The Role of Non-Coding RNAs in Uveal Melanoma


Abstract
:Simple Summary
Abstract
1. Introduction
2. MicroRNAs in UM
2.1. Dysregulated Expression of MicroRNAs
2.1.1. miRNA Tumor Suppressors
2.1.2. Oncogenic miRNAs
2.2. miRNAs in Clinical Applications
2.2.1. miRNAs as Potential Biomarkers for UM
Studies on Tissue Samples/Cell Lines
miRNAs in Serum or Plasma
2.2.2. miRNAs as Immune Regulators
3. Long Non-Coding RNAs in UM
3.1. Plasmacytoma Variant Translocation 1 (PVT1)
3.2. CASC15-New-Transcript 1
3.3. Metastasis-Associated Lung Adenocarcinoma Transcript 1 (MALAT1)
3.4. Homeobox A11 Antisense (HOXA11-AS)
3.5. Small Nucleolar RNA Host Gene 7 (SNHG7)
3.6. Ferrtin Heavy-Chain 1 Pseudogene 3 (FTH1P3)
3.7. Purinergic Receiver P2X7 Variant3 (P2RX7-V3)
3.8. Rhophilin RHO Gtpase Binding Protein 1 Antisense 1 (RHPN1-AS1)
3.9. Pax6 Upstream Antisense RNA (PAUPAR)
3.10. Other lncRNAs
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hattori, M. Finishing the euchromatic sequence of the human genome. Tanpakushitsu Kakusan Koso 2005, 50, 162–168. [Google Scholar] [PubMed]
- Consortium, E.P. An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 489, 57–74. [Google Scholar] [CrossRef] [PubMed]
- Kapranov, P.; Drenkow, J.; Cheng, J.; Long, J.; Helt, G.; Dike, S.; Gingeras, T.R. Examples of the complex architecture of the human transcriptome revealed by RACE and high-density tiling arrays. Genome Res. 2005, 15, 987–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taft, R.J.; Pang, K.C.; Mercer, T.R.; Dinger, M.; Mattick, J.S. Non-coding RNAs: Regulators of disease. J. Pathol. 2010, 220, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Guzel, E.; Okyay, T.M.; Yalcinkaya, B.; Karacaoglu, S.; Gocmen, M.; Akcakuyu, M.H. Tumor suppressor and oncogenic role of long non-coding RNAs in cancer. North. Clin. Istanb. 2020, 7, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Staby, K.M.; Gravdal, K.; Mork, S.J.; Heegaard, S.; Vintermyr, O.K.; Krohn, J. Prognostic impact of chromosomal aberrations and GNAQ, GNA11 and BAP1 mutations in uveal melanoma. Acta Ophthalmol. 2018, 96, 31–38. [Google Scholar] [CrossRef]
- Damato, B.; Duke, C.; Coupland, S.E.; Hiscott, P.; Smith, P.A.; Campbell, I.; Douglas, A.; Howard, P. Cytogenetics of uveal melanoma: A 7-year clinical experience. Ophthalmology 2007, 114, 1925–1931. [Google Scholar] [CrossRef]
- Shain, A.H.; Bagger, M.M.; Yu, R.; Chang, D.; Liu, S.; Vemula, S.; Weier, J.F.; Wadt, K.; Heegaard, S.; Bastian, B.C.; et al. The genetic evolution of metastatic uveal melanoma. Nat. Genet. 2019, 51, 1123–1130. [Google Scholar] [CrossRef]
- Rodrigues, M.J.; Stern, M.H. Genetic landscape of uveal melanoma. J. Fr. D’ophtalmologie 2015, 38, 522–525. [Google Scholar] [CrossRef]
- Friedman, R.C.; Farh, K.K.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Croce, C.M. The role of MicroRNAs in human cancer. Signal Transduct. Target. Ther. 2016, 1, 15004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liz, J.; Esteller, M. lncRNAs and microRNAs with a role in cancer development. Biochimica et Biophysica Acta 2016, 1859, 169–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, D.D.; Zhao, X.; Li, W.T. Identification of differentially expressed metastatic genes and their signatures to predict the overall survival of uveal melanoma patients by bioinformatics analysis. Int. J. Ophthalmol. 2020, 13, 1046–1053. [Google Scholar] [CrossRef] [PubMed]
- Griewank, K.G.; Yu, X.; Khalili, J.; Sozen, M.M.; Stempke-Hale, K.; Bernatchez, C.; Wardell, S.; Bastian, B.C.; Woodman, S.E. Genetic and molecular characterization of uveal melanoma cell lines. Pigment Cell Melanoma Res. 2012, 25, 182–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nareyeck, G.; Zeschnigk, M.; Prescher, G.; Lohmann, D.R.; Anastassiou, G. Establishment and characterization of two uveal melanoma cell lines derived from tumors with loss of one chromosome 3. Exp. Eye Res. 2006, 83, 858–864. [Google Scholar] [CrossRef] [PubMed]
- Jager, M.J.; Magner, J.A.; Ksander, B.R.; Dubovy, S.R. Uveal Melanoma Cell Lines: Where do they come from? (An American Ophthalmological Society Thesis). Trans. Am. Ophthalmol. Soc. 2016, 114, T5. [Google Scholar]
- Yu, X.; Ambrosini, G.; Roszik, J.; Eterovic, A.K.; Stempke-Hale, K.; Seftor, E.A.; Chattopadhyay, C.; Grimm, E.; Carvajal, R.D.; Hendrix, M.J.; et al. Genetic analysis of the ‘uveal melanoma’ C918 cell line reveals atypical BRAF and common KRAS mutations and single tandem repeat profile identical to the cutaneous melanoma C8161 cell line. Pigment Cell Melanoma Res. 2015, 28, 357–359. [Google Scholar] [CrossRef]
- Amirouchene-Angelozzi, N.; Nemati, F.; Gentien, D.; Nicolas, A.; Dumont, A.; Carita, G.; Camonis, J.; Desjardins, L.; Cassoux, N.; Piperno-Neumann, S.; et al. Establishment of novel cell lines recapitulating the genetic landscape of uveal melanoma and preclinical validation of mTOR as a therapeutic target. Mol. Oncol. 2014, 8, 1508–1520. [Google Scholar] [CrossRef]
