Host-Species Variation and Environment Influence Endophyte Symbiosis and Mycotoxin Levels in Chinese Oxytropis Species


 and
and
Abstract
:1. Introduction
2. Results
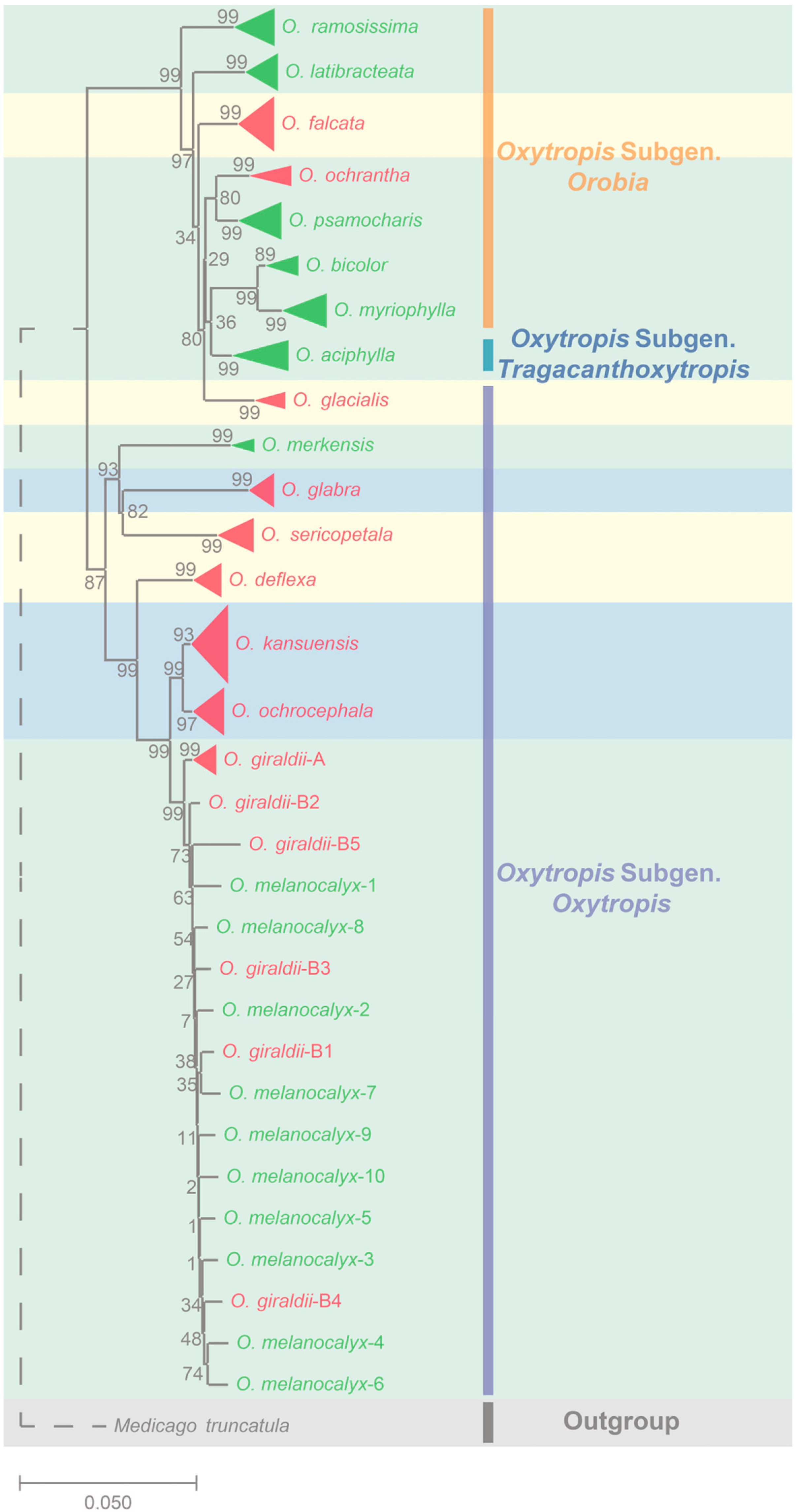
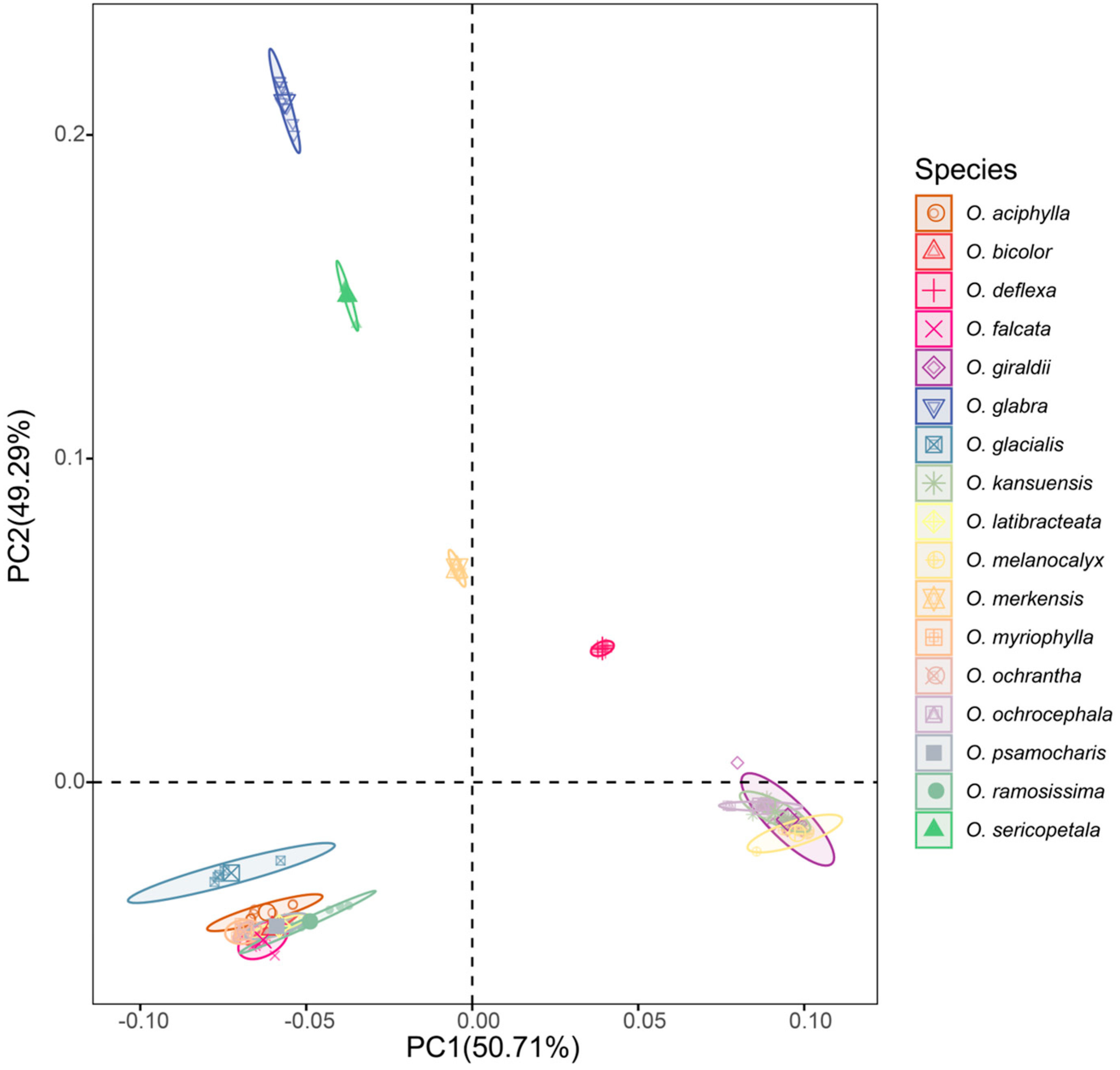
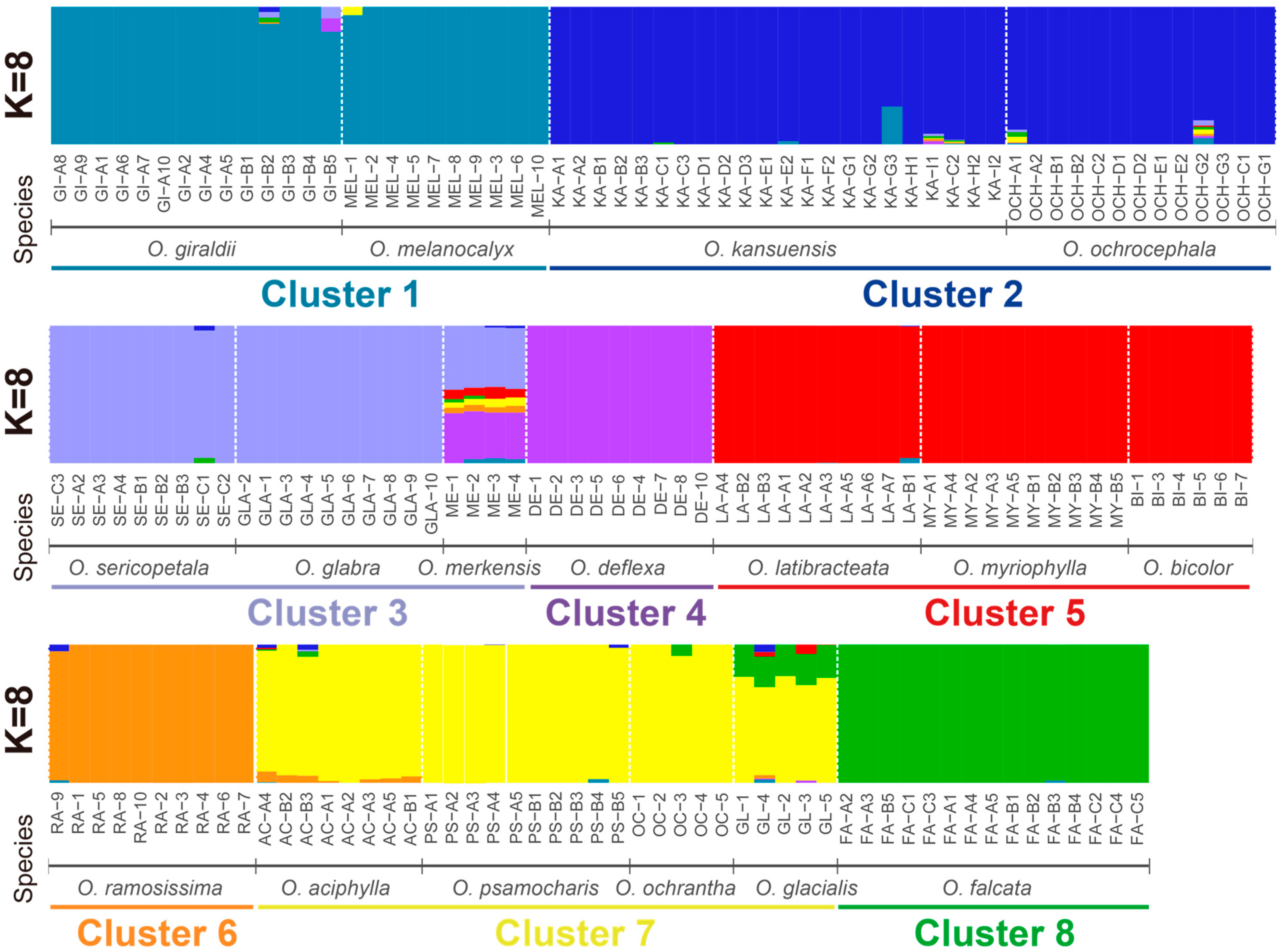
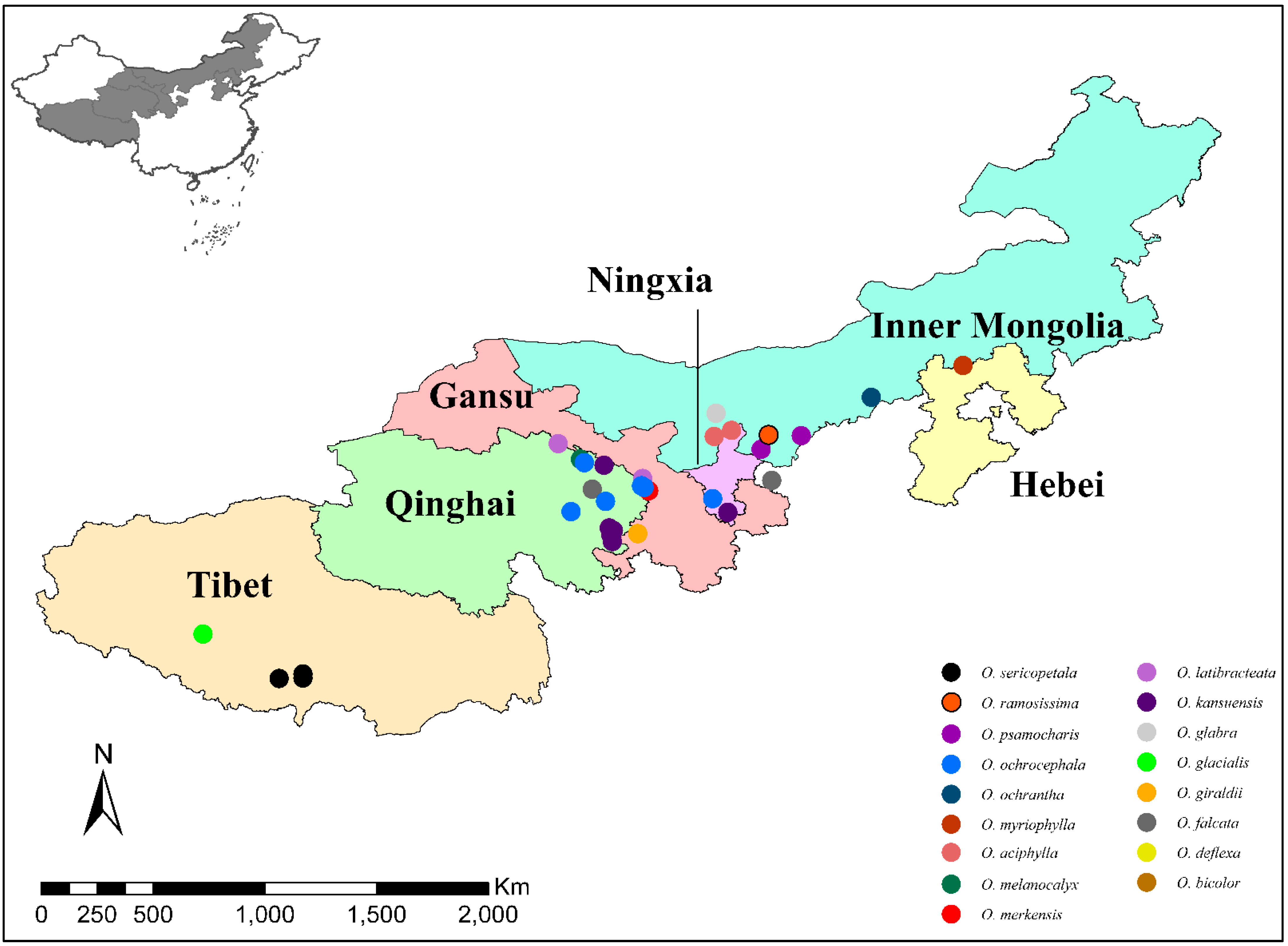
2.1. Genetic Evolution of the Natural Population of Oxytropis spp.
2.2. Quantification of A. oxytropis and Swainsonine of Oxytropis Plants
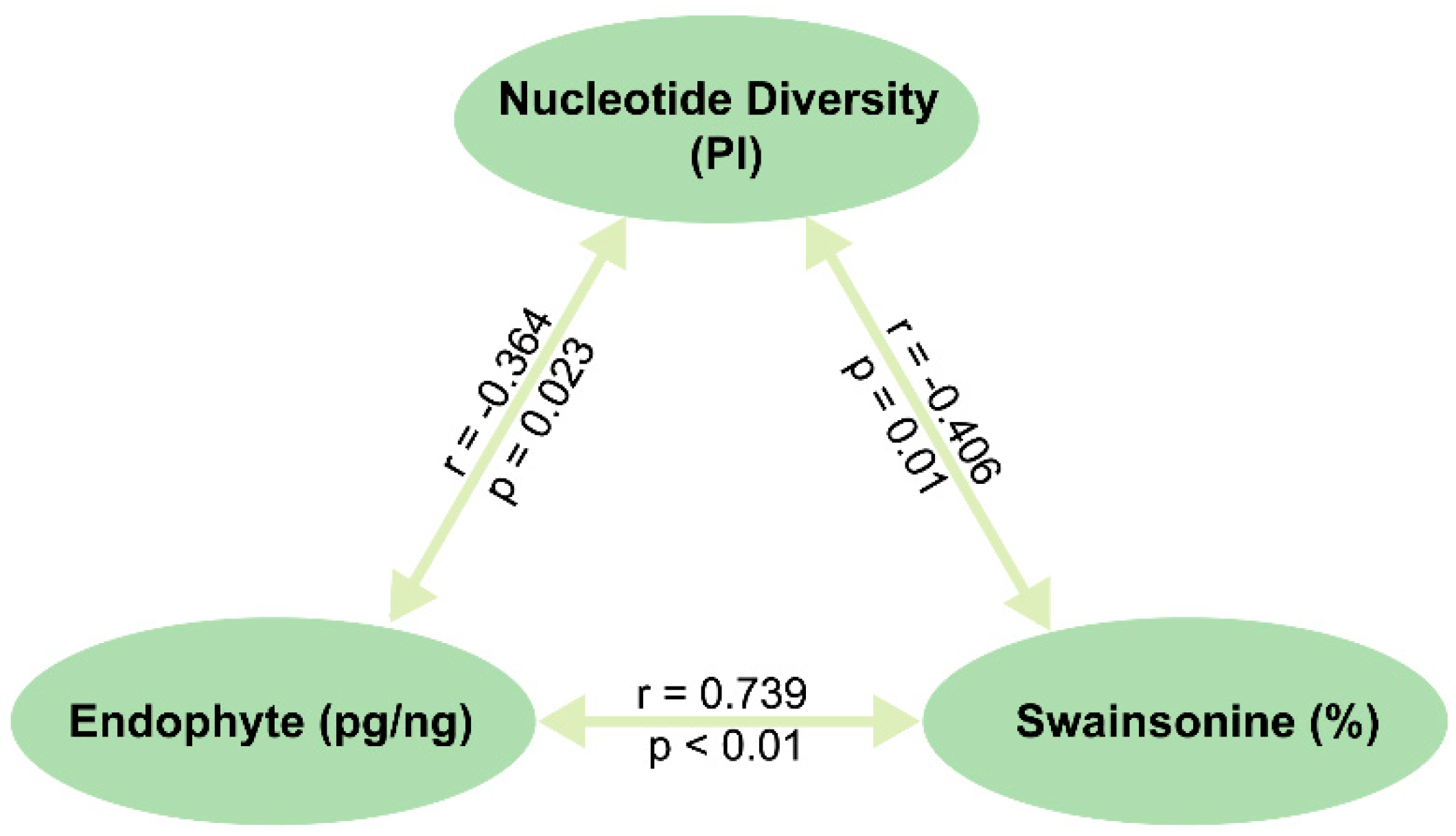
2.3. Interaction between Genetics and Environment and Their Integrated Effect on Endophyte Symbiosis and Swainsonine Production
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Materials and Genomic DNA Extraction
5.2. SLAF Sequencing and SNP Calling
5.3. Characterisation of Genetic Diversity and Genetic Structure
5.4. Determination of Endophyte and Swainsonine Concentration
5.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rodriguez, R.J.; White, J.F., Jr.; Arnold, A.E.; Redman, R.S. Fungal endophytes: Diversity and functional roles. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef]
- Saikkonen, K.; Young, C.A.; Helander, M.; Schardl, C.L. Endophytic Epichloё species and their grass hosts: From evolution to applications. Plant Mol. Biol. 2016, 90, 665–675. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.M.; Uroz, S.; Barker, D.G. Ancestral alliances: Plant mutualistic symbioses with fungi and bacteria. Science 2017, 356, eaad4501. [Google Scholar] [CrossRef]
- Zhao, Z.; Li, X.; Liu, M.F.; Merckx, V.S.F.T.; Saunders, R.M.K.; Zhang, D. Specificity of assemblage, not fungal partner species, explains mycorrhizal partnerships of mycoheterotrophic Burmannia plants. ISME J. 2021, 15, 1614–1627. [Google Scholar] [CrossRef]
- Harrison, J.G.; Grifin, E.A. The diversity and distribution of endophytes across biomes, plant phylogeny and host tissues: How far have we come and where do we go from here? Environ. Microbiol. 2020, 22, 2107–2123. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Gao, X.; Wang, J.; He, X.; Han, B. A review of the most economically important poisonous plants to the livestock industry on temperate grasslands of China. J. Appl. Toxicol. 2013, 33, 9–17. [Google Scholar] [CrossRef]
