Targeting Programmed Fusobacterium nucleatum Fap2 for Colorectal Cancer Therapy



1
Food Science and Technology Program, Beijing Normal University-Hong Kong Baptist University United International College, Zhuhai 519087, China
2
Department of Microbial and Biochemical Pharmacy, School of Pharmaceutical Sciences, Sun Yat-Sen University, Guangzhou 510006, China
3
Laboratory for Translational Oncology and Personalized Medicine, Rashid Latif Medical College, Lahore 44000, Pakistan
*
Authors to whom correspondence should be addressed.
Cancers 2019, 11(10), 1592; https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11101592
Submission received: 28 September 2019
/
Revised: 13 October 2019
/
Accepted: 14 October 2019
/
Published: 18 October 2019
(This article belongs to the Special Issue Targeting Therapy for Colon Cancer)
Abstract
:Colorectal patients generally have the maximum counts of Fusobacterium nucleatum (F. nucleatum) in tumors and elevate colorectal adenomas and carcinomas, which show the lowest rate of human survival. Hence, F. nucleatum is a diagnostic marker of colorectal cancer (CRC). Studies demonstrated that targeting fusobacterial Fap2 or polysaccharide of the host epithelium may decrease fusobacteria count in the CRC. Attenuated F. nucleatum-Fap2 prevents transmembrane signals and inhibits tumorigenesis inducing mechanisms. Hence, in this review, we hypothesized that application of genetically programmed fusobacterium can be skillful and thus reduce fusobacterium in the CRC. Genetically programmed F. nucleatum is a promising antitumor strategy.
1. Introduction
Colorectal cancer (CRC) is the third-highest widespread malignant neoplasm and the fourth extremely common cause of malignancy death globally, and the five-year existence rate is less than 65 percent [1]. It is accountable for 694,000 demises yearly worldwide. It is a multifaceted disease, caused by genetic predisposition, lifestyle (sedentary, smoking), or diet (alcohol or red and processed meat consumption), and environmental exposure to various xenobiotics that could result in CRC development. Gut microbiota seems to be the mediator of this causal relationship, being disturbed by the exposure to such compounds, thus causing tumorigenic phenomena, a fact that constitutes a novel topic of research [2]. The mortality rate of CRC is based on the availability of medical resources [3]. In recent years, accumulating evidence greatly suggested a relationship between gut microbiota composition and CRC [4,5,6]. The human intestinal tract contains more than 100 trillion microorganisms that play a great significant function in human health. These organisms regulate gut homeostasis by maintaining several biological activities, including mucosal barrier, metabolic and immune functions [3,7,8]. Any disturbances occur in gut equilibrium, it may cause various intestinal illnesses, including Crohn’s disease, ulcerative colitis, and colorectal neoplasms [9,10]. There is great evidence that disturbance of gut microbiota can also lead to various systemic diseases such as diabetes, obesity, cancer, cardiovascular, and central nervous system disorders [11,12]. Increasing evidence confirmed that the gut microbiota is extremely connected with colorectal neoplasms [3,13,14,15].
Numerous investigations have confirmed that the levels of Bacteroides, Prevotella, Leptotrichia, Clostridium difficile, Streptococcus gallolyticus, Bacteroides fragilis, Enterococcus faecalis, Campylobacter spp., Escherichia coli, Fusobacterium nucleatum, and Streptococcus bovis are considerably higher in CRC compared to those in nearby normal tissue [16,17,18,19]. Studies in fecal samples containing Hungatella hathewayi, F. nucleatum, Clostridium asparagiforme, Klebsiella oxytoca, E. coli, Bilophila wadsworthia, and the genera Lactococcus, Porphyromonas, Odoribacter, Bilophila, and Pyramidobacter found to be enriched in patients with CRC [20,21]. F. nucleatum is a Gram-negative anaerobic bacterium found to be considerably higher and synergistically associated with other Gram-negative bacteria that promote the incidence and pathogenesis of CRC [22,23,24]. Gathering evidence suggested that the tumor tissues and fecal specimens of CRC patients have significantly increased counts of F. nucleatum [25,26,27]. This higher count of the organism may contribute to the development of CRC [28,29]. In a recent review regarding the role of oral bacteria and intestinal dysbiosis in CRC, it has been indicated that F. nucleatum merely resembles a passenger rather than a driver of intestinal dysbiosis in CRC according to the “driver-passenger model” about microbiota-driven CRC pathogenesis [30]. Nevertheless, the exact role of this specific bacterium in CRC progression is still an ongoing debate. It would be important at this point to include a brief summary of the “driver-passenger” model of microbiota dysbiosis, as introduced by Tjalsma et al. [31] since it is a crucial pathogenic concept regarding CRC. Hence, the F. nucleatum is a diagnostic marker of CRC. The positive detection rates of F. nucleatum in CRC patients testified by diverse study groups listed in Table 1. Further investigations have also confirmed that a higher count of F. nucleatum in CRC related to shorter survival rates [26,29].
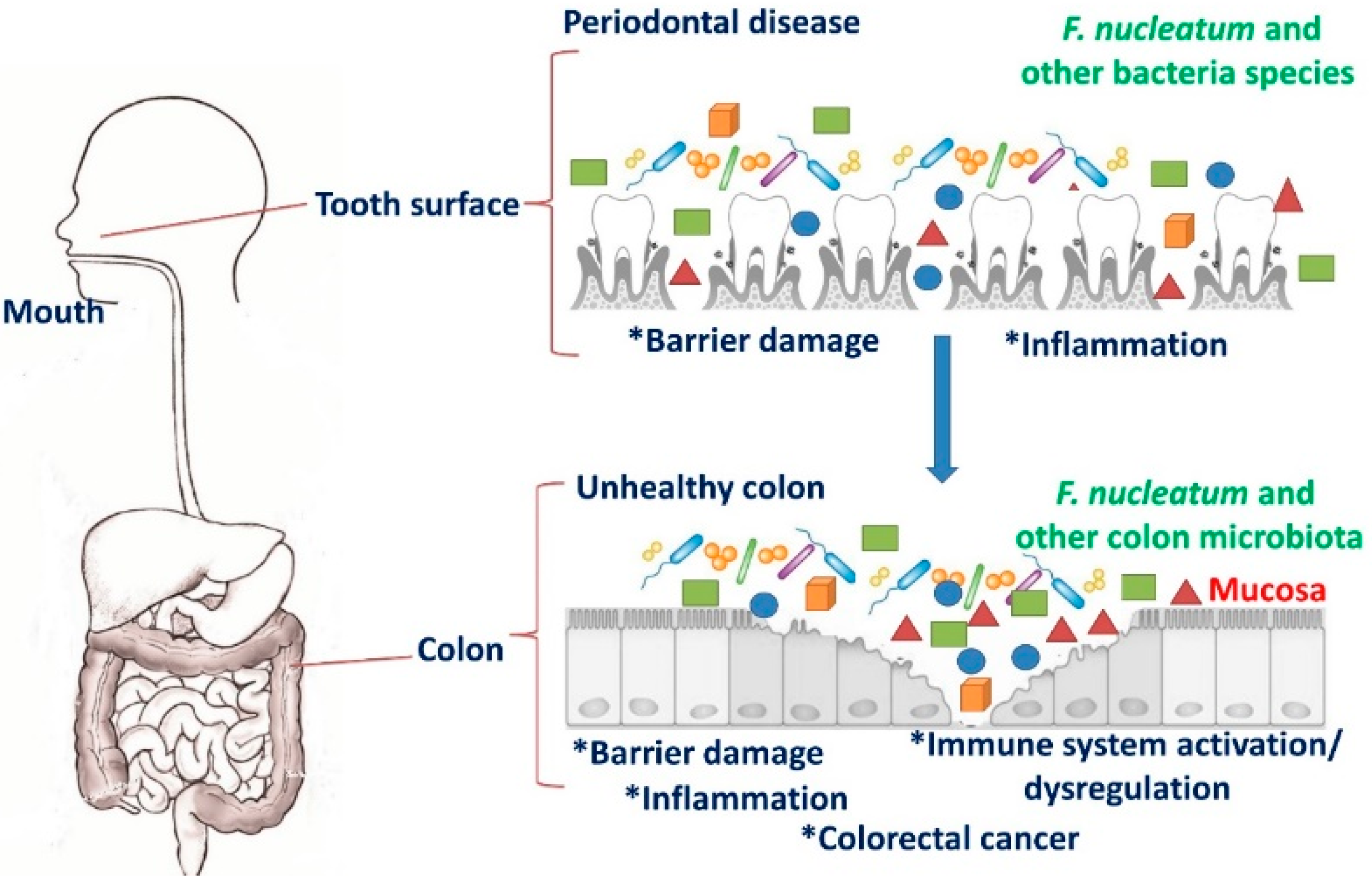
F. nucleatum normally lives in the oral cavity of human and is commonly associated with diseases such as gingivitis, periodontal plaque, tonsillitis, sinusitis, chronic periodontitis, liver abscess, and appendicitis [19,24,50]. In addition to oral infections, this organism has been implicated in respiratory tract infections, cardiovascular disease, arthritis, Alzheimer’s disease, adverse pregnancy outcomes, and the development of various cancers including esophageal, gastric, and colon [51]. It exhibits high levels of homology with other Fusobacterium species including, F. alocis, F. periodonticum, and F. simiae. All these Fusobacterium species reside in oral cavities [51]. It is usually absent or habitually not found elsewhere in the body under normal conditions [52]. Until recently, F. nucleatum was thought to primarily be a component of the human oral microbiota and only an occasional resident of the gut. However, this premise was built on a culture-based examination of stool, which usually does not contain high numbers of live, epithelium-associated bacteria. FISH was used to elegantly demonstrate an association between invasive Fusobacterium spp. (including F. nucleatum) cells with inflamed appendix tissues, suggesting that the gut could be a hitherto unrecognized niche for this pathogen [53]. The mechanism of migration of these bacteria from the oral cavity to intestines to promote CRC are illustrated in Figure 1.
CRC constitutes a huge global economic problem and vigorous action should be taken to reduce the financial cost of this disease [54]. CRC is one of the leading and genetically categorized malignancies, with definite somatic mutations, oncogenes, and tumor suppressor genes. These mutations and other cellular regulators are essential for the development of adenomatous lesions to hostile carcinoma [25,55]. Accumulating evidence showed that F. nucleatum is among the most abundant species of bacteria in CRC tissues [43,56]. The outer membrane of this organism contains variable thickness of lipopolysaccharide (LPS) that may present pathogenic properties [52], and are vital for the evasion of the immune system in the human [50].
In recent years, F. nucleatum has been recognized to be a possible causative agent of CRC, tumor development and promotes colorectal tumorigenesis in Apcmin/+ mice [29,57,58]. In situ hybridization studies have also confirmed that F. nucleatum is largely connected with malignancy cells in the metastatic lesions [58]. It stimulates tumor cell proliferation in CRC by activating β-catenin signaling and stimulating upstream regulation of oncogenic gene expression via the adhesive membrane virulence factor, includes protein adhesins, toxins, and enzymes [28]. Previously, Fecal F. nucleatum infection has been recognized as a significant diagnostic marker for CRC [47]. Our recent study also confirmed that among the important probiotics-Faecalibacterium and Bifidobacterium, F. nucleatum has identified as biomarkers for early CRC screening [59]. Taken together, these investigations demonstrated that F. nucleatum plays a significant role in the prime causes, diagnostic markers and progression of CRC and development.
