.D. A novel behavioral test battery to assess global drug effects using the zebrafish. Int. J. Comp. Psychol. 2008, 21, 19–34. [Google Scholar]
- Papke, R.L.; Horenstein, N.A.; Stokes, C. Nicotinic activity of arecoline, the psychoactive element of “Betel Nuts”, suggests a basis for habitual use and anti-inflammatory activity. PLoS ONE 2015, 10, e0140907. [Google Scholar] [CrossRef] [PubMed]
- Javed, F.; Bello Correra, F.O.; Chotai, M.; Tappuni, A.R.; Almas, K. Systemic conditions associated with areca nut usage: A literature review. Scand. J. Public Health 2010, 38, 838–844. [Google Scholar] [CrossRef] [PubMed]
- Chu, N.S. Neurological aspects of areca and betel chewing. Addict. Biol. 2002, 7, 111–114. [Google Scholar] [CrossRef]
- Klee, E.W.; Ebbert, J.O.; Schneider, H.; Hurt, R.D.; Ekker, S.C. Zebrafish for the study of the biological effects of nicotine. Nicotine Tob. Res. 2011, 13, 301–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levin, E.D.; Bencan, Z.; Cerutti, D.T. Anxiolytic effects of nicotine in zebrafish. Physiol. Behav. 2007, 90, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Serikuly, N.; Alpyshov, E.T.; Wang, D.; Wang, J.; Yang, L.; Hu, G.; Yan, D.; Demin, K.A.; Kolesnikova, T.O.; Galstyan, D. Effects of acute and chronic arecoline in adult zebrafish: Anxiolytic-like activity, elevated brain monoamines and the potential role of microglia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2020, 104, 109977. [Google Scholar]
- Richendrfer, H.; Pelkowski, S.; Colwill, R.; Creton, R. On the edge: Pharmacological evidence for anxiety-related behavior in zebrafish larvae. Behav. Brain Res. 2012, 228, 99–106. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Lin, K.; Miao, Y.; Dong, Q.; Huang, C.; Wang, H.; Guo, M.; Cui, X. Toxicity assessment of zebrafish following exposure to CdTe QDs. J. Hazard. Mater. 2012, 213, 413–420. [Google Scholar] [CrossRef]
- Ulhaq, M.; Örn, S.; Carlsson, G.; Morrison, D.A.; Norrgren, L. Locomotor behavior in zebrafish (Danio rerio) larvae exposed to perfluoroalkyl acids. Aquat. Toxicol. 2013, 144, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Calogero, A.E.; Kamilaris, T.C.; GOMEZ, M.T.; Johnson, E.O.; Tartaglia, M.E.; Gold, P.W.; Chrousos, G.P. The muscarinic cholinergic agonist arecoline stimulates the rat hypothalamic-pituitary-adrenal axis through a centrally-mediated corticotropin-releasing hormone-dependent mechanism. Endocrinology 1989, 125, 2445–2453. [Google Scholar] [CrossRef] [PubMed]
- Ghelardini, C.; Galeotti, N.; Lelli, C.; Bartolini, A. M1 receptor activation is a requirement for arecoline analgesia. II Farmaco 2001, 56, 383–385. [Google Scholar] [CrossRef]
- Nakatani, Y.; Takeda, H.; Kohara, Y.; Morishita, S. Reconstruction of the vertebrate ancestral genome reveals dynamic genome reorganization in early vertebrates. Genome Res. 2007, 17, 1254–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaillon, O.; Aury, J.-M.; Brunet, F.; Petit, J.-L.; Stange-Thomann, N.; Mauceli, E.; Bouneau, L.; Fischer, C.; Ozouf-Costaz, C.; Bernot, A. Genome duplication in the teleost fish Tetraodon nigroviridis reveals the early vertebrate proto-karyotype. Nature 2004, 431, 946–957. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, J.E.; Bergqvist, C.A.; Larhammar, D. Evolution of the muscarinic acetylcholine receptors in vertebrates. eNeuro 2018, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Pei, J.; Lai, L. Binding site detection and druggability prediction of protein targets for structure-based drug design. Curr. Pharm. Des. 2013, 19, 2326–2333. [Google Scholar] [CrossRef]
