2.1. Cytotoxic Effects of Protein Toxins on Cancer and Normal Cells
Among several human cancer cells, human liver cancer cell line, HepG2, was reported to be a highly susceptible cell line to PS2 [
11]. Therefore, to explore the cytotoxic effects of Bin toxin on human cancer cell lines compared with PS2, HepG2 cell line was used as a model system in this study. Previously, Bin proteins (BinA, BinB and BinA + BinB) at high concentrations have been reported to inhibit the cell proliferation of A549 and HepG2 cells [
11,
12]. In the present study, the cytotoxic effects on human cells were investigated at comparatively lower concentrations of protein toxins. The morphological alterations induced by protein toxins during 24 h incubation in Hs68 and HepG2 cell lines were observed by inverted light microscopy as presented in
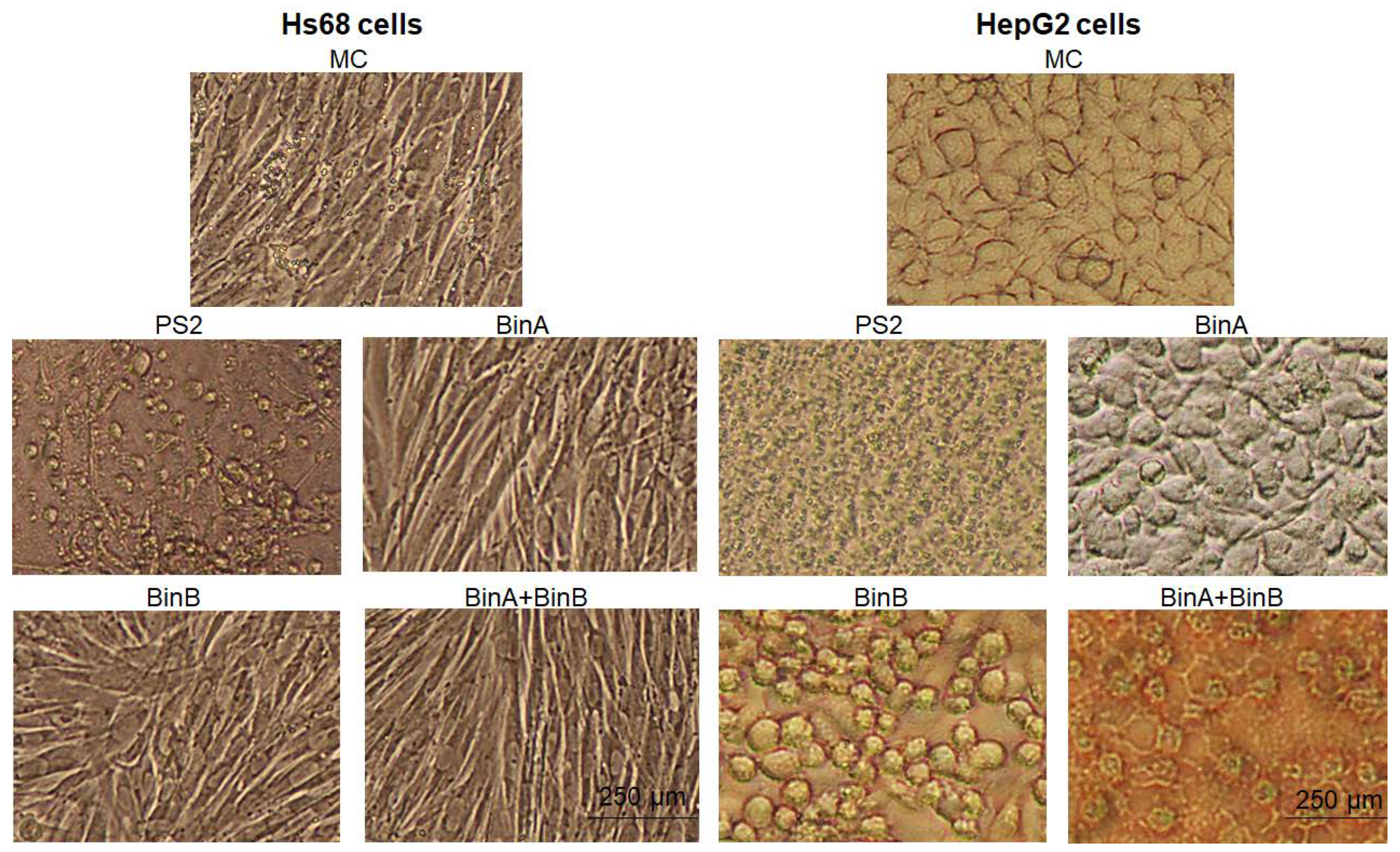
Figure 1. No cytopathic change was observed in Hs68 cells upon Bin toxin treatment when compared with mock control (MC)-treated cells, whereas PS2 (0.5 µg/mL), which is known to cause cell killing via creating pores in the plasma membrane of cells [
13], caused cell blebbing and clustering in Hs68 cells. On the other hand, complete cell necrosis and lysis were observed in HepG2 cells treated with PS2. The treatment with BinA (50 µg/mL) seemed to inhibit the proliferation of HepG2 cells when compared with MC-treated cells, whereas both BinB (50 µg/mL) and BinA + BinB (1:1) treatments altered the cell morphology by forming cell clumping and shrinkage along with an antiproliferative effect on HepG2 cells. Mesh formation was also observed in HepG2 cells treated with BinA + BinB. These results suggest that Bin and PS2 exert different cytotoxic patterns towards the target cells.
