
Handling Detection Limits of Multiplex Lateral Flow Immunoassay by Choosing the Order of Binding Zones


Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Materials
2.2. Determination of Constants of Immune Interaction Using an Enzyme-Linked Immunosorbent Assay
2.3. Synthesis of AuNPs and Their Conjugation with GAMI
2.4. Preparation of the Test Strips
2.5. LFIA of Mycotoxins
2.6. Data Processing
3. Results and Discussion
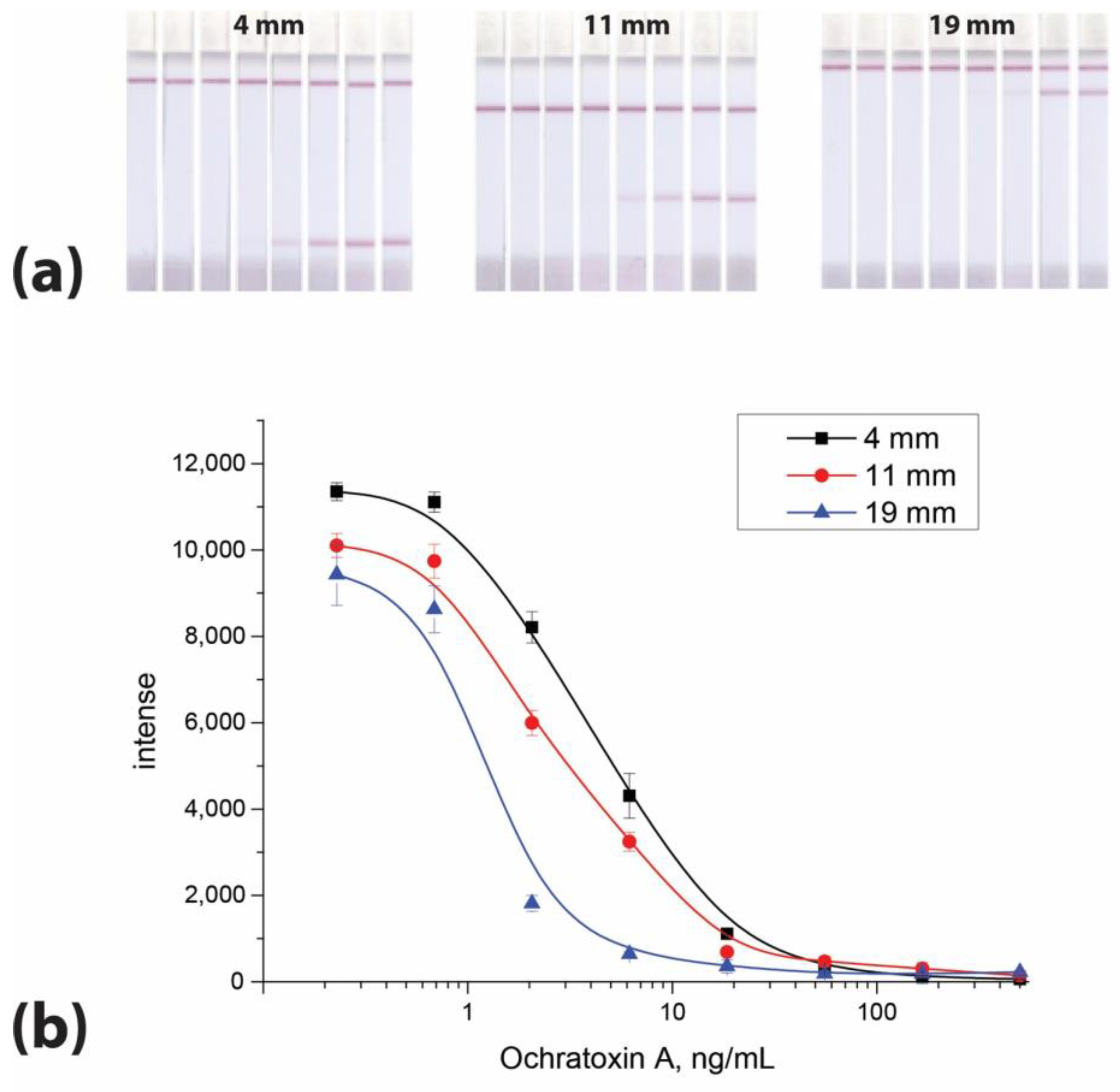
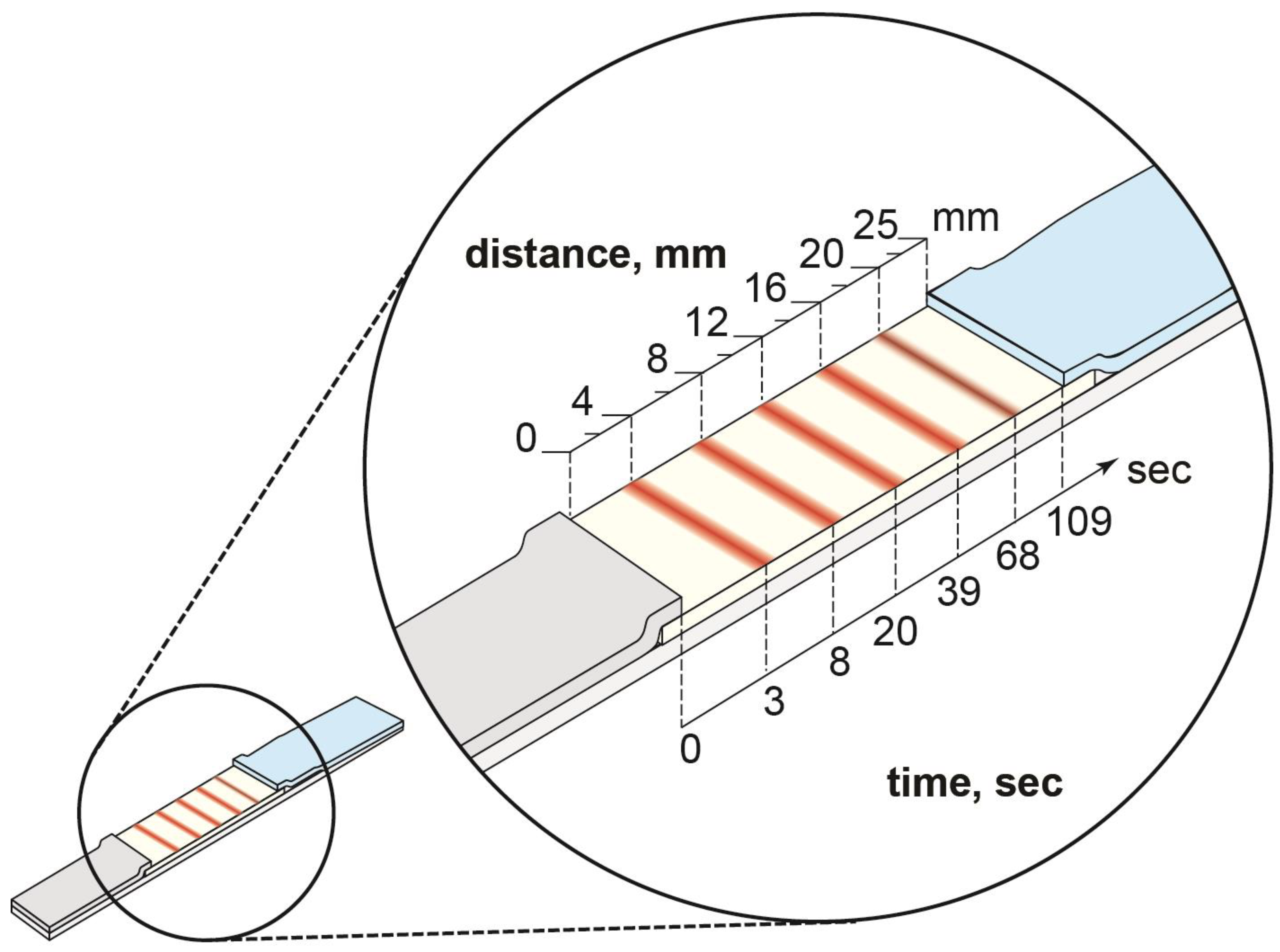
3.1. Dependence of Signal Intensity on the Location of the Binding Zone in the LFIA
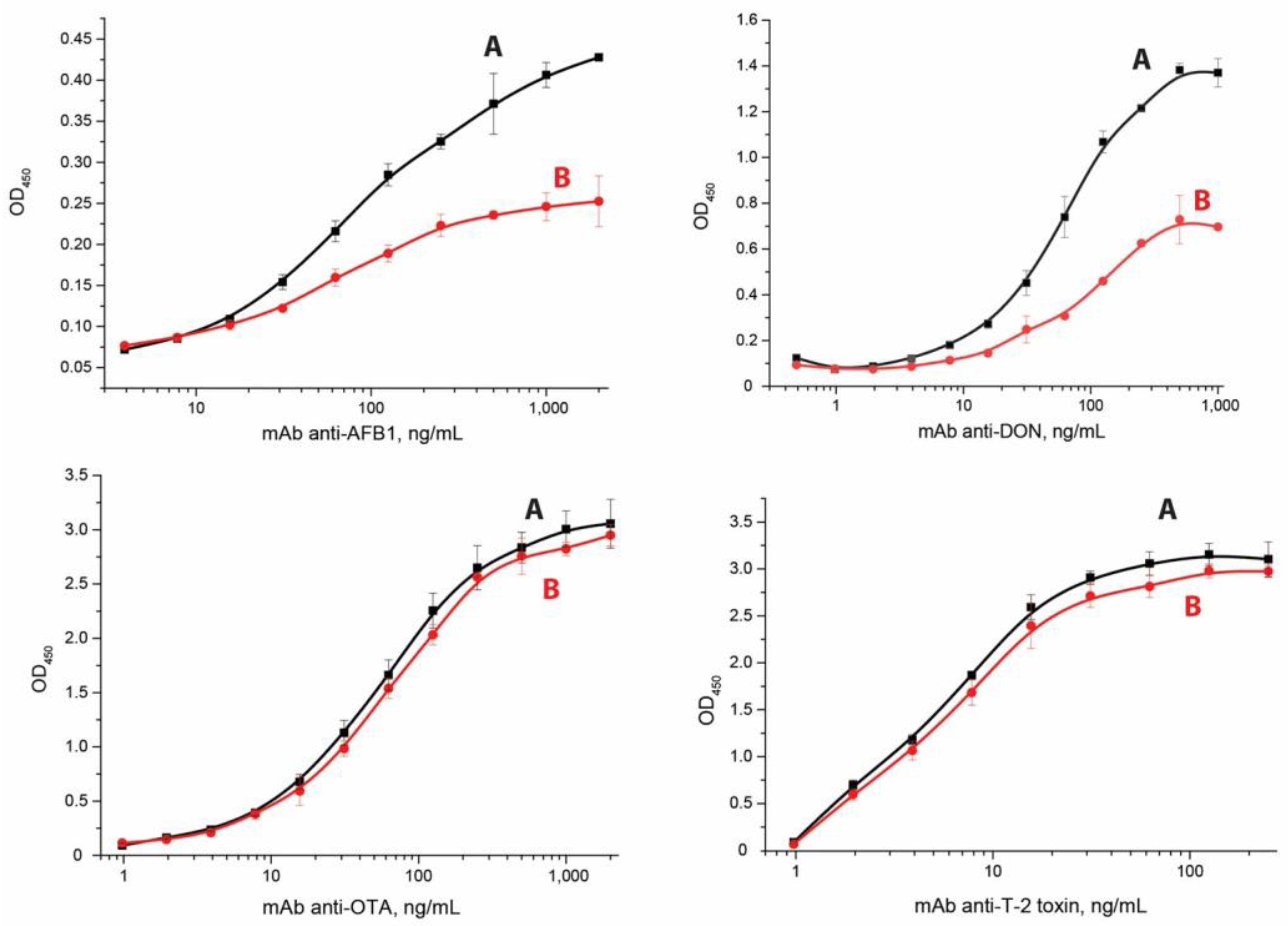
3.2. Determination of Equilibrium and Kinetic Constants
3.3. Factors Affecting Complexation in the Multiplex LFIA
3.4. Multiplex LFIA
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Parolo, C.; Sena-Torralba, A.; Bergua, J.F.; Calucho, E.; Fuentes-Chust, C.; Hu, L.; Rivas, L.; Álvarez-Diduk, R.; Nguyen, E.P.; Cinti, S.; et al. Tutorial: Design and fabrication of nanoparticle-based lateral-flow immunoassays. Nat. Protoc. 2020, 15, 3788–3816. [Google Scholar] [CrossRef]
- Boehringer, H.R.; O’Farrell, B.J. Lateral flow assays in infectious disease diagnosis. Clin. Chem. 2022, 68, 52–58. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, D.; Zhou, X.; Yu, Y.; Liu, J.; Hu, N.; Wang, H.; Li, G.; Wu, Y. Recent progress in the construction of nanozyme-based biosensors and their applications to food safety assay. Trends Anal. Chem. 2019, 121, 115668. [Google Scholar] [CrossRef]
- Di Nardo, F.; Chiarello, M.; Cavalera, S.; Baggiani, C.; Anfossi, L. Ten years of lateral flow immunoassay technique applications: Trends, challenges and future perspectives. Sensors 2021, 21, 5185. [Google Scholar] [CrossRef]
