Evaluation of the Susceptibility of Lotus Seeds (Nelumbo nucifera Gaertn.) to Aspergillus flavus Infection and Aflatoxin Contamination

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
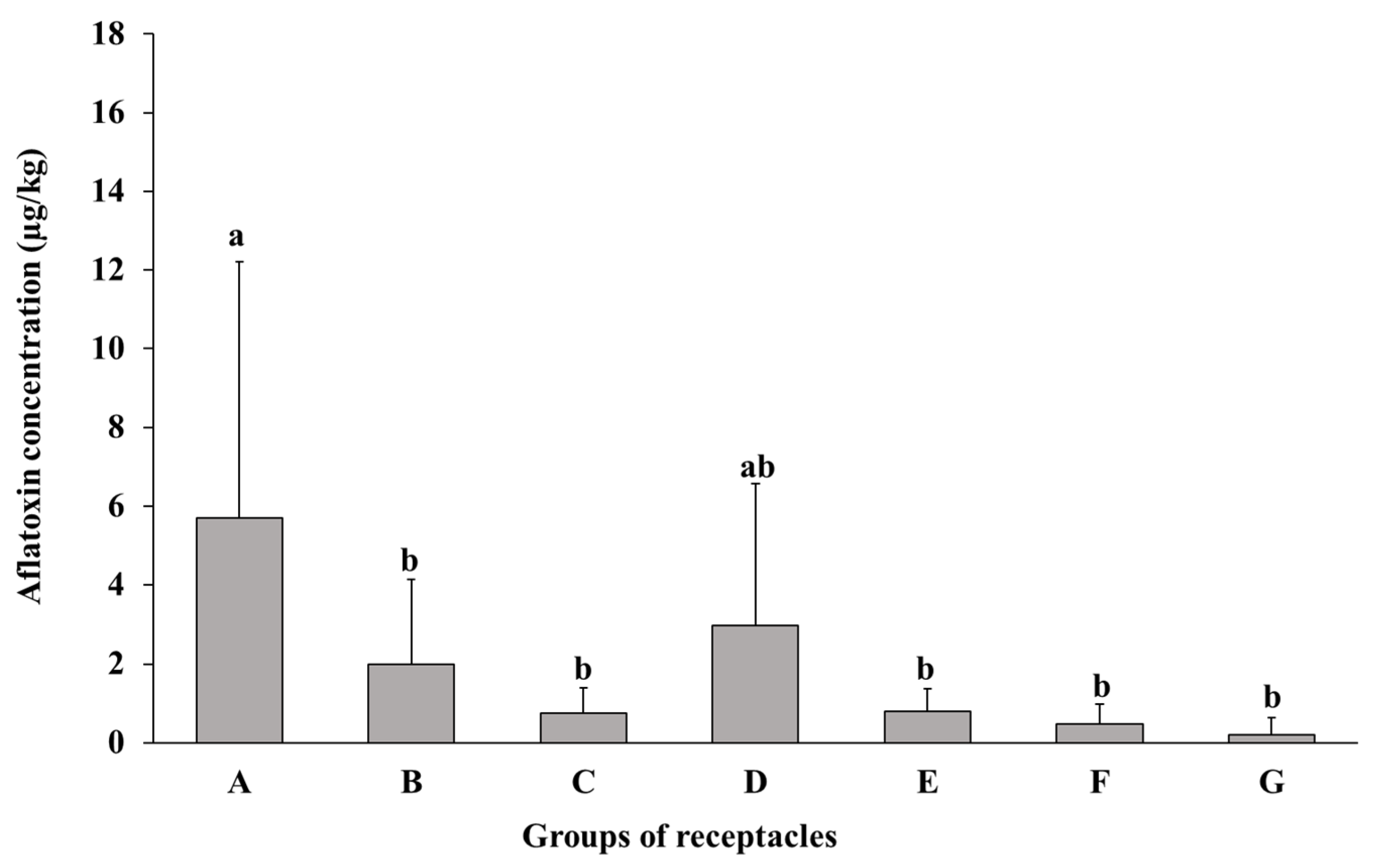
2.1. Susceptibility of Lotus Seeds to AF Contamination
2.1.1. Susceptibility of Whole Seeds at Different Maturity Stages to AF Contamination
2.1.2. Susceptibility of Different Parts of Lotus Seeds to AF Contamination
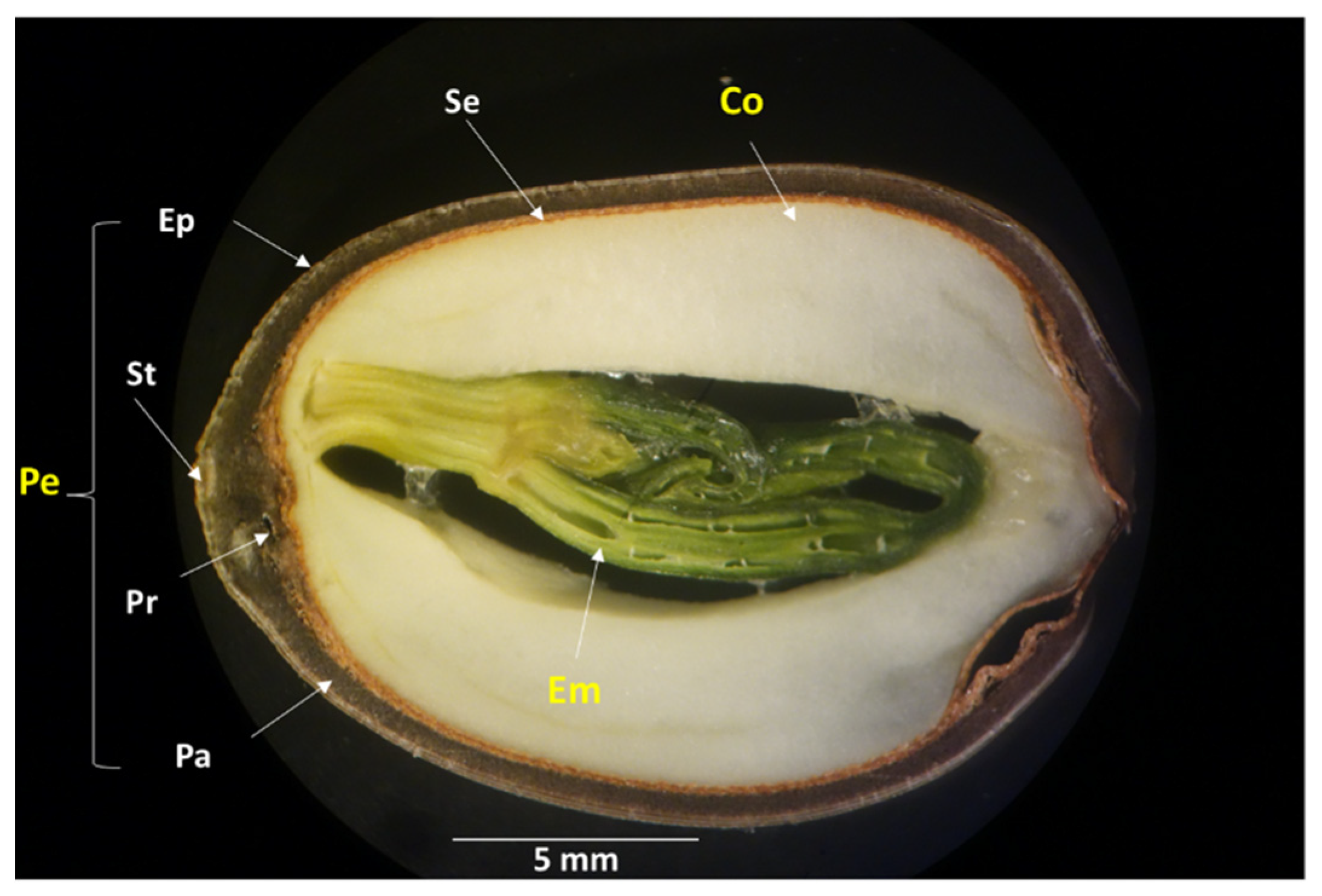
2.2. Changes in the Water-Gap Structure during Maturation
2.3. Path of Fungal Mycelial Penetration of Lotus Seeds
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Sample Preparation
5.2. AF Quantification using HPLC
5.3. Preparation of Longitudinal Sections
5.4. LM and SEM
5.5. Stereomicroscope (SM)
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.; Lu, X.; Zeng, S.; Huang, X.; Guo, Z.; Zheng, Y.; Tian, Y.; Zheng, B. Nutritional composition, physiological functions and processing of lotus (Nelumbo nucifera Gaertn.) seeds: A review. Phytochem. Rev. 2015, 14, 321–334. [Google Scholar] [CrossRef]
- Bangar, S.P.; Dunno, K.; Kumar, M.; Mostafa, H.; Maqsood, S. A comprehensive review on lotus seeds (Nelumbo nucifera Gaertn.): Nutritional composition, health-related bioactive properties, and industrial applications. J. Funct. Foods 2022, 89, 104937. [Google Scholar] [CrossRef]
- Wei, F.; Liu, X.; Liao, X.; Shi, L.; Zhang, S.; Lu, J.; Zhou, L.; Kong, W. Simultaneous determination of 19 mycotoxins in lotus seed using a multimycotoxin UFLC-MS/MS method. J. Pharm. Pharmacol. 2019, 71, 1172–1183. [Google Scholar] [CrossRef] [PubMed]
- Ashoka, S.; Revann, M.L. Physico chemical and functional properties of lotus (Nelumbo nucifera) Seed. Mysore J. Agric. Sci. 2022, 56, 61–67. [Google Scholar]