2. F. nucleatum Mediate CRC and Inhibits Host Immune Response
F. nucleatum has been linked to immune suppression, through the promotion of lymphocytic apoptosis [60], and the abundance of F. nucleatum has been found to be inversely proportional to cluster of differentiation 3 (CD3+) T-cell density [61]. Recently, growing evidence demonstrated a high relationship between the infection of F. nucleatum and various cancers thus proposing innovative approaches in malignancy prevention by targeting F. nucleatum [29,56]. The earlier investigation has also established that F. nucleatum induces a noteworthy humoral immune response in chronic oral infection [62,63]. Recently, in our lab, we confirmed that F. nucleatum infection provoked high-level serum antibodies to F. nucleatum in CRC patients [64]. Using the sera of CRC patients to probe the bacteria protein extract, we found a robust reactive antigen, alkyl hydroperoxide reductase subunit C, activates the anti- F. nucleatum immune response [64].
F. nucleatum is a facultative intracellular anaerobic microorganism, possibly activating, proliferating, and migrating macrophages/monocytes that can provoke CRC development [34,65,66]. F. nucleatum can invade into endothelial and epithelial cells, induces the synthesis of pro-inflammatory cytokines, inflammatory lesions ultimately leading to CRC [29,67,68]. A contemporary study has also provided insights into the connection between the gut microbiota and the capacity of inflammatory cytokine production [69]. F. nucleatum promotes local inflammation and elevates the expression of inflammatory cytokines (Interleukins-IL-6, IL-8, tumor necrosis factor-alpha (TNF-α), and cyclooxygenase (COX-2), contributing to tumorigenic effects in CRC [28,29,69]. It can also induce chemokine C-C-motif ligand 20 (CCL20) expression in CRC while they are treated with F. nucleatum [66].
Nuclear factor kappa B (NF-κB) is a transcription factor that participates in regulating many gene expressions and promoting tumor development and progression [70]. It is well recognized as having crucial relationship with inflammation and cancer. The enrichment of the F. nucleatum triggers NF-κB activation that can be significantly involved in CRC development [71]. The expression of NF-κB is quite often triggering in F. nucleatum -enriched CRC [27,29]. Consistent with this study, Rubinstein et al. [28] have also confirmed that wild-type F. nucleatum continuously induces the expression of NF-κB in a human colon tumor (HCT116) xenograft model. Recently, two outer membrane proteins from F. nucleatum have been taken for attention, namely, fibroblast activation protein 2 (Fap2) and Fusobacterium adhesin A (FadA). Fap2 is a galactose-sensitive hemagglutinin and adhesive protein, which contributes to the invasive ability of F. nucleatum into the human cells that bind to D-galactose-β (1-3)-N-acetyl-D-galactosamine (Gal-GalNAc) [72,73,74,75]. FadA is another surface adhesive protein, exclusively present in F. nucleatum that plays a vital function in the mechanism of cell–cell attachment [75]. These two described proteins have greatly participated in the host cell attachment that modulates the expression of CRC signaling [28,76].
Yang et al. [27] found the massive quantities of F. nucleatum in the CRC tissues and the team further observed the increased invasion rate, proliferation, and xenograft tumors in humanized mice. F. nucleatum (specifically LPS) binds to TLR4, which trough myeloid differentiation primary response gene 88 (MYD88) signaling activates the NF-κΒ pathway, which enhances the gene expression of miR-21 [30]. Toll like receptor 4 (TLR4) is a chief receptor for bacterial LPS that overexpresses in CRC and plays a vital function in tumor development [77]. miR-21 may serve as a key promotor of colitis-associated colon cancer [78,79]. Earlier studies revealed that patients with a higher count of F. nucleatum and miR-21 exhibited the lowest survival rate [27]. Antibiotic treatment of CRC in mice bearing xenograft models reduced the count of Fusobacterium and prevent the proliferation of all tumor growth. However, these findings debate with the clinical investigation of antimicrobial interventions as effective management with F. nucleatum -associated CRC [80]. Moreover, our study also identified that F. nucleatum is sensitive to the tryptophan-depleted microenvironments and kynurenine could inhibit the growth of F. nucleatum, suggesting that tryptophan metabolism plays a role during the infection [65]. In addition, our study further confirmed that the subunit vaccine for F. nucleatum, such as alkyl hydroperoxide reductase subunit C can reduce F. nucleatum load in the intestinal tract [59,81].
Gut microbiota of humans comprises a diverse range of microbial strains in which certain strains are well recognized as carcinogenic agents [76]. In 2012, two different studies confirmed that Fusobacterium species or F. nucleatum in specific is excessively habituated in CRC tissues when compared to the normal mucosa of the gut [25,58]. Fusobacterium species are normal inhabitants of the oral cavity and poor invader to the healthy intestine and however, during an unhealthy tumor environment, these organisms can be reached to the gut [75]. These outcomes were effectively confirmed by McCoy et al. [57] and these authors have demonstrated an excess of Fusobacterium species, acknowledged as CRC precursors, which are highly populated in the colorectal adenomas (CRAs) when compared to the normal gut mucosa. Subsequently, numerous investigations have also proved the connection between Fusobacterium and CRA [82]. To recognize the molecular pathway associated with F. nucleatum, inflammation, and CRC, Kostic et al. [29] have demonstrated using animals (ApcMin/+) and found the genetic vulnerability of emerging colonic cancers. These animals were administered with invasive organisms of F. nucleatum initially obtained from the patient’s intestine of inflammatory bowel disease. The outcome showed elevated tumorigenesis, caused due to elevating infiltrating cells of the myeloid lineage (dendritic cells, macrophages, and granulocytes) [29]. Eventually, F. nucleatum induced inflammation as well as tumor proliferation through regulating tumor-immune microenvironment [29]. These animal trials confirmed that human colonic strains strongly correlated with the richness of F. nucleatum and pro-inflammatory markers expression [29,57]. F. nucleatum has been known to suppress anti-tumor immunity by preventing tumor-killing cells (natural killer cells and tumor-infiltrating lymphocytes) [74]. Based on these experimental outcomes, F. nucleatum is not only inhabited and developed in CRAs and adenocarcinomas but also augments tumor progression and existence through the tumor-immune microenvironment.
3. Fap2 in F. nucleatum Mediates CRC through Host Gal-GalNAc
Fibroblast activation protein 2 (Fap2) plays a critical function in mediating CRC development through binding with acetylgalactosamine (Gal-GalNAc), which is overexpressed in human metastases and colorectal adenocarcinoma [83]. Fap2 is a galactose-sensitive hemagglutinin and adhesin that possibly plays a function in the virulence of Fusobacterium. It is a 390-KDa protein encoded by the Fap2 gene of F. nucleatum [73]. The outer membrane composed of 3692 amino acid, which is recognized to stimulate apoptosis in human lymphocytes [48]. Earlier study demonstrated that Fap2 has participated in the binding of F. nucleatum to malignant cells and interacts with the immunoglobulin and immunoreceptor tyrosine based inhibitory motif (ITIM) domain receptor mainly expressed on natural killer cells (NK), regulatory T cells (Treg), cluster of differentiation-CD8+, and CD4+ T cells [75]. The binding of Fap2 to T cell immuneoreceptor with Immunoglobulin G (TIGIT) was found to inhibit the activity of NK cells against the tumor cells, thus causing the development of CRC [74].
Abed et al. [84] have confirmed the quantities of fusobacterial lectin and host polysaccharide that explicates fusobacterial abundance in the CRC. This study further explains that host epithelial Gal-GalNAc over-expressed in the CRC, which is identified by fusobacterial Fap2, providing its role as a Gal-GalNAc lectin [84]. High Gal-GalNAc levels are also found in CRC and are connected with fusobacterial genomic DNA occurrence in these metastases, representing the ability of fusobacteria to colonize CRC [85]. The team has also further confirmed in orthotopic rectal colon tumor (CT26) adenocarcinoma model that intravascular injection of oral F. nucleatum strain favors inhabiting in CRC tissue. This transmission route is mediated through the binding of Fap2 to Gal-GalNAc. Hence, all these supporting ideas have provided oral fusobacteria that may colonize CRC via a hematogenous route [84].
Gal-GalNAc lectin is universally familiar by the immune sera of patients with amoebic liver abscess and various other diseases [86]. It plays a crucial role in cytolysis and phagocytosis of human and rat colonic mucin glycoproteins. Earlier study related to the uptake of L. pneumophila by H. vermiformis was specifically inhibited by Gal-GalNAc against the lectin of E. histolytica. Remarkably, the inhibition of invasion by Gal-GalNAc was connected with inhibition of bacterial-induced tyrosine dephosphorylation of H. vermiformis proteins [87]. Normally, the functions of the lectin comprise the host cell binding, cytotoxicity, complement resistance, induction of encystation, and generation of the cyst wall. In addition, the functions of the lectin in both differentiation and virulence suggest that it may be a pivotal molecule that determines the severity of the infection from a commensal state resulting from increased encystation to an invasive state [88]. Earlier studies, high levels of the tumor expressed Gal-GalNAc moieties have shown in adenocarcinomas of various organs such as stomach [89], prostate [90], ovary [91], colon [92], uterus [93], pancreas [94], breast [89], lung [95], and esophagus [96].
Clinical fusobacteria strains that present lacking Fap2 or inactivated Fap2 mutants demonstrate reduced binding to Gal-GalNAc-expressing CRC cells [84]. Providing the tumorigenic role of fusobacteria and its immune evasion potential, the removal of fusobacteria might be a promising and improve treatment outcome of the CRC [85]. Moreover, fusobacteria seems to explicitly bind to Gal-GalNAc-displaying tumors, it must be programmed as a stage for the treating CRC. As immunosuppression is undesired in the future fusobacterial-based tumor therapy, the interesting novelty of detecting and inactivation of the Fap2 TIGIT receptor-activating domain highly required.