- Xing, K.-Y.; Shan, S.; Liu, D.-F.; Lai, W.-H. Recent advances of lateral flow immunoassay for mycotoxins detection. Trends Anal. Chem. 2020, 133, 116087. [Google Scholar] [CrossRef]
- Anfossi, L.; Di Nardo, F.; Cavalera, S.; Giovannoli, C.; Baggiani, C. Multiplex lateral flow immunoassay: An overview of strategies towards high-throughput point-of-need testing. Biosensors 2019, 9, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Liu, Y.; Wang, M.; Chen, Y.; Jiang, W. A multiplex immunochromatographic test using gold nanoparticles for the rapid and simultaneous detection of four nitrofuran metabolites in fish samples. Anal. Bioanal. Chem. 2018, 410, 223–233. [Google Scholar] [CrossRef]
- Liu, B.; Gong, H.; Wang, Y.; Zhang, X.; Li, P.; Qiu, Y.; Wang, L.; Hua, X.; Guo, Y.; Wang, M. A gold immunochromatographic assay for simultaneous detection of parathion and triazophos in agricultural products. Anal. Methods 2018, 10, 422–428. [Google Scholar] [CrossRef]
- Zangheri, M.; Di Nardo, F.; Anfossi, L.; Giovannoli, C.; Baggiani, C.; Roda, A.; Mirasoli, M. A multiplex chemiluminescent biosensor for type B-fumonisins and aflatoxin B1 quantitative detection in maize flour. Analyst 2015, 140, 358–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartosh, A.V.; Sotnikov, D.V.; Hendrickson, O.D.; Zherdev, A.V.; Dzantiev, B.B. Design of multiplex lateral flow tests: A case study for simultaneous detection of three antibiotics. Biosensors 2020, 10, 17. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Wang, Y.; Liu, L.; Kuang, H.; Li, A.; Xu, C. Multiplex lateral flow immunoassay for five antibiotics detection based on gold nanoparticle aggregations. RSC Adv. 2016, 6, 7798–7805. [Google Scholar] [CrossRef]