- Brooks, B.R.; Bruccoleri, R.E.; Olafson, B.D.; States, D.J.; Swaminathan, S.A.; Karplus, M. CHARMM: A program for macromolecular energy, minimization, and dynamics calculations. J. Comput. Chem. 1983, 4, 187–217. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- MacPhail, R.; Brooks, J.; Hunter, D.; Padnos, B.; Irons, T.; Padilla, S. Locomotion in larval zebrafish: Influence of time of day, lighting and ethanol. Neurotoxicology 2009, 30, 52–58. [Google Scholar] [CrossRef]
- Mathur, P.; Guo, S. Differences of acute versus chronic ethanol exposure on anxiety-like behavioral responses in zebrafish. Behav. Brain Res. 2011, 219, 234–239. [Google Scholar] [CrossRef] [Green Version]
- Van Woudenberg, A.B.; Wolterbeek, A.; te Brake, L.; Snel, C.; Menke, A.; Rubingh, C.; de Groot, D.; Kroese, D. A category approach to predicting the developmental (neuro) toxicity of organotin compounds: The value of the zebrafish (Danio rerio) embryotoxicity test (ZET). Reprod. Toxicol. 2013, 41, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Hill, B.N.; Saari, G.N.; Steele, W.B.; Corrales, J.; Brooks, B.W. Nutrients and salinity influence Prymnesium parvum (UTEX LB 2797) elicited sublethal toxicity in Pimephales promelas and Danio rerio. Harmful Algae 2020, 93, 101795. [Google Scholar] [CrossRef]
- Steele, W.B.; Mole, R.A.; Brooks, B.W. Experimental protocol for examining behavioral response profiles in larval fish: Application to the neuro-stimulant caffeine. JoVE J. Vis. Exp. 2018, 137, e57938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kristofco, L.A.; Cruz, L.C.; Haddad, S.P.; Behra, M.L.; Chambliss, C.K.; Brooks, B.W. Age matters: Developmental stage of Danio rerio larvae influences photomotor response thresholds to diazinion or diphenhydramine. Aquat. Toxicol. 2016, 170, 344–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kokel, D.; Bryan, J.; Laggner, C.; White, R.; Cheung, C.Y.J.; Mateus, R.; Healey, D.; Kim, S.; Werdich, A.A.; Haggarty, S.J. Rapid behavior-based identification of neuroactive small molecules in the zebrafish. Nat. Chem. Biol. 2010, 6, 231–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araujo, D.M.; Lapchak, P.A.; Quirion, R. Heterogeneous binding of [3H] 4-DAMP to muscarinic cholinergic sites in the rat brain: Evidence from membrane binding and autoradiographic studies. Synapse 1991, 9, 165–176. [Google Scholar] [CrossRef]
- Liu, Y.-J.; Peng, W.; Hu, M.-B.; Xu, M.; Wu, C.-J. The pharmacology, toxicology and potential applications of arecoline: A review. Pharm. Biol. 2016, 54, 2753–2760. [Google Scholar] [CrossRef]
- Sun, Y.P.; Liu, Q.; Luo, J.; Guo, P.; Chen, F.; Lawrence, A.J.; Liang, J.H. Systemic Administration of Arecoline Reduces Ethanol-Induced Sleeping Through Activation of Central Muscarinic Receptor in Mice. Alcohol. Clin. Exp. Res. 2010, 34, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Retz, K.C.; Trimmer, C.K.; Forster, M.J.; Lal, H. Motor responses of autoimmune NZB/B1NJ and C57BL/6NNia mice to arecoline and nicotine. Pharmacol. Biochem. Behav. 1987, 28, 275–282. [Google Scholar] [CrossRef]