- Luyten, G.P.; Naus, N.C.; Mooy, C.M.; Hagemeijer, A.; Kan-Mitchell, J.; Van Drunen, E.; Vuzevski, V.; De Jong, P.T.; Luider, T.M. Establishment and characterization of primary and metastatic uveal melanoma cell lines. Int. J. Cancer 1996, 66, 380–387. [Google Scholar] [CrossRef]
- Liu, N.; Sun, Q.; Chen, J.; Li, J.; Zeng, Y.; Zhai, S.; Li, P.; Wang, B.; Wang, X. MicroRNA-9 suppresses uveal melanoma cell migration and invasion through the NF-kappaB1 pathway. Oncol. Rep. 2012, 28, 961–968. [Google Scholar] [CrossRef]
- Wu, S.; Chen, H.; Han, N.; Zhang, C.; Yan, H. Long Noncoding RNA PVT1 Silencing Prevents the Development of Uveal Melanoma by Impairing MicroRNA-17-3p-Dependent MDM2 Upregulation. Investig. Ophthalmol. Vis. Sci. 2019, 60, 4904–4914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Luo, Y.; Guan, W.; Zhao, H. Role of miR-23a/Zeb1 negative feedback loop in regulating epithelial-mesenchymal transition and tumorigenicity of intraocular tumors. Oncol. Lett. 2018, 16, 2462–2470. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Zhou, X.; Chen, X.; Hu, D.N.; Dong, X.D.; Wang, J.; Lu, F.; Tu, L.; Qu, J. MicroRNA-34a inhibits uveal melanoma cell proliferation and migration through downregulation of c-Met. Investig. Ophthalmol. Vis. Sci. 2009, 50, 1559–1565. [Google Scholar] [CrossRef] [PubMed]
- Hou, Q.; Han, S.; Yang, L.; Chen, S.; Chen, J.; Ma, N.; Wang, C.; Tang, J.; Chen, X.; Chen, F.; et al. The Interplay of MicroRNA-34a, LGR4, EMT-Associated Factors, and MMP2 in Regulating Uveal Melanoma Cells. Investig. Ophthalmol. Vis. Sci. 2019, 60, 4503–4510. [Google Scholar] [CrossRef] [Green Version]
- Dong, F.; Lou, D. MicroRNA-34b/c suppresses uveal melanoma cell proliferation and migration through multiple targets. Mol. Vis. 2012, 18, 537–546. [Google Scholar]
- Amaro, A.; Croce, M.; Ferrini, S.; Barisione, G.; Gualco, M.; Perri, P.; Pfeffer, U.; Jager, M.J.; Coupland, S.E.; Mosci, C.; et al. Potential Onco-Suppressive Role of miR122 and miR144 in Uveal Melanoma through ADAM10 and C-Met Inhibition. Cancers 2020, 12, 1468. [Google Scholar] [CrossRef]
- Chen, X.; He, D.; Dong, X.D.; Dong, F.; Wang, J.; Wang, L.; Tang, J.; Hu, D.N.; Yan, D.; Tu, L. MicroRNA-124a is epigenetically regulated and acts as a tumor suppressor by controlling multiple targets in uveal melanoma. Investig. Ophthalmol. Vis. Sci. 2013, 54, 2248–2256. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Wang, J.; Shen, H.; Lu, J.; Li, C.; Hu, D.N.; Dong, X.D.; Yan, D.; Tu, L. Epigenetics, microRNAs, and carcinogenesis: Functional role of microRNA-137 in uveal melanoma. Investig. Ophthalmol. Vis. Sci. 2011, 52, 1193–1199. [Google Scholar] [CrossRef]
- Peng, D.; Dong, J.; Zhao, Y.; Peng, X.; Tang, J.; Chen, X.; Wang, L.; Hu, D.N.; Reinach, P.S.; Qu, J.; et al. miR-142-3p suppresses uveal melanoma by targeting CDC25C, TGFbetaR1, GNAQ, WASL, and RAC1. Cancer Manag. Res. 2019, 11, 4729–4742. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Bian, G.; Meng, Z.; Dang, G.; Shi, D.; Mi, S. MiR-144 Inhibits Uveal Melanoma Cell Proliferation and Invasion by Regulating c-Met Expression. PLoS ONE 2015, 10, e0124428. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Luo, J.T.; Liu, Y.M.; Wei, W.B. miRNA-145/miRNA-205 inhibits proliferation and invasion of uveal melanoma cells by targeting NPR1/CDC42. Int. J. Ophthalmol. 2020, 13, 718–724. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Dong, X.D.; Chen, X.; Yao, S.; Wang, L.; Wang, J.; Wang, C.; Hu, D.N.; Qu, J.; Tu, L. Role of microRNA-182 in posterior uveal melanoma: Regulation of tumor development through MITF, BCL2 and cyclin D2. PLoS ONE 2012, 7, e40967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Huo, Y.; Wang, D.; Tai, Y.; Li, J.; Pang, D.; Zhang, Y.; Zhao, W.; Du, N.; Huang, Y. MiR-216a-5p/Hexokinase 2 axis regulates uveal melanoma growth through modulation of Warburg effect. Biochem. Biophys. Res. Commun. 2018, 501, 885–892. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, X.; Li, C.; Wang, W. miR-224-5p inhibits proliferation, migration, and invasion by targeting PIK3R3/AKT3 in uveal melanoma. J. Cell. Biochem. 2019, 120, 12412–12421. [Google Scholar] [CrossRef] [PubMed]
- Venza, I.; Visalli, M.; Beninati, C.; Benfatto, S.; Teti, D.; Venza, M. IL-10Ralpha expression is post-transcriptionally regulated by miR-15a, miR-185, and miR-211 in melanoma. BMC Med. Genom. 2015, 8, 81. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Jiang, J.; Wang, S.; Xia, X. Oncogenic role of microRNA20a in human uveal melanoma. Mol. Med. Rep. 2016, 14, 1560–1566. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.C.; Yang, X.; Wei, W.B.; Xu, X.L. Role of microRNA-21 in uveal melanoma cell invasion and metastasis by regulating p53 and its downstream protein. Int. J. Ophthalmol. 2018, 11, 1258–1268. [Google Scholar] [CrossRef]