- Li, X.; Li, P.; Zhang, Q.; Li, R.; Zhang, W.; Zhang, Z.; Ding, X.; Tang, X. Multi-component immunochromatographic assay for simultaneous detection of aflatoxin B1, ochratoxin A and zearalenone in agro-food. Biosens. Bioelectron. 2013, 49, 426–432. [Google Scholar] [CrossRef] [PubMed]
- Cavalera, S.; Di Nardo, F.; Chiarello, M.; Serra, T.; Colitti, B.; Guiotto, C.; Fagioli, F.; Cagnazzo, C.; Denina, M.; Palazzo, A.; et al. Bacterial ligands as flexible and sensitive detectors in rapid tests for antibodies to SARS-CoV-2. Anal. Bioanal. Chem. 2022, 414, 5473–5482. [Google Scholar] [CrossRef]
- Lin, K.W.; Chang, Y.C. Use of the Taguchi method to optimize an immunodetection system for quantitative analysis of a rapid test. Diagnostics 2021, 11, 1179. [Google Scholar] [CrossRef]
- Rushing, B.R.; Selim, M.I. Aflatoxin B1: A review on metabolism, toxicity, occurrence in food, occupational exposure, and detoxification methods. Food Chem. Toxicol. 2019, 124, 81–100. [Google Scholar] [CrossRef] [PubMed]
- Sobrova, P.; Adam, V.; Vasatkova, A.; Beklova, M.; Zeman, L.; Kizek, R. Deoxynivalenol and its toxicity. Interdiscip. Toxicol. 2010, 3, 94–99. [Google Scholar] [CrossRef]
- Yang, X.; Liu, P.; Cui, Y.; Xiao, B.; Liu, M.; Song, M.; Huang, W.; Li, Y. Review of the reproductive toxicity of T-2 toxin. J. Agric. Food Chem. 2020, 68, 727–734. [Google Scholar] [CrossRef]
- Tao, Y.; Xie, S.; Xu, F.; Liu, A.; Wang, Y.; Chen, D.; Pan, Y.; Huang, L.; Peng, D.; Wang, X. Ochratoxin A: Toxicity, oxidative stress and metabolism. Food Chem. Toxicol. 2018, 112, 320–331. [Google Scholar] [CrossRef]
- Yao, X.; Wang, Z.; Dou, L.; Zhao, B.; He, Y.; Wang, J.; Sun, J.; Li, T.; Zhang, D. An innovative immunochromatography assay for highly sensitive detection of 17β-estradiol based on an indirect probe strategy. Sens. Actuators B Chem. 2019, 289, 48–55. [Google Scholar] [CrossRef]