- Cook, D.; Gardner, D.R.; Lee, S.T.; Pfister, J.A.; Stonecipher, C.A.; Welsh, S.L. A swainsonine survey of North American Astragalus and Oxytropis taxa implicated as locoweeds. Toxicon 2016, 118, 104–111. [Google Scholar] [CrossRef]
- Wang, Q.; Nagao, H.; Li, Y.; Wang, H.; Kakishima, M. Embellisia Oxytropis, a new species isolated from Oxytropis kansuensis in China. Mycotaxon 2006, 95, 255–260. [Google Scholar]
- Pryor, B.; Creamer, R.; Shoemaker, R.; McLain-Romero, J.; Hambleton, S. Undifilum, a new genus for endophytic Embellisia oxytropis and parasitic Helminthosporium bornmuelleri on legumes. Botany 2009, 87, 178–194. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Groenewald, J.Z.; Binder, M.; Crous, P.W. Alternaria redefined. Stud Mycol 2013, 75, 171–212. [Google Scholar] [CrossRef]
- Molyneux, R.; James, L. Loco intoxication: Indolizidine alkaloids of spotted locoweed (Astragalus lentiginosus). Science 1982, 216, 190–191. [Google Scholar] [CrossRef] [PubMed]
- Colegate, S.M.; Dorling, P.R.; Huxtable, C.R. A spectroscopic investigation of swainsonine: An α-Mannosidase inhibitor isolated from Swainsona canescens. Aust. J. Chem. 1979, 32, 2257–2264. [Google Scholar] [CrossRef]
- Zhao, B.; Liu, Z.; Wan, X.; Huo, X.; Guo, X.; Wang, J.; Liu, Z.; Sun, L.; Shi, Z. Damage and control of poisonous-weeds in Chinese western grassland. Sci. Agric. Sin. 2008, 41, 3094–3103. [Google Scholar]
- Cook, D.; Ralphs, M.H.; Welch, K.D.; Stegelmeier, B.L. Locoweed poisoning in livestock. Rangelands 2009, 31, 16–21. [Google Scholar] [CrossRef]
- Lu, H.; Cao, D.; Ma, F.; Wang, S.; Yang, X.; Wang, W.; Zhou, Q.; Zhao, B. Characterisation of locoweeds and their effect on livestock production in the western rangelands of China: A review. Rangel. J. 2014, 36, 121–131. [Google Scholar] [CrossRef]
