Discovery of the Relationship between Distribution and Aflatoxin Production Capacity of Aspergillusspecies and Soil Types in Peanut Planting Areas

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Isolation and Verification of Aspergillus Strains from Two Types of Soil
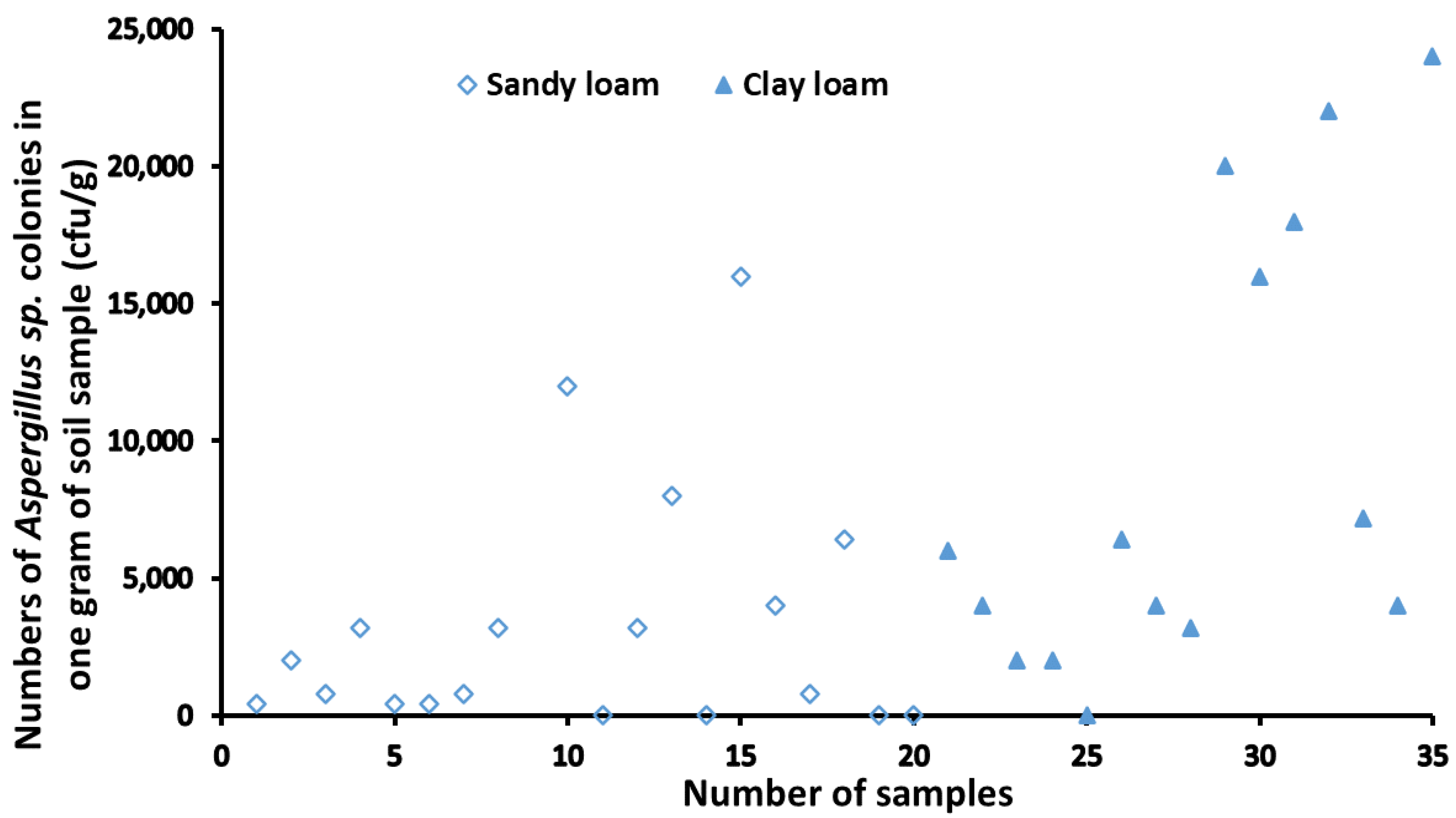
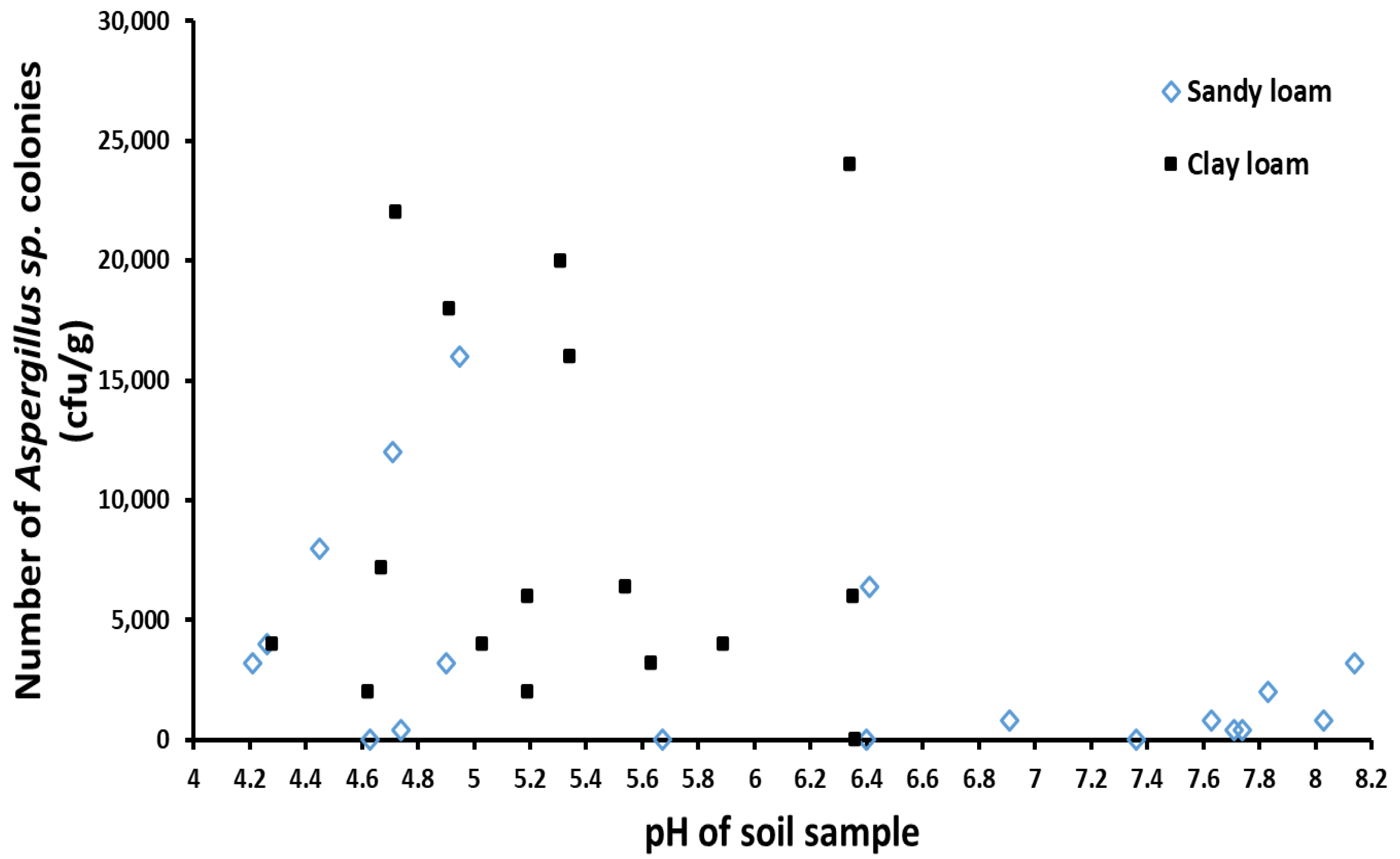
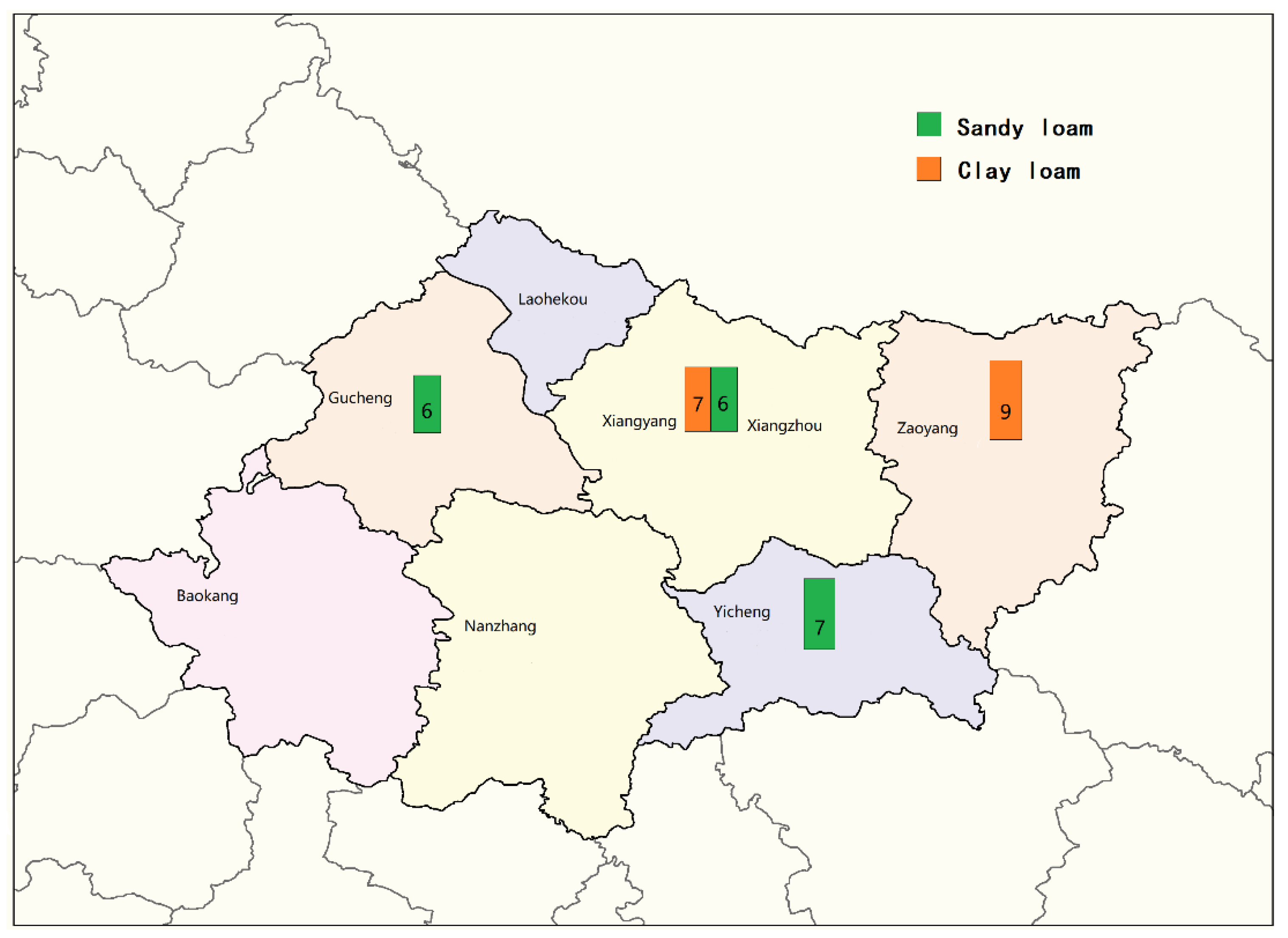
2.2. Relationship between Soil Types and Distribution of Aspergillus Isolates
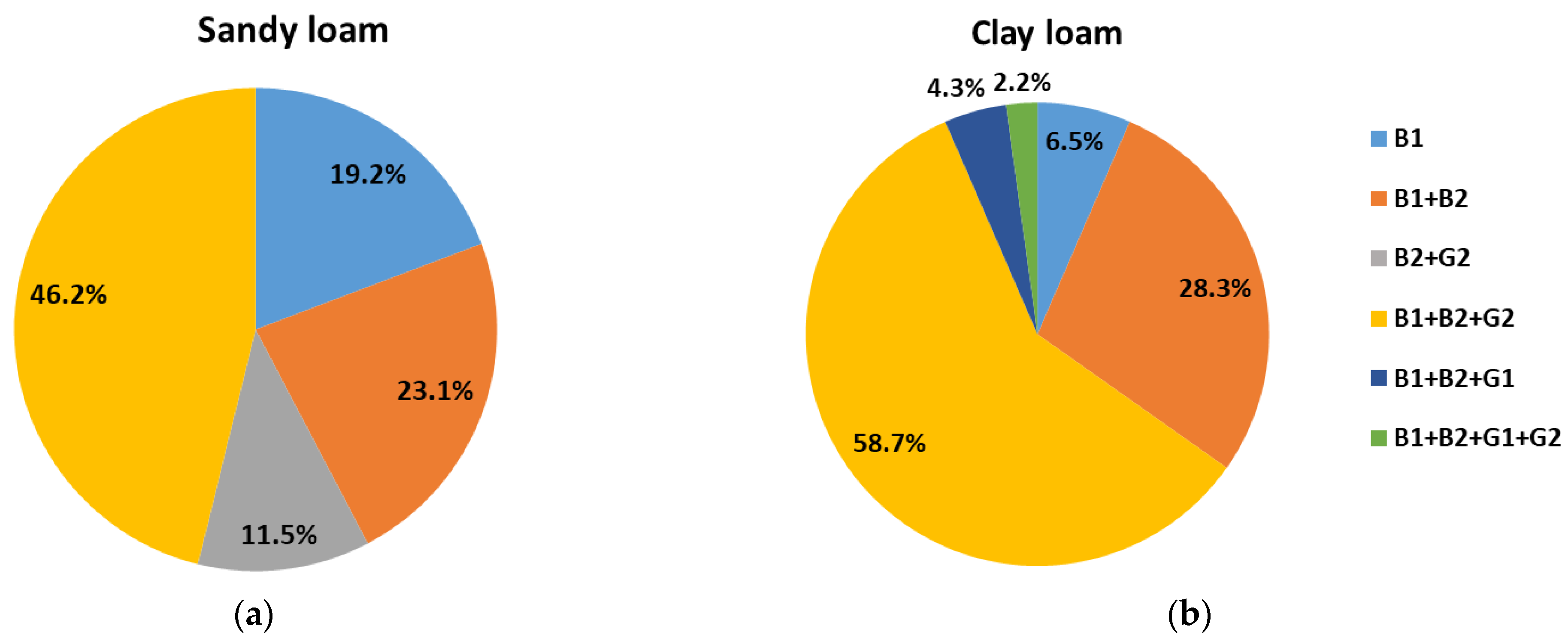
2.3. Aflatoxin-Producing Types in Different Soil Types
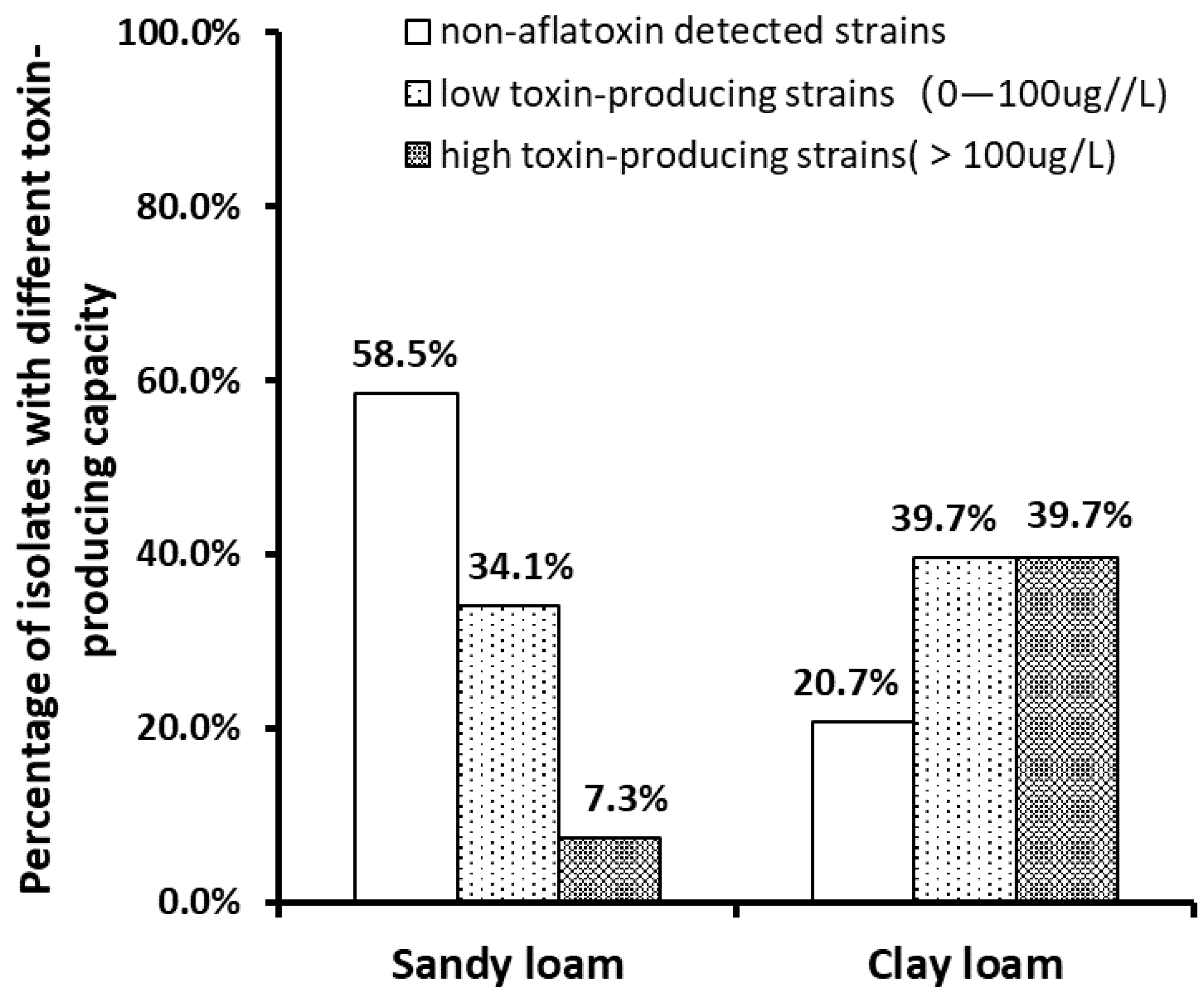
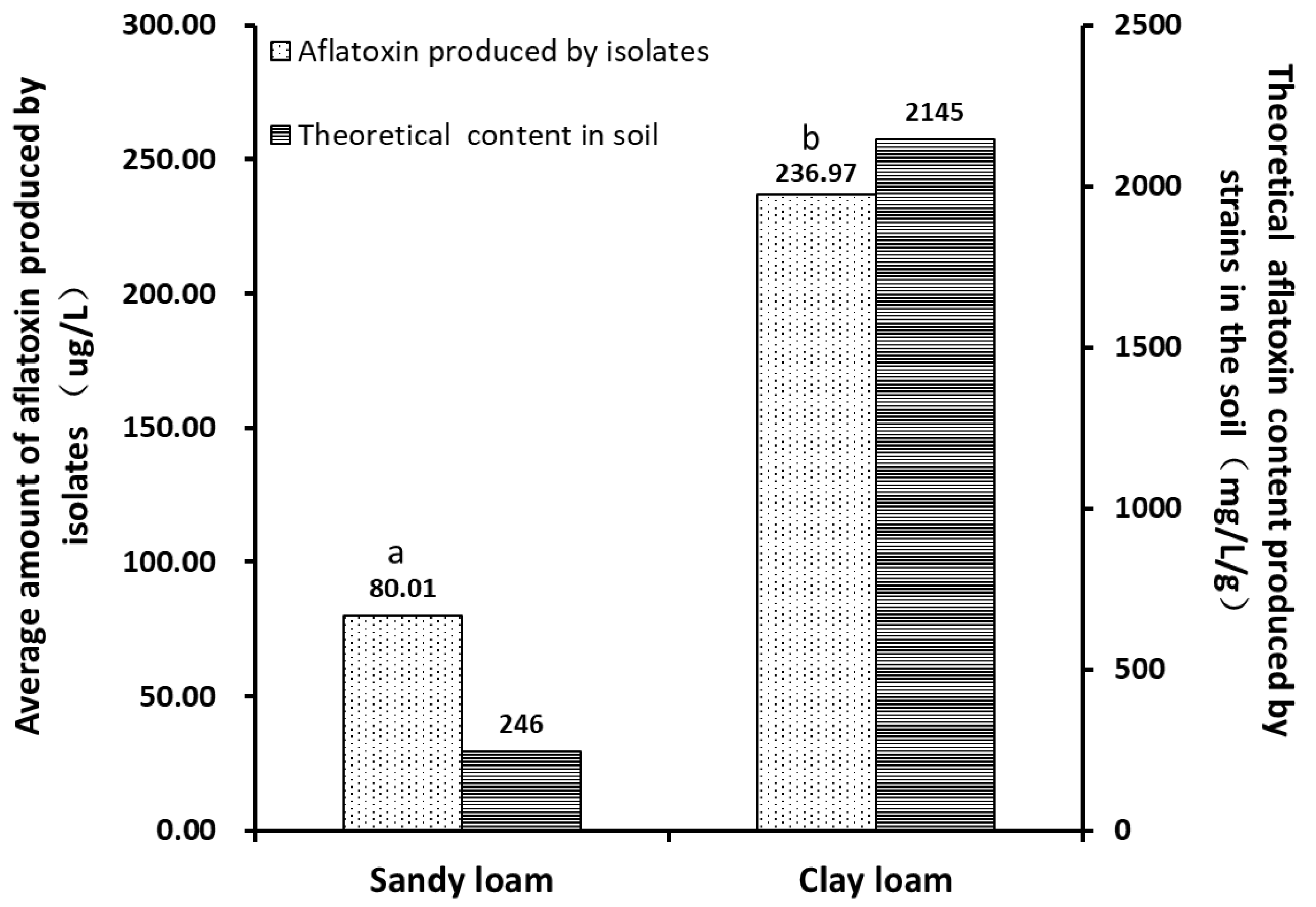
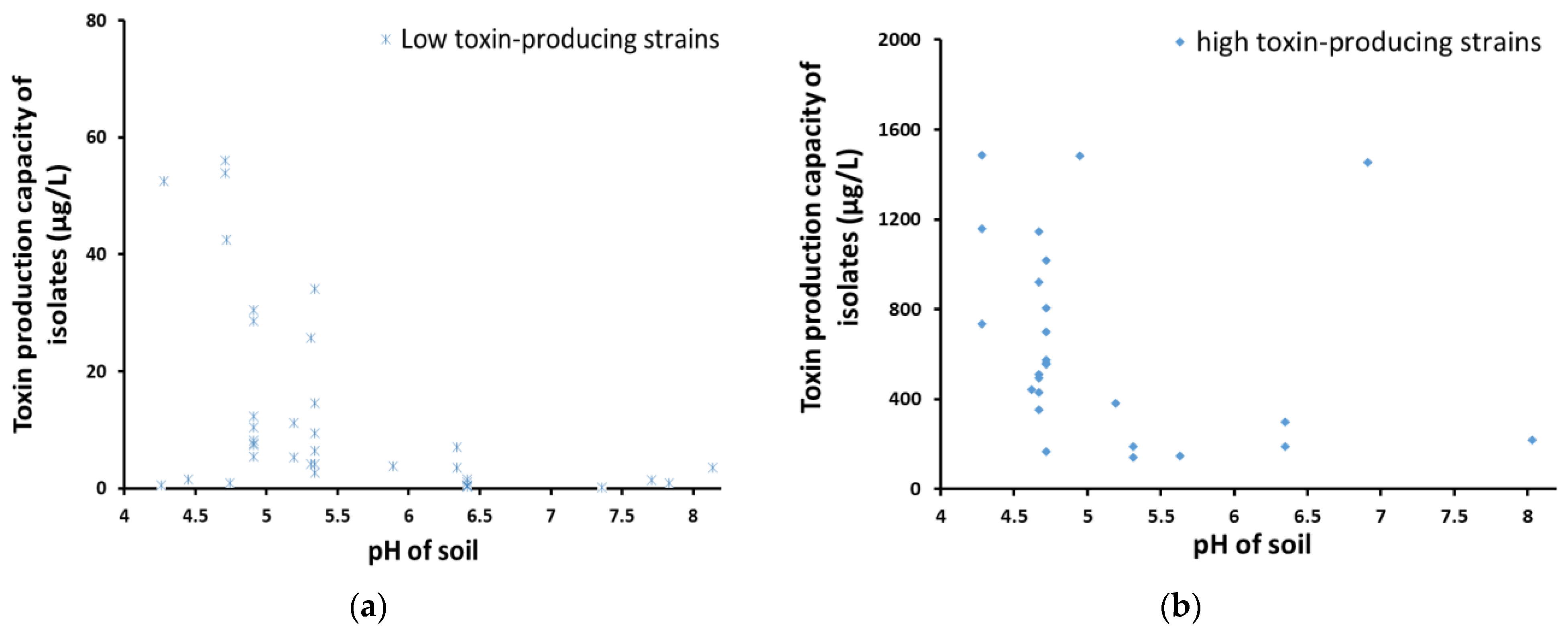
2.4. Aflatoxin Production Capacity of Aspergillus Isolates in Different Types of Soils
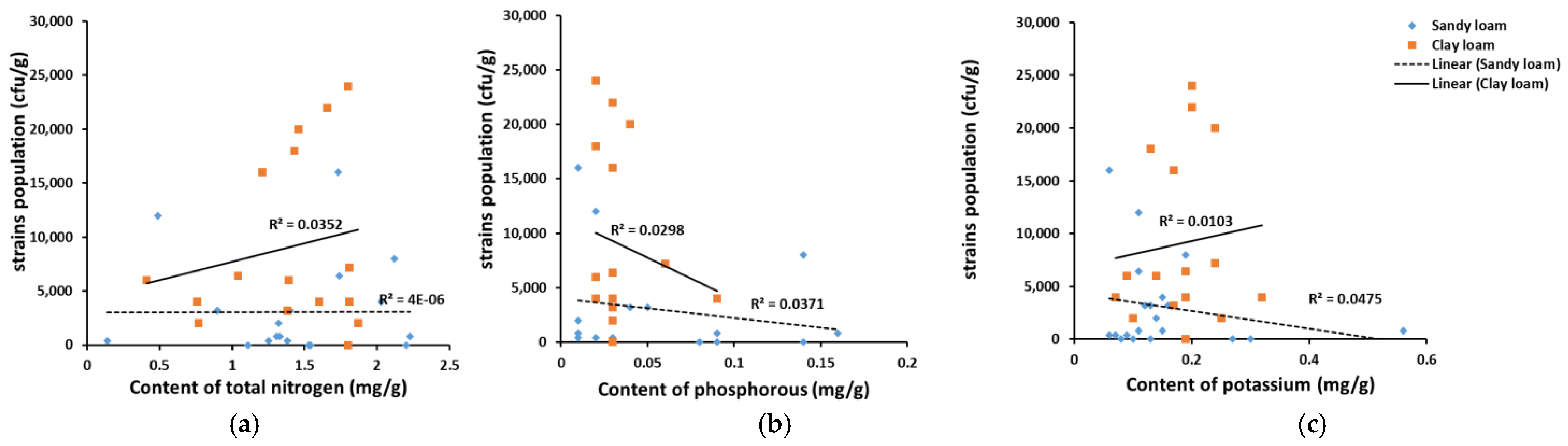
2.5. The Relationship between Soil Properties and the Number of Aspergillus sp. Colonies and Toxin Production Capacity
3. Discussion
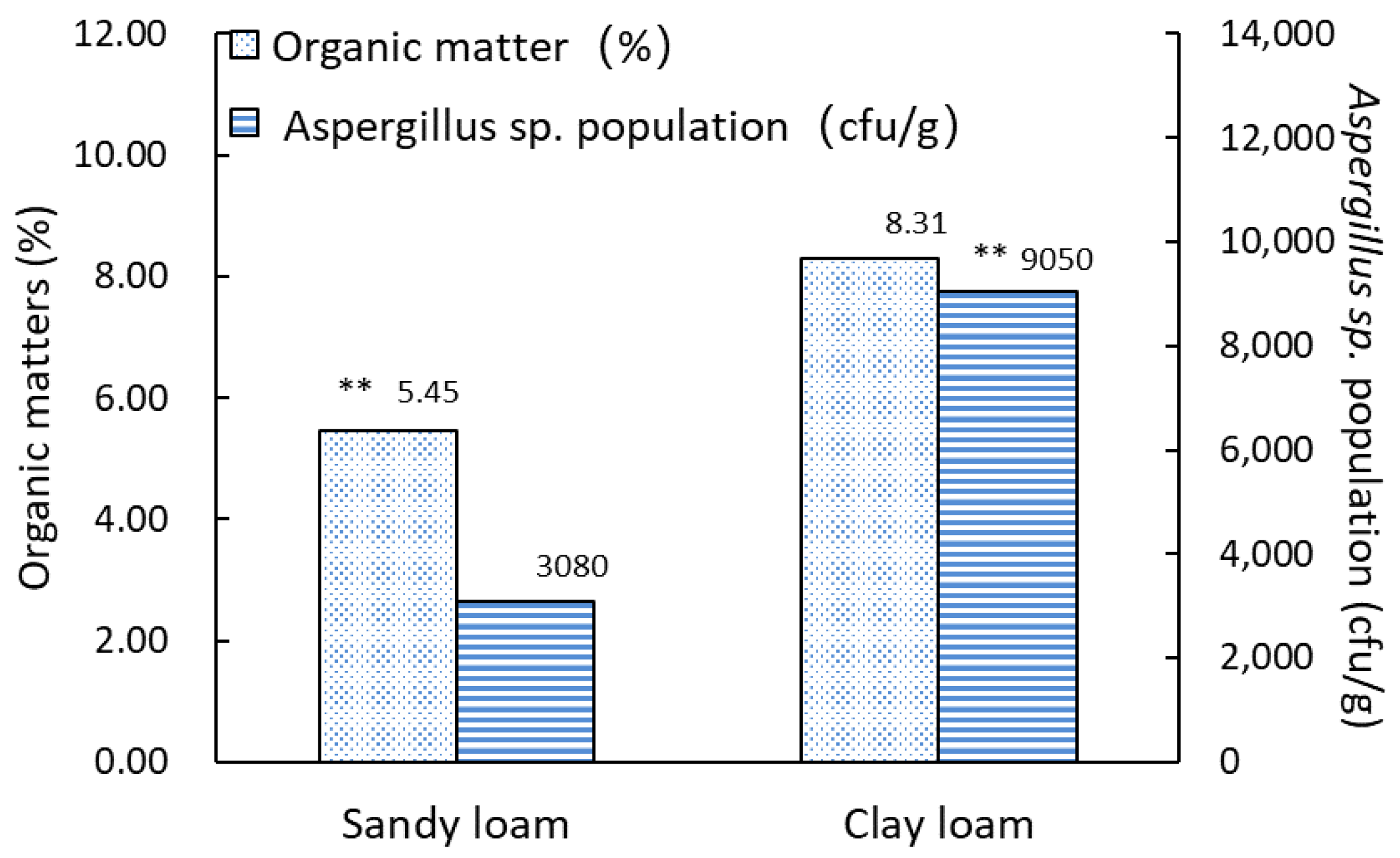
3.1. Characteristics of Aspergillus Isolates Differ in Two Typical Soil Types
3.2. The Properties of Soil May Affect the Number of Colonies and the Formation of Strain Group Structure
4. Conclusions
5. Materials and Methods
5.1. Materials
5.1.1. Reagents
5.1.2. Instruments
5.1.3. Samples
5.2. Methods
5.2.1. Isolation and Identification of A. flavus
5.2.2. Aflatoxin-Producing Culture
5.2.3. Detection of Aflatoxin
5.2.4. Calculation