- Venza, M.; Visalli, M.; Beninati, C.; Benfatto, S.; Teti, D.; Venza, I. miR-92a-3p and MYCBP2 are involved in MS-275-induced and c-myc-mediated TRAIL-sensitivity in melanoma cells. Int. Immunopharmacol. 2016, 40, 235–243. [Google Scholar] [CrossRef]
- Peng, J.; Liu, H.; Liu, C. MiR-155 Promotes Uveal Melanoma Cell Proliferation and Invasion by Regulating NDFIP1 Expression. Technol. Cancer Res. Treat. 2017, 16, 1160–1167. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; He, X.; Li, F.; Pan, H.; Huang, X.; Wen, X.; Zhang, H.; Li, B.; Ge, S.; Xu, X.; et al. The miR-181 family promotes cell cycle by targeting CTDSPL, a phosphatase-like tumor suppressor in uveal melanoma. J. Exp. Clin. Cancer Res. CR 2018, 37, 15. [Google Scholar] [CrossRef]
- Cheng, Y.; Cheng, T.; Zhao, Y.; Qu, Y. HMGA1 exacerbates tumor progression by activating miR-222 through PI3K/Akt/MMP-9 signaling pathway in uveal melanoma. Cell. Signal. 2019, 63, 109386. [Google Scholar] [CrossRef] [PubMed]
- Ling, J.W.; Lu, P.R.; Zhang, Y.B.; Jiang, S.; Zhang, Z.C. miR-367 promotes uveal melanoma cell proliferation and migration by regulating PTEN. Genet. Mol. Res. 2017, 16. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wang, Q.; Gao, X.; Shi, D.; Mi, S.; Han, Q. MicroRNA-454 functions as an oncogene by regulating PTEN in uveal melanoma. FEBS Lett. 2015, 589, 2791–2796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, Z.; Yang, C.; Yang, X.; Wu, S.; Feng, Z.; Qu, L.; Chen, X.; Liu, L.; Ma, Y. miR-652 Promotes Proliferation and Migration of Uveal Melanoma Cells by Targeting HOXA9. Med Sci. Monit. 2019, 25, 8722–8732. [Google Scholar] [CrossRef]
- Abdel-Rahman, M.H.; Boru, G.; Massengill, J.; Salem, M.M.; Davidorf, F.H. MET oncogene inhibition as a potential target of therapy for uveal melanomas. Investig. Ophthalmol. Vis. Sci. 2010, 51, 3333–3339. [Google Scholar] [CrossRef] [Green Version]
- Eedunuri, V.K.; Rajapakshe, K.; Fiskus, W.; Geng, C.; Chew, S.A.; Foley, C.; Shah, S.S.; Shou, J.; Mohamed, J.S.; Coarfa, C.; et al. miR-137 Targets p160 Steroid Receptor Coactivators SRC1, SRC2, and SRC3 and Inhibits Cell Proliferation. Mol. Endocrinol. 2015, 29, 1170–1183. [Google Scholar] [CrossRef] [Green Version]
- Mocellin, S.; Marincola, F.M.; Young, H.A. Interleukin-10 and the immune response against cancer: A counterpoint. J. Leukoc. Biol. 2005, 78, 1043–1051. [Google Scholar] [CrossRef]
- Wang, X.; Hu, Y.; Cui, J.; Zhou, Y.; Chen, L. Coordinated targeting of MMP-2/MMP-9 by miR-296-3p/FOXCUT exerts tumor-suppressing effects in choroidal malignant melanoma. Mol. Cell. Biochem. 2018, 445, 25–33. [Google Scholar] [CrossRef]
- Maldonado, M.D.M.; Dharmawardhane, S. Targeting Rac and Cdc42 GTPases in Cancer. Cancer Res. 2018, 78, 3101–3111. [Google Scholar] [CrossRef] [Green Version]
- Okayama, H.; Schetter, A.J.; Harris, C.C. MicroRNAs and inflammation in the pathogenesis and progression of colon cancer. Dig. Dis. 2012, 30 (Suppl. S2), 9–15. [Google Scholar] [CrossRef]
- Williams, M.D.; Zhang, X.; Belton, A.S.; Xian, L.; Huso, T.; Park, J.J.; Siems, W.F.; Gang, D.R.; Resar, L.M.; Reeves, R.; et al. HMGA1 drives metabolic reprogramming of intestinal epithelium during hyperproliferation, polyposis, and colorectal carcinogenesis. J. Proteome Res. 2015, 14, 1420–1431. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Xie, M.; He, H.; Shi, Y.; Luo, B.; Gong, G.; Li, J.; Wang, J.; Wu, X.; Wen, J. Increases urinary HMGA1 in serous epithelial ovarian cancer patients. Cancer Biomark. Sect. A Dis. Markers 2015, 15, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Rahman, M.H.; Yang, Y.; Zhou, X.P.; Craig, E.L.; Davidorf, F.H.; Eng, C. High frequency of submicroscopic hemizygous deletion is a major mechanism of loss of expression of PTEN in uveal melanoma. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2006, 24, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Guan, X.; Sun, Y.; Mi, J.; Shu, X.; Liu, F.; Li, C. miR-92a family and their target genes in tumorigenesis and metastasis. Exp. Cell Res. 2014, 323, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, S.; Gregory, R.I. MicroRNA biogenesis pathways in cancer. Nat. Rev. Cancer 2015, 15, 321–333. [Google Scholar] [CrossRef]
- Calin, G.A.; Croce, C.M. MicroRNA signatures in human cancers. Nat. Rev. Cancer 2006, 6, 857–866. [Google Scholar] [CrossRef] [PubMed]
- Gilad, S.; Meiri, E.; Yogev, Y.; Benjamin, S.; Lebanony, D.; Yerushalmi, N.; Benjamin, H.; Kushnir, M.; Cholakh, H.; Melamed, N.; et al. Serum microRNAs are promising novel biomarkers. PLoS ONE 2008, 3, e3148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosaka, N.; Iguchi, H.; Ochiya, T. Circulating microRNA in body fluid: A new potential biomarker for cancer diagnosis and prognosis. Cancer Sci. 2010, 101, 2087–2092. [Google Scholar] [CrossRef]