- Kakar, M.U.; Karim, H.; Shabir, G.; Iqbal, I.; Akram, M.; Ahmad, S.; Shafi, M.; Gul, P.; Riaz, S.; Ur-Rehman, R.; et al. A review on extraction, composition, structure, and biological activities of polysaccharides from different parts of Nelumbo nucifera. Food Sci Nutr. 2023, 11, 3655–3674. [Google Scholar] [CrossRef] [PubMed]
- Bhat, R.; Sridhar, K.R.; Karim, A.A. Microbial quality evaluation and effective decontamination of nutraceutically valued lotus seeds by electron beams and gamma irradiation. Radiat. Phys. Chem. 2010, 79, 976–981. [Google Scholar] [CrossRef]
- Liao, X.; Sun, C.; Wei, F.; Zhou, L.; Kong, W. Exploration of the safe water content and activity control points for medicinal and edible lotus seeds from mildew. AMB Expr. 2020, 10, 89. [Google Scholar] [CrossRef]
- Ałtyn, I.; Twarużek, M. Mycotoxin contamination concerns of herbs and medicinal plants. Toxins 2020, 12, 182. [Google Scholar] [CrossRef]
- Tulayakul, P.; Sugita-Konishi, Y. Mycotoxin contamination in foodstuffs and feeds-health concerns in Thailand. JJVR 2017, 65, 173–183. [Google Scholar]
- Namjoo, M.; Salamat, F.; Rajabli, N.; Hajihoseeini, R.; Niknejad, F.; Kohsar, F.; Joshaghani, H. Quantitative determination of aflatoxin by high performance liquid chromatography in wheat silos in Golestan province, north of Iran. Iran J. Public Health 2016, 45, 905–910. [Google Scholar]
- Alshannaq, A.F.; Yu, J.-H. A liquid chromatographic method for rapid and sensitive analysis of aflatoxins in laboratory fungal cultures. Toxins 2020, 12, 93. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Qiu, F.; Kong, W.; Wei, J.; Xiao, X.; Yang, M. Development and validation of an accurate and rapid LC-ESI-MS/MS method for the simultaneous quantification of aflatoxin B1, B2, G1 and G2 in lotus seeds. Food Control 2013, 29, 156–161. [Google Scholar] [CrossRef]
- Elamin, A.; Enomoto, H.; Watanabe, M.; Sakuda, S. The Mechanism of ochratoxin contamination of artificially inoculated licorice roots. Toxins 2023, 15, 219. [Google Scholar] [CrossRef] [PubMed]