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yan’e, S.H.A.N.G.; Weimin, Y.A.N.G. Variation analysis of cereals mycotoxin limit standards of CAC, EU, USA and China. J. Food Sci. Technol. 2019, 37, 10–15. [Google Scholar]
- Xing, F.; Ding, N.; Liu, X.; Selvaraj, J.N.; Wang, L.; Zhou, L.; Zhao, Y.; Wang, Y.; Liu, Y. Variation in fungal microbiome (mycobiome) and aflatoxins during simulated storage of in-shell peanuts and peanut kernels. Sci. Rep. 2016, 6, 25930. [Google Scholar] [CrossRef] [PubMed]
- Pankaj, S.K.; Shi, H.; Keener, K.M. A review of novel physical and chemical decontamination technologies for aflatoxin in food. Trends Food Sci. Technol. 2018, 71, 73–83. [Google Scholar] [CrossRef]
- Valencia-Quintana, R.; Milić, M.; Jakšić, D.; Šegvić Klarić, M.; Tenorio-Arvide, M.G.; Pérez-Flores, G.A.; Bonassi, S.; Sánchez-Alarcón, J. Environment Changes, Aflatoxins, and Health Issues, a Review. Int. J. Environ. Res. Public Health 2020, 17, 7850. [Google Scholar] [CrossRef]
- Benkerroum, N. Mycotoxins in dairy products: A review. Int. Dairy J. 2016, 62, 63–75. [Google Scholar] [CrossRef]
- Milićević, D.R.; Škrinjar, M.; Baltić, T. Real and Perceived Risks for Mycotoxin Contamination in Foods and Feeds: Challenges for Food Safety Control. Toxins 2010, 2, 572–592. [Google Scholar] [CrossRef] [Green Version]