- China National Animal Husbandry Station. China’s Grassland Biological Disasters; China Agriculture Press: Beijing, China, 2018. [Google Scholar]
- Creamer, R.; Baucom, D. Fungal endophytes of locoweeds: A commensal relationship? J. Plant Physiol. Pathol. 2013, 1, 2. [Google Scholar] [CrossRef]
- Cook, D.; Shi, L.; Gardner, D.R.; Pfister, J.A.; Grum, D.; Welch, K.D.; Ralphs, M.H. Influence of phenological stage on swainsonine and endophyte concentrations in Oxytropis sericea. J. Chem. Ecol. 2012, 38, 195–203. [Google Scholar] [CrossRef]
- Cook, D.; Grum, D.S.; Gardner, D.R.; Welch, K.D.; Pfister, J.A. Influence of endophyte genotype on swainsonine concentrations in Oxytropis sericea. Toxicon 2013, 61, 105–111. [Google Scholar] [CrossRef]
- Cook, D.; Donzelli, B.G.G.; Creamer, R.; Baucom, D.L.; Gardner, D.R.; Pan, J.; Moore, N.; Krasnoff, S.B.; Jaromczyk, J.W.; Schardl, C.L. Swainsonine biosynthesis genes in diverse symbiotic and pathogenic fungi. G3 Genes|Genomes|Genet. 2017, 7, 1791–1797. [Google Scholar] [CrossRef]
- Cook, D.; Gardner, D.R.; Grum, D.; Pfister, J.A.; Ralphs, M.H.; Welch, K.D.; Green, B.T. Swainsonine and endophyte relationships in Astragalus mollissimus and Astragalus lentiginosus. J. Agric. Food Chem. 2011, 59, 1281–1287. [Google Scholar] [CrossRef]
- Ralphs, M.H.; Creamer, R.; Baucom, D.; Gardner, D.R.; Welsh, S.L.; Graham, J.D.; Hart, C.; Cook, D.; Stegelmeier, B.L. Relationship between the endophyte Embellisia spp. and the toxic alkaloid swainsonine in major locoweed species (Astragalus and Oxytropis). J. Chem. Ecol. 2008, 34, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Cook, D.; Gardner, D.R.; Ralphs, M.H.; Pfister, J.A.; Welch, K.D.; Green, B.T. Swainsoninine concentrations and endophyte amounts of Undifilum oxytropis in different plant parts of Oxytropis sericea. J. Chem. Ecol. 2009, 35, 1272–1278. [Google Scholar] [CrossRef] [PubMed]
- Ralphs, M.H.; Cook, D.; Gardner, D.R.; Grum, D.S. Transmission of the locoweed endophyte to the next generation of plants. Fungal Ecol. 2011, 4, 251–255. [Google Scholar] [CrossRef]