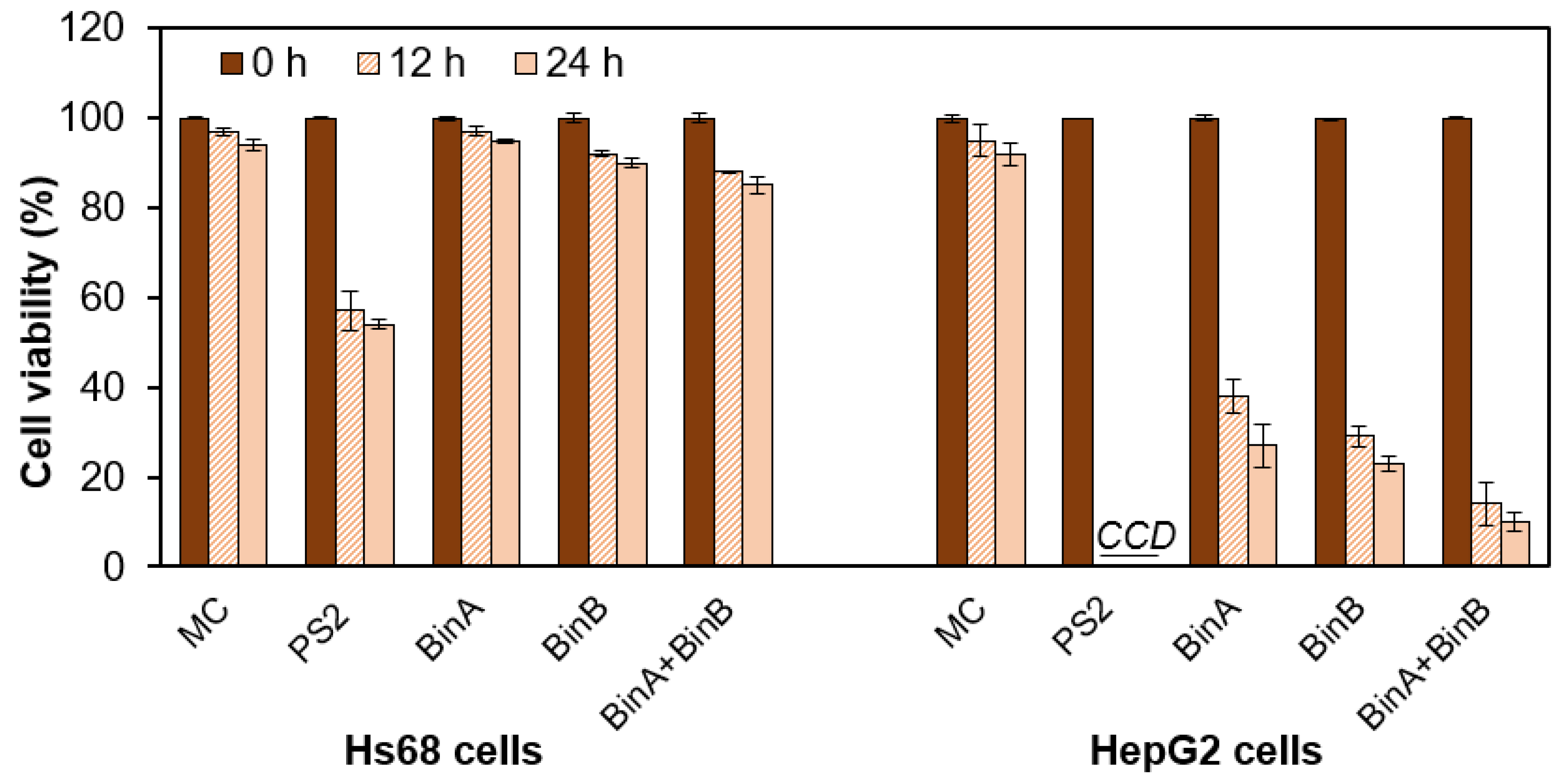
Activated Bin toxin from
L. sphaericus significantly affected the HepG2 cell viability as examined from one-dose trypan blue exclusion assay for 12 and 24 h treatment (
Figure 2). Up to 12 h after inoculation, BinB caused a decrease in cell viability resulting in ~29% cell survival rate in HepG2 cells, and BinA caused ~38% cell survival rate. Notably, BinA + BinB (1:1) treatment decreased the cell viability to ~14%. In contrast, the viability of Hs68 cells remained completely unaffected upon BinA treatment, whereas BinB and BinA + BinB had minor cytotoxic effects (~5–9% when compared to MC-treated Hs68 cells). Cells showed >94% viability when treated with MC. When the incubation period was extended to 24 h, the viability of HepG2 cells further decreased to ~11%, 6% and 4% for BinA, BinB and BinA + BinB treatments, respectively. However, the viability of Hs68 cells was mildly affected by extending the incubation period to 24 h with protein toxins. The results obtained from the cytotoxic effects of Bin proteins on cancer cells were found concomitant with the cell morphological changes induced by the toxin and also confirm the non-cytotoxicity of Bin proteins on normal cells. As compared to the cytotoxicity induced by Bin toxin, HepG2 cells were found to be more sensitive to PS2, which caused complete cell death. This agrees with the previous studies on HepG2 cells, showing the involvement of efficient cytocidal action of PS2 protein via pore formation in the membrane that leads to organelle fragmentation, cell swelling and ultimately cell lysis [
9,
10]. However, in the present study, it was observed that PS2 caused significant cell death (~46% reduction in cell survival rate) in Hs68 cells, which was quite contrary to various previous reports that suggested that normal cells such as normal lymphocytes, Vero (monkey kidney normal cells), normal mouse splenocytes and thymocytes show either no or minimal sensitivity toward parasporin proteins [
14,
15].