- Xianfeng, R. Simultaneous Detection and Trichoderma’s Biocontrol on Aspergillus flavus and Aflatoxin in Grains and Oil. Ph.D. Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2020. [Google Scholar]
- Xu, Y.; Liang, C.; Lei, Z.; Lin, X.; Wang, D. Research progress on detection and biological prevention of aflatioxins. J. Anhui Agric. Sci. 2007, 32, 10210–10228. [Google Scholar]
- Zhu, T.; Yue, X.; Bai, Y.; Ding, X.; Li, P.; Zhang, Q.; Zhang, W. Distribution, aflatoxinproduction of Aspergillus flavus in soils of typical peanuts planting areas in Hubei province. Chin. J. Oil Crop Sci. 2019, 41, 255–260. [Google Scholar]
- Chushu, Z. Study on Distribution, Aflatoxin Production and Genetic Diversity of Aspergillus flavus in Soils of Peanut Fields in Four Agroecological Zones of China. Ph.D. Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2013. [Google Scholar]
- Xing, Z. Study on Distribution, Toxigenicity and Infection of Aspergillus flavus in Typical Peanuts Producing Areas in China. Master’s Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2019. [Google Scholar]
- Houmiao, W. Mechanism of Resistance to Aflaitoxin Production in Peanuts. Ph.D. Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2016. [Google Scholar]
- Khan, R.; Ghazali, F.M.; Mahyudin, N.A.; Samsudin, N.I.P. Biocontrol of Aflatoxins Using Non-Aflatoxigenic Aspergillus flavus: A Literature Review. J. Fungi 2021, 7, 381. [Google Scholar] [CrossRef]