- Reiniger, I.W.; Schaller, U.C.; Haritoglou, C.; Hein, R.; Bosserhoff, A.K.; Kampik, A.; Mueller, A.J. “Melanoma inhibitory activity” (MIA): A promising serological tumour marker in metastatic uveal melanoma. Graefes Arch. Clin. Exp. Ophthalmol. 2005, 243, 1161–1166. [Google Scholar] [CrossRef]
- Bande, M.F.; Santiago, M.; Mera, P.; Piulats, J.M.; Blanco, M.J.; Rodriguez-Alvarez, M.X.; Capeans, C.; Pineiro, A.; Pardo, M. ME20-S as a Potential Biomarker for the Evaluation of Uveal Melanoma. Investig. Ophthalmol. Vis. Sci. 2015, 56, 7007–7011. [Google Scholar] [CrossRef] [Green Version]
- Worley, L.A.; Long, M.D.; Onken, M.D.; Harbour, J.W. Micro-RNAs associated with metastasis in uveal melanoma identified by multiplexed microarray profiling. Melanoma Res. 2008, 18, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Venkatesan, N.; Kanwar, J.; Deepa, P.R.; Khetan, V.; Crowley, T.M.; Raguraman, R.; Sugneswari, G.; Rishi, P.; Natarajan, V.; Biswas, J.; et al. Clinico-Pathological Association of Delineated miRNAs in Uveal Melanoma with Monosomy 3/Disomy 3 Chromosomal Aberrations. PLoS ONE 2016, 11, e0146128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, A.G.; Shih, J.; Yau, C.; Gibb, E.A.; Oba, J.; Mungall, K.L.; Hess, J.M.; Uzunangelov, V.; Walter, V.; Danilova, L.; et al. Integrative Analysis Identifies Four Molecular and Clinical Subsets in Uveal Melanoma. Cancer Cell 2018, 33, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smit, K.N.; Chang, J.; Derks, K.; Vaarwater, J.; Brands, T.; Verdijk, R.M.; Wiemer, E.A.C.; Mensink, H.W.; Pothof, J.; de Klein, A.; et al. Aberrant MicroRNA Expression and Its Implications for Uveal Melanoma Metastasis. Cancers 2019, 11, 815. [Google Scholar] [CrossRef] [Green Version]
- Wroblewska, J.P.; Lach, M.S.; Ustaszewski, A.; Kulcenty, K.; Ibbs, M.; Jagiello, I.; Suchorska, W.M.; Marszalek, A. The Potential Role of Selected miRNA in Uveal Melanoma Primary Tumors as Early Biomarkers of Disease Progression. Genes 2020, 11, 271. [Google Scholar] [CrossRef] [Green Version]
- Radhakrishnan, A.; Badhrinarayanan, N.; Biswas, J.; Krishnakumar, S. Analysis of chromosomal aberration (1, 3, and 8) and association of microRNAs in uveal melanoma. Mol. Vis. 2009, 15, 2146–2154. [Google Scholar]
- Falzone, L.; Romano, G.L.; Salemi, R.; Bucolo, C.; Tomasello, B.; Lupo, G.; Anfuso, C.D.; Spandidos, D.A.; Libra, M.; Candido, S. Prognostic significance of deregulated microRNAs in uveal melanomas. Mol. Med. Rep. 2019, 19, 2599–2610. [Google Scholar] [CrossRef] [Green Version]
- Xin, X.; Zhang, Y.; Ling, F.; Wang, L.; Sheng, X.; Qin, L.; Zhao, X. Identification of a nine-miRNA signature for the prognosis of Uveal Melanoma. Exp. Eye Res. 2019, 180, 242–249. [Google Scholar] [CrossRef]
- Larsen, A.C.; Holst, L.; Kaczkowski, B.; Andersen, M.T.; Manfe, V.; Siersma, V.D.; Kolko, M.; Kiilgaard, J.F.; Winther, O.; Prause, J.U.; et al. MicroRNA expression analysis and Multiplex ligation-dependent probe amplification in metastatic and non-metastatic uveal melanoma. Acta Ophthalmol. 2014, 92, 541–549. [Google Scholar] [CrossRef]
- Stark, M.S.; Gray, E.S.; Isaacs, T.; Chen, F.K.; Millward, M.; McEvoy, A.; Zaenker, P.; Ziman, M.; Soyer, H.P.; Glasson, W.J.; et al. A Panel of Circulating MicroRNAs Detects Uveal Melanoma with High Precision. Transl. Vis. Sci. Technol. 2019, 8, 12. [Google Scholar] [CrossRef]
- Russo, A.; Caltabiano, R.; Longo, A.; Avitabile, T.; Franco, L.M.; Bonfiglio, V.; Puzzo, L.; Reibaldi, M. Increased Levels of miRNA-146a in Serum and Histologic Samples of Patients with Uveal Melanoma. Front. Pharmacol. 2016, 7, 424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achberger, S.; Aldrich, W.; Tubbs, R.; Crabb, J.W.; Singh, A.D.; Triozzi, P.L. Circulating immune cell and microRNA in patients with uveal melanoma developing metastatic disease. Mol. Immunol. 2014, 58, 182–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ragusa, M.; Barbagallo, C.; Statello, L.; Caltabiano, R.; Russo, A.; Puzzo, L.; Avitabile, T.; Longo, A.; Toro, M.D.; Barbagallo, D.; et al. miRNA profiling in vitreous humor, vitreal exosomes and serum from uveal melanoma patients: Pathological and diagnostic implications. Cancer Biol. Ther. 2015, 16, 1387–1396. [Google Scholar] [CrossRef] [PubMed]
- Triozzi, P.L.; Achberger, S.; Aldrich, W.; Crabb, J.W.; Saunthararajah, Y.; Singh, A.D. Association of tumor and plasma microRNA expression with tumor monosomy-3 in patients with uveal melanoma. Clin. Epigenetics 2016, 8, 80. [Google Scholar] [CrossRef] [Green Version]
- Forloni, M.; Dogra, S.K.; Dong, Y.; Conte, D., Jr.; Ou, J.; Zhu, L.J.; Deng, A.; Mahalingam, M.; Green, M.R.; Wajapeyee, N. miR-146a promotes the initiation and progression of melanoma by activating Notch signaling. eLife 2014, 3, e01460. [Google Scholar] [CrossRef]