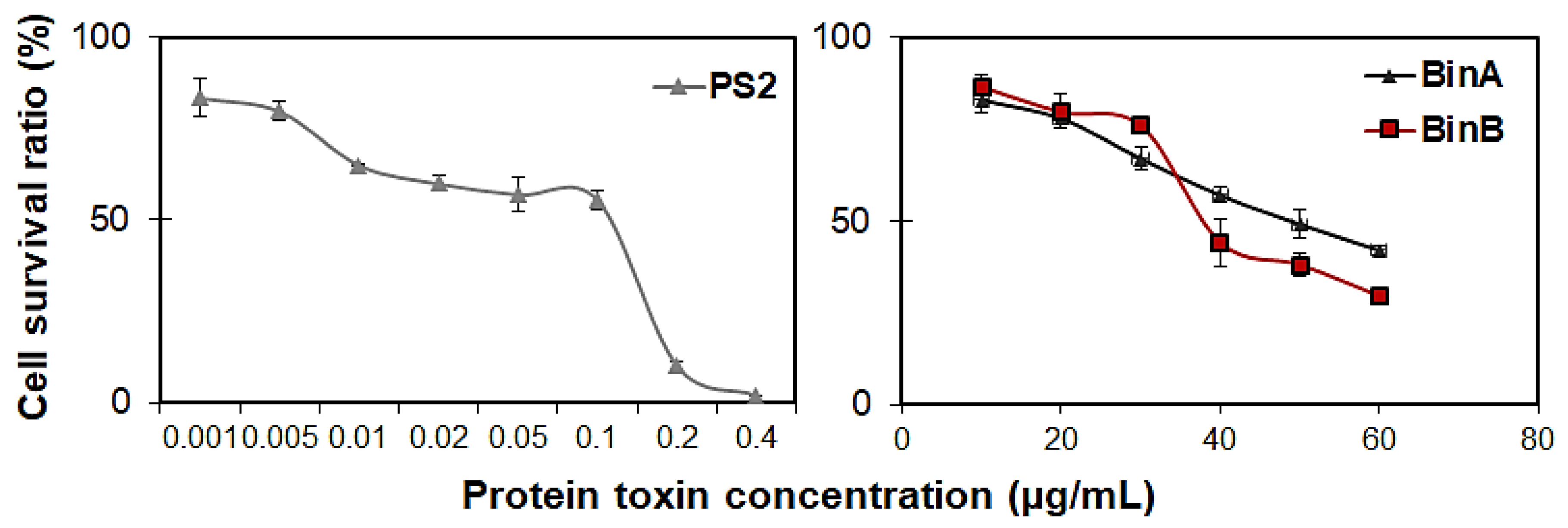
The results of the concentration–response study conducted by MTT assay in HepG2 cells indicated that the cell survival rate is significantly affected by protein toxins in a dose-dependent manner (
Figure 3). The EC
50 value for PS2 protein deduced from concentration–response analyses at 24 h after inoculation was 24 ng/mL, whereas, BinA and BinB had EC
50 values of 46.56 and 39.72 µg/mL against HepG2 cells, respectively (
Table 1).
The decrease in the cytotoxic effect on HepG2 cells caused by the combination of Bin subunits in comparison to individual Bin toxin subunits observed in the present study was in concordance with a previous report by Chankamngoen et al. [
11]. In fact, Surya et al. [
16] indicated that change in conformational state of Bin toxin monomeric subunits to heterodimeric form occurs when present together in an equimolar ratio in an aqueous solution. The formation of heterodimers in BinA + BinB might affect the interaction of Bin toxin with membrane or with the targeted receptor in susceptible cancer cell lines, resulting in the difference in cytotoxic effects. On the other hand, despite the structural similarities of Bin toxin and PS2, the results from our study indicate that the cytopathological effects of Bin toxin on HepG2 are possibly caused in a manner distinct from that of PS2. To further test our hypothesis, the effects of protein toxins on plasma membrane integrity and apoptotic characteristics of human cells were assessed.