- Pradhan, S.; Dutta, S. Behavioral effects of arecoline in rats. Psychopharmacologia 1970, 17, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K.; Holloway, H.H.; Larson, D.M.; Soncrant, T.T. Effect of acute and chronic arecoline treatment on cerebral metabolism and blood flow in the conscious rat. Brain Res. 1994, 641, 65–75. [Google Scholar] [CrossRef]
- Hodgkin, A.L.; Huxley, A.F. A quantitative description of membrane current and its application to conduction and excitation in nerve. J. Physiol. 1952, 117, 500–544. [Google Scholar] [CrossRef]
- Saint-Amant, L.; Drapeau, P. Time course of the development of motor behaviors in the zebrafish embryo. J. Neurobiol. 1998, 37, 622–632. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Patterson, J.; Kimmel, R.O. The development and behavioral characteristics of the startle response in the zebra fish. Dev. Psychobiol. 1974, 7, 47–60. [Google Scholar] [CrossRef]
- Mickey, J.; Tate, R.; Lefkowitz, R. Subsensitivity of adenylate cyclase and decreased beta-adrenergic receptor binding after chronic exposure to (minus)-isoproterenol in vitro. J. Biol. Chem. 1975, 250, 5727–5729. [Google Scholar] [CrossRef]
- Morrow, A.L.; Montpied, P.; Lingford-Hughes, A.; Paul, S.M. Chronic ethanol and pentobarbital administration in the rat: Effects on GABAA receptor function and expression in brain. Alcohol 1990, 7, 237–244. [Google Scholar] [CrossRef]
- González, S.; Fernández-Ruiz, J.; Sparpaglione, V.; Parolaro, D.; Ramos, J.A. Chronic exposure to morphine, cocaine or ethanol in rats produced different effects in brain cannabinoid CB1 receptor binding and mRNA levels. Drug Alcohol Depend. 2002, 66, 77–84. [Google Scholar] [CrossRef]
- Molinengo, L.; Cassone, M.C.; Orsetti, M. Action of arecoline on the levels of acetylcholine, norepinephrine and dopamine in the mouse central nervous system. Pharmacol. Biochem. Behav. 1986, 24, 1801–1803. [Google Scholar] [CrossRef]
- Gerlai, R.; Lee, V.; Blaser, R. Effects of acute and chronic ethanol exposure on the behavior of adult zebrafish (Danio rerio). Pharmacol. Biochem. Behav. 2006, 85, 752–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerlai, R.; Chatterjee, D.; Pereira, T.; Sawashima, T.; Krishnannair, R. Acute and chronic alcohol dose: Population differences in behavior and neurochemistry of zebrafish. Genes Brain Behav. 2009, 8, 586–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sackerman, J.; Donegan, J.J.; Cunningham, C.S.; Nguyen, N.N.; Lawless, K.; Long, A.; Benno, R.H.; Gould, G.G. Zebrafish behavior in novel environments: Effects of acute exposure to anxiolytic compounds and choice of Danio rerio line. Int. J. Comp. Psychol. 2010, 23, 43. [Google Scholar]
- Tran, S.; Gerlai, R. Time-course of behavioural changes induced by ethanol in zebrafish (Danio rerio). Behav. Brain Res. 2013, 252, 204–213. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Busillo, J.M.; Benovic, J.L. M3 muscarinic acetylcholine receptor-mediated signaling is regulated by distinct mechanisms. Mol. Pharmacol. 2008, 74, 338–347. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, D.J.Y.; Liao, C.F. Zebrafish M2 muscarinic acetylcholine receptor: Cloning, pharmacological characterization, expression patterns and roles in embryonic bradycardia. Br. J. Pharmacol. 2002, 137, 782–792. [Google Scholar] [CrossRef] [Green Version]
- Johnston, G.; Krogsgaard-Larsen, P.; Stephanson, A. Betel nut constituents as inhibitors of γ-aminobutyric acid uptake. Nature 1975, 258, 627. [Google Scholar] [CrossRef]