- Jaime-Garcia, R.; Cotty, P.J. Crop rotation and soil temperature influence the community structure of Aspergillus flavus in soil. Soil Biol. Biochem. 2010, 42, 1842–1847. [Google Scholar] [CrossRef]
- Abbas, H.; Wilkinson, J.; Zablotowicz, R.; Accinelli, C.; Abel, C.; Bruns, H.; Weaver, M. Ecology of Aspergillus flavus, regulation of aflatoxin production, and management strategies to reduce aflatoxin contamination of corn. Toxin Rev. 2009, 28, 142–153. [Google Scholar] [CrossRef]
- Dorner, J.W. Management and prevention of mycotoxins in peanuts. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2008, 25, 203–208. [Google Scholar] [CrossRef]
- Horn, B.W.; Dorner, J.W. Regional Differences in Production of Aflatoxin B1 and Cyclopiazonic Acid by Soil Isolates of Aspergillus flavus along a Transect within the United States. Appl. Environ. Microbiol. 1999, 65, 1444–1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran-Dinh, N.; Kennedy, I.; Bui, T.; Carter, D. Survey of Vietnamese Peanuts, Corn and Soil for the Presence of Aspergillus flavus and Aspergillus parasiticus. Mycopathologia 2009, 168, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Linxia, W. Researches on Risk Prediction Model of Aflatoxin Contamination in China. Ph.D. Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2019. [Google Scholar]