2.2. Effect of Protein Toxins on Plasma Membrane Permeability and Caspase Activation in Human Cells
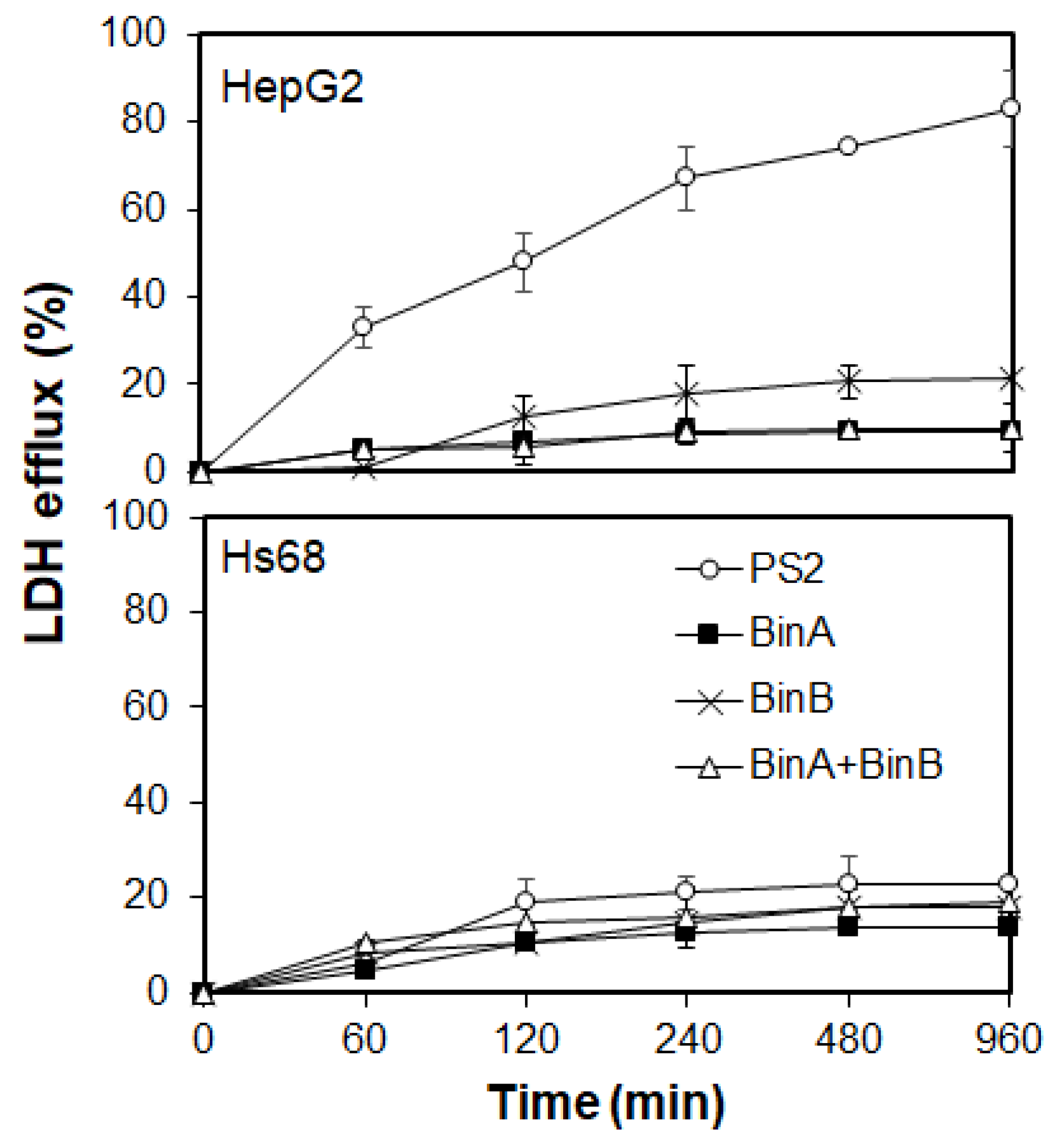
Cytocidal actions induced by various types of β-PFTs involve different mechanisms such as apoptotic cell death or pore formation in the plasma membrane of cells [
13,
14]. Studies emphasize that cytocidal actions induced by Bt-derived Cry toxins and Bin toxin in target cells of insect larvae are caused by receptor-mediated membrane interactions and cytoplasmic leakage leading to ultimate cell lysis [
15,
17], a mechanism similar to the cytocidal activity of β-PFTs. However, it is not clear whether the Bin-toxin-mediated cytocidal action on HepG2 cells is elicited by cell lysis via membrane disruption or cellular internalization resulting in intracellular cytotoxicity or apoptotic cell death. To confirm the mode of cell death, additional characterization of intracellular events in HepG2 cells treated with Bin proteins was completed to investigate the possible involvement of apoptosis. The effect of protein toxins on plasma membrane permeability of Hs68 and HepG2 cells was investigated by monitoring the LDH leakage from the cytoplasm of cells. LDH present in all cell types is instantly released into the cell culture medium upon plasma membrane damage, and its extracellular levels could be determined colorimetrically. The rate of LDH efflux accounts for the population of damaged cells and hence determines the cytotoxicity levels caused by disruption of plasma membrane permeability. In the present study, both cell lines were treated with Bin proteins and PS2 as described in
Section 3.3 and incubated for 1 to 16 h at 37 °C prior to performing LDH assay. The LDH efflux increased in PS2- and BinB-treated HepG2 cells with the increase in incubation time, as shown in
Figure 4. The LDH leakage from BinB-treated HepG2 cells was slightly higher than that from cells treated with BinA and BinA + BinB, although the overall LDH leakage was not too high in both of the Bin-toxin-treated HepG2 and Hs68 cells, suggesting that Bin toxin does not induce noticeable damage to the membrane permeability. A significant escalation in LDH efflux in PS2-treated HepG2 cells was detected within 60 min, depicting substantial damage caused by PS2 to the plasma membrane permeability leading to cell lysis and death corresponding to the cytotoxic effects explained earlier in this study (
Figure 1 and
Figure 2). Our results are supported by the previous study of Kitada et al. [
13] on the cytocidal effects of PS2 on HepG2 cells, elucidating that this toxin causes rapid membrane depolarization in HepG2 cells following protein efflux from the cells. However, PS2 did not seem to cause significant damage to the plasma membrane of Hs68 cells, as apparent from the lower LDH efflux. These findings suggest that Bin toxin might not cause pore formation in the plasma membrane of susceptible cancer cells as compared with PS2 that causes noticeable damage to the plasma membrane permeability, manifesting a typical characteristic of the β-PFT family.