- Li, G.; Xu, L.; Wu, W.; Wang, D.; Jiang, J.; Chen, X.; Zhang, W.; Poapolathep, S.; Poapolathep, A.; Zhang, Z.; et al. On-site ultrasensitive detection paper for multiclass chemical contaminants via universal bridge-antibody labeling: Mycotoxin and illegal additives in milk as an example. Anal. Chem. 2019, 91, 1968–1973. [Google Scholar] [CrossRef]
- Urusov, A.E.; Zherdev, A.V.; Dzantiev, B.B. Use of gold nanoparticle-labeled secondary antibodies to improve the sensitivity of an immunochromatographic assay for aflatoxin B1. Microchim. Acta 2014, 181, 1939–1946. [Google Scholar] [CrossRef]
- Petrakova, A.V.; Urusov, A.E.; Zherdev, A.V.; Dzantiev, B.B. Gold nanoparticles of different shape for bicolor lateral flow test. Anal. Biochem. 2019, 568, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Urusov, A.E.; Petrakova, A.V.; Zherdev, A.V.; Dzantiev, B.B. “Multistage in one touch” design with a universal labelling conjugate for high-sensitive lateral flow immunoassays. Biosens. Bioelectron. 2016, 86, 575–579. [Google Scholar] [CrossRef] [PubMed]
- Karakus, C.; Salih, B.A. Comparison of the lateral flow immunoassays (LFIA) for the diagnosis of Helicobacter pylori infection. J. Immunol. Methods 2013, 396, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Urusov, A.E.; Petrakova, A.V.; Zherdev, A.V.; Zvereva, E.A. Indirect labeling of antibodies as a universal approach to increase sensitivity of lateral flow tests: A case study for mycotoxins detection. Open Biotechnol. J. 2019, 13, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Byzova, N.A.; Zvereva, E.A.; Zherdev, A.V.; Eremin, S.A.; Sveshnikov, P.G.; Dzantiev, B. Pretreatment-free immunochromatographic assay for the detection of streptomycin and its application to the control of milk and dairy products. Anal. Chim. Acta 2011, 701, 209–217. [Google Scholar] [CrossRef]
