Enhanced Expression of IGFBP-3 Reduces Radiosensitivity and Is Associated with Poor Prognosis in Oral Squamous Cell Carcinoma

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Clinical Significance of IGFBP-3 Expression in Patients with OSCC
2.2. Relationships between IGFBP-3 Expression and Survival
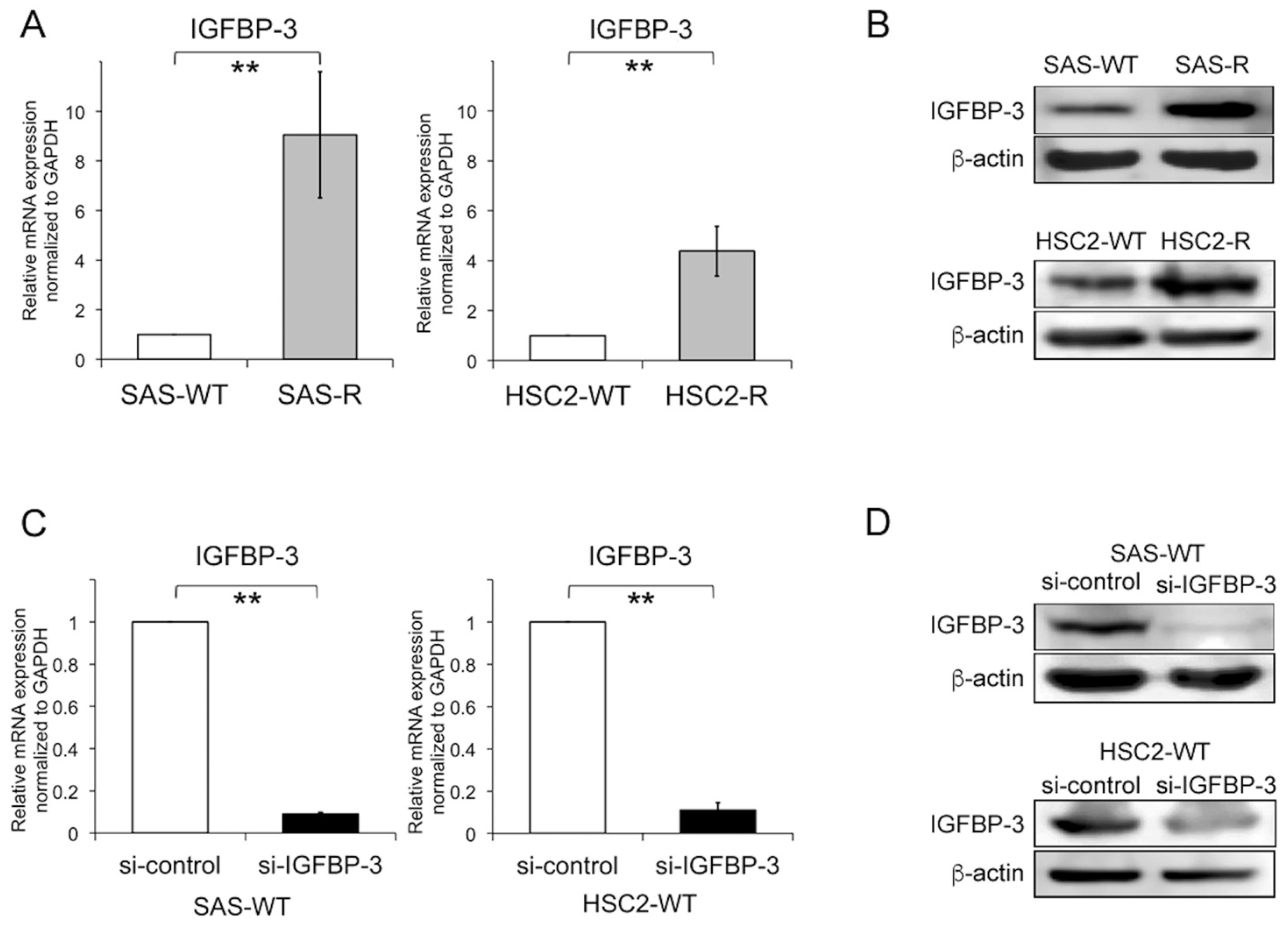
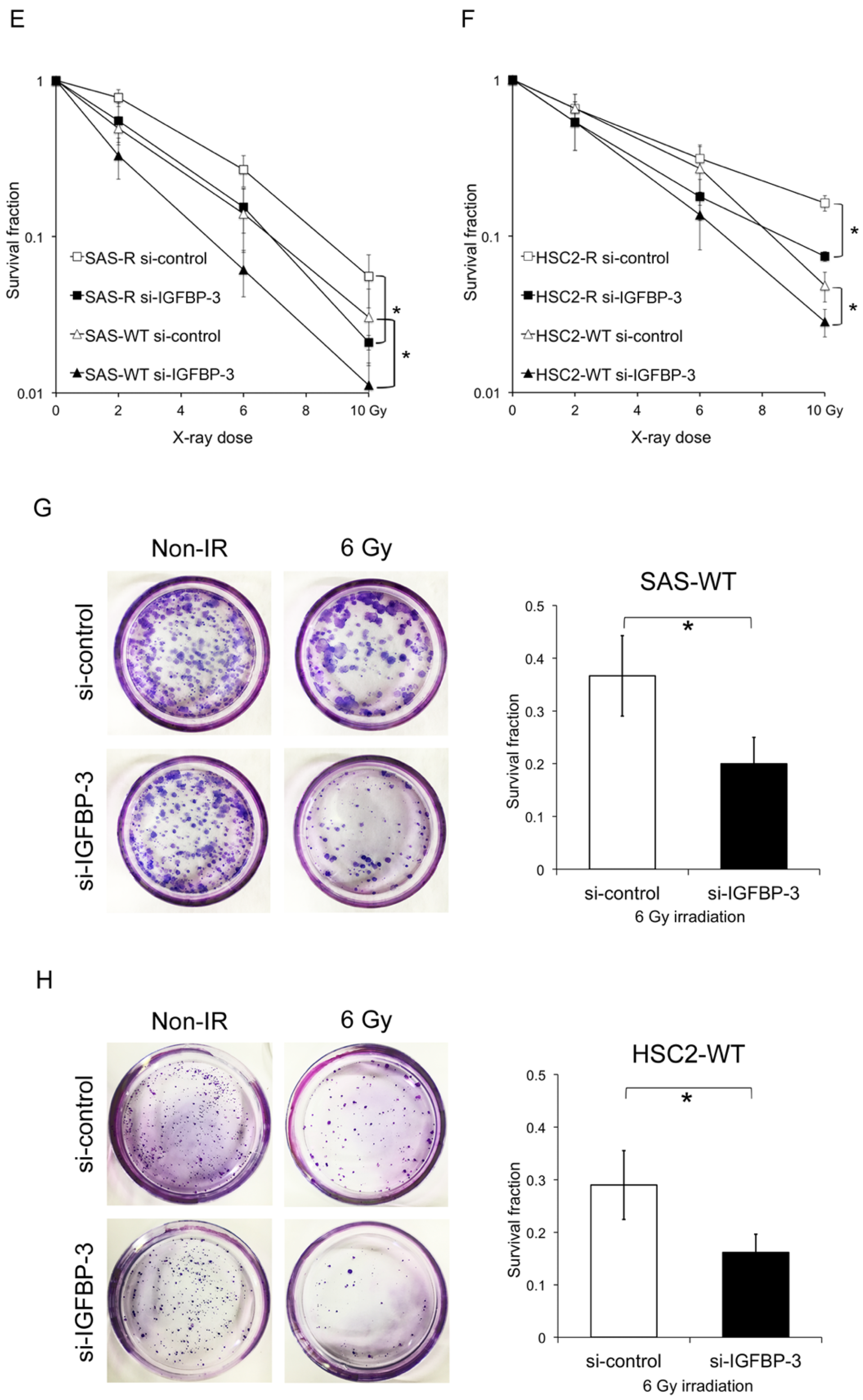