- Baskin, J.M.; Baskin, C.C. Evolutionary consideration of claims of physical dormancy-break by microbial action and abrasion by soil particles. Seed Sci. Res. 2000, 10, 409–413. [Google Scholar] [CrossRef]
- Jayasuriya, K.M.G.; Baskin, J.M.; Geneve, R.L.; Baskin, C.C. Morphology and anatomy of physical dormancy in Ipomoea lacunosa: Identification of the water gap in seeds of Convolvulaceae (solanales). Ann. Bot. 2007, 100, 13–22. [Google Scholar] [CrossRef]
- Karaki, T.; Watanabe, Y.; Kondo, T.; Koike, T. Strophiole of seeds of the black locust acts as a water gap. Plant Species Biol. 2012, 27, 226–232. [Google Scholar] [CrossRef]
- Tajkarimi, M.; Shojaee, M.H.; Yazdanpanah, H.; Ibrahim, S.A. Aflatoxin in agricultural commodities and herbal medicine. In Aflatoxins—Biochemistry and Molecular Biology; Guevara-Gonzalez, R.G., Ed.; InTechOpen: Rijeka, Croatia, 2011. [Google Scholar]
- Chen, A.J.; Jiao, X.; Hu, Y.; Lu, X.; Gao, W. Mycobiota and mycotoxins in traditional medicinal seeds from China. Toxins 2015, 7, 3858–3875. [Google Scholar] [CrossRef]
- Gama-Arachchige, N.S.; Baskin, J.M.; Geneve, R.L.; Baskin, C.C. Identification and characterization of ten new water gaps in seeds and fruits with physical dormancy and classification of water-gap complexes. Ann. Bot. 2013, 112, 69–84. [Google Scholar] [CrossRef]
- Dam, S.; Laursen, B.S.; Ørnfelt, J.H.; Jochimsen, B.; Stærfeldt, H.H.; Friis, C.; Nielsen, K.; Goffard, N.; Besenbacher, S.; Krusell, L.; et al. The proteome of seed development in the model legume Lotus japonicus. Plant Physiol. 2009, 149, 1325–1340. [Google Scholar] [CrossRef]
- Jaganathan, G.K.; Song, D.; Liu, W.; Han, Y.; Liu, B. Relationship between seed moisture content and acquisition of impermeability in Nelumbo nucifera (Nelumbonaceae). Acta Bot. Bras. 2017, 31, 639–644. [Google Scholar] [CrossRef]