- Zhang, C.; Selvaraj, J.N.; Yang, Q.; Liu, Y. A Survey of Aflatoxin-Producing Aspergillus sp. from Peanut Field Soils in Four Agroecological Zones of China. Toxins 2017, 9, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaime-Garcia, R.; Cotty, P.J. Aspergillus flavus in Soils and Corncobs in South Texas: Implications for Management of Aflatoxins in Corn-Cotton Rotations. Plant Dis. 2004, 88, 1366–1371. [Google Scholar] [CrossRef] [Green Version]
- Proctor, R.H. Fusarium toxins: Trichothecenes and fumonisins. In Microbial Foodborne Diseases: Mechanisms of Pathogenesis and Toxin Synthesis; Cary, J.W., Linz, J.E., Bhatnagar, D., Eds.; Technomic Publishing Co., Inc.: Lancaster, PA, USA, 2000; pp. 368–381. [Google Scholar]
- Bhatnagar, D.; Ehrlich, K.C.; Cleveland, T.E. Molecular genetic analysis and regulation of aflatoxin biosynthesis. Appl. Microbiol. Biotechnol. 2003, 61, 83–93. [Google Scholar] [CrossRef]
- Zablotowicz, R.M.; Abbas, H.K.; Locke, M.A. Population ecology of Aspergillus flavus associated with Mississippi Delta soils. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2007, 24, 1102–1108. [Google Scholar] [CrossRef]
- Wicklow, D.; Donahue, J.E. Sporogenic germination of sclerotia in Aspergillus flavus and A. parasiticus. Trans. Br. Mycol. Soc. 1984, 82, 621–624. [Google Scholar] [CrossRef]
- Winter, G.; Pereg, L. A review on the relation between soil and mycotoxins: Effect of aflatoxin on field, food and finance. Eur. J. Soil Sci. 2019, 70, 882–897. [Google Scholar] [CrossRef]