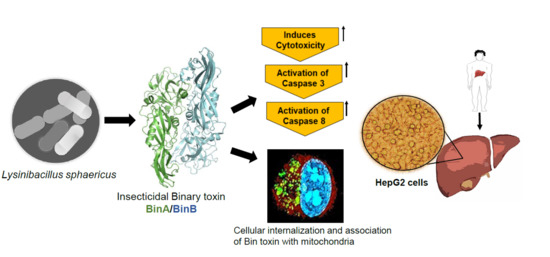
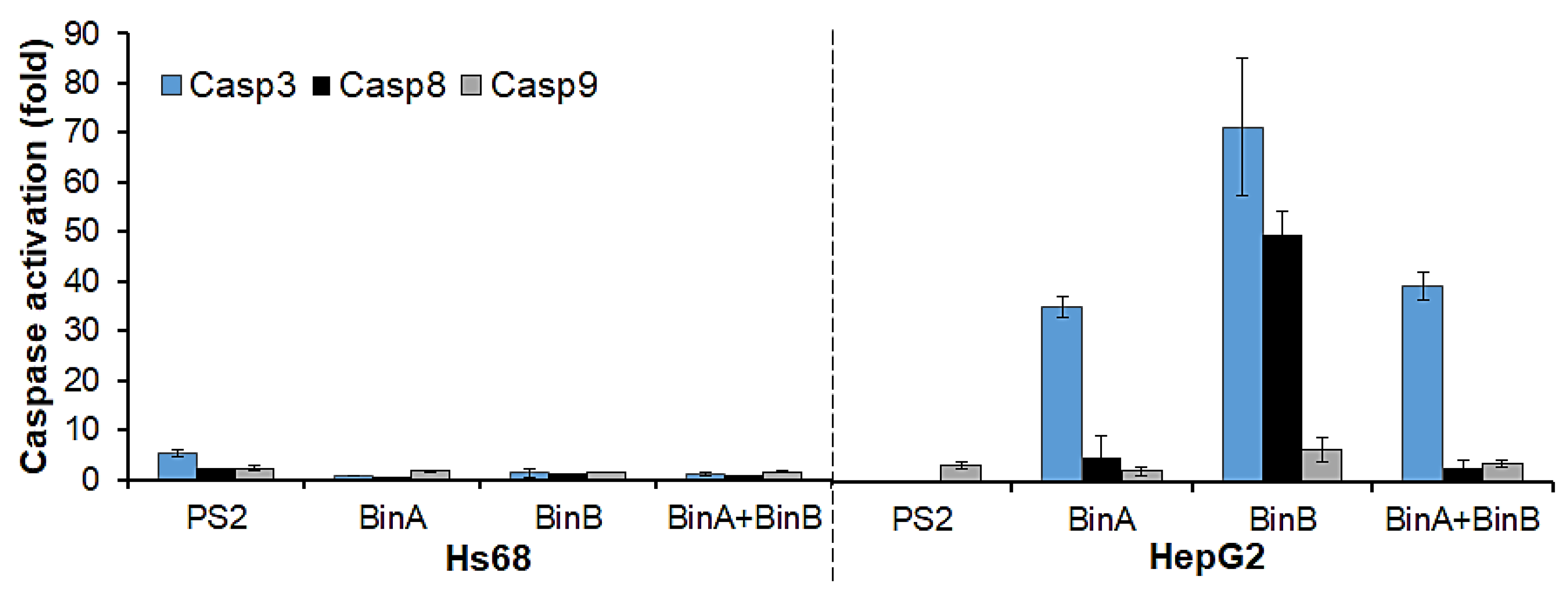
To further test the possibility of apoptosis induction by protein toxins in Hs68 and HepG2 cells, the levels of apoptotic mediators (caspases 3, 8 and 9) were determined using Caspase 3, 8 and 9 Multiplex Activity Assay. The activity levels (
Figure 5) of initiator Casp8 and executioner Casp3 were highly elevated in BinA- and BinB-treated HepG2 cells. The Casp3 and 8 activities were increased by ~35- and 5-fold in BinA-treated cells, respectively. Respective increments of ~71- and 50-fold in Casp3 and 8 activity levels were observed in BinB-treated cells. The activity level of Casp9 was increased up to ~6.5-fold in response to BinB treatment. On the other hand, BinA + BinB treatment resulted in an increase in Casp3 activity up to ~39-fold, but little increase in activity levels of Casp8 (about 3-fold) and Casp9 (about 4-fold) was observed. The PS2 treatment in HepG2 cells tends to enhance the activity of Casp9 by ~3.4-fold, whereas activities of Casp3 and 8 were not detected. These results are in line with the previous study by Brasseur et al. [
9] in which the presence of cleaved Casp9 via Western blot analysis in PS2-treated HepG2 cells was reported but Casp8 cleavage/activation was not observed. It was also noticed that no caspase activity could be seen in HepG2 cells treated for 24 h with PS2 at higher concentration (0.5 µg/mL) (data not shown), as the cells had already undergone necrosis and complete lysis prior to induction in caspase activity owing to significant plasma membrane damage caused by PS2. Even though the cell damage had apparently proceeded, apoptotic processes do occur in an elusive manner. It was also mentioned previously [
13] that caspase activation is not observed in HepG2 cells treated with PS2 at higher doses. On the other hand, Bin proteins did not seem to alter the plasma membrane integrity of HepG2 cells as much as PS2 (
Figure 4) and instead reduced the cell viability by causing the intracellular apoptotic events after translocating inside the cells, which is in concordance with the decline in cell survival rate observed in the cytotoxicity assay (
Figure 2). In the case of Hs68 cells treated with Bin proteins, no significant increase in caspase activity could be measured by the caspase assay. However, in this cell line, PS2 treatment induced the Casp3 activity up to ~5-fold. The cytocidal action of PS2 on Hs68 cells was never studied before, but other normal tissue cell lines that were studied were not found susceptible to PS2 [