- Elamin, A.; Sakuda, S. Evaluation of the susceptibility of Ziziphus jujuba var. spinosa fruit to aflatoxin contamination and infection of aflatoxigenic fungus based on ripening stages and fruit parts. JSM Mycotoxins 2021, 71, 63–67. [Google Scholar] [CrossRef]
- Shen-Miller, J. Sacred lotus, the long-living fruits of China antique. Seed Sci. Res. 2002, 12, 131–143. [Google Scholar] [CrossRef]
- Zhang, D.; Liu, T.; Sheng, J.; Lv, S.; Ren, L. TMT-based quantitative proteomic analysis reveals the physiological regulatory networks of embryo dehydration protection in lotus (Nelumbo nucifera). Front. Plant Sci. 2021, 12, 792057. [Google Scholar] [CrossRef] [PubMed]
- Shen-Miller, J.; Aung, L.H.; Turek, J.; Schopf, J.W.; Tholandi, M.; Yang, M.; Czaj, A. Centuries-old viable fruit of sacred lotus Nelumbo nucifera Gaertn var. China antique. Trop. Plant Biol. 2013, 6, 53–68. [Google Scholar] [CrossRef]
- Moro, C.F.; Fukao, Y.; Shibato, J.; Rakwal, R.; Agrawal, G.K.; Shioda, S.; Kouzuma, K.; Yonekura, M. Immature seed endosperm and embryo proteomics of the lotus (Nelumbo Nucifera Gaertn.) by one-dimensional gel-based tandem mass spectrometry and a comparison with the mature endosperm proteome. Proteomes 2015, 3, 184–235. [Google Scholar] [CrossRef]
- Priyadarshini, E.; Tulpule, P.G. Effect of free fatty acids on aflatoxin production in a synthetic medium. Food Cosmet. Toxicol. 1980, 18, 367–369. [Google Scholar] [CrossRef]
- Uppala, S.S.; Bowen, K.L.; Woods, F.M. Pre-harvest aflatoxin contamination and soluble sugars of peanut. Peanut Sci. 2013, 40, 40–51. [Google Scholar] [CrossRef]
- Wang, L.; Fu, J.; Li, M.; Fragner, L.; Weckwerth, W.; Yang, P. Metabolomic and proteomic profiles reveal the dynamics of primary metabolism during seed development of lotus (Nelumbo nucifera). Front Plant Sci 2016, 7, 750. [Google Scholar] [CrossRef]
- Wu, J.-Z.; Zheng, Y.-B.; Chen, T.-Q.; Yi, J.; Qin, L.-P.; Rahman, K.; Lin, W.-X. Evaluation of the quality of lotus seed of Nelumbo nucifera Gaertn from outer space mutation. Food Chem. 2007, 105, 540–547. [Google Scholar] [CrossRef]