- Seifert, R.; Schirmer, B. A simple mechanistic terminology of psychoactive drugs: A proposal. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2020, 393, 1331–1339. [Google Scholar] [CrossRef]
- Xie, D.; Chen, L.; Liu, C.; Zhang, C.; Liu, K.; Wang, P.S. Arecoline Excites the Colonic Smooth Muscle Motility via M~ 3 Receptor in Rabbits. Chin. J. Physiol. 2004, 47, 89. [Google Scholar]
- Chandra, J.N.S.; Malviya, M.; Sadashiva, C.; Subhash, M.; Rangappa, K.S. Effect of novel arecoline thiazolidinones as muscarinic receptor 1 agonist in Alzheimer’s dementia models. Neurochem. Int. 2008, 52, 376–383. [Google Scholar] [CrossRef] [PubMed]
- Messer, W.S., Jr.; Ellerbrock, B.; Price, M.; Hoss, W. Autoradiographic analyses of agonist binding to muscarinic receptor subtypes. Biochem. Pharmacol. 1989, 38, 837–850. [Google Scholar] [CrossRef]
- Soncrant, T.T.; Holloway, H.W.; Greig, N.H.; Rapoport, S.I. Regional brain metabolic responsivity to the muscarinic cholinergic agonist arecoline is similar in young and aged Fischer-344 rats. Brain Res. 1989, 487, 255–266. [Google Scholar] [CrossRef]
- Han, R.; Sun, Y.; Li, J.; Liang, J. Effects of arecoline on morphine-induced behavioral sensitization in mice. Chin. J. Drug Depend. 2005, 14, 197. [Google Scholar]
- Sheffler, D.J.; Williams, R.; Bridges, T.M.; Xiang, Z.; Kane, A.S.; Byun, N.E.; Jadhav, S.; Mock, M.M.; Zheng, F.; Lewis, L.M. A novel selective muscarinic acetylcholine receptor subtype 1 antagonist reduces seizures without impairing hippocampus-dependent learning. Mol. Pharmacol. 2009, 76, 356–368. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.H.; El-Fakahany, E.E. Mixed Competitive and Allosteric Antagonism by Gallamine of Muscarinic Receptor-Mediated Second Messenger Responses in N1E-115 Neuroblastoma Cells. J. Neurochem. 1989, 53, 1300–1308. [Google Scholar] [CrossRef] [PubMed]
- Ehlert, F.J. The interaction of 4-DAMP mustard with subtypes of the muscarinic receptor. Life Sci. 1996, 58, 1971–1978. [Google Scholar] [CrossRef]
- Dei, S.; Bellucci, C.; Ghelardini, C.; Romanelli, M.N.; Spampinato, S. Synthesis, characterization and pharmacological profile of tropicamide enantiomers. Life Sci. 1996, 58, 2147–2153. [Google Scholar] [CrossRef]
- McKinney, M.; Miller, J.; Gibson, V.; Nickelson, L.; Aksoy, S. Interactions of agonists with M2 and M4 muscarinic receptor subtypes mediating cyclic AMP inhibition. Mol. Pharmacol. 1991, 40, 1014–1022. [Google Scholar]
- Felder, C.C. Muscarinic acetylcholine receptors: Signal transduction through multiple effectors. FASEB J. 1995, 9, 619–625. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, S.E.; Loose, M.D.; Qi, M.; Levey, A.I.; Hille, B.; McKnight, G.S.; Idzerda, R.L.; Nathanson, N.M. Disruption of the m1 receptor gene ablates muscarinic receptor-dependent M current regulation and seizure activity in mice. Proc. Natl. Acad. Sci. USA 1997, 94, 13311–13316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pisani, A.; Bernardi, G.; Ding, J.; Surmeier, D.J. Re-emergence of striatal cholinergic interneurons in movement disorders. Trends Neurosci. 2007, 30, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Riker, W.F., Jr.; Wescoe, W.C. The pharmacology of Flaxedil, with observations on certain analogs. Ann. N. Y. Acad. Sci. 1951, 54, 373–394. [Google Scholar] [CrossRef] [PubMed]
- Brown, B.R.; Crout, J.R. The sympathomimetic effect of gallamine on the heart. J. Pharmacol. Exp. Ther. 1970, 172, 266–273. [Google Scholar] [PubMed]