- Dadzie, M.A.; Oppong, A.; Ofori, K.; Eleblu, J.S.Y.; Ifie, B.E.; Blay, E.T.; Obeng-Bio, E.; Appiah-Kubi, Z.; Warburton, M.L. Distribution and genetic diversity among Aspergillus flavus isolates across three agro-ecologies essential for maize cultivation in Ghana. Plant Pathol. 2019, 68, 1565–1576. [Google Scholar] [CrossRef]
- Luo, Z.; Qin, Y.; Xu, Y.; Xu, T. Recent progress in the biosynthesis, metabolism and toxincity of aflatoxins. Food Sci. 2015, 36, 250–257. [Google Scholar]
- Mehan, V.K.; Mayee, C.D.; Jayanthi, S.; McDonald, D. Preharvest seed infection by Aspergillus flavus group fungi and subsequent aflatoxin contamination in groundnuts in relation to soil types. Plant Soil 1991, 136, 239–248. [Google Scholar] [CrossRef]
- Torres, A.M.; Barros, G.G.; Palacios, S.A.; Chulze, S.N.; Battilani, P. Review on pre- and post-harvest management of peanuts to minimize aflatoxin contamination. Food Res. Int. 2014, 62, 11–19. [Google Scholar] [CrossRef]
- Windham, G.L.; Williams, W.P.; Hawkins, L.K.; Brooks, T.D. Effect of Aspergillus flavus inoculation methods and environmental conditions on aflatoxin accumulation in corn hybrids. Toxin Rev. 2009, 28, 70–78. [Google Scholar] [CrossRef]
- Zhang, X.; Yue, X.; Ding, X.; Li, P.; Yu, Q.; Xie, H.; Zhang, Q.; Zhang, W. Distribution and aflatoxin contamination by Aspergillus flavus in peanut from the southwest China. Chin. J. Oil Crop Sci. 2019, 41, 773–780. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Type | Quantity of Soil Sample | Range of Colony Count (cfu/g) | Average of Colony Count (cfu/g) | Standard Deviation (SD) |