- Grum, D.S.; Cook, D.; Gardner, D.R.; Roper, J.M.; Pfister, J.A.; Ralphs, M.H. Influence of seed endophyte amounts on swainsonine concentrations in Astragalus and Oxytropis locoweeds. J. Agric. Food Chem. 2012, 60, 8083–8089. [Google Scholar] [CrossRef] [PubMed]
- Harrison, J.G.; Parchman, T.L.; Cook, D.; Gardner, D.R.; Forister, M.L. A heritable symbiont and host-associated factors shape fungal endophyte communities across spatial scales. J. Ecol. 2018, 106, 2274–2286. [Google Scholar] [CrossRef]
- He, W.; Guo, L.; Wang, L.; Zhao, Q.; Guo, L.; Cao, W.; Mur, L.A.J.; Wei, Y. Host genotype and precipitation influence of fungal endophyte symbiosis and mycotoxin abundance in a locoweed. Int. J. Mol. Sci. 2019, 20, 5285. [Google Scholar] [CrossRef] [PubMed]
- Gardner, D.R.; Mclain-Romero, J.M.; Ralphs, M.H.; Creamer, R. Correlation of an endophytic fungus (Alternaria spp.) with the presence of swainsonine in lambert locoweed (Oxytropis lambertii). Poisonous Plants Relat. Toxins 2004, 3, 32–37. [Google Scholar]
- Kholina, A.B.; Koren’, O.G.; Zhuravlev, Y.N. High polymorphism and autotetraploid origin of the rare endemic species Oxytropis chankaensis Jurtz. (Fabaceae) inferred from allozyme data. Russ. J. Genet. 2004, 40, 393–400. [Google Scholar] [CrossRef]
- Schönswetter, P.; Tribsch, A.; Niklfeld, H. Amplified Fragment Length Polymorphism (AFLP) reveals no genetic divergence of the Eastern Alpine endemic Oxytropis campestris subsp. tiroliensis (Fabaceae) from widespread subsp. campestris. Plant Syst. Evol. 2004, 244, 245–255. [Google Scholar] [CrossRef]
- Artyukova, E.V.; Kholina, A.B.; Kozyrenko, M.M.; Zhuravlev, Y.N. Analysis of genetic variation in rare endemic species Oxytropis chankaensis Jurtz. (Fabaceae) using RAPD markers. Russ. J. Genet. 2004, 40, 710–716. [Google Scholar] [CrossRef]
- Chung, M.; Gelembiuk, G.; Givnish, T.J. Population genetics and phylogeography of endangered Oxytropis campestris var. chartacea and relatives: Arctic-alpine disjuncts in eastern North America. Mol. Ecol. 2004, 13, 3657–3673. [Google Scholar] [CrossRef] [PubMed]