- Berlina, A.N.; Bartosh, A.V.; Zherdev, A.V.; Xu, C.; Dzantiev, B.B. Development of immunochromatographic assay for determination of tetracycline in human serum. Antibiotics 2018, 7, 99. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Meng, X.; Zhu, Y.; Shen, M.; Lu, Y.; Cheng, J.; Xu, Y. Rapid detection of four mycotoxins in corn using a microfluidics and microarray-based immunoassay system. Talanta 2018, 186, 299–305. [Google Scholar] [CrossRef]
- Tijssen, P. Practice and Theory of Enzyme Immunoassays; Elsevier Science: Amsterdam, The Netherlands, 1985; pp. 167–172. [Google Scholar]
- EMD Millipore Corporation. Rapid Lateral Flow Test Strips: Considerations for Product Development; EMD Millipore Corporation: Billerica, MA, USA, 2013; p. 36. [Google Scholar]
- Mansfield, M.A. Nitrocellulose membranes for lateral flow immunoassays: A technical treatise. In Lateral Flow Immunoassay; Humana Press: Totowa, NJ, USA, 2009; pp. 1–19. [Google Scholar] [CrossRef]
- Joshi, M.; Kaur, S.; Shergill, B.; Sood, S.; Tulika, M. Isolation of multidrug-resistant bacteria from the hospital environment. Int. J. Res. Pharm. Sci. 2019, 10, 990–996. [Google Scholar] [CrossRef]
- Wang, M.; Cai, C.; Zhang, B.; Liu, H. Characterization and mechanism analysis of lincomycin biodegradation with Clostridium sp. strain LCM-B isolated from lincomycin mycelial residue (LMR). Chemosphere 2018, 193, 611–617. [Google Scholar] [CrossRef]