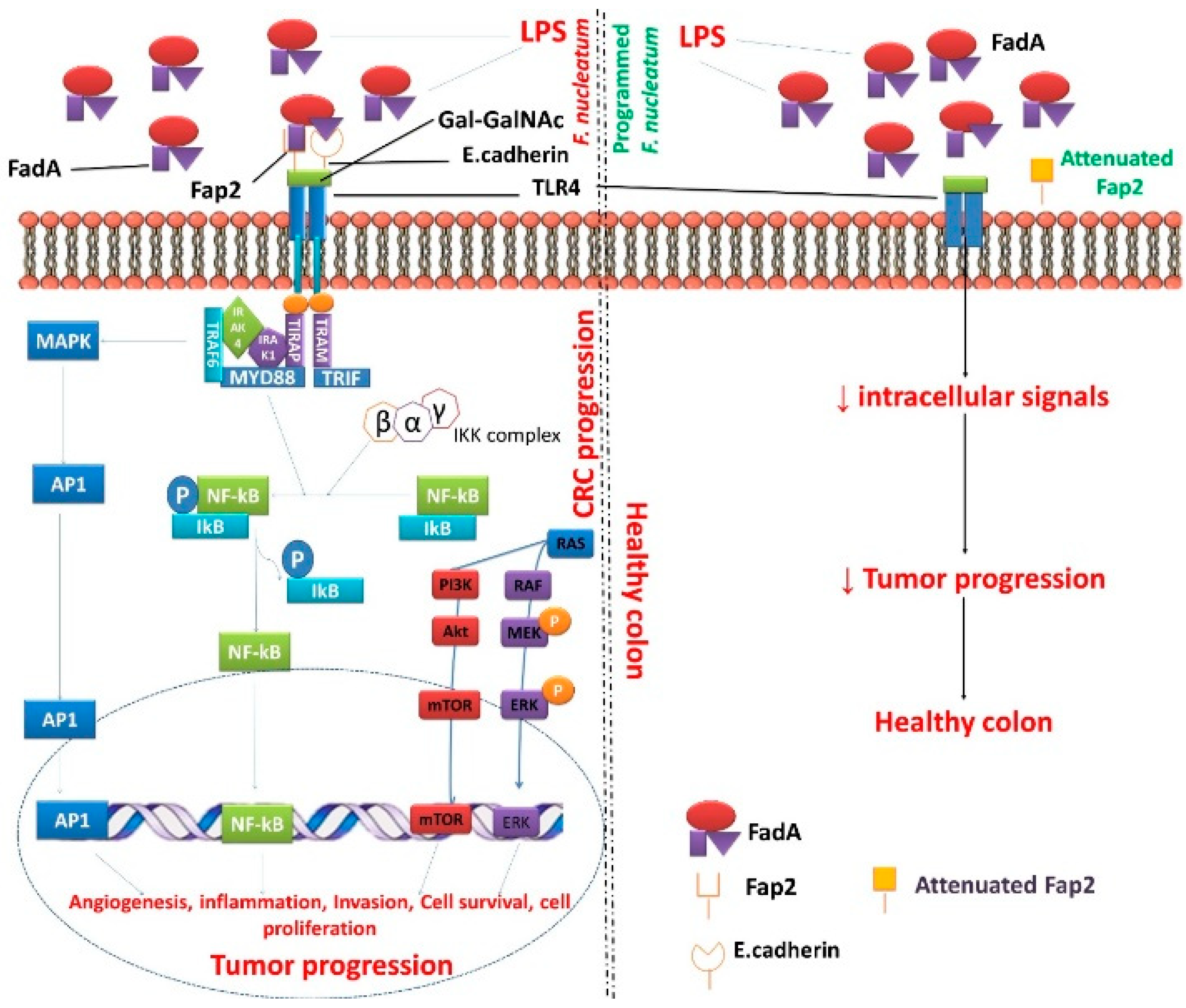
F. nucleatum infects the oral cavity and reaches the colon, whereby it causes tumor progression. However, the proposal of the programmed F. nucleatum promotes a healthy colon and prevents tumor prevention in the host. Outer membrane protein of F. nucleatum- Fap2 contributes to the invasive ability of F. nucleatum into the human that binds to E-cadherin and Gal-GalNAc. Fap2 binds only to Gal-GalNAc. FadA binds to E-cadherin [97]. Through Toll-like receptor 4, the F. nucleatum actively triggers the signals to MYD88 that stimulate activating protein-1 (AP1), NF-κB, mammalian target of rapamycin (mTOR), and extracellular signal-regulated kinase (ERK) pathways and eventually cause tumorigenesis. Although these facts are generally true, the activation of TLR4 is due to binding with LPS, not Fap2 [77]. The attenuated F. nucleatum-Fap2 prevents transmembrane signals and inhibit tumorigenesis inducing mechanisms. The genetically programmed mechanism can be achieved in fusobacterial Fap2 by the following steps: mutagenesis, rDNA techniques, attenuation of oncogenic materials performed by using vector encoded siRNA and shRNAs; induction of tumor suppressor genes, and immunogenic peptides. Targeting programmed fusobacterial Fap2 may reduce fusobacterium count in the CRC and promotes a healthy colon. Thus, the programmed F. nucleatum Fap2 is a promising anti-tumor activity against CRC.
Adhering to the gut epithelium by the various cell surface proteins, FadA, Fap2, and role of radiation genes (RadD) expressed by F. nucleatum can cause CRC in humans and produce inflammatory factors in the tumor microenvironment [98]. Rubinstein et al. [28] demonstrated that F. nucleatum enters into the host and persuades inflammation and oncogenic responses that proliferate CRC cells via its FadA attachment. FadA is a surface adhesive protein localized in F. nucleatum that functions as cell binding [75]. These FadA proteins fix with host E-cadherin (cellular adhesion) to trigger Wnt/β-catenin signaling pathways and regulate the inflammation as well as oncogenic responses (Figure 2). FadA interacts with E-cadherin at a locality of the 11-amino-acid region [28]. However, this amino acid proximately synthesizes a new peptide in the human, which terminates FadA induced cell proliferation, inflammation, and oncogenic responses in the colon [28]. In patients with adenomas and adenocarcinomas, FadA levels are 10–100 times higher than normal subjects [57]. These elevated FadA levels in CRC mostly associate with augmented inflammatory signaling and tumorigenic responses [28]. This investigation further reveals a key mechanism of F. nucleatum that can potentially regulate CRC and recognizes FadA as a possible diagnosis and therapeutic marker for CRC [76]. F. nucleatum binds to the patient’s adenocarcinomas associated with Gal-GalNAc expression that has been diminished upon O-glycanase treatment [84]. Therefore, targeting Fap2 in F. nucleatum and or host epithelial Gal-GalNAc could shorten fusobacteria potentiation in the CRC.
Researchers have strongly believed that F. nucleatum is prevalent and resident in high numbers during tooth brushing as well as a periodontal disease [99]. There are high chances that this transient oral fusobacteria may transmit through the circulatory system to spread CRA and CRC spots. Since F. nucleatum transition to tumor spots is a detrimental effect to the host [74], new avenues to block the enrichment of F. nucleatum in tumor sites or CRC would be therapeutically beneficial.
4. Bacteria-Mediated Cancer Treatment: Alternative to Surgery
Surgical treatment is the utmost general cancer therapy, and it has been practiced for several centuries [100]. Nevertheless, surgical procedure is not an actual treatment for metastatic conditions since they need radiation and chemotherapy [101]. The surgical procedure has lots of hitches that might offer incomplete elimination of tumor growth and possible reappearance [102]. The potential treatment of radiotherapy mostly affects tissue oxygen levels, however hypoxic environments occur in cancerous spots that result in failure of the treatment [101]. Moreover, the drug transition is therapeutically effective for chemotherapy; and deprived vasculature in cancer spots weakens drug delivery, which reduces the efficacy of the drugs, particularly in hypoxic and necrotic environments [102]. The usage of microbes in malignancy treatment has been practiced several times [102], and it is not well documented as an effective therapeutic tactic. Efficient knowledge and scientific progress have permitted generation of genetically programmed bacteria that drive harmless and effective application in cancer therapy. Bacteria-mediated cancer treatment facilitates facultative anaerobes that can able to live even in hypoxic and necrotic environments. It aids drug transition all over the tumor sites [103]. Nowadays, the sum of available bacterial therapy articles has been swiftly improved, in which the use of bacterial therapy of Salmonella has increased significantly [104].
5. Programmed Bacteria
Synthetic biology is dynamic to a new era of medicine through the genetic programming of living cells [105]. This transformative method allows for the creation of engineered systems and adding specificity and effectiveness that encompasses beyond the competences of molecular-based therapeutics [106,107]. One specific area of attention has been the programming of bacteria as therapeutic delivery systems to selectively discharge therapeutic cargos in vivo [105,106,108,109]. Chowdhury et al. [105] engineered a non-pathogenic Escherichia coli strain encoded with CD47nb, which elevates activation of tumor-infiltrating T cells, induces quick tumor regression, averts metastasis, and provides long-term survival in a syngeneic tumor mouse model. Harimoto et al. [110] screened Salmonella typhimurium strains expressing and carrying antitumor therapeutic molecules through various programmed gene circuits. In addition, these research groups have identified the candidates exhibiting noteworthy tumor reduction in a syngeneic mouse model [110]. This platform can be aided to identify the programmed diverse microbial species such as Listeria monocytogenes, Proteus mirabilis, and Escherichia coli in several host cell types [110].
Yoon et al. [111] also indicated that inherently adapted attenuated Salmonella typhimurium, that produces interferon-gamma (IFN-γ) as a tumoricidal agent, could elevate the therapeutic efficiency. In 1891, one of the greatest bone sarcoma surgeon named William B. Coley inoculated streptococcal organisms into a cancer patient, and those organisms successfully cured the patient of malignancy [104]. The anti-tumor activity of attenuated bacteria was achieved in various animal models. It has been achieved by antigen-specific tumor inhibition [112], reduced tumor mass [113,114], reduced proinflammatory stimulation [115,116], suppression of angiogenesis and pulmonary metastasized tumors [117], and eventually increases the survival time [118]. The list of programmed bacteria expressed as anti-tumor agents in various animal models are given in Table 2.
6. Mechanism of Tumoricidal Properties of Programmed Bacteria
Indeed, bacteria synthesize exotoxins through their type 1 secretion systems (T1SS) for their survival in the rigid environment [138]. T1SS is generally chaperone-dependent machinery employing proteins expressed by hly and tol genes [113]. Clinically significant T1SS cargo comprises of proteins, polysaccharides, ions, and small molecules, which are termed as exotoxin. For instance, uropathogenic E. coli contains a virulence factor of exotoxin: a-hemolysin (HlyA). This set of exotoxin normally aids pores on the host cells and consequently has the capacity to break down blood cells as well as cancer cells [139]. The studies showed that T1SS machinery activates chimeric human prostate-specific antigen (PSA), which elevates CD8+ cell-mediated reactions against a mouse mastocytoma [135]. Employing T1SS machinery in recent investigations showed a reduction of cancerous proliferation when hlyE was secreted by recombinant S. typhimurium using arabinose-inducible [113] and hypoxia-inducible bacterial promoter [114] in syngeneic hosts, even though the nature of anti-tumor reaction performed mainly not reliant on CD8+ cells.
The promoter of recombinant S. typhimurium [131] established to synthesize Shiga toxin in the tumor microenvironment, which causes tumor necrosis. When E. coli expresses a chimeric heterologous bacterial toxin, Listeriolysin-O (LLO), it can be elevated CD8+ cell-mediated antitumor response [125]. LLO is an exotoxin secreted by the bacterium Listeria monocytogenes, similar to hemolysin of E. coli. Likewise, when S. typhimurium synthesized a chimeric heterologous antigen, CD8+ cell-mediated anti-tumor reactions can be aroused [112]. Based on this experimental methodology, anti-tumor responses were initiated and experiments were performed using nude mice to control experimental tumors [140,141]. This high-throughput framework may serve to accelerate synthetic biology for translational medicine and for understanding the host-microbe interactions in disease sites.
7. Targeting Programmed F. nucleatum Fap2 for Colorectal Cancer
According to current studies, the tumor microenvironment and feces samples of patients with CRC are enriched by F. nucleatum [98]. Therefore, F. nucleatum is projected as one of the risk factors in the commencement and progression of CRC. The most significant mechanisms of F. nucleatum participated in CRC carcinogenesis are virulence factors (FadA, Fap2, RadD), miR-21, immunomodulation (inhibitory receptors of NK cells and elevating myeloid-derived suppressor cells), and metabolism of bacteria [98,142]. Studies showed that a host polysaccharide and lectin of fusobacterium that explains CRC comprises the richness of fusobacterium [28]. The host epithelium sugars of Gal-GalNAc are generally expressed higher in CRC, which is recognized by binding with fusobacterial Fap2. Thus, targeting fusobacterial Fap2 or host epithelium polysaccharide of Gal-GalNAc may decrease fusobacteria count in the CRC. Enormous studies previously described that bacteria act as anti-tumorigenic or anti-cancer agents [27,107,142]. The entire bacterium or the immunogenic properties of peptides carried by the bacteria show an effective anti-tumorigenic potent in the animal models of various cancers [106,107]. In this context, the application of attenuated host strains resulting from mutagenesis, rDNA techniques could also be practiced in bacteria that aid to eliminate tumor colonization. Vector encoded siRNA and shRNAs that target oncogenic materials, induction of tumor suppressor genes, and immunogenic peptides could also be developed. These methodologies show that the genetically programmed F. nucleatum has promising anti-tumor activity against CRC.
8. Conclusions
F. nucleatum infects the oral cavity to the colon that interrelates with the host immune system, ultimately cause inflammation and tumor progression. F. nucleatum exhibited the ability to cause CRC by Fap2-Gal-GalNAc complex specifically. However, attenuated F. nucleatum-Fap2 prevent transmembrane cellular signals and avert tumorigenesis persuading mechanisms. The genetically programmed practice can be performed in fusobacterial Fap2 by mutagenesis, rDNA techniques, attenuation of oncogenic materials, stimulation of immunogenic peptides and tumor suppressor genes. Targeting programmed fusobacterial Fap2 may reduce fusobacterium count in the CRC and promotes a healthy colon. Thus, the development of genetically programmed F. nucleatum Fap2 is a promising anti-tumor activity that will provide more methods for bacteria-based cancer treatment.