- Kholina, A.B.; Kozyrenko, M.M.; Artyukova, E.V.; Sandanov, D.V.; Andrianova, E.A. Phylogenetic relationships of the species of Oxytropis DC. subg. Oxytropis and Phacoxytropis (Fabaceae) from Asian Russia inferred from the nucleotide sequence analysis of the intergenic spacers of the chloroplast genome. Russ. J. Genet. 2016, 52, 780–793. [Google Scholar] [CrossRef]
- Tekpinar, A.D.; Erkul, S.K.; Aytaç, Z.; Kaya, Z. Phylogenetic relationships between Oxytropis DC. and Astragalus L. species native to an Old World diversity center inferred from nuclear ribosomal ITS and plastid matK gene sequences. Turk. J. Biol. 2016, 40, 250–263. [Google Scholar] [CrossRef]
- Almerekova, S.; Mukhitdinov, N.; Abugalieva, S. Phylogenetic study of the endemic species Oxytropis almaatensis (Fabaceae) based on nuclear ribosomal DNA ITS sequences. BMC Plant Biol. 2017, 17, 173. [Google Scholar] [CrossRef] [PubMed]
- Shavvon, R.S.; Osaloo, S.K.; Maassoumii, A.A.; Moharrek, F.; Erkul, S.K.; Lemmon, A.R.; Lemmon, E.M.; Michalak, I.; Muellner-Riehl, A.N.; Favre, A. Increasing phylogenetic support for explosively radiating taxa: The promise of high-throughput sequencing for Oxytropis (Fabaceae). J. Syst. Evol. 2017, 55, 385–404. [Google Scholar] [CrossRef]
- Archambault, A.; Strömvik, M. Evolutionary relationships in Oxytropis species, as estimated from the nuclear ribosomal internal transcribed spacer (ITS) sequences point to multiple expansions into the Arctic. Botany 2012, 90, 770–779. [Google Scholar] [CrossRef]
- Knief, C. Analysis of plant microbe interactions in the era of next generation sequencing technologies. Front. Plant Sci. 2014, 5, 216. [Google Scholar] [CrossRef]
- Kozurahova, E.; Richards, A.J. Breeding systems of rare and endemic Asytagalus, Oxytropis and Onobrychis species (Fabaceae) tested with alternative methods. C. R. I’académie Bulg. Des. Sci. 2016, 69, 1571–1580. [Google Scholar]