- Yajima, I.; Kumasaka, M.Y.; Thang, N.D.; Goto, Y.; Takeda, K.; Iida, M.; Ohgami, N.; Tamura, H.; Yamanoshita, O.; Kawamoto, Y.; et al. Molecular Network Associated with MITF in Skin Melanoma Development and Progression. J. Ski. Cancer 2011, 2011, 730170. [Google Scholar] [CrossRef] [Green Version]
- Trotta, R.; Chen, L.; Ciarlariello, D.; Josyula, S.; Mao, C.; Costinean, S.; Yu, L.; Butchar, J.P.; Tridandapani, S.; Croce, C.M.; et al. miR-155 regulates IFN-gamma production in natural killer cells. Blood 2012, 119, 3478–3485. [Google Scholar] [CrossRef] [Green Version]
- Cichocki, F.; Felices, M.; McCullar, V.; Presnell, S.R.; Al-Attar, A.; Lutz, C.T.; Miller, J.S. Cutting edge: MicroRNA-181 promotes human NK cell development by regulating Notch signaling. J. Immunol. 2011, 187, 6171–6175. [Google Scholar] [CrossRef] [Green Version]
- Djirackor, L.; Kalirai, H.; Coupland, S.E.; Petrovski, G. CD166high Uveal Melanoma Cells Represent a Subpopulation With Enhanced Migratory Capacity. Investig. Ophthalmol. Vis. Sci. 2019, 60, 2696–2704. [Google Scholar] [CrossRef] [Green Version]
- Joshi, P.; Kooshki, M.; Aldrich, W.; Varghai, D.; Zborowski, M.; Singh, A.D.; Triozzi, P.L. Expression of natural killer cell regulatory microRNA by uveal melanoma cancer stem cells. Clin. Exp. Metastasis 2016, 33, 829–838. [Google Scholar] [CrossRef]
- Chatila, W.M.; Criner, G.J.; Hancock, W.W.; Akimova, T.; Moldover, B.; Chang, J.K.; Cornwell, W.; Santerre, M.; Rogers, T.J. Blunted expression of miR-199a-5p in regulatory T cells of patients with chronic obstructive pulmonary disease compared to unaffected smokers. Clin. Exp. Immunol. 2014, 177, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Zhang, M.; Jiang, X.; Zhang, Z.; Dai, L.; Min, S.; Wu, X.; He, Q.; Liu, J.; Zhang, Y.; et al. miR-223 suppresses differentiation of tumor-induced CD11b(+) Gr1(+) myeloid-derived suppressor cells from bone marrow cells. Int. J. Cancer 2011, 129, 2662–2673. [Google Scholar] [CrossRef]
- Chu, C.; Qu, K.; Zhong, F.L.; Artandi, S.E.; Chang, H.Y. Genomic maps of long noncoding RNA occupancy reveal principles of RNA-chromatin interactions. Mol. Cell 2011, 44, 667–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prensner, J.R.; Chinnaiyan, A.M. The emergence of lncRNAs in cancer biology. Cancer Discov. 2011, 1, 391–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batista, P.J.; Chang, H.Y. Long noncoding RNAs: Cellular address codes in development and disease. Cell 2013, 152, 1298–1307. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.T. Epigenetic regulation by long noncoding RNAs. Science 2012, 338, 1435–1439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitz, S.U.; Grote, P.; Herrmann, B.G. Mechanisms of long noncoding RNA function in development and disease. Cell. Mol. Life Sci. 2016, 73, 2491–2509. [Google Scholar] [CrossRef] [Green Version]
- Moran, V.A.; Perera, R.J.; Khalil, A.M. Emerging functional and mechanistic paradigms of mammalian long non-coding RNAs. Nucleic Acids Res. 2012, 40, 6391–6400. [Google Scholar] [CrossRef]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA hypothesis: The Rosetta Stone of a hidden RNA language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.M.; Shi, S.S.; Jian, T.M.; Tang, D.R.; Wu, T.; Sun, F.Y. LncRNA PVT1 knockdown affects proliferation and apoptosis of uveal melanoma cells by inhibiting EZH2. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 2880–2887. [Google Scholar] [CrossRef]
- Villanueva, M.T. Anticancer drugs: All roads lead to EZH2 inhibition. Nat. Rev. Drug Discov. 2017, 16, 239. [Google Scholar] [CrossRef] [PubMed]
- Su, M.; Xiao, Y.; Tang, J.; Wu, J.; Ma, J.; Tian, B.; Zhou, Y.; Wang, H.; Yang, D.; Liao, Q.J.; et al. Role of lncRNA and EZH2 Interaction/Regulatory Network in Lung Cancer. J. Cancer 2018, 9, 4156–4165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, T.; Wang, Y.; Zhou, F.; Gao, G.; Ren, S.; Zhou, C. Prognostic value of high EZH2 expression in patients with different types of cancer: A systematic review with meta-analysis. Oncotarget 2016, 7, 4584–4597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Q.; Zhao, N.; Zha, G.; Wang, H.; Tong, Q.; Xin, S. LncRNA HOXA11-AS Exerts Oncogenic Functions by Repressing p21 and miR-124 in Uveal Melanoma. DNA Cell Biol. 2017, 36, 837–844. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Yuan, Y.; Ma, R.; Xu, B.; Zhang, R. lncRNA SNHG7 affects malignant tumor behaviors through downregulation of EZH2 in uveal melanoma cell lines. Oncol. Lett. 2020, 19, 1505–1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Gong, J.; Liu, H. High expression of lncRNA PVT1 independently predicts poor overall survival in patients with primary uveal melanoma. PLoS ONE 2017, 12, e0189675. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.; Tang, H.; Zhao, X.; Sun, Y.; Jiang, Y.; Liu, Y. Long non-coding RNA FTH1P3 facilitates uveal melanoma cell growth and invasion through miR-224-5p. PLoS ONE 2017, 12, e0184746. [Google Scholar] [CrossRef] [Green Version]