- European Union. Commission regulation (Ec) no 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants In foodstuffs. Off. J. Eur. Union 2006, 49, L364. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analyte | Ka, M−1 | Kd, M | ka, M−1·s−1 | kd, s−1 |
|---|---|---|---|---|
| OTA | 4.0 × 109 | 2.5 × 10−10 | 9.6 × 104 | 2.4 × 10−5 |
| T-2 | 1.7 × 1010 | 5.8 × 10−11 | 4.0 × 105 | 2.3 × 10−5 |
| AFB1 | 1.6 × 109 | 6.2 × 10−10 | 1.7 × 105 | 1.1 × 10−4 |
| DON | 2.1 × 109 | 4.8 × 10−10 | 3.3 × 105 | 1.6 × 10−4 |
| Analyte | Test Strip Design | Position | Instrumental LOD, ng/mL | IC50, ng/mL |
|---|---|---|---|---|
| AFB1 | Optimized | 1 | 2.9 ± 0.4 | 7.6 ± 0.8 |
| AFB1 | Non-optimized | 4 | 2.1 ± 0.6 | 4.3 ± 0.3 |
| T-2 | Non-optimized | 1 | 0.7 ± 0.3 | 1.4 ± 0.3 |
| T-2 | Optimized | 4 | 0.8 ± 0.1 | 1.5 ± 0.1 |
| DON | Optimized | 2 | 0.3 ± 0.1 | 2.9 ± 1.5 |
| DON | Non-optimized | 3 | 8.1 ± 2.5 | 19.9 ± 4.0 |
| OTA | Non-optimized | 2 | 2.5 ± 0.3 | 9.5 ± 1.4 |
| OTA | Optimized | 3 | 1.5 ± 0.6 | 4.7 ± 1.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bartosh, A.V.; Sotnikov, D.V.; Zherdev, A.V.; Dzantiev, B.B. Handling Detection Limits of Multiplex Lateral Flow Immunoassay by Choosing the Order of Binding Zones. Micromachines 2023, 14, 333. https://0-doi-org.brum.beds.ac.uk/10.3390/mi14020333
Bartosh AV, Sotnikov DV, Zherdev AV, Dzantiev BB. Handling Detection Limits of Multiplex Lateral Flow Immunoassay by Choosing the Order of Binding Zones. Micromachines. 2023; 14(2):333. https://0-doi-org.brum.beds.ac.uk/10.3390/mi14020333
Chicago/Turabian StyleBartosh, Anastasiya V., Dmitriy V. Sotnikov, Anatoly V. Zherdev, and Boris B. Dzantiev. 2023. "Handling Detection Limits of Multiplex Lateral Flow Immunoassay by Choosing the Order of Binding Zones" Micromachines 14, no. 2: 333. https://0-doi-org.brum.beds.ac.uk/10.3390/mi14020333