- Li, Y.; Lan, F.; Chang, Z.; Guo, Z. Molecular phylogeny of Oxytropis DC.of Qinghai-Tibetan Plateau by ITS and trnL-F sequences. J. Northwest A F Univ. (Nat. Sci. Ed.) 2011, 39, 187–193. [Google Scholar] [CrossRef]
- Schirrmann, M.K.; Zoller, S.; Croll, D.; Stukenbrock, E.H.; Leuchtmann, A.; Fior, S. Genomewide signatures of selection in Epichloё reveal candidate genes for host specialization. Mol. Ecol. 2018, 27, 3070–3086. [Google Scholar] [CrossRef]
- Gladieux, P. What makes a specialized endophyte special? Mol. Ecol. 2018, 27, 3037–3039. [Google Scholar] [CrossRef] [PubMed]
- Oldrup, E.; McLain-Romero, J.; Padilla, A.; Moya, A.; Gardner, D.; Creamer, R. Localization of endophytic Undifilum fungi in locoweed seed and influence of environmental parameters on a locoweed in vitro culture system. Botany 2010, 88, 512–521. [Google Scholar] [CrossRef]
- Saikkonen, K.; Wäli, P.; Helander, M.; Faeth, S.H. Evolution of endophyte-plant symbioses. Trends Plant Sci. 2004, 9, 275–280. [Google Scholar] [CrossRef]
- Sun, X.; Liu, D.; Zhang, X.; Li, W.; Liu, H.; Hong, W.; Jiang, C.; Guan, N.; Ma, C.; Zeng, H.; et al. SLAF-seq: An efficient method of large-scale de novo SNP discovery and genotyping using high-throughput sequencing. PLoS ONE 2013, 8, e58700. [Google Scholar] [CrossRef] [PubMed]
- Kent, W.J. Blat-the BLAST-like alignment tool. Genome Res. 2001, 12, 656–664. [Google Scholar] [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The genome analysis toolkit: A mapreduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. Mega7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Pickrell, J.K.; Pritchard, J.K. Inference of population splits and mixtures from genome-wide allele frequency data. PLoS Genet. 2012, 8, e1002967. [Google Scholar] [CrossRef]
- Gardner, D.R.; Molyneux, R.J.; Ralphs, M.H. Analysis of swainsonine: Extraction methods, detection, and measurement in populations of locoweeds (Oxytropis spp.). J. Agric. Food Chem. 2001, 49, 4573–4580. [Google Scholar] [CrossRef]