- Guan, Y.; Kuo, W.L.; Stilwell, J.L.; Takano, H.; Lapuk, A.V.; Fridlyand, J.; Mao, J.H.; Yu, M.; Miller, M.A.; Santos, J.L.; et al. Amplification of PVT1 contributes to the pathophysiology of ovarian and breast cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2007, 13, 5745–5755. [Google Scholar] [CrossRef] [Green Version]
- Iden, M.; Fye, S.; Li, K.; Chowdhury, T.; Ramchandran, R.; Rader, J.S. The lncRNA PVT1 Contributes to the Cervical Cancer Phenotype and Associates with Poor Patient Prognosis. PLoS ONE 2016, 11, e0156274. [Google Scholar] [CrossRef] [Green Version]
- Xing, Y.; Wen, X.; Ding, X.; Fan, J.; Chai, P.; Jia, R.; Ge, S.; Qian, G.; Zhang, H.; Fan, X. CANT1 lncRNA Triggers Efficient Therapeutic Efficacy by Correcting Aberrant lncing Cascade in Malignant Uveal Melanoma. Mol. Ther. J. Am. Soc. Gene Ther. 2017, 25, 1209–1221. [Google Scholar] [CrossRef]
- Li, F.; Li, X.; Qiao, L.; Liu, W.; Xu, C.; Wang, X. MALAT1 regulates miR-34a expression in melanoma cells. Cell Death Dis. 2019, 10, 389. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Cheng, H.; Wang, G.; Yu, G.; Zhang, D.; Wang, Y.; Fan, W.; Yang, W. Deregulation of miR-183 promotes melanoma development via lncRNA MALAT1 regulation and ITGB1 signal activation. Oncotarget 2017, 8, 3509–3518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Sun, P.; Zhou, Q.Y.; Gao, X.; Han, Q. Long noncoding RNA MALAT1 promotes uveal melanoma cell growth and invasion by silencing of miR-140. Am. J. Transl. Res. 2016, 8, 3939–3946. [Google Scholar] [PubMed]
- Wu, S.; Chen, H.; Zuo, L.; Jiang, H.; Yan, H. Suppression of long noncoding RNA MALAT1 inhibits the development of uveal melanoma via microRNA-608-mediated inhibition of HOXC4. Am. J. Physiol. Cell Physiol. 2020, 318, C903–C912. [Google Scholar] [CrossRef] [PubMed]
- Di Sanzo, M.; Aversa, I.; Santamaria, G.; Gagliardi, M.; Panebianco, M.; Biamonte, F.; Zolea, F.; Faniello, M.C.; Cuda, G.; Costanzo, F. FTH1P3, a Novel H-Ferritin Pseudogene Transcriptionally Active, Is Ubiquitously Expressed and Regulated during Cell Differentiation. PLoS ONE 2016, 11, e0151359. [Google Scholar] [CrossRef]
- Pan, H.; Ni, H.; Zhang, L.; Xing, Y.; Fan, J.; Li, P.; Li, T.; Jia, R.; Ge, S.; Zhang, H.; et al. P2RX7-V3 is a novel oncogene that promotes tumorigenesis in uveal melanoma. Tumour Biol. J. Int. Soc. Oncodevelopmental Biol. Med. 2016, 37, 13533–13543. [Google Scholar] [CrossRef]
- Lu, L.; Yu, X.; Zhang, L.; Ding, X.; Pan, H.; Wen, X.; Xu, S.; Xing, Y.; Fan, J.; Ge, S.; et al. The Long Non-Coding RNA RHPN1-AS1 Promotes Uveal Melanoma Progression. Int. J. Mol. Sci. 2017, 18, 226. [Google Scholar] [CrossRef] [Green Version]
- Ding, X.; Wang, X.; Lin, M.; Xing, Y.; Ge, S.; Jia, R.; Zhang, H.; Fan, X.; Li, J. PAUPAR lncRNA suppresses tumourigenesis by H3K4 demethylation in uveal melanoma. FEBS Lett. 2016, 590, 1729–1738. [Google Scholar] [CrossRef]
- Moriyama, M.; Osawa, M.; Mak, S.S.; Ohtsuka, T.; Yamamoto, N.; Han, H.; Delmas, V.; Kageyama, R.; Beermann, F.; Larue, L.; et al. Notch signaling via Hes1 transcription factor maintains survival of melanoblasts and melanocyte stem cells. J. Cell Biol. 2006, 173, 333–339. [Google Scholar] [CrossRef]
- Asnaghi, L.; Ebrahimi, K.B.; Schreck, K.C.; Bar, E.E.; Coonfield, M.L.; Bell, W.R.; Handa, J.; Merbs, S.L.; Harbour, J.W.; Eberhart, C.G. Notch signaling promotes growth and invasion in uveal melanoma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2012, 18, 654–665. [Google Scholar] [CrossRef] [Green Version]
- Cheng, G.; He, J.; Zhang, L.; Ge, S.; Zhang, H.; Fan, X. HIC1 modulates uveal melanoma progression by activating lncRNA-numb. Tumour Biol. J. Int. Soc. Oncodevelopmental Biol. Med. 2016, 37, 12779–12789. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Li, X.F.; Wu, Q.; Ma, R.Q.; Qian, J.; Zhang, R. LncRNA SNHG15 predicts poor prognosis in uveal melanoma and its potential pathways. Int. J. Ophthalmol. 2020, 13, 1195–1201. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhang, L.; Fan, J.; Jia, R.; Song, X.; Xu, X.; Dai, L.; Zhuang, A.; Ge, S.; Fan, X. Let-7b overexpression leads to increased radiosensitivity of uveal melanoma cells. Melanoma Res. 2015, 25, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Rossi, E.; Pagliara, M.M.; Orteschi, D.; Dosa, T.; Sammarco, M.G.; Caputo, C.G.; Petrone, G.; Rindi, G.; Zollino, M.; Blasi, M.A.; et al. Pembrolizumab as first-line treatment for metastatic uveal melanoma. Cancer Immunol. Immunother. 2019, 68, 1179–1185. [Google Scholar] [CrossRef] [Green Version]