Author Contributions
K.G. and B.X. conceived and designed the review; K.G., S.G., S.F. and G.Z. wrote the paper; and S.G., S.F. and B.X. critically revised and improved the manuscript.
Funding
This research was funded by National Natural Science Foundation of China (No. 81673005 and National Natural Science Foundation of Guangdong Province (No.2017A030313746).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Shang, F.-M.; Liu, H.-L. Fusobacterium nucleatum and colorectal cancer: A review. World J. Gastrointest. Oncol. 2018, 10, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wang, P.; Yu, Y.; Huang, B.; Zhang, X.; Xu, C.; Zhao, X.; Yin, Z.; He, Z.; Jin, M.; et al. Tumor-preventing activity of aspirin in multiple cancers based on bioinformatic analyses. PeerJ 2018, 6, e5667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, F.; Cai, Y. Study insights into gastrointestinal cancer through the gut microbiota. BioMed Res. Int. 2019, 2019, 8721503. [Google Scholar] [CrossRef] [PubMed]
- Jobin, C. Colorectal cancer: Looking for answers in the microbiota. Cancer Discov. 2013, 3, 384–387. [Google Scholar] [CrossRef] [PubMed]
- Mira-Pascual, L.; Cabrera-Rubio, R.; Ocon, S.; Costales, P.; Parra, A.; Suarez, A.; Moris, F.; Rodrigo, L.; Mira, A.; Collado, M.C. Microbial mucosal colonic shifts associated with the development of colorectal cancer reveal the presence of different bacterial and archaeal biomarkers. J. Gastroenterol. 2014, 50, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, K.; Xu, B. Molecular targets of vitexin and isovitexin in cancer therapy: A critical review. Ann. N. Y. Acad. Sci. 2017, 1401, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Fu, C.; Zhao, Y.; He, J. Intervention with α-ketoglutarate ameliorates colitis-related colorectal carcinoma via modulation of the gut microbiome. BioMed Res. Int. 2019, 2019, 8020785. [Google Scholar] [CrossRef]
- Ganesan, K.; Chung, S.K.; Vanamala, J.; Xu, B. Causal relationship between diet-induced gut microbiota changes and diabetes: A novel strategy to transplant Faecalibacterium prausnitzii in preventing diabetes. Int. J. Mol. Sci. 2018, 19, 3720. [Google Scholar] [CrossRef]
- Arthur, J.C.; Jobin, C. The complex interplay between inflammation, the microbiota and colorectal cancer. Gut Microbes 2013, 4, 253–258. [Google Scholar] [CrossRef] [Green Version]
- Candela, M. Inflammation and colorectal cancer, when microbiota-host mutualism breaks. World J. Gastroenterol. 2014, 20, 908. [Google Scholar] [CrossRef]
- Sender, R.; Fuchs, S.; Milo, R. Revised estimates for the number of human and bacteria cells in the body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [PubMed]
- Belizario, J.E.; Faintuch, J. Microbiome and gut dysbiosis. Exp. Suppl. (2012) 2018, 109, 459–476. [Google Scholar] [CrossRef]
- Viljoen, K.S.; Dakshinamurthy, A.; Goldberg, P.; Blackburn, J.M. Quantitative profiling of colorectal cancer-associated bacteria reveals associations between Fusobacterium spp., enterotoxigenic Bacteroides fragilis (ETBF) and clinicopathological features of colorectal cancer. PLoS ONE 2015, 10, e0119462. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; He, H.; Xu, H.; Li, Y.; Li, Z.; Du, Y.; He, J.; Zhou, Y.; Wang, H.; Nie, Y. Association of oncogenic bacteria with colorectal cancer in South China. Oncotarget 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.-E.; Chiu, C.-T.; Rayner, C.K.; Wu, K.-L.; Chiu, Y.-C.; Hu, M.-L.; Chuah, S.-K.; Tai, W.-C.; Liang, C.-M.; Wang, H.-M. Associated factors in Streptococcus bovis bacteremia and colorectal cancer. Kaohsiung J. Med. Sci. 2016, 32, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Sobhani, I.; Tap, J.; Roudot-Thoraval, F.; Roperch, J.P.; Letulle, S.; Langella, P.; Corthier, G.; Van Nhieu, J.T.; Furet, J.P. Microbial dysbiosis in colorectal cancer (CRC) patients. PLoS ONE 2011, 6, e16393. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, S.; Eslick, G.D. Streptococcus bovisinfection and colorectal neoplasia: A meta-analysis. Colorectal Dis. 2014, 16, 672–680. [Google Scholar] [CrossRef]
- Gagnière, J. Gut microbiota imbalance and colorectal cancer. World J. Gastroenterol. 2016, 22, 501. [Google Scholar] [CrossRef]
- Yang, Y.; Misra, B.B.; Liang, L.; Bi, D.; Weng, W.; Wu, W.; Cai, S.; Qin, H.; Goel, A.; Li, X.; et al. Integrated microbiome and metabolome analysis reveals a novel interplay between commensal bacteria and metabolites in colorectal cancer. Theranostics 2019, 9, 4101–4114. [Google Scholar] [CrossRef]
- Song, M.; Chan, A.T.; Sun, J. Influence of the gut microbiome, diet, and environment on risk of colorectal cancer. Gastroenterology 2019, in press. [Google Scholar] [CrossRef]
- Yazici, C.; Wolf, P.G.; Kim, H.; Cross, T.L.; Vermillion, K.; Carroll, T.; Augustus, G.J.; Mutlu, E.; Tussing-Humphreys, L.; Braunschweig, C.; et al. Race-dependent association of sulfidogenic bacteria with colorectal cancer. Gut 2017, 66, 1983–1994. [Google Scholar] [CrossRef]
- Kharrat, N.; Assidi, M.; Abu-Elmagd, M.; Pushparaj, P.N.; Alkhaldy, A.; Arfaoui, L.; Naseer, M.I.; Omri, A.E.; Messaoudi, S.; Buhmeida, A.; et al. Data mining analysis of human gut microbiota links Fusobacterium spp. with colorectal cancer onset. Bioinformation 2019, 15, 372–379. [Google Scholar] [CrossRef]
- Warren, R.L.; Freeman, D.J.; Pleasance, S.; Watson, P.; Moore, R.A.; Cochrane, K.; Allen-Vercoe, E.; Holt, R.A. Co-occurrence of anaerobic bacteria in colorectal carcinomas. Microbiome 2013, 1, 16. [Google Scholar] [CrossRef]
- Yu, J.; Feng, Q.; Wong, S.H.; Zhang, D.; Liang, Q.y.; Qin, Y.; Tang, L.; Zhao, H.; Stenvang, J.; Li, Y.; et al. Metagenomic analysis of faecal microbiome as a tool towards targeted non-invasive biomarkers for colorectal cancer. Gut 2015, 66, 70–78. [Google Scholar] [CrossRef]
- Castellarin, M.; Warren, R.L.; Freeman, J.D.; Dreolini, L.; Krzywinski, M.; Strauss, J.; Barnes, R.; Watson, P.; Allen-Vercoe, E.; Moore, R.A.; et al. Fusobacterium nucleatum infection is prevalent in human colorectal carcinoma. Genome Res. 2011, 22, 299–306. [Google Scholar] [CrossRef]
- Mima, K.; Nishihara, R.; Qian, Z.R.; Cao, Y.; Sukawa, Y.; Nowak, J.A.; Yang, J.; Dou, R.; Masugi, Y.; Song, M.; et al. Fusobacterium nucleatumin colorectal carcinoma tissue and patient prognosis. Gut 2015, 65, 1973–1980. [Google Scholar] [CrossRef]
- Yang, Y.; Weng, W.; Peng, J.; Hong, L.; Yang, L.; Toiyama, Y.; Gao, R.; Liu, M.; Yin, M.; Pan, C.; et al. Fusobacterium nucleatum increases proliferation of colorectal cancer cells and tumor development in mice by activating toll-like receptor 4 signaling to nuclear factor-κB, and up-regulating expression of microRNA-21. Gastroenterology 2017, 152, 851–866.e824. [Google Scholar] [CrossRef]
- Rubinstein, M.R.; Wang, X.; Liu, W.; Hao, Y.; Cai, G.; Han, Y.W. Fusobacterium nucleatum promotes colorectal carcinogenesis by modulating E-cadherin/β-catenin signaling via its FadA adhesin. Cell Host Microbe 2013, 14, 195–206. [Google Scholar] [CrossRef]
- Kostic, A.D.; Chun, E.; Robertson, L.; Glickman, J.N.; Gallini, C.A.; Michaud, M.; Clancy, T.E.; Chung, D.C.; Lochhead, P.; Hold, G.L.; et al. Fusobacterium nucleatum potentiates intestinal tumorigenesis and modulates the tumor-immune microenvironment. Cell Host Microbe 2013, 14, 207–215. [Google Scholar] [CrossRef]
- Koliarakis, I.; Messaritakis, I.; Nikolouzakis, T.K.; Hamilos, G.; Souglakos, J.; Tsiaoussis, J. Oral bacteria and intestinal dysbiosis in colorectal cancer. Int. J. Mol. Sci. 2019, 20, 4146. [Google Scholar] [CrossRef]
- Tjalsma, H.; Boleij, A.; Marchesi, J.R.; Dutilh, B.E. A bacterial driver–passenger model for colorectal cancer: Beyond the usual suspects. Nat. Rev. Microbiol. 2012, 10, 575–582. [Google Scholar] [CrossRef]
- Dejea, C.M.; Wick, E.C.; Hechenbleikner, E.M.; White, J.R.; Mark Welch, J.L.; Rossetti, B.J.; Peterson, S.N.; Snesrud, E.C.; Borisy, G.G.; Lazarev, M.; et al. Microbiota organization is a distinct feature of proximal colorectal cancers. Proc. Natl. Acad. Sci. USA 2014, 111, 18321–18326. [Google Scholar] [CrossRef] [Green Version]
- Marchesi, J.R.; Dutilh, B.E.; Hall, N.; Peters, W.H.M.; Roelofs, R.; Boleij, A.; Tjalsma, H. Towards the human colorectal cancer microbiome. PLoS ONE 2011, 6, e20447. [Google Scholar] [CrossRef]
- Park, C.H.; Han, D.S.; Oh, Y.-H.; Lee, A.R.; Lee, Y.-R.; Eun, C.S. Role of Fusobacteria in the serrated pathway of colorectal carcinogenesis. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Komiya, Y.; Shimomura, Y.; Higurashi, T.; Sugi, Y.; Arimoto, J.; Umezawa, S.; Uchiyama, S.; Matsumoto, M.; Nakajima, A. Patients with colorectal cancer have identical strains of Fusobacterium nucleatum in their colorectal cancer and oral cavity. Gut 2019, 68, 1335–1337. [Google Scholar] [CrossRef]
- Gao, Z.; Guo, B.; Gao, R.; Zhu, Q.; Qin, H. Microbiota disbiosis is associated with colorectal cancer. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef]
- Gao, R.; Kong, C.; Huang, L.; Li, H.; Qu, X.; Liu, Z.; Lan, P.; Wang, J.; Qin, H. Mucosa-associated microbiota signature in colorectal cancer. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 2073–2083. [Google Scholar] [CrossRef]