- Meirmans, P.G. Using the AMOVA framework to estimate a standardized genetic differentiation measure. Evolution 2006, 60, 2399–2402. [Google Scholar] [CrossRef]
- Cook, D.; Gardner, D.R.; Welch, K.D.; Roper, J.M.; Ralphs, M.H.; Green, B.T. Quantitative PCR method to measure the fungal endophyte in locoweeds. J. Agric. Food Chem. 2009, 57, 6050–6054. [Google Scholar] [CrossRef] [PubMed]
- Gardner, D.R.; Cook, D. A comparison of alternative sample preparation procedures for the analysis of swainsonine using LC-MS/MS. Phytochem. Anal. 2011, 22, 124–127. [Google Scholar] [CrossRef] [PubMed]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | d.f. | Sum of Squares | Variance Components | Percentage of Variation | Fixation Indices |
|---|---|---|---|---|---|
| Among 17 species | 16 | 30,048.556 | 175.96679 Va | 67.66 | Fsc = 0.13165 *** |
| Among populations within each species | 22 | 2388.915 | 11.07451 Vb | 4.26 | Fst = 0.71915 *** |
| Among individuals within each of the 39 populations | 131 | 9569.124 | 73.04675 Vc | 28.09 | Fct = 0.67657 *** |
| Total | 169 | 42,006.594 | 260.08805 | 100 |
| Species | Number of Samples | Number of Endophyte + Samples | Endophyte Concn. (Mean pg/ng ± S.E.) | Swainsonine (Mean % ± S.E.) | Chemotype |
|---|---|---|---|---|---|
| O. glacialis | 5 | 1 | 0.03 ± 0.03 | 1.1900 ± 0.3278 | Chemotype I |
| O. deflexa | 9 | 9 | 0.99 ± 0.14 | 0.3267 ± 0.0788 | Chemotype I |
| O. sericopetala | 9 | 9 | 1.00 ± 0.05 | 0.2140 ± 0.0445 | Chemotype I |
| O. falcata | 15 | 13 | 0.60 ± 0.08 | 0.1643 ± 0.0423 | Chemotype I |
| O. glabra | 10 | 5 | 0.51 ± 0.17 | 0.0668 ± 0.0164 | Intermediate |
| O. ochrocephala | 13 | 9 | 0.26 ± 0.09 | 0.0479 ± 0.0177 | Intermediate |
| O. kansuensis | 22 | 6 | 0.06 ± 0.02 | 0.0145 ± 0.0042 | Intermediate |
| O. giraldii | 14 | 4 | 0.22 ± 0.11 | 0.0030 ± 0.0019 | Chemotype II |
| O. ochrantha | 5 | 3 | 0.10 ± 0.04 | 0.0022 ± 0.0012 | Chemotype II |
| O. latibracteata | 10 | n.d. | n.d. | 0.0022 ± 0.0008 | Chemotype II |