|---|---|---|---|---|
| Sandy loam | 19 | 0–12,000 | 3080 | 4403 |
| Clay loam | 16 | 0–24,000 | 9050 | 8003 |
| p value | 0.0074 ** |
| Soil Types | Numbers of Strains | The Average Amount of Aflatoxin Produced by the Strains (μg/L) | The Range of the Amount of Aflatoxin Produced by the Strains (μg/L) | The Theoretical Amount of Aflatoxin Produced in the Soil (mg/L/g) | ||
|---|---|---|---|---|---|---|
| Non-Aflatoxin Detected Strains | Toxin-Producing Strains | Average | SD | |||
| Sandy loam | 24 | 17 | 80.01 | 320.53 | 0–1482.81 | 246.44 |
| Clay loam | 12 | 46 | 236.97 | 336.09 | 0–1485.16 | 2144.58 |
| p values | 0.029 * | |||||
| Soil Type | Organic Matter (%) | Total Nitrogen (mg/g) | Phosphorous (mg/g) | Potassium (mg/g) | pH | Average of Colony Count (cfu/g) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Average | SD | Average | SD | Average | SD | Average | SD | Average | SD | Average | SD | |
| Sandy loam | 5.45 | 1.71 | 1.45 | 0.53 | 0.05 | 0.05 | 0.15 | 0.11 | 6.07 | 1.48 | 3080 | 4403 |
| Clay loam | 8.31 | 0.86 | 1.39 | 0.44 | 0.03 | 0.02 | 0.18 | 0.07 | 5.34 | 0.65 | 9050 | 8003 |
| p value | 0.0001 ** | 0.7726 | 0.1331 | 0.4205 | 0.0784 | 0.0074 ** | ||||||
| pH of Soils | Numbers of Strains | Number of Colonies | The Average Amount of Aflatoxin Produced by the Strains (μg/L) | The Range of the Amount of Aflatoxin Produced by the Strains (μg/L) | |||