- Burns, M.B.; Lynch, J.; Starr, T.K.; Knights, D.; Blekhman, R. Virulence genes are a signature of the microbiome in the colorectal tumor microenvironment. Genome Med. 2015, 7. [Google Scholar] [CrossRef]
- Feng, Q.; Liang, S.; Jia, H.; Stadlmayr, A.; Tang, L.; Lan, Z.; Zhang, D.; Xia, H.; Xu, X.; Jie, Z.; et al. Gut microbiome development along the colorectal adenoma–carcinoma sequence. Nat. Commun. 2015, 6. [Google Scholar] [CrossRef]
- Amitay, E.L.; Werner, S.; Vital, M.; Pieper, D.H.; Höfler, D.; Gierse, I.-J.; Butt, J.; Balavarca, Y.; Cuk, K.; Brenner, H. Fusobacterium and colorectal cancer: Causal factor or passenger? Results from a large colorectal cancer screening study. Carcinogenesis 2017, 38, 781–788. [Google Scholar] [CrossRef]
- Ahn, J.; Sinha, R.; Pei, Z.; Dominianni, C.; Wu, J.; Shi, J.; Goedert, J.J.; Hayes, R.B.; Yang, L. Human gut microbiome and risk for colorectal cancer. J. Natl. Cancer Inst. 2013, 105, 1907–1911. [Google Scholar] [CrossRef] [PubMed]
- Vogtmann, E.; Hua, X.; Zeller, G.; Sunagawa, S.; Voigt, A.Y.; Hercog, R.; Goedert, J.J.; Shi, J.; Bork, P.; Sinha, R. Colorectal cancer and the human gut microbiome: Reproducibility with whole-genome shotgun sequencing. PLoS ONE 2016, 11, e0155362. [Google Scholar] [CrossRef]
- Yamaoka, Y.; Suehiro, Y.; Hashimoto, S.; Hoshida, T.; Fujimoto, M.; Watanabe, M.; Imanaga, D.; Sakai, K.; Matsumoto, T.; Nishioka, M.; et al. Fusobacterium nucleatum as a prognostic marker of colorectal cancer in a Japanese population. J. Gastroenterol. 2018, 53, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y.; Ge, Q.X.; Cao, J.; Zhou, Y.J.; Du, Y.L.; Shen, B.; Wan, Y.J.; Nie, Y.Q. Association of Fusobacterium nucleatum infection with colorectal cancer in Chinese patients. World J. Gastroenterol. 2016, 22, 3227–3233. [Google Scholar] [CrossRef] [PubMed]
- Tahara, T.; Yamamoto, E.; Suzuki, H.; Maruyama, R.; Chung, W.; Garriga, J.; Jelinek, J.; Yamano, H.O.; Sugai, T.; An, B.; et al. Fusobacterium in colonic flora and molecular features of colorectal carcinoma. Cancer Res. 2014, 74, 1311–1318. [Google Scholar] [CrossRef] [PubMed]
- Suehiro, Y.; Sakai, K.; Nishioka, M.; Hashimoto, S.; Takami, T.; Higaki, S.; Shindo, Y.; Hazama, S.; Oka, M.; Nagano, H.; et al. Highly sensitive stool DNA testing of Fusobacterium nucleatum as a marker for detection of colorectal tumours in a Japanese population. Ann. Clin. Biochem. 2017, 54, 86–91. [Google Scholar] [CrossRef]
- Wong, S.H.; Kwong, T.N.Y.; Chow, T.C.; Luk, A.K.C.; Dai, R.Z.W.; Nakatsu, G.; Lam, T.Y.T.; Zhang, L.; Wu, J.C.Y.; Chan, F.K.L.; et al. Quantitation of faecal Fusobacterium improves faecal immunochemical test in detecting advanced colorectal neoplasia. Gut 2017, 66, 1441–1448. [Google Scholar] [CrossRef]
- Nosho, K.; Sukawa, Y.; Adachi, Y.; Ito, M.; Mitsuhashi, K.; Kurihara, H.; Kanno, S.; Yamamoto, I.; Ishigami, K.; Igarashi, H.; et al. Association of Fusobacterium nucleatum with immunity and molecular alterations in colorectal cancer. World J. Gastroenterol. 2016, 22, 557–566. [Google Scholar] [CrossRef]
- Ito, M.; Kanno, S.; Nosho, K.; Sukawa, Y.; Mitsuhashi, K.; Kurihara, H.; Igarashi, H.; Takahashi, T.; Tachibana, M.; Takahashi, H.; et al. Association of Fusobacterium nucleatum with clinical and molecular features in colorectal serrated pathway. Int. J. Cancer 2015, 137, 1258–1268. [Google Scholar] [CrossRef]
- Purcell, R.V.; Visnovska, M.; Biggs, P.J.; Schmeier, S.; Frizelle, F.A. Distinct gut microbiome patterns associate with consensus molecular subtypes of colorectal cancer. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Han, Y.W. Fusobacterium nucleatum: A commensal-turned pathogen. Curr. Opin. Microbiol. 2015, 23, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Allen-Vercoe, E.; Strauss, J.; Chadee, K. Fusobacterium nucleatum. Gut Microbes 2011, 2, 294–298. [Google Scholar] [CrossRef] [PubMed]
- Swidsinski, A.; Dorffel, Y.; Loening-Baucke, V.; Theissig, F.; Ruckert, J.C.; Ismail, M.; Rau, W.A.; Gaschler, D.; Weizenegger, M.; Kuhn, S.; et al. Acute appendicitis is characterised by local invasion with Fusobacterium nucleatum/necrophorum. Gut 2009, 60, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2014, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Irrazábal, T.; Belcheva, A.; Girardin, S.E.; Martin, A.; Philpott, D.J. The multifaceted role of the intestinal microbiota in colon cancer. Mol. Cell 2014, 54, 309–320. [Google Scholar] [CrossRef]
- Yamamura, K.; Baba, Y.; Miyake, K.; Nakamura, K.; Shigaki, H.; Mima, K.; Kurashige, J.; Ishimoto, T.; Iwatsuki, M.; Sakamoto, Y.; et al. Fusobacterium nucleatum in gastroenterological cancer: Evaluation of measurement methods using quantitative polymerase chain reaction and a literature review. Oncol. Lett. 2017, 14, 6373–6378. [Google Scholar] [CrossRef]
- McCoy, A.N.; Araújo-Pérez, F.; Azcárate-Peril, A.; Yeh, J.J.; Sandler, R.S.; Keku, T.O. Fusobacterium is associated with colorectal adenomas. PLoS ONE 2013, 8, e53653. [Google Scholar] [CrossRef]
- Kostic, A.D.; Gevers, D.; Pedamallu, C.S.; Michaud, M.; Duke, F.; Earl, A.M.; Ojesina, A.I.; Jung, J.; Bass, A.J.; Tabernero, J.; et al. Genomic analysis identifies association of Fusobacterium with colorectal carcinoma. Genome Res. 2011, 22, 292–298. [Google Scholar] [CrossRef]
- Guo, S.-H.; Wang, H.-F.; Nian, Z.-G.; Wang, Y.-D.; Zeng, Q.-Y.; Zhang, G. Immunization with alkyl hydroperoxide reductase subunit C reduces Fusobacterium nucleatum load in the intestinal tract. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Park, H.E.; Kim, J.H.; Cho, N.-Y.; Lee, H.S.; Kang, G.H. Intratumoral Fusobacterium nucleatum abundance correlates with macrophage infiltration and CDKN2A methylation in microsatellite-unstable colorectal carcinoma. Virchows Arch. 2017, 471, 329–336. [Google Scholar] [CrossRef]
- Mima, K.; Sukawa, Y.; Nishihara, R.; Qian, Z.R.; Yamauchi, M.; Inamura, K.; Kim, S.A.; Masuda, A.; Nowak, J.A.; Nosho, K.; et al. Fusobacterium nucleatumand T cells in colorectal carcinoma. Jama Oncol. 2015, 1, 653. [Google Scholar] [CrossRef] [PubMed]
- Mikuls, T.R.; Payne, J.B.; Yu, F.; Thiele, G.M.; Reynolds, R.J.; Cannon, G.W.; Markt, J.; McGowan, D.; Kerr, G.S.; Redman, R.S.; et al. Periodontitis and Porphyromonas gingivalis in patients with rheumatoid arthritis. Arthritis Rheumatol. 2014, 66, 1090–1100. [Google Scholar] [CrossRef] [PubMed]
- Velsko, I.M.; Chukkapalli, S.S.; Rivera-Kweh, M.F.; Chen, H.; Zheng, D.; Bhattacharyya, I.; Gangula, P.R.; Lucas, A.R.; Kesavalu, L. Fusobacterium nucleatum alters atherosclerosis risk factors and enhances inflammatory markers with an atheroprotective immune response in ApoE(null) mice. PLoS ONE 2015, 10, e0129795. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.F.; Li, L.F.; Guo, S.H.; Zeng, Q.Y.; Ning, F.; Liu, W.L.; Zhang, G. Evaluation of antibody level against Fusobacterium nucleatum in the serological diagnosis of colorectal cancer. Sci. Rep. 2016, 6, 33440. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Xiao, H.; Guo, S.; Xu, B.; Liao, Y.; Wu, Y.; Zhang, G. Indoleamine 2,3-dioxygenase expression regulates the survival and proliferation of Fusobacterium nucleatum in THP-1-derived macrophages. Cell Death Dis. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Wang, R.; Bhattacharya, R.; Boulbes, D.R.; Fan, F.; Xia, L.; Adoni, H.; Ajami, N.J.; Wong, M.C.; Smith, D.P.; et al. Fusobacterium nucleatum subspecies animalis influences proinflammatory cytokine expression and monocyte activation in human colorectal tumors. Cancer Prev. Res. 2017, 10, 398–409. [Google Scholar] [CrossRef] [PubMed]
- Strauss, J.; Kaplan, G.G.; Beck, P.L.; Rioux, K.; Panaccione, R.; DeVinney, R.; Lynch, T.; Allen-Vercoe, E. Invasive potential of gut mucosa-derived Fusobacterium nucleatum positively correlates with IBD status of the host. Inflamm. Bowel Dis. 2011, 17, 1971–1978. [Google Scholar] [CrossRef]
- Manson McGuire, A.; Cochrane, K.; Griggs, A.D.; Haas, B.J.; Abeel, T.; Zeng, Q.; Nice, J.B.; MacDonald, H.; Birren, B.W.; Berger, B.W.; et al. Evolution of invasion in a diverse set of Fusobacterium species. mBio 2014, 5. [Google Scholar] [CrossRef]
- Schirmer, M.; Smeekens, S.P.; Vlamakis, H.; Jaeger, M.; Oosting, M.; Franzosa, E.A.; Horst, R.t.; Jansen, T.; Jacobs, L.; Bonder, M.J.; et al. Linking the human gut microbiome to inflammatory cytokine production capacity. Cell 2016, 167, 1897. [Google Scholar] [CrossRef]
- Karin, M. Nuclear factor-κB in cancer development and progression. Nature 2006, 441, 431–436. [Google Scholar] [CrossRef]
- DiDonato, J.A.; Mercurio, F.; Karin, M. NF-κB and the link between inflammation and cancer. Immunol. Rev. 2012, 246, 379–400. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, C.W.; Ma, X.; Paranjpe, A.; Jewett, A.; Lux, R.; Kinder-Haake, S.; Shi, W. Fusobacterium nucleatum outer membrane proteins Fap2 and RadD induce cell death in human lymphocytes. Infect. Immun. 2010, 78, 4773–4778. [Google Scholar] [CrossRef] [PubMed]