| O. melanocalyx | 10 | n.d. | n.d. | 0.0021 ± 0.0014 | Chemotype II |
| O. myriophylla | 10 | n.d. | n.d. | 0.0015 ± 0.0007 | Chemotype II |
| O. merkensis | 4 | n.d. | n.d. | 0.0010 ± 0.0005 | Chemotype II |
| O. psamocharis | 10 | n.d. | n.d. | 0.0008 ± 0.0001 | Chemotype II |
| O. bicolor | 6 | n.d. | n.d. | 0.0007 ± 0.0001 | Chemotype II |
| O. ramosissima | 10 | n.d. | n.d. | 0.0007 ± 0.0001 | Chemotype II |
| O. aciphylla | 8 | n.d. | n.d. | 0.0006 ± 0.0001 | Chemotype II |
| Nucleotide Diversity (PI) | Endophyte (pg/ng) | Swainsonine (%) | |
|---|---|---|---|
| Annual average precipitation (mm) | −0.062 | −0.090 | −0.077 |
| Annual average air pressure (hpa) | −0.004 | −0.004 | −0.196 |
| Annual average wind speed (m/s) | 0.037 | −0.004 | 0.063 |
| Annual average temperature (°C) | −0.146 | 0.301 * | −0.038 |
| Annual average water vapor pressure (hpa) | −0.115 | 0.072 | −0.286 * |
| Annual relative humidity (%) | −0.012 | −0.322 * | −0.433 ** |
| Annual sunlight percentage (%) | 0.087 | 0.213 | 0.363 ** |
| Altitude (m) | 0.036 | −0.060 | 0.341 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, C.; Zhang, L.; Zhao, Q.; Beckmann, M.; Phillips, H.; Meng, H.; Mo, C.; Mur, L.A.J.; He, W. Host-Species Variation and Environment Influence Endophyte Symbiosis and Mycotoxin Levels in Chinese Oxytropis Species. Toxins 2022, 14, 181. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14030181
Guo C, Zhang L, Zhao Q, Beckmann M, Phillips H, Meng H, Mo C, Mur LAJ, He W. Host-Species Variation and Environment Influence Endophyte Symbiosis and Mycotoxin Levels in Chinese Oxytropis Species. Toxins. 2022; 14(3):181. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14030181
Chicago/Turabian StyleGuo, Chenchen, Li Zhang, Qianqian Zhao, Manfred Beckmann, Helen Phillips, Huizhen Meng, Chonghui Mo, Luis A. J. Mur, and Wei He. 2022. "Host-Species Variation and Environment Influence Endophyte Symbiosis and Mycotoxin Levels in Chinese Oxytropis Species" Toxins 14, no. 3: 181. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14030181