|---|---|---|---|---|---|---|---|
| Non-Aflatoxin Detected Strains | Toxin-Producing Strains | Average | SD | Average | SD | ||
| <7 (weakly acidic) | 34 | 58 | 7100 | 6885 | 182.6 | 363.5 | 0–1485.16 |
| >7 (weakly alkaline) | 2 | 5 | 1086 | 1124 | 32.3 | 82.7 | 0–219.8 |
| p value | 0.015 * | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Wang, X.; Wang, D.; Chu, Q.; Zhang, Q.; Yue, X.; Zhu, M.; Dong, J.; Li, L.; Jiang, X.; et al. Discovery of the Relationship between Distribution and Aflatoxin Production Capacity of Aspergillusspecies and Soil Types in Peanut Planting Areas. Toxins 2022, 14, 425. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14070425
Zhang S, Wang X, Wang D, Chu Q, Zhang Q, Yue X, Zhu M, Dong J, Li L, Jiang X, et al. Discovery of the Relationship between Distribution and Aflatoxin Production Capacity of Aspergillusspecies and Soil Types in Peanut Planting Areas. Toxins. 2022; 14(7):425. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14070425
Chicago/Turabian StyleZhang, Shujuan, Xue Wang, Dun Wang, Qianmei Chu, Qian Zhang, Xiaofeng Yue, Mengjie Zhu, Jing Dong, Li Li, Xiangguo Jiang, and et al. 2022. "Discovery of the Relationship between Distribution and Aflatoxin Production Capacity of Aspergillusspecies and Soil Types in Peanut Planting Areas" Toxins 14, no. 7: 425. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14070425