- Jakubík, J.; Bačáková, L.; El-Fakahany, E.E.; Tuček, S. Constitutive activity of the M1–M4 subtypes of muscarinic receptors in transfected CHO cells and of muscarinic receptors in the heart cells revealed by negative antagonists. FEBS Lett. 1995, 377, 275–279. [Google Scholar] [CrossRef] [Green Version]
- De Amici, M.; Dallanoce, C.; Holzgrabe, U.; Tränkle, C.; Mohr, K. Allosteric ligands for G protein-coupled receptors: A novel strategy with attractive therapeutic opportunities. Med. Res. Rev. 2010, 30, 463–549. [Google Scholar] [CrossRef]
- Stamatiou, R.; Paraskeva, E.; Vasilaki, A.; Hatziefthimiou, A. The muscarinic antagonist gallamine induces proliferation of airway smooth muscle cells regardless of the cell phenotype. Pharmacol. Rep. 2019, 71, 225–232. [Google Scholar] [CrossRef]
- Jakubík, J.; Bacakova, L.; Lisá, V.; El-Fakahany, E.E.; Tucek, S. Activation of muscarinic acetylcholine receptors via their allosteric binding sites. Proc. Natl. Acad. Sci. USA 1996, 93, 8705–8709. [Google Scholar] [CrossRef] [Green Version]
- Schrag, A.; Schelosky, L.; Scholz, U.; Poewe, W. Reduction of parkinsonian signs in patients with Parkinson’s disease by dopaminergic versus anticholinergic single-dose challenges. Mov. Disord. 1999, 14, 252–255. [Google Scholar] [CrossRef]
- Lazareno, S.; Buckley, N.J.; Roberts, F.F. Characterization of muscarinic M4 binding sites in rabbit lung, chicken heart, and NG108-15 cells. Mol. Pharmacol. 1990, 38, 805–815. [Google Scholar] [PubMed]
- Garcia, N.; Santafé, M.; Salon, I.; Lanuza, M.; Tomàs, J. Expression of muscarinic acetylcholine receptors (M1-, M2-, M3-and M4-type) in the neuromuscular junction of the newborn and adult rat. Histol. Histopathol. 2005, 20, 733–744. [Google Scholar] [PubMed]
- Lazartigues, E.; Brefel-Courbon, C.; Tran, M.; Montastruc, J.; Rascol, O. Spontaneously hypertensive rats cholinergic hyper-responsiveness: Central and peripheral pharmacological mechanisms. Br. J. Pharmacol. 1999, 127, 1657–1665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crans, R.A.; Wouters, E.; Valle-León, M.; Taura, J.; Massari, C.M.; Fernández-Dueñas, V.; Stove, C.P.; Ciruela, F. Striatal Dopamine D2-Muscarinic Acetylcholine M1 Receptor–Receptor Interaction in a Model of Movement Disorders. Front. Pharmacol. 2020, 11, 194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Airhart, M.J.; Lee, D.H.; Wilson, T.D.; Miller, B.E.; Miller, M.N.; Skalko, R.G. Movement disorders and neurochemical changes in zebrafish larvae after bath exposure to fluoxetine (PROZAC). Neurotoxicol. Teratol. 2007, 29, 652–664. [Google Scholar] [CrossRef]
- Avdesh, A.; Chen, M.; Martin-Iverson, M.T.; Mondal, A.; Ong, D.; Rainey-Smith, S.; Taddei, K.; Lardelli, M.; Groth, D.M.; Verdile, G. Regular care and maintenance of a zebrafish (Danio rerio) laboratory: An introduction. JoVE J. Vis. Exp. 2012, 69, e4196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarvis, R.; Knowles, J. DNA damage in zebrafish larvae induced by exposure to low-dose rate γ-radiation: Detection by the alkaline comet assay. Mutat. Res. Genet. Toxicol. Environ. Mutagenesis 2003, 541, 63–69. [Google Scholar] [CrossRef]