- Coppenhagen-Glazer, S.; Sol, A.; Abed, J.; Naor, R.; Zhang, X.; Han, Y.W.; Bachrach, G. Fap2 of Fusobacterium nucleatum is a galactose-inhibitable adhesin involved in coaggregation, cell adhesion, and preterm birth. Infect. Immun. 2015, 83, 1104–1113. [Google Scholar] [CrossRef] [PubMed]
- Gur, C.; Ibrahim, Y.; Isaacson, B.; Yamin, R.; Abed, J.; Gamliel, M.; Enk, J.; Bar-On, Y.; Stanietsky-Kaynan, N.; Coppenhagen-Glazer, S.; et al. Binding of the Fap2 protein of Fusobacterium nucleatum to human inhibitory receptor TIGIT protects tumors from immune cell attack. Immunity 2015, 42, 344–355. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Cai, S.; Ma, Y. Association between Fusobacterium nucleatum and colorectal cancer: Progress and future directions. J. Cancer 2018, 9, 1652–1659. [Google Scholar] [CrossRef]
- Xu, M.; Yamada, M.; Li, M.; Liu, H.; Chen, S.G.; Han, Y.W. Fad A from Fusobacterium nucleatum utilizes both secreted and nonsecreted forms for functional oligomerization for attachment and invasion of host cells. J. Biol. Chem. 2007, 282, 25000–25009. [Google Scholar] [CrossRef]
- Santaolalla, R.; Sussman, D.A.; Ruiz, J.R.; Davies, J.M.; Pastorini, C.; España, C.L.; Sotolongo, J.; Burlingame, O.; Bejarano, P.A.; Philip, S.; et al. TLR4 activates the β-catenin pathway to cause intestinal neoplasia. PLoS ONE 2013, 8, e63298. [Google Scholar] [CrossRef]
- Shi, C.; Yang, Y.; Xia, Y.; Okugawa, Y.; Yang, J.; Liang, Y.; Chen, H.; Zhang, P.; Wang, F.; Han, H.; et al. Novel evidence for an oncogenic role of microRNA-21 in colitis-associated colorectal cancer. Gut 2015, 65, 1470–1481. [Google Scholar] [CrossRef]
- Chen, Y.; Peng, Y.; Yu, J.; Chen, T.; Wu, Y.; Shi, L.; Li, Q.; Wu, J.; Fu, X. Invasive Fusobacterium nucleatum activates beta-catenin signaling in colorectal cancer via a TLR4/P-PAK1 cascade. Oncotarget 2017, 8. [Google Scholar] [CrossRef]
- Bullman, S.; Pedamallu, C.S.; Sicinska, E.; Clancy, T.E.; Zhang, X.; Cai, D.; Neuberg, D.; Huang, K.; Guevara, F.; Nelson, T.; et al. Analysis of Fusobacterium persistence and antibiotic response in colorectal cancer. Science 2017, 358, 1443–1448. [Google Scholar] [CrossRef]
- Guo, S.; Li, L.; Xu, B.; Li, M.; Zeng, Q.; Xiao, H.; Xue, Y.; Wu, Y.; Wang, Y.; Liu, W.; et al. A simple and novel fecal biomarker for colorectal cancer: Ratio of Fusobacterium nucleatum to probiotics populations, based on their antagonistic effect. Clin. Chem. 2018, 64, 1327–1337. [Google Scholar] [CrossRef] [PubMed]
- Bashir, A.; Miskeen, A.Y.; Bhat, A.; Fazili, K.M.; Ganai, B.A. Fusobacterium nucleatum. Eur. J. Cancer Prev. 2015, 24, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Hussan, H.; Clinton, S.K.; Roberts, K.; Bailey, M.T. Fusobacterium’s link to colorectal neoplasia sequenced: A systematic review and future insights. World J. Gastroenterol. 2017, 23, 8626–8650. [Google Scholar] [CrossRef] [PubMed]
- Abed, J.; Emgard, J.E.M.; Zamir, G.; Faroja, M.; Almogy, G.; Grenov, A.; Sol, A.; Naor, R.; Pikarsky, E.; Atlan, K.A.; et al. Fap2 mediates Fusobacterium nucleatum colorectal adenocarcinoma enrichment by binding to tumor-expressed Gal-GalNAc. Cell Host Microbe 2016, 20, 215–225. [Google Scholar] [CrossRef]
- Abed, J.; Maalouf, N.; Parhi, L.; Chaushu, S.; Mandelboim, O.; Bachrach, G. Tumor targeting by Fusobacterium nucleatum: A pilot study and future perspectives. Front. Cell. Infect. Microbiol. 2017, 7. [Google Scholar] [CrossRef]
- Rawal, S.; Majumdar, S.; Dhawan, V.; Vohra, H. Entamoeba histolytica Gal/GalNAc lectin depletes antioxidant defences of target epithelial cells. Parasitology 2004, 128, 617–624. [Google Scholar] [CrossRef]
- Sanchez, V.; Serrano-Luna, J.; Ramirez-Moreno, E.; Tsutsumi, V.; Shibayama, M. Entamoeba histolytica: Overexpression of the gal/galnac lectin, ehcp2 and ehcp5 genes in an in vivo model of amebiasis. Parasitol. Int. 2016, 65, 665–667. [Google Scholar] [CrossRef]
- Frederick, J.R.; Petri, W.A., Jr. Roles for the galactose-/N-acetylgalactosamine-binding lectin of Entamoeba in parasite virulence and differentiation. Glycobiology 2005, 15, 53r–59r. [Google Scholar] [CrossRef]
- Burchell, J.M.; Beatson, R.; Graham, R.; Taylor-Papadimitriou, J.; Tajadura-Ortega, V. O-linked mucin-type glycosylation in breast cancer. Biochem. Soc. Trans. 2018, 46, 779–788. [Google Scholar] [CrossRef]
- Tzeng, S.-F.; Tsai, C.-H.; Chao, T.-K.; Chou, Y.-C.; Yang, Y.-C.; Tsai, M.-H.; Cha, T.-L.; Hsiao, P.-W. O-Glycosylation–mediated signaling circuit drives metastatic castration-resistant prostate cancer. FASEB J. 2018, 32, 6869–6882. [Google Scholar] [CrossRef]
- Arend, P. Early ovariectomy reveals the germline encoding of natural anti-A- and Tn-cross-reactive immunoglobulin M (IgM) arising from developmentalO-GalNAc glycosylations. (Germline-encoded natural anti-A/Tn cross-reactive IgM). Cancer Med. 2017, 6, 1601–1613. [Google Scholar] [CrossRef] [PubMed]
- Hung, J.-S.; Huang, J.; Lin, Y.-C.; Huang, M.-J.; Lee, P.-H.; Lai, H.-S.; Liang, J.-T.; Huang, M.-C. C1GALT1 overexpression promotes the invasive behavior of colon cancer cells through modifying O-glycosylation of FGFR2. Oncotarget 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Peng, R.-Q.; Wan, H.-Y.; Li, H.-F.; Liu, M.; Li, X.; Tang, H. MicroRNA-214 suppresses growth and invasiveness of cervical cancer cells by targeting UDP-N-acetyl-α-d-galactosamine: Polypeptide N-acetylgalactosaminyltransferase 7. J. Biol. Chem. 2012, 287, 14301–14309. [Google Scholar] [CrossRef] [PubMed]
- Chugh, S.; Barkeer, S.; Rachagani, S.; Nimmakayala, R.K.; Perumal, N.; Pothuraju, R.; Atri, P.; Mahapatra, S.; Thapa, I.; Talmon, G.A.; et al. Disruption of C1galt1 gene promotes development and metastasis of pancreatic adenocarcinomas in mice. Gastroenterology 2018, 155, 1608–1624. [Google Scholar] [CrossRef]
- Jiang, X.-N.; Dang, Y.-F.; Gong, F.-L.; Guo, X.-L. Role and regulation mechanism of Gal-3 in non-small cell lung cancer and its potential clinical therapeutic significance. Chem. Biol. Interact. 2019, 309, 108724. [Google Scholar] [CrossRef]
- Tian, Y.; Denda-Nagai, K.; Kamata-Sakurai, M.; Nakamori, S.; Tsukui, T.; Itoh, Y.; Okada, K.; Yi, Y.; Irimura, T. Mucin 21 in esophageal squamous epithelia and carcinomas: Analysis with glycoform-specific monoclonal antibodies. Glycobiology 2012, 22, 1218–1226. [Google Scholar] [CrossRef]
- Han, Y.W.; Ikegami, A.; Rajanna, C.; Kawsar, H.I.; Zhou, Y.; Li, M.; Sojar, H.T.; Genco, R.J.; Kuramitsu, H.K.; Deng, C.X. Identification and characterization of a novel adhesin unique to oral Fusobacteria. J. Bacteriol. 2005, 187, 5330–5340. [Google Scholar] [CrossRef]
- Wu, J.; Li, Q.; Fu, X. Fusobacterium nucleatum contributes to the carcinogenesis of colorectal cancer by inducing inflammation and suppressing host immunity. Transl. Oncol. 2019, 12, 846–851. [Google Scholar] [CrossRef]
- Ashare, A.; Stanford, C.; Hancock, P.; Stark, D.; Lilli, K.; Birrer, E.; Nymon, A.; Doerschug, K.C.; Hunninghake, G.W. Chronic liver disease impairs bacterial clearance in a human model of induced Bacteremia. Clin. Transl. Sci. 2009, 2, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Ghidini, M.; Petrillo, A.; Salati, M.; Khakoo, S.; Varricchio, A.; Tomasello, G.; Grossi, F.; Petrelli, F. Surgery or locoregional approaches for hepatic oligometastatic pancreatic cancer: Myth, hope, or reality? Cancers 2019, 11, 1095. [Google Scholar] [CrossRef]
- Spałek, M.; Michalski, K.; Bujko, K.; Wyrwicz, L. Association between preoperative pelvic irradiation and toxicity of subsequent chemotherapy in rectal cancer. Oncol. Res. Treat. 2019, 42, 497–504. [Google Scholar] [CrossRef]
- Mitchell, D.; Puckett, Y.; Nguyen, Q.N. Literature review of current management of colorectal liver metastasis. Cureus 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Brennan, C.A.; Garrett, W.S. Gut Microbiota, inflammation, and colorectal cancer. Annu. Rev. Microbiol. 2016, 70, 395–411. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.H.; Min, J.-J. Salmonella-mediated cancer therapy: Roles and potential. Nucl. Med. Mol. Imaging 2016, 51, 118–126. [Google Scholar] [CrossRef]
- Chowdhury, S.; Castro, S.; Coker, C.; Hinchliffe, T.E.; Arpaia, N.; Danino, T. Programmable bacteria induce durable tumor regression and systemic antitumor immunity. Nat. Med. 2019, 25, 1057–1063. [Google Scholar] [CrossRef]
- Dougan, M.; Dougan, S.K. Programmable bacteria as cancer therapy. Nat. Med. 2019, 25, 1030–1031. [Google Scholar] [CrossRef]
- Wu, M.-R.; Jusiak, B.; Lu, T.K. Engineering advanced cancer therapies with synthetic biology. Nat. Rev. Cancer 2019, 19, 187–195. [Google Scholar] [CrossRef]
- Chien, T.; Doshi, A.; Danino, T. Advances in bacterial cancer therapies using synthetic biology. Curr. Opin. Syst. Biol. 2017, 5, 1–8. [Google Scholar] [CrossRef]
- Lim, W.A.; June, C.H. The principles of engineering immune cells to treat cancer. Cell 2017, 168, 724–740. [Google Scholar] [CrossRef]