13,
14]. However, our results seem to contradict the previous studies [
9,
13,
14] stating that PS2 has no or little cytotoxic effect on normal tissue cells; in the present study, a cell death rate of approximately 46% was accompanied by marked cytopathic changes (
Figure 1 and
Figure 2), and Casp3 activation (
Figure 5) was noticed in Hs68 cells in response to 0.5 µg/mL of PS2.
In caspase-dependent apoptosis (programmed cell death), Casp3, the effector caspase, requires initiator caspases from either the intrinsic (Casp9) or extrinsic (Casp8) pathway for its activation [
18]. Our results indicated that activity levels of Casp3 were very high in BinA- and BinB-treated cancer cells and corresponded to the activity levels of Casp8, reinforcing the idea of the induction of the extrinsic apoptosis pathway by individual Bin toxin subunits. Moreover, the BinA + BinB treatment increased the activities of Casp3 and Casp9 (which had activity slightly higher than Casp8) in HepG2 cells, indicating the induction of the mitochondrial apoptosis pathway [
19]. The association of Bin toxin with HepG2 mitochondria is discussed in the next section.
2.3. Intracellular Localization and Mitochondrial Association of Bin Toxin in HepG2 Cells
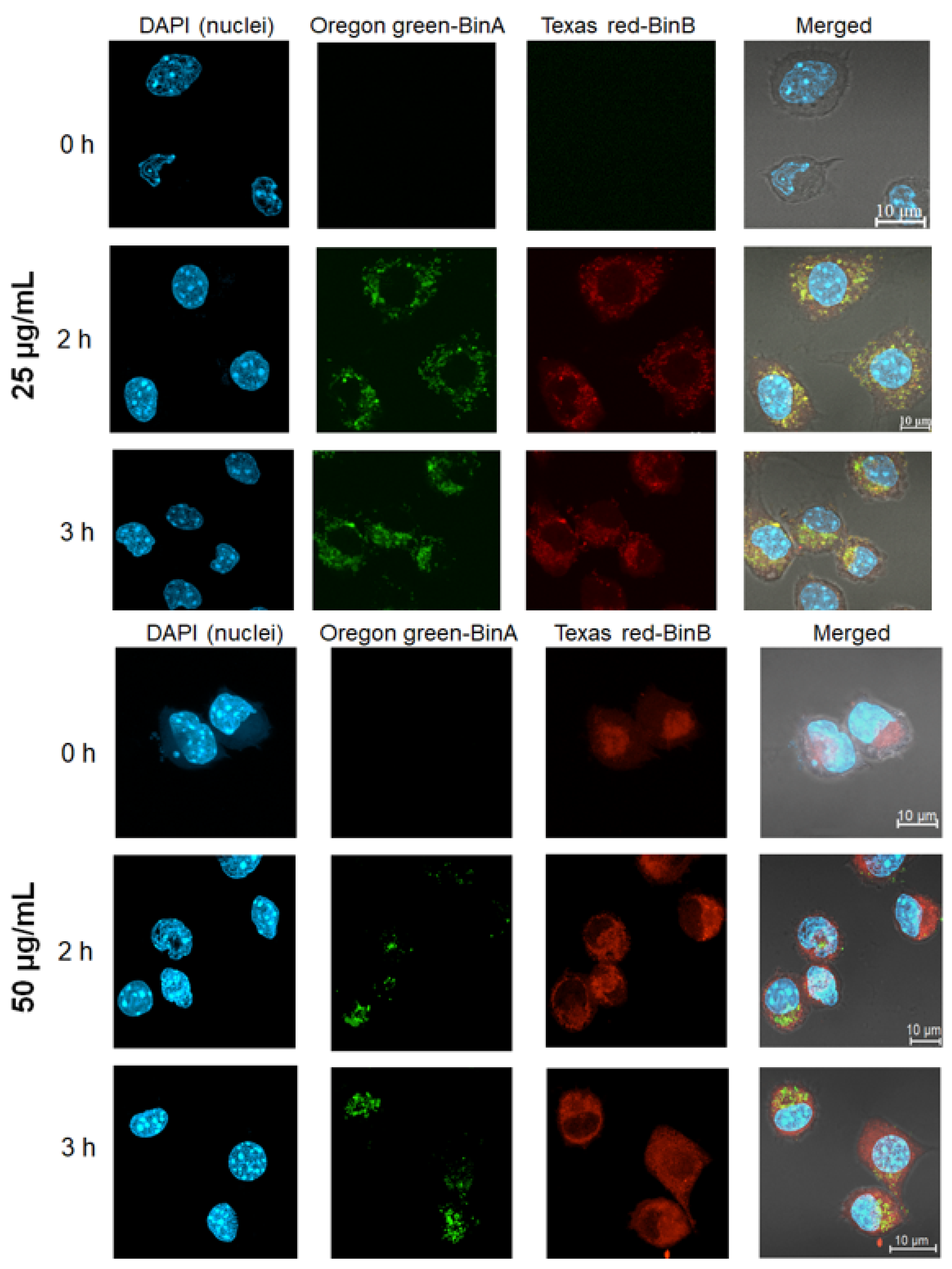
To gain an in-depth understanding of the cytotoxic activity of Bin in the HepG2 cell line, BinA and BinB proteins fluorescently labeled with Oregon Green and Texas Red, respectively, were tracked for their internalization and co-localization patterns in the cells. The analysis of co-localization pattern was conducted with a combination of Oregon Green labeled BinA and Texas Red labeled BinB (1:1) at 25 µg/mL and 50 µg/mL, and the efficiency of labeling was calculated by Beer–Lambert law. The degree of labeling for Oregon Green labeled BinA and Texas Red labeled BinB was calculated as 1.42 and 1.59, respectively. Both Texas Red and Oregon Green succinimidyl ester dyes, also known as amine-reactive dyes, bind with the amine groups of the proteins. The biological activity of Bin toxin with a similar fluorescent labeling was previously analyzed via in vivo biological assay by feeding Bin toxin to
Culex quinquefasciatus larvae that showed comparable LC
50 values of fluorescently-labeled and unlabeled Bin toxin, indicating no functional alteration upon fluorescent labeling [
3]. The intracellular co-localization of 25 µg/mL of Oregon Green labeled BinA/Texas Red labeled BinB was observed as yellow fluorescence after 2 h of toxin treatment, as shown in