- Alderton, W.; Berghmans, S.; Butler, P.; Chassaing, H.; Fleming, A.; Golder, Z.; Richards, F.; Gardner, I. Accumulation and metabolism of drugs and CYP probe substrates in zebrafish larvae. Xenobiotica 2010, 40, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Best, J.D.; Berghmans, S.; Hunt, J.J.; Clarke, S.C.; Fleming, A.; Goldsmith, P.; Roach, A.G. Non-associative learning in larval zebrafish. Neuropsychopharmacology 2008, 33, 1206–1215. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, C. The husbandry of zebrafish (Danio rerio): A review. Aquaculture 2007, 269, 1–20. [Google Scholar] [CrossRef]
- Webb, B.; Sali, A. Comparative Protein Structure Modeling Using MODELLER. Curr. Protoc. Bioinform. 2016, 54, 5.6.1–5.6.37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Accelrys_Software_Inc. Discovery Studio Modeling Environment; Release 3.0; Accelrys Software Inc.: San Diego, CA, USA, 2007. [Google Scholar]
- Shen, M.Y.; Sali, A. Statistical potential for assessment and prediction of protein structures. Protein Sci. A Publ. Protein Soc. 2006, 15, 2507–2524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- John, B.; Sali, A. Comparative protein structure modeling by iterative alignment, model building and model assessment. Nucleic Acids Res. 2003, 31, 3982–3992. [Google Scholar] [CrossRef] [PubMed]
- Venkatachalam, C.M.; Jiang, X.; Oldfield, T.; Waldman, M. LigandFit: A novel method for the shape-directed rapid docking of ligands to protein active sites. J. Mol. Graph. Model. 2003, 21, 289–307. [Google Scholar] [CrossRef]
- Vogt, G.F., Jr.; Coon, T.G. A comparison of the foraging behavior of two darter (Etheostoma) species. Copeia 1990, 1, 41–49. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Uniprot (ID) | Length (a.a.) | PDB ID/Organism/Identity with D. rerio | Dock Score |
|---|---|---|---|---|
| chrm1a | A0A140LG95 | 465 | 6OIJ/human (Homo sapiens)/74.05% | 35.909 |
| chrm2a | B3DKN8 | 495 | 6OIK/human (Homo sapiens)/90.29% | 36.896 |
| chrm2b | F8W634 | 466 | 5ZK8/human (Homo sapiens)/82.12% | 34.155 |
| chrm3a | X1WHZ7 | 595 | 4DAJ/brown rat (Rattus norvegicus)/81.48% | 38.108# |
| chrm3b | U3JAM0 | 494 | 4U15/brown rat (Rattus norvegicus)/80.59% | 38.419# |
| chrm4a | E7F3U8 | 513 | 5DSG/human (Homo sapiens)/87.50% | 34.285 |
| chrm5a | B3DJA3 | 490 | 6OIJ/human (Homo sapiens)/74.91% | 26.621 |
| chrm5b | A0A2R8RMF6 | 505 | 4U14/brown rat (Rattus norvegicus)/74.33% | 1.839 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siregar, P.; Audira, G.; Feng, L.-Y.; Lee, J.-H.; Santoso, F.; Yu, W.-H.; Lai, Y.-H.; Li, J.-H.; Lin, Y.-T.; Chen, J.-R.; et al. Pharmaceutical Assessment Suggests Locomotion Hyperactivity in Zebrafish Triggered by Arecoline Might Be Associated with Multiple Muscarinic Acetylcholine Receptors Activation. Toxins 2021, 13, 259. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13040259
Siregar P, Audira G, Feng L-Y, Lee J-H, Santoso F, Yu W-H, Lai Y-H, Li J-H, Lin Y-T, Chen J-R, et al. Pharmaceutical Assessment Suggests Locomotion Hyperactivity in Zebrafish Triggered by Arecoline Might Be Associated with Multiple Muscarinic Acetylcholine Receptors Activation. Toxins. 2021; 13(4):259. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13040259
Chicago/Turabian StyleSiregar, Petrus, Gilbert Audira, Ling-Yi Feng, Jia-Hau Lee, Fiorency Santoso, Wen-Hao Yu, Yu-Heng Lai, Jih-Heng Li, Ying-Ting Lin, Jung-Ren Chen, and et al. 2021. "Pharmaceutical Assessment Suggests Locomotion Hyperactivity in Zebrafish Triggered by Arecoline Might Be Associated with Multiple Muscarinic Acetylcholine Receptors Activation" Toxins 13, no. 4: 259. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13040259