- Harimoto, T.; Singer, Z.S.; Velazquez, O.S.; Zhang, J.; Castro, S.; Hinchliffe, T.E.; Mather, W.; Danino, T. Rapid Screening of Engineered Microbial Therapies in a 3-D Multicellular Model; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY, USA, 2018. [Google Scholar] [CrossRef]
- Yoon, W.; Yoo, Y.; Chae, Y.S.; Kee, S.H.; Kim, B.M. Therapeutic advantage of genetically engineered Salmonella typhimurium carrying short hairpin RNA against inhibin alpha subunit in cancer treatment. Ann. Oncol. 2018, 29, 2010–2017. [Google Scholar] [CrossRef]
- Panthel, K.; Meinel, K.M.; Sevil Domènech, V.E.; Geginat, G.; Linkemann, K.; Busch, D.H.; Rüssmann, H. Prophylactic anti-tumor immunity against a murine fibrosarcoma triggered by the Salmonella type III secretion system. Microbes Infect. 2006, 8, 2539–2546. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.H.; Kim, H.S.; Ha, J.M.; Hong, Y.; Choy, H.E.; Min, J.J. Genetically engineered Salmonella typhimurium as an imageable therapeutic probe for cancer. Cancer Res. 2009, 70, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Ryan, R.M.; Green, J.; Williams, P.J.; Tazzyman, S.; Hunt, S.; Harmey, J.H.; Kehoe, S.C.; Lewis, C.E. Bacterial delivery of a novel cytolysin to hypoxic areas of solid tumors. Gene Ther. 2009, 16, 329–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loeffler, M.; Le’Negrate, G.; Krajewska, M.; Reed, J.C. Salmonella typhimurium engineered to produce CCL21 inhibit tumor growth. Cancer Immunol. Immunother. 2008, 58, 769–775. [Google Scholar] [CrossRef]
- Loeffler, M.; Le’Negrate, G.; Krajewska, M.; Reed, J.C. Attenuated Salmonella engineered to produce human cytokine LIGHT inhibit tumor growth. Proc. Natl. Acad. Sci. USA 2007, 104, 12879–12883. [Google Scholar] [CrossRef]
- Xiang, R.; Mizutani, N.; Luo, Y.; Chiodoni, C.; Zhou, H.; Mizutani, M.; Ba, Y.; Becker, J.C.; Reisfeld, R.A. A DNA vaccine targeting survivin combines apoptosis with suppression of angiogenesis in lung tumor eradication. Cancer Res. 2005, 65, 553–561. [Google Scholar]
- Agorio, C.; Schreiber, F.; Sheppard, M.; Mastroeni, P.; Fernandez, M.; Martinez, M.A.; Chabalgoity, J.A. Live attenuated Salmonella as a vector for oral cytokine gene therapy in melanoma. J. Gene Med. 2007, 9, 416–423. [Google Scholar] [CrossRef]
- Wen, L.J.; Gao, L.F.; Jin, C.S.; Zhang, H.J.; Ji, K.; Yang, J.P.; Zhao, X.J.; Wen, M.J.; Guan, G.F. Small interfering RNA survivin and GRIM-19 co-expression salmonella plasmid inhibited the growth of laryngeal cancer cells in vitro and in vivo. Int. J. Clin. Exp. Pathol. 2013, 6, 2071–2081. [Google Scholar]
- Liu, Y.-B.; Zhang, L.; Guo, Y.-X.; Gao, L.-F.; Liu, X.-C.; Zhao, L.-J.; Guo, B.-F.; Zhao, L.-J.; Zhao, X.-J.; Xu, D.-Q. Plasmid-based Survivin shRNA and GRIM-19 carried by attenuated Salmonella suppresses tumor cell growth. Asian J. Androl. 2012, 14, 536–545. [Google Scholar] [CrossRef] [Green Version]
- Li, Z. Anti-angiogenesis and anticancer effects of a plasmid expressing both ENDO-VEGI151 and small interfering RNA against survivin. Int. J. Mol. Med. 2011, 29, 485–490. [Google Scholar] [CrossRef]
- Nishikawa, H. In vivo antigen delivery by a Salmonella typhimurium type III secretion system for therapeutic cancer vaccines. J. Clin. Investig. 2006, 116, 1946–1954. [Google Scholar] [CrossRef] [PubMed]
- Loeffler, M.; Le’Negrate, G.; Krajewska, M.; Reed, J.C. Inhibition of tumor growth using Salmonella expressing Fas ligand. JNCI J. Natl. Cancer Inst. 2008, 100, 1113–1116. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Chu, L.; Han, X.; Liu, X.; Ren, D. Synergistic antitumoral effects of human telomerase reverse transcriptase-mediated dual-apoptosis-related gene vector delivered by orally attenuated Salmonella enterica Serovar Typhimurium in murine tumor models. J. Gene Med. 2008, 10, 690–701. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.-S.; Nitcheu-Tefit, J.; Alcock, S.; Ramirez-Jimenez, F.; Chao, T.-Y.; Baril, P.; Rocha, M.; Brett, S.J.; Stauss, H.J.; Vassaux, G. Development of an Escherichia coli expressing Listeriolysin-O vaccine against wilms tumor gene 1-expressing tumors. J. Immunother. 2009, 32, 845–855. [Google Scholar] [CrossRef]
- Yang, N.; Zhu, X.; Chen, L.; Li, S.; Ren, D. Oral administration of attenuated S. typhimurium carrying shRNA-expressing vectors as a cancer therapeutic. Cancer Biol. Ther. 2008, 7, 145–151. [Google Scholar] [CrossRef]
- Li, X.; Li, Y.; Hu, J.; Wang, B.; Zhao, L.; Ji, K.; Guo, B.; Yin, D.; Du, Y.; Kopecko, D.J.; et al. Plasmid-based E6-specific siRNA and co-expression of wild-type p53 suppresses the growth of cervical cancer in vitro and in vivo. Cancer Lett. 2013, 335, 242–250. [Google Scholar] [CrossRef] [Green Version]
- Sorenson, B.S.; Banton, K.L.; Frykman, N.L.; Leonard, A.S.; Saltzman, D.A. Attenuated Salmonella typhimurium with interleukin 2 gene prevents the establishment of pulmonary metastases in a model of osteosarcoma. J. Pediatric Surg. 2008, 43, 1153–1158. [Google Scholar] [CrossRef]
- Jiang, T.; Zhou, C.; Gu, J.; Liu, Y.; Zhao, L.; Li, W.; Wang, G.; Li, Y.; Cai, L. Enhanced therapeutic effect of cisplatin on the prostate cancer in tumor-bearing mice by transfecting the attenuated Salmonella carrying a plasmid co-expressing p53 gene and mdm2 siRNA. Cancer Lett. 2013, 337, 133–142. [Google Scholar] [CrossRef]
- Ganai, S.; Arenas, R.B.; Forbes, N.S. Tumour-targeted delivery of TRAIL using Salmonella typhimurium enhances breast cancer survival in mice. Br. J. Cancer 2009, 101, 1683–1691. [Google Scholar] [CrossRef]
- Flentie, K.; Kocher, B.; Gammon, S.T.; Novack, D.V.; McKinney, J.S.; Piwnica-Worms, D. A Bioluminescent transposon reporter-trap identifies tumor-specific microenvironment-induced promoters in Salmonella for conditional bacterial-based tumor therapy. Cancer Discov. 2012, 2, 624–637. [Google Scholar] [CrossRef]
- Tian, Y.; Guo, B.; Jia, H.; Ji, K.; Sun, Y.; Li, Y.; Zhao, T.; Gao, L.; Meng, Y.; Kalvakolanu, D.V.; et al. Targeted therapy via oral administration of attenuated Salmonella expression plasmid-vectored Stat3-shRNA cures orthotopically transplanted mouse HCC. Cancer Gene Ther. 2012, 19, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Gao, L.; Zhao, L.; Guo, B.; Ji, K.; Tian, Y.; Wang, J.; Yu, H.; Hu, J.; Kalvakolanu, D.V.; et al. Intratumoral delivery and suppression of prostate tumor growth by attenuated Salmonella enterica serovar typhimurium carrying plasmid-based small interfering RNAs. Cancer Res. 2007, 67, 5859–5864. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, Y.; Wang, B.; Ji, K.; Liang, Z.; Guo, B.; Hu, J.; Yin, D.; Du, Y.; Kopecko, D.J.; et al. Delivery of the co-expression plasmid pEndo-Si-Stat3 by attenuated Salmonella serovar typhimurium for prostate cancer treatment. J. Cancer Res. Clin. Oncol. 2013, 139, 971–980. [Google Scholar] [CrossRef] [PubMed]
- Fensterle, J.; Bergmann, B.; Yone, C.L.R.P.; Hotz, C.; Meyer, S.R.; Spreng, S.; Goebel, W.; Rapp, U.R.; Gentschev, I. Cancer immunotherapy based on recombinant Salmonella enterica serovar Typhimurium aroA strains secreting prostate-specific antigen and cholera toxin subunit B. Cancer Gene Ther. 2007, 15, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Loeffler, M.; Le’Negrate, G.; Krajewska, M.; Reed, J.C. IL-18-producing Salmonella inhibit tumor growth. Cancer Gene Ther. 2008, 15, 787–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niethammer, A.G.; Xiang, R.; Becker, J.C.; Wodrich, H.; Pertl, U.; Karsten, G.; Eliceiri, B.P.; Reisfeld, R.A. A DNA vaccine against VEGF receptor 2 prevents effective angiogenesis and inhibits tumor growth. Nat. Med. 2002, 8, 1369–1375. [Google Scholar] [CrossRef]
- Green, E.R.; Mecsas, J. Bacterial secretion systems: An overview. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef]
- Wu, X.Y.; Chapman, T.; Trott, D.J.; Bettelheim, K.; Do, T.N.; Driesen, S.; Walker, M.J.; Chin, J. Comparative analysis of virulence genes, genetic diversity, and phylogeny of commensal and enterotoxigenic Escherichia coli isolates from weaned pigs. Appl. Environ. Microbiol. 2007, 73, 83–91. [Google Scholar] [CrossRef]
- Luo, X.; Li, Z.; Lin, S.; Le, T.; Ittensohn, M.; Bermudes, D.; Runyab, J.D.; Shen, S.y.; Chen, J.; King, I.C.; et al. Antitumor effect of VNP20009, an attenuated Salmonella, in murine tumor models. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2001, 12, 501–508. [Google Scholar] [CrossRef]
- Sznol, M.; Lin, S.L.; Bermudes, D.; Zheng, L.-m.; King, I. Use of preferentially replicating bacteria for the treatment of cancer. J. Clin. Investig. 2000, 105, 1027–1030. [Google Scholar] [CrossRef] [Green Version]
- Hashemi Goradel, N.; Heidarzadeh, S.; Jahangiri, S.; Farhood, B.; Mortezaee, K.; Khanlarkhani, N.; Negahdari, B. Fusobacterium nucleatum and colorectal cancer: A mechanistic overview. J. Cell. Physiol. 2018, 234, 2337–2344. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Migration of F. nucleatum from the oral cavity to colon that promote colorectal cancer (CRC).
Figure 1.
Migration of F. nucleatum from the oral cavity to colon that promote colorectal cancer (CRC).

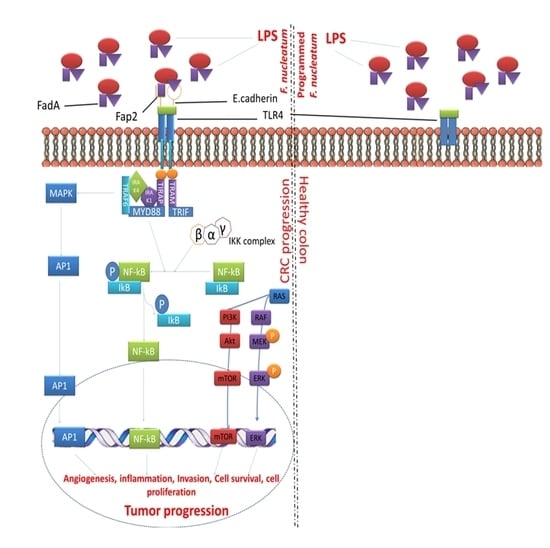
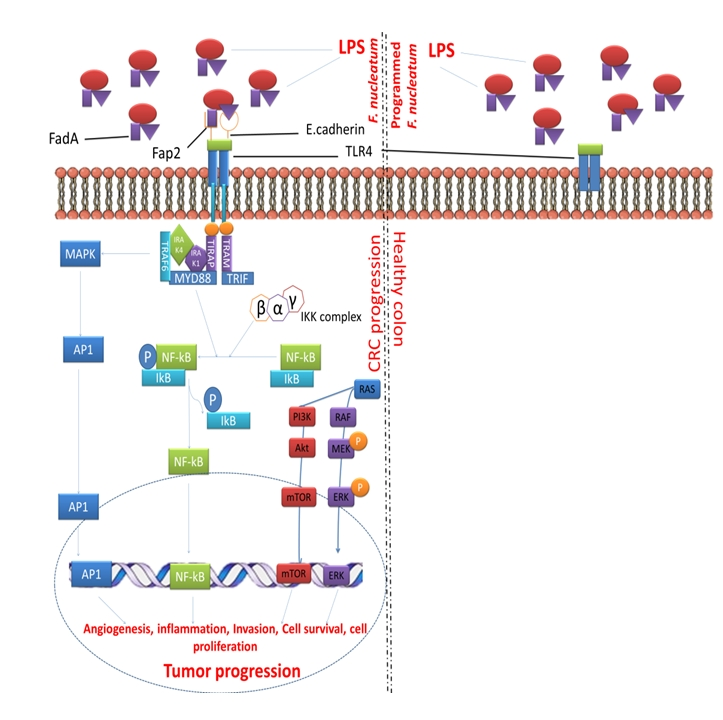
Figure 2.