- Sathithon, P.; Yan-bin, X. Effect of sprouting on the chemical and nutritional qualities and phenolic alkaloid content of lotus (Nelumbo nucifera Gaertn.) seeds. Afr. J. Food Sci. 2012, 6, 204–211. [Google Scholar]
- Pandey, M.K.; Kumar, R.; Pandey, A.K.; Soni, P.; Gangurde, S.S.; Sudini, H.K.; Fountain, J.C.; Liao, B.; Desmae, H.; Okori, P.; et al. Mitigating aflatoxin contamination in groundnut through a combination of genetic resistance and post-harvest management practices. Toxins 2019, 11, 315. [Google Scholar] [CrossRef] [PubMed]
- Elamin, A.; Takatori, K.; Matsuda, Y.; Tsukada, M.; Kirino, F. Fungal biodeterioration of artificial aged linen textile: Evaluation by microscopic, spectroscopic and viscometric methods. MAA 2018, 18, 103–120. [Google Scholar]
- Enomoto, H. Adhesive film applications help to prepare strawberry fruit sections for desorption electrospray ionization-mass spectrometry imaging. Biosci. Biotechnol. Biochem. 2021, 85, 1341–1347. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elamin, A.; Sultana, S.; Sakuda, S. Evaluation of the Susceptibility of Lotus Seeds (Nelumbo nucifera Gaertn.) to Aspergillus flavus Infection and Aflatoxin Contamination. Toxins 2024, 16, 29. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins16010029
Elamin A, Sultana S, Sakuda S. Evaluation of the Susceptibility of Lotus Seeds (Nelumbo nucifera Gaertn.) to Aspergillus flavus Infection and Aflatoxin Contamination. Toxins. 2024; 16(1):29. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins16010029
Chicago/Turabian StyleElamin, Abdelrahman, Sharmin Sultana, and Shohei Sakuda. 2024. "Evaluation of the Susceptibility of Lotus Seeds (Nelumbo nucifera Gaertn.) to Aspergillus flavus Infection and Aflatoxin Contamination" Toxins 16, no. 1: 29. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins16010029