- Audrito, V.; Serra, S.; Stingi, A.; Orso, F.; Gaudino, F.; Bologna, C.; Neri, F.; Garaffo, G.; Nassini, R.; Baroni, G.; et al. PD-L1 up-regulation in melanoma increases disease aggressiveness and is mediated through miR-17-5p. Oncotarget 2017, 8, 15894–15911. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Johnston, N.; Zheng, X.; Wang, H.; Zhang, X.; Gao, D.; Min, W. miR-28 modulates exhaustive differentiation of T cells through silencing programmed cell death-1 and regulating cytokine secretion. Oncotarget 2016, 7, 53735–53750. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Biswas, A.; Liu, H.; Sen, S.; Paruchuri, A.; Katsonis, P.; Lichtarge, O.; Chand Dakal, T.; Maulik, U.; Gromiha, M.M.; et al. Mutational Landscape of the BAP1 Locus Reveals an Intrinsic Control to Regulate the miRNA Network and the Binding of Protein Complexes in Uveal Melanoma. Cancers 2019, 11, 1600. [Google Scholar] [CrossRef] [Green Version]
- Bande Rodriguez, M.F.; Fernandez Marta, B.; Lago Baameiro, N.; Santiago-Varela, M.; Silva-Rodriguez, P.; Blanco-Teijeiro, M.J.; Pardo Perez, M.; Pineiro Ces, A. Blood Biomarkers of Uveal Melanoma: Current Perspectives. Clin. Ophthalmol. 2020, 14, 157–169. [Google Scholar] [CrossRef] [Green Version]
- Telonis, A.G.; Rigoutsos, I. Race Disparities in the Contribution of miRNA Isoforms and tRNA-Derived Fragments to Triple-Negative Breast Cancer. Cancer Res. 2018, 78, 1140–1154. [Google Scholar] [CrossRef] [Green Version]
- Telonis, A.G.; Loher, P.; Jing, Y.; Londin, E.; Rigoutsos, I. Beyond the one-locus-one-miRNA paradigm: MicroRNA isoforms enable deeper insights into breast cancer heterogeneity. Nucleic Acids Res. 2015, 43, 9158–9175. [Google Scholar] [CrossRef] [Green Version]
- Londin, E.; Magee, R.; Shields, C.L.; Lally, S.E.; Sato, T.; Rigoutsos, I. IsomiRs and tRNA-derived fragments are associated with metastasis and patient survival in uveal melanoma. Pigment Cell Melanoma Res. 2020, 33, 52–62. [Google Scholar] [CrossRef] [PubMed]



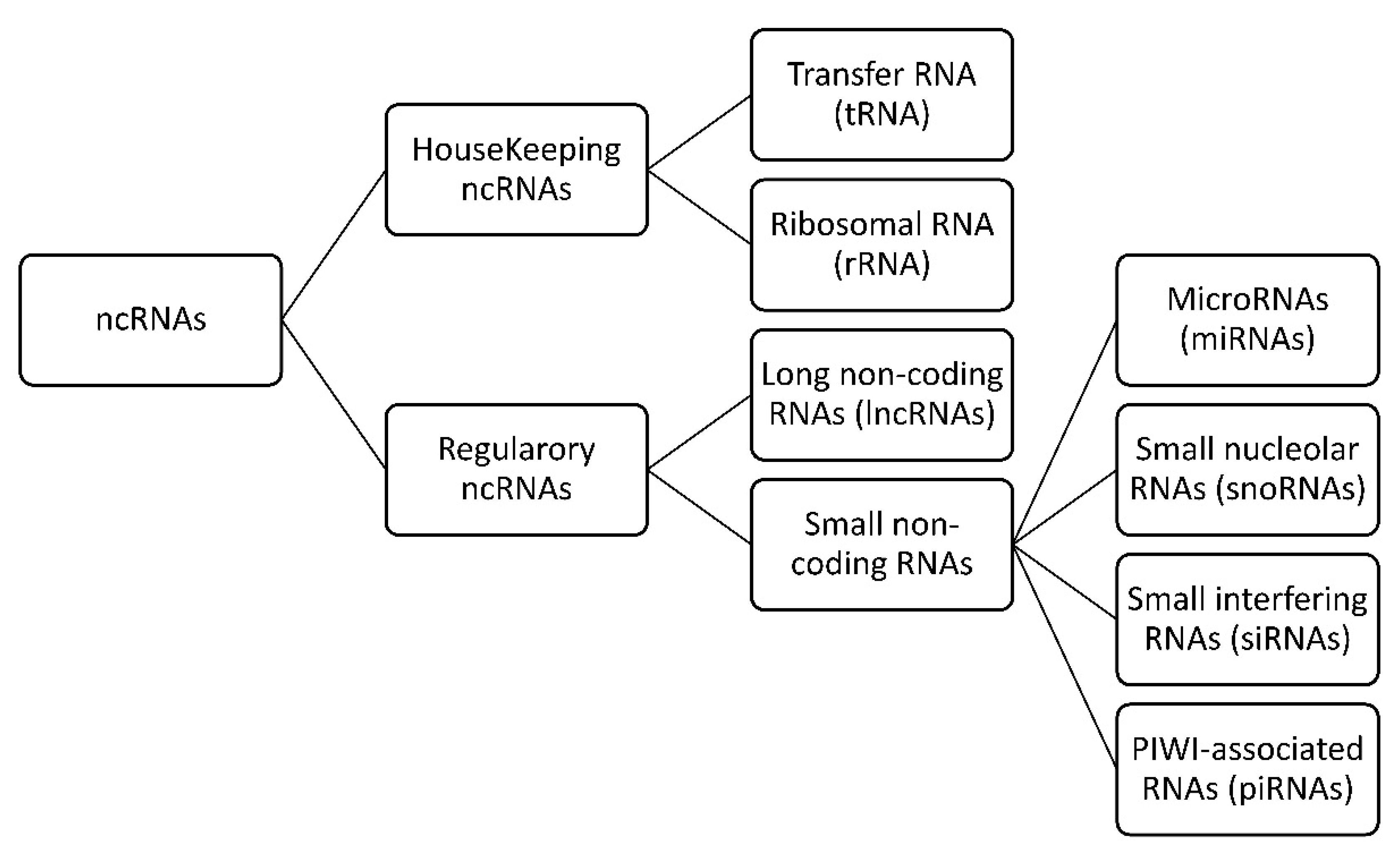{kind=link}
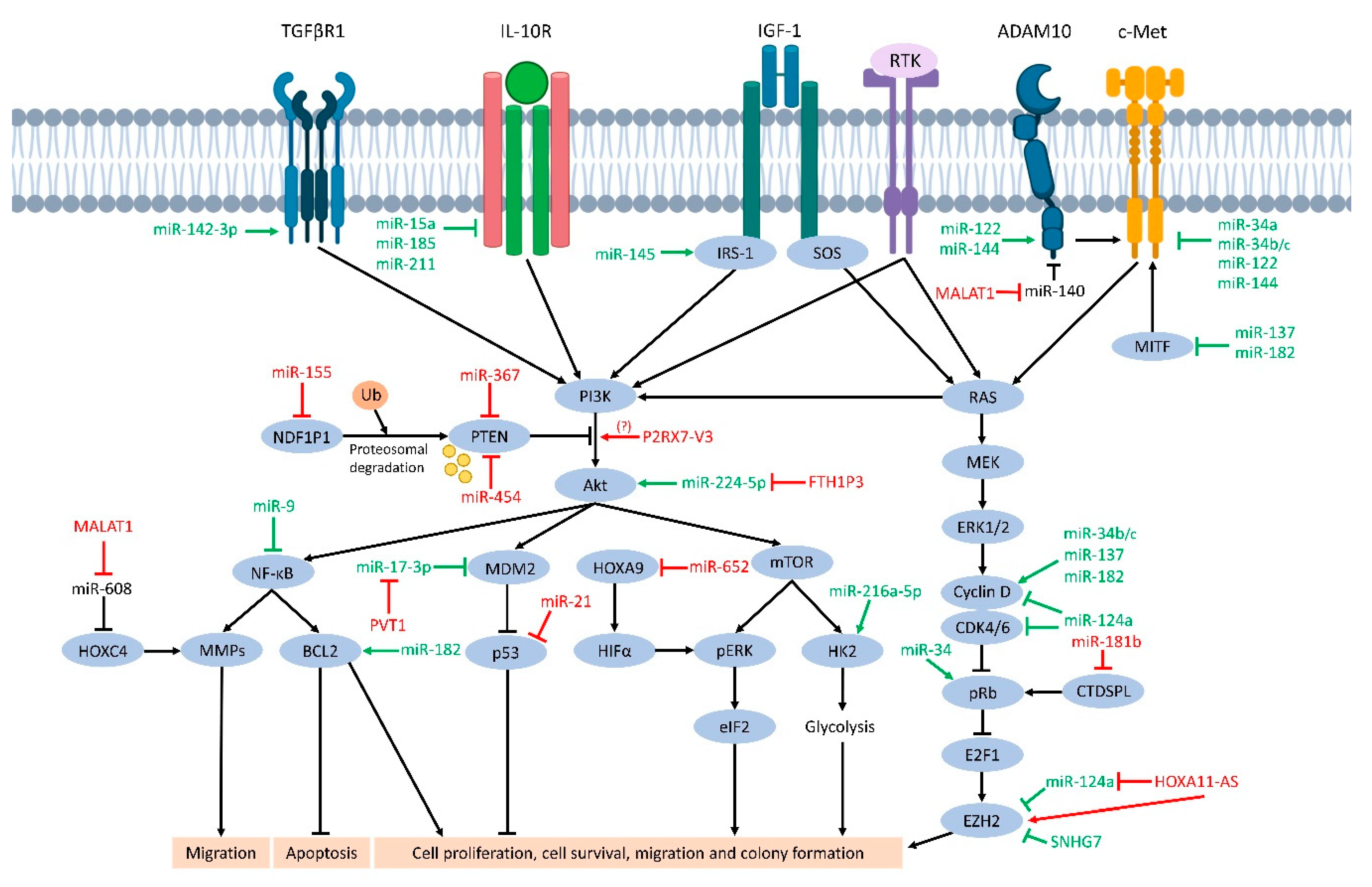{kind=link}
| miRNA | Location † | Sample Type | Inhibitory Events | Validation | Target | Reference |
|---|---|---|---|---|---|---|
| miR-9 | 1q22 | Highly invasive cell lines: MUM2B and C918 | Cell migration and invasion | MA, IA, PA, RT-PCR, WB, FRA | NF-kB1 | [20] |
| miR-17-3p | 13q31.3 | UM patient tumor tissues; UM cell lines: OCM-1A, MUM-2C, C918, and MUM-2B | Cell proliferation and metastasis | RT-PCR, PA, MA, IA, Dual LUCA, WB, ATM | MDM2 | [21] |
| miR-23a | 19p13.12 | UM cell lines: OCM-1; UM patient tumor tissues | Cell migration | RT-PCR, MA IF, WB | Zeb1 | [22] |
| miR-34a | 1p36.22 | UM cell lines: M17, M21, M23, and SP6.5; UM patient tumor tissues | Cell proliferation and migration | PA, MA, IA, Dual LUCA, IF, WB | C-MET LGR4 | [23,24] |
| miR-34b/c | 11q23.1 | UM cell lines: SP6.5; UM patient tumor tissues (upregulated in doxorubicin- and epigenetic drug-treated SP6.5 cells) | Cell proliferation and migration | PA, AA, MA, LUCA, WB | C-MET | [25] |
| miR-122 | 18q21.31 | UM cell lines: 92.1, MEL270, OMM2.5, UPMM2, and UPMM3; TCGA UM dataset, UM patient tumor tissues | Cell proliferation and migration | RT-PCR, LUCA, WB | C-MET, ADAM10 | [26] |
| miR-124a | 3 loci: 8p23.1 8q12.3 20q13.33 | UM cell lines: M17, M21, M23, and SP6.5; UM patient tumor tissues | Cell proliferation, migration, and invasion | PA, MA, IA, RT-PCR, ATM, WB | CDK4, CDK6, cyclin D2, EZH2 | [27] |
| miR-137 | 1p21.3 | UM cell lines: OMM1.3, Mel202, 92.1, OMM1, OCM1, and OCM3 | Cell proliferation | RT-PCR, PA, LUCA, WB | MITF CDK6 | [28] |
| miR-142-3p | 17q22 | UM cell lines: M17 and SP6.5; UM patient tumor tissues | Cell proliferation, migration, and invasion | RT-PCR, PA, MA, IA, AA, LUCA, WB | CDC25C, TGFβR1, GNAQ, WASL, RAC1 | [29] |
| miR-144 | 17q11.2 | UM cell lines: MUM-2B, C918, MUM-2C, and OCM-1A; UM patient tumor tissues | Cell proliferation and migration | LUCA, WB, PA, IA | C-MET | [30] |
| miR-145 | 5q32 | UM cell lines: MUM-2B and OCM-1; UM patient tumor tissues | Cell proliferation and promotes apoptosis | RT-PCR, PA, IA, Dual LUCA, WB | IRS-1 NPR1 | [31] |
| miR-182 | 7q32.2 | UM cell lines: M23 and SP6.5; UM patient tumor tissues;(upregulated by p53 activation) | Cell proliferation, migration, and invasion | RT-PCR, PA, MA, LUCA, ATM, WB | MITF, BCL2, cyclin D2 | [32] |
| miR-205 | 1q32.2 | UM patient tumor tissues (both high- and low invasive UM) | Cell proliferation and promotes apoptosis | PA, IA, Dual LUCA, WB | NPR1 | [31] |
| miR-216a-5p | 2p16.1 | UM cell line: MUM-2B; UM patient tumor tissues | Cell proliferation | PA, RT-PCR, LUCA, GA, WB, ATM, IHC | HK2 | [33] |
| miR-224-5p | Xq28 | UM cell line: OCM-1A; UM patient tumor tissues | Cell proliferation, migration, and invasion | RT-PCR, PA, MA, IA, LUCA, WB | PIK3R3/AKT3, FTH1P3, RAC1, Fizzled 5 | [34] |
| miR-15a, miR-185, miR-211 | 13q14.2 22q11.21 15q13.3 | Cell line: OCM-1; UM patient tumor tissues | Cell proliferation | RT-PCR, PA, WB | IL-10Rα | [35] |
| miRNA | Location † | Sample Type | Pro-Tumorigenic Events | Validation | Target | Reference |
|---|---|---|---|---|---|---|