Figure 6. It was found that Texas Red labeled BinB was quickly internalized in the HepG2 cytoplasm; in comparison, Oregon Green labeled BinA was mostly localized in the cell membrane (data not shown). Similar internalization of Bin toxin was previously observed in susceptible mosquito gut cells, where BinA was detected only on the cell membrane but could not be detected inside the cytoplasm of mosquito gut cells, whereas BinB was detected inside the cytoplasm of susceptible mosquito midgut cells, suggesting that BinB alone can enter into mosquito larvae gut cells [
3]. The toxin internalization signals were intensified with the increase in the concentration to 50 µg/mL and the increase in the incubation time up to 3 h. It is not clear yet whether the BinA/BinB complex was transiently formed during membrane translocation, as BinA and BinB free molecules were also detected after localization in HepG2 cell cytoplasm. It might be possible that the BinA/BinB complex transiently formed prior to internalization undergoes dissociation after intracellular localization.
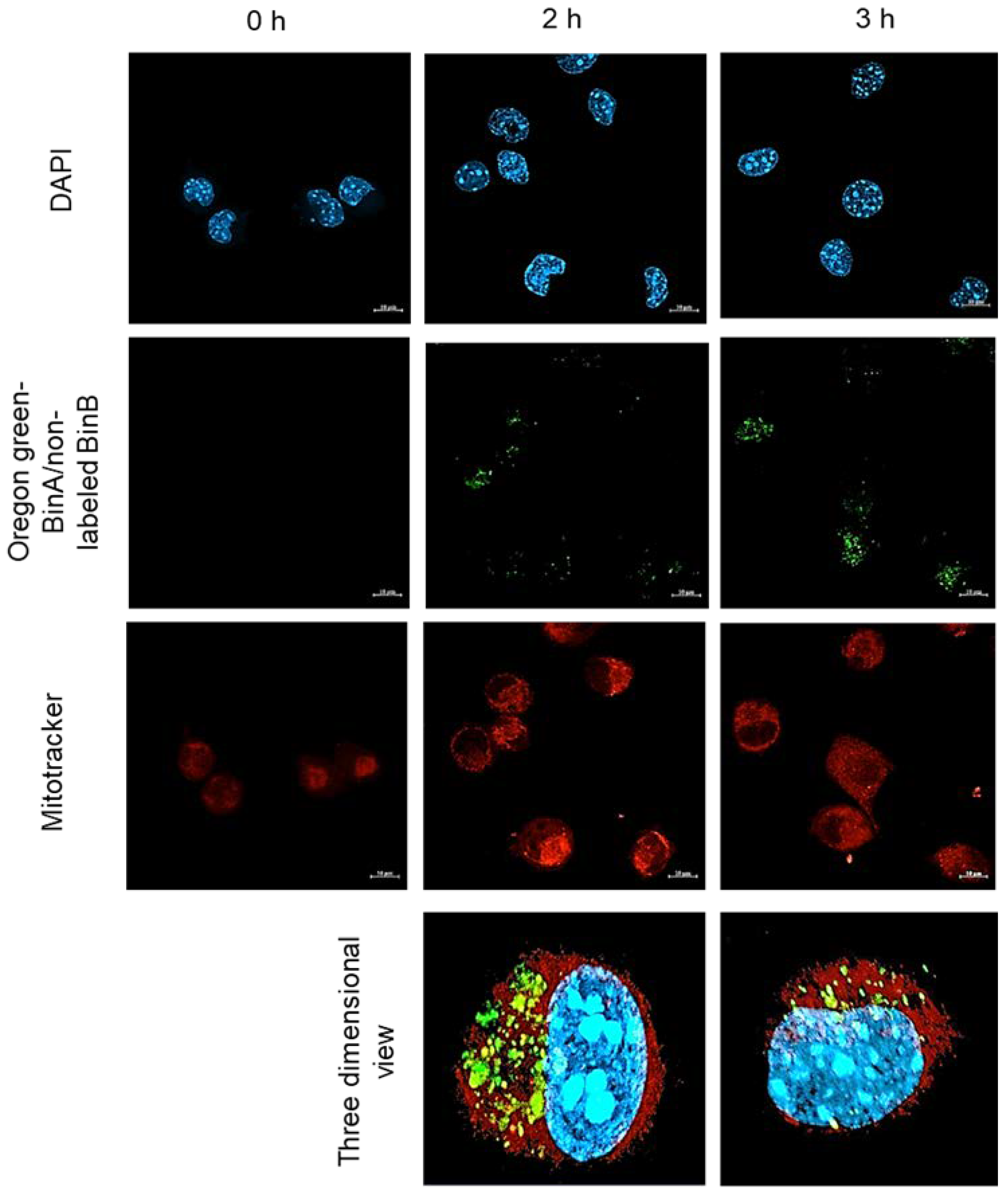
The association of internalized Bin toxin with mitochondria of HepG2 cells was investigated by treating the cells with a 1:1 combination of 50 µg/mL of Oregon Green labeled BinA and non-labeled BinB. The mitochondria were labeled with Mitotracker Red to detect the co-localized toxin in yellow fluorescent signal. The confocal images showed that the mitochondria were stained with red signal that was distributed throughout the cytoplasm of HepG2 cells (
Figure 7). The fluorescent signal of Oregon Green labeled BinA/nonlabeled BinB was detected after 2 and 3 h of toxin treatment, and the signal was slightly co-localized with mitochondria giving a yellow fluorescent signal. Results revealed that BinB toxin was internalized and localized in the cytoplasm of HepG2 cells after 2 h of incubation as the signal could be seen throughout the cytoplasm. However, the co-localization of BinA with mitochondria was not as specific and strong as that of BinB as analyzed from Pearson’s coefficient values (data not shown). It was reported previously that Bin toxin induces cell vacuolization and mitochondrial swelling in
Culex larval gut cells [
20,
21,
22]. Although BinB seems to facilitate the internalization of BinA, the BinA/BinB complex might undergo dissociation after internalization in the cytoplasm of HepG2 cells, resulting in a lower association of BinA with mitochondria.