Programmed/attenuated F. nucleatum Fap2 as a potent factor for the treatment of CRC. Abbreviation: Akt-protein kinase B; Ap1-activating protein-1; ERK-extracellular signal-regulated kinase; FadA- Fusobacterium adhesin A; Fap2-fibroblast activation protein 2; Gal-GalNAc-acetylgalactosamine; IkB-Ikappa B proteins; IKK-Ikappa B kinase; IRAK1-Interleukin-1; receptor-associated kinase 1; IRAK4-Interleukin-1 receptor-associated kinase 4; LPS-lipopolysaccharide; MAPK-mitogen-activated protein kinase; MEK-MAPK-ERK-kinase; mTOR-mammalian target of rapamycin; MYD88-myeloid differentiation primary response gene 88; NF-kB-nuclear factor kappa-B; p-phosphorylated; PI3K-phosphatidylinositol 3-kinase; RAF-rapidly accelerated fibrosarcoma; RAS-rat sarcoma; TLR4-Toll-like receptor 4; TRAF6-Tumor necrosis factor receptor associated factor 6; TRAM-Transverse rectus abdominis myocutaneous; TRAP-thyroid hormone receptor associated protein.
Figure 2.
Programmed/attenuated F. nucleatum Fap2 as a potent factor for the treatment of CRC. Abbreviation: Akt-protein kinase B; Ap1-activating protein-1; ERK-extracellular signal-regulated kinase; FadA- Fusobacterium adhesin A; Fap2-fibroblast activation protein 2; Gal-GalNAc-acetylgalactosamine; IkB-Ikappa B proteins; IKK-Ikappa B kinase; IRAK1-Interleukin-1; receptor-associated kinase 1; IRAK4-Interleukin-1 receptor-associated kinase 4; LPS-lipopolysaccharide; MAPK-mitogen-activated protein kinase; MEK-MAPK-ERK-kinase; mTOR-mammalian target of rapamycin; MYD88-myeloid differentiation primary response gene 88; NF-kB-nuclear factor kappa-B; p-phosphorylated; PI3K-phosphatidylinositol 3-kinase; RAF-rapidly accelerated fibrosarcoma; RAS-rat sarcoma; TLR4-Toll-like receptor 4; TRAF6-Tumor necrosis factor receptor associated factor 6; TRAM-Transverse rectus abdominis myocutaneous; TRAP-thyroid hormone receptor associated protein.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Detection of F. nucleatum in colorectal cancer (CRC) patients.
| Sample Tested | Total Number of Clinical Samples | Detection Method | Positive Percentage | References |
|---|---|---|---|---|
| FFPE tissue | 6 | 16S rRNA | 32 | [32] |
| FFPE tissue | 6 | 16S rRNA | 100 | [33] |
| FFPE tissue | 8 | 16S rRNA | 100 | [34] |
| Feces | 14 | qPCR | 57 | [35] |
| FFPE tissue | 31 | 16S rRNA | 10 | [36] |
| FFPE tissue | 37 | 16S rRNA | 9 | [37] |
| FFPE tissue | 44 | 16S rRNA | 100 | [38] |
| FFPE tissue | 46 | 16S rRNA | 100 | [39] |
| FFPE tissue | 46 | 16S rRNA | 54 | [40] |
| FFPE tissue | 47 | 16S rRNA | 32 | [41] |
| FFPE tissue | 52 | 16S rRNA | 77 | [42] |
| Feces | 72 | qPCR | 64 | [43] |
| FFPE tissue | 97 | 16S rRNA | 72 | [14] |
| Frozen tissue and FFPE tissue | 101 | FISH and FQ-PCR | 87 | [44] |
| Genomic DNA | 149 | qPCR | 74 | [45] |
| Feces | 158 | ddPCR | 54 | [46] |
| FFPE tissue | 309 | qPCR | 34 | [47] |
| FFPE tissue | 511 | qPCR | 9 | [48] |
| FFPE tissue | 504 | qPCR | 56 | [49] |
| FFPE tissue | 598 | qPCR | 13 | [26] |
Abbreviation: ddPCR: droplet digital polymerase chain reaction; FFPE: formalin-fixed paraffin-embedded; FISH: fluorescence in situ hybridization; rRNA: ribosomal ribonucleic acid; FQ-PCR: fluorescent quantitative polymerase chain reaction; qPCR: quantitative real-time polymerase chain reaction.
Table 2.
Programmed bacteria expressed as anti-tumor agents in various animal models.
| Bacterial Species | Agent(s) | Host | Origin of the Tumor | Tumor(s) | Effector(s) | Results | References |
|---|---|---|---|---|---|---|---|
| S. typhimurium | Bacterial antigen S. typhimurium secreting L. monocytogenes Iap217–225 (Lm-p60) | BALB/c | Bones | WEHI-164 (Fibrosarcoma) cells expressing Lm-p60 | CD8+ cell-mediated | Antigen-specific tumor inhibition | [112] |
| S. typhimurium | Bacterial toxin S. typhimurium secreting HlyE | BALB/c | Breast | CT-26, 4T1 | Not reported | Reduction in tumor mass | [113,114] |
| S. typhimurium | Birc5 (Survivin) | C57BL/6 | Lungs | D121 | CD8+ cell-mediated | Suppression of angiogenesis and pulmonary metastasized tumors | [117] |
| S. typhimurium | BIRC5 shRNA NDUFA13 (GRIM-19) | Nude Mice | Larynx, prostate | Hep-2 (Laryngeal cancer) DU145 (PC-Xenograft) | Apoptosis | Tumor growth reduced | [119,120] |
| S. typhimurium | BIRC5 shRNA TNFSF15 (VEGI) | Nude mice | Breast | MDA-MB-231 (BC-Xenograft) | Apoptosis | Tumor growth reduced | [121] |
| S. typhimurium | ccl21 | BALB/C | Breast | D2F2, CT-26 | CD4+ and CD8+ cell mediated | Tumor-limited inflammatory reaction with a substantial reduction in tumor burden | [115] |
| S. typhimurium | CTAG1B (NY-ESO-1) | BALB/c | Skin | CMS5 cells expressing human NY-ESO-1 | CD8+ cell-mediated | NY-ESO-1-positive tumors are eliminated | [122] |
| S. typhimurium | Cytokine ccl21 | C57BL/6 | Lungs | D121 (LC-Syngeneic) | CD8+ cell mediated | Suppression of angiogenesis and growth of pulmonary metastasized tumors | [117] |
| S. typhimurium | Death inducer S. typhimurium secreting murine Fasl | BALB/c | Breast, colon | CT-26 D2F2 (BC-Syngeneic) | Neutrophils | Reduction in tumor mass | [123] |
| S. typhimurium | Diablo/Trail | BALB/c C57BL/6 | Liver, spleen, kidney | 4T1 LL/2 (LC-Syngeneic) B16F10 (Melanoma) | Apoptosis | Tumor growth inhibition with prolonged survival | [124] |
| E. coli | E. coli expressing LLO | C57BL/6 | Blood | MBL2 (Leukemia-Syngeneic) TRAMP-C (PC-Syngeneic) | CD8+ cell-mediated | Reduction in tumor mass | [125] |
| S. typhimurium | Growth inhibitor(s) Bcl2 shRNA | C57BL/6 | Liver, spleen, skin | B16F10 | Apoptosis | Survival time of tumor-bearing mice Prolonged Complete tumor regression not observed | [126] |
| S. typhimurium | HPV E6 shRNA TP53 | Nude mice | Cervix | SiHa | Apoptosis | Tumor growth reduced | [127] |
| S. typhimurium | IL 18 | C57BL/6 | Skin | B16F1A (Melanoma) | Not reported | Increased survival time | [118] |
| BALB/C | Skin, colon | D2F2, CT-26 | Granulocyte, NK, CD4+, CD8+ cell mediated | Reduced tumor growth and pulmonary metastases | [111] | ||
| S. typhimurium | IL2 | C57BL/6 | Liver | MCA-38 (Adenocarcinoma Syngeneic) | NK cells | Hepatic metastases reduced | [116] |
| BALB/C | Bone, lungs | K7M2 (Osteosarcoma –Syngeneic) | NK cells | Pulmonary metastases reduced compared to saline control | [128] | ||
| S. typhimurium | IL4 | C57BL/6 | Skin | B16F1A (Melanoma) | Not reported | Increased survival time | [118] |
| MDM2 shRNA TP53-Cisplatin | Nude Mice | Prostate | PC3 | Apoptosis | Tumor growth reduced | [129] | |
| S. typhimurium | S. typhimurium secreting murine Trail | BALB/c | Breast | 4T1 | Apoptosis | Tumor growth reduced | [130] |
| S. typhimurium | S. typhimurium secreting Stx2 | Nude Mice | Skin, colon | B16, HCT116, HeLa | Necrosis | Reduction in tumor mass | [131] |
| S. typhimurium | Stat3 shRNA | C57BL/6 | Bone | H22 | Apoptosis and CD8+ cell mediated | Tumor growth reduced | [132] |
| S. typhimurium | Stat3 shRNA Col18A1Endo | C57BL/6 | Prostate | RM1 | Apoptosis | Tumor growth reduced | [133,134] |
| S. typhimurium | Target antigen KLK3 (PSA) | DBA/2 | Prostate | P815 cells expressing human PSA | CD8+ cell-mediated | Direct i.m. DNA vaccination was better than Serovar typhimurium-delivered immunogen | [135] |
| S. typhimurium | Tnfsf14 (LIGHT) | BALB/C | Breast | D2F2, CT-26 | NK, CD4+, CD8+ cell- mediated | Primary and metastatic tumor growth inhibited | [136] |
| S. typhimurium | Vegfr2 (Kdr or Flk1) full-length Protein | BALB/c C57BL/6 | Skin | CT-26 B16G3.26 (Melanoma) D121 MC-38 (CRC-Syngeneic) | CD8+ cell-mediated | Microvessel destruction retarded tumor growth and metastases. Healing of skin wounds slightly delayed. Immunological memory persisted at 120 days post-immunization | [137] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ganesan, K.; Guo, S.; Fayyaz, S.; Zhang, G.; Xu, B. Targeting Programmed Fusobacterium nucleatum Fap2 for Colorectal Cancer Therapy. Cancers 2019, 11, 1592. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11101592
AMA Style
Ganesan K, Guo S, Fayyaz S, Zhang G, Xu B. Targeting Programmed Fusobacterium nucleatum Fap2 for Colorectal Cancer Therapy. Cancers. 2019; 11(10):1592. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11101592
Chicago/Turabian StyleGanesan, Kumar, Songhe Guo, Sundaz Fayyaz, Ge Zhang, and Baojun Xu. 2019. "Targeting Programmed Fusobacterium nucleatum Fap2 for Colorectal Cancer Therapy" Cancers 11, no. 10: 1592. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11101592
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.