| miR-20a | 13q31.3 | MUM-2B and MUM-2C metastatic UM cells; UM patient tumor tissues | Cell proliferation, migration, and invasion | PA, IA, MA, RT-PCR | Not validated | [36] |
| miR-21 | 17q23.1 | OCM-1, M619 and MUM-2B UM cells | Cell proliferation, migration, and invasion | PA, AA, MA, Dual LUCA, WB | p53 | [37] |
| miR-92a-3p | 2 loci: 13q31.3 Xq26.2 | OCM-1 UM cells | Inhibition of apoptosis | LUCA, WB | MYCBP2 | [38] |
| miR-155 | 21q21.3 | OCM-1A, MUM-2C, C918, and MUM-2B UM cells; UM patient tumor tissues | Cell proliferation and invasion | PA, IA, LUCA, WB | NDFIP1 | [39] |
| miR-181 (family) | Chr 1, 9, and 19 | SP6.5, VUP, OCM1, MUM2b, and 92-1; UM patient tumor tissues | Cell proliferation | Dual LUCA, WB, RT-PCR | CTDSPL | [40] |
| miR-222 | Xp11.3 | C918 and MUM-2B UM cells | Cell proliferation and migration | AA, PA, MA, WB | PI3K/Akt/MMP-9 | [41] |
| miR-367 | 4q25 | UM cell lines: M17, M23, MUM-2B, and C918; UM patient tumor tissues | Cell proliferation and migration | LUCA, PA, MA, WB | PTEN | [42] |
| miR-454 | 17q22 | UM cell lines: OCM-1A, MUM-2B, MUM-2C, and C918; UM patients tumor tissues | Cell proliferation, colony formation, and invasion | RT-PCR, PA, IA, LUCA, WB | PTEN | [43] |
| miR-652 | Xq23 | MUM-2B and MEL270 UM cell lines; UM patient tumor tissues | Cell proliferation and migration | PA, MA, Dual LUCA, RT-PCR, WB | HOXA9 | [44] |
| Sample Type | Over Expressed | Under Expressed | Reference | |
|---|---|---|---|---|
| High Risk UM Metastatic | Low Risk UM Metastatic | High Risk UM Metastatic | ||
| 24 UM patient frozen tumor tissues | let-7b, miR-143, miR-193b, miR-199a, miR-199 *, miR-652 | [61] | ||
| 60 UM patient tumor tissues and 11 control tissues | miR-33a, miR-99a, miR-135b, miR-196a, miR-325, miR-497, miR-512-5p, miR-549, miR-556-5p, miR-585, miR-640, miR-885-5p † | let-7e, miR-1, miR-10a, miR-18a, miR-19b-1, miR- 26a-2, miR-34c-5p, miR-129, miR-133a, miR-154, miR-181a, miR-218, miR-369-3p, miR-377, miR-376c, miR-493, miR-495, miR-586 †† | [66] | |
| 26 UM patient tumor tissues | miR-16-5p, miR-17-5p, miR-21-5p, miR-132-5p, miR-151a-3p ‡ | let-7c-5p, miR-99a-3p, miR-99a-5p, miR-101-3p, miR-181a-2-3p, miR-181b-5p, miR-378d ‡, miR-1537-3p | [64] | |
| 50 UM patient tumor tissues | miR-346, miR-592, miR-1247 | miR-506 and miR-513c | [65] | |
| 86 UM patient tumor tissues | miR-134, miR-143, miR-146b, miR-149, miR-199a, miR-214 | [62] | ||
| TCGA UM database | Let-7b, miR-21, miR-29b, miR-142, miR-143, miR-144, miR-145, miR-150, miR-155, miR-199a, miR-199b, miR-330, miR-486 ‡ | miR-30c, miR-181a-5p, miR-335, miR-140, Xq27.3 group | [63] | |
| TCGA UM database | miR-195, miR-224, miR-365a, miR-365b, miR-452, miR-4709, miR-7702 | miR-513c, miR-873 | [68] | |
| TCGA UM database | let-7b(5p/3p), miR-199a-5p, miR-224-5p, miR-452-5p, miR-592 | miR-211-5p, miR-508-3p, miR-509–3-5p, miR-513a-5p, miR-513c-5p, miR-514a-3p | [67] | |
| Sample Type | miRNA | Levels | Differentiate (Ratio) | |
|---|---|---|---|---|
| Vitreous humor and serum (± exosomes) | Vitreous humor (+ exosomes): miR-21, miR-34a, miR-146a Serum (+ exosomes): miR-146a | ↑ | Primary/Control | [73] |
| Serum (14 patients with UM vs. 14 controls) | miR-146a | ↑ | Primary/Control | [71] |
| miR-19a, miR-30d ‡, miR-127, miR-451, miR-518f, miR-1274b | ↓ | |||
| Plasma (33 UM M3/22 UM D3/26 controls) | miR-92b, miR-199a-5p, miR-223 | ↑ | M3/D3 M3/Control | [74] |
| Plasma (6 patients with UM vs. 26 controls) | miR-20a, miR-125b, miR-146a, miR-155, miR-181a, miR-223 | ↑ | Primary/Control | [72] |
| miR-20a, miR-125b, miR-146a, miR-155, miR-223 | ↑ | Metastatic/Primary | ||
| miR-181a | ↓ | |||
| Blood (10 uveal nevus, 50 localized UM, 5 metastatic UM) | miR-16, miR-145, miR-146a, miR-204, miR-211, miR-363-3p | ↑ | Primary/Metastatic Primary/Nevus | [70] |
| miR-211 | ↑ | Metastatic/Primary | ||
| miR-204 | ↓ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bande, M.; Fernandez-Diaz, D.; Fernandez-Marta, B.; Rodriguez-Vidal, C.; Lago-Baameiro, N.; Silva-Rodríguez, P.; Paniagua, L.; Blanco-Teijeiro, M.J.; Pardo, M.; Piñeiro, A. The Role of Non-Coding RNAs in Uveal Melanoma. Cancers 2020, 12, 2944. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12102944
Bande M, Fernandez-Diaz D, Fernandez-Marta B, Rodriguez-Vidal C, Lago-Baameiro N, Silva-Rodríguez P, Paniagua L, Blanco-Teijeiro MJ, Pardo M, Piñeiro A. The Role of Non-Coding RNAs in Uveal Melanoma. Cancers. 2020; 12(10):2944. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12102944
Chicago/Turabian StyleBande, Manuel, Daniel Fernandez-Diaz, Beatriz Fernandez-Marta, Cristina Rodriguez-Vidal, Nerea Lago-Baameiro, Paula Silva-Rodríguez, Laura Paniagua, María José Blanco-Teijeiro, María Pardo, and Antonio Piñeiro. 2020. "The Role of Non-Coding RNAs in Uveal Melanoma" Cancers 12, no. 10: 2944. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12102944