Considering the lower cytotoxicity of Bin toxin than that of PS2, the structural diversities may contribute to their different cytotoxic mechanisms. It has been proposed that the
N-terminal domains of Bin and PS2 toxins play a major role in recognizing the target receptors based on their lectin-like structures. Several exposed aromatic residues are located on the surface of the
N-terminal domain of PS2 and are predicted to interact with the carbohydrate groups of the target receptor on the cancer cell surface [
7]. Although some clusters of aromatic residues are present in the
N-terminal domains of Bin proteins [
5], high structural diversity between Bin and PS2 is found in these domains. It was also noted that Bin toxin does not form the SDS-resistant oligomers such as aerolysin protein when exposed to detergents or lipid bilayers [
16], indicating the difference between the conformational states of Bin toxin and PS2 when exposed to the plasma membrane of targeted cells. Despite the localization of Oregon Green labeled BinA on the cell membrane of HepG2 cells, the induction of apoptotic events via caspase activation was evident, as shown earlier in
Figure 5, indicating the possible interaction of BinA with cell membrane receptors. This characteristic is similar to some other protein toxins such as aerolysin produced by a Gram-negative bacterium
Aeromonas hydrophila [
23], which is reported to cause apoptosis in T cells by forming a small number of channels in the cell membrane, leading to a rapid increase in intracellular calcium levels. As a variety of receptors take part in caspase-dependent apoptosis, identification of the target cell receptors and receptor-binding motifs is expected to provide further insight into the mechanisms of target specificity and cytotoxicity of Bin and PS2 toxins.
In conclusion, we report that the L. sphaericus originated Bin toxin, having preferential cytotoxicity towards cancer cells, acts by mechanisms different than those of PS2. Our results suggest that Bin toxin triggers the apoptotic events by caspase induction in HepG2 cells. Altogether, our findings suggest that Bin toxin could be potentially engineered, e.g., by tagging with the tumor-specific peptides, and might serve as an intracellular delivery vector of anticancer therapeutics to target mitochondrial compartments to trigger apoptotic cell death of human cancer cells.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}