2.3. Effect of IGFBP-3 on Radiosensitivity in OSCC Cells
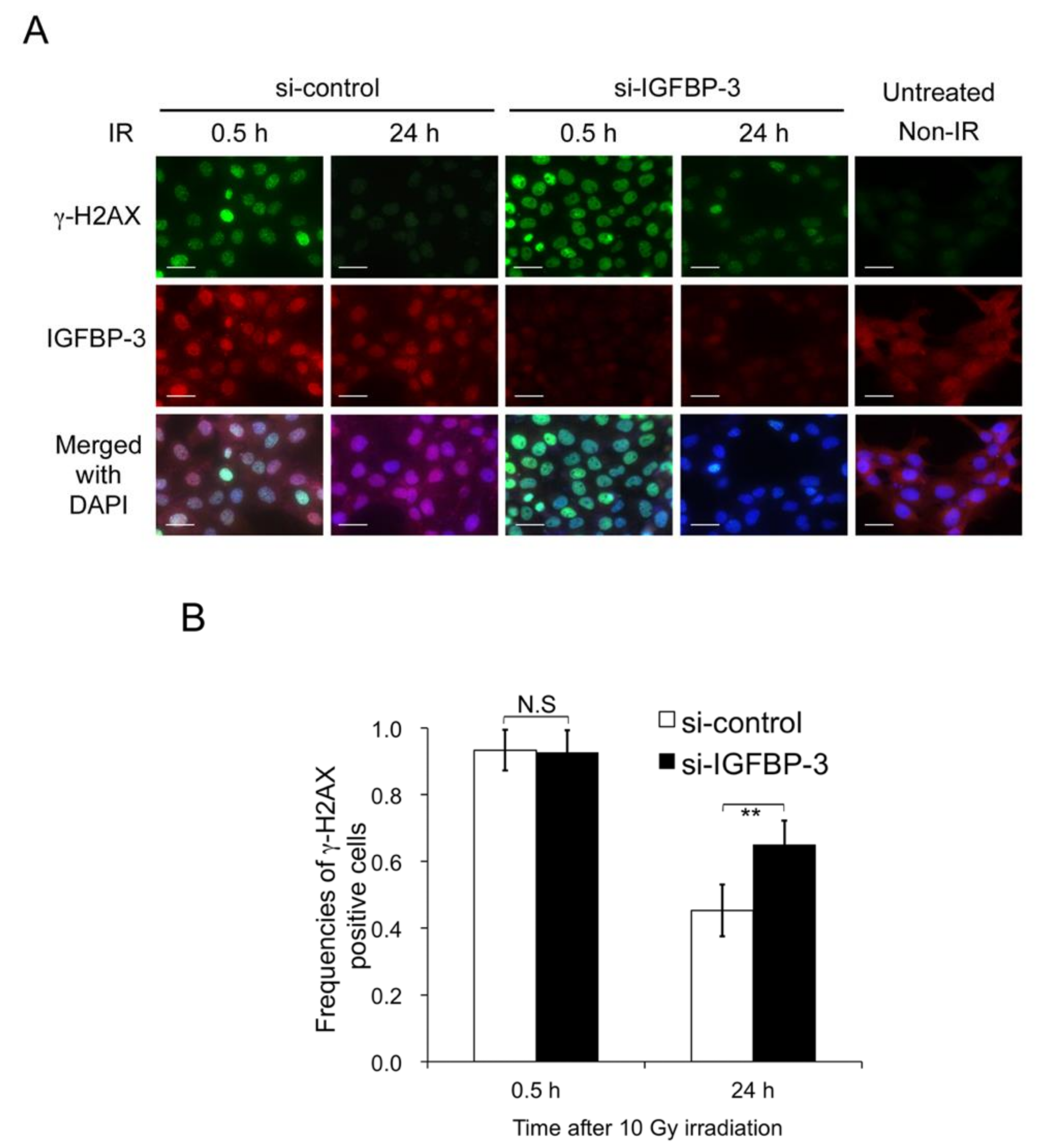
2.4. Contribution of IGFBP-3 to Decreases in DNA Damage in Irradiated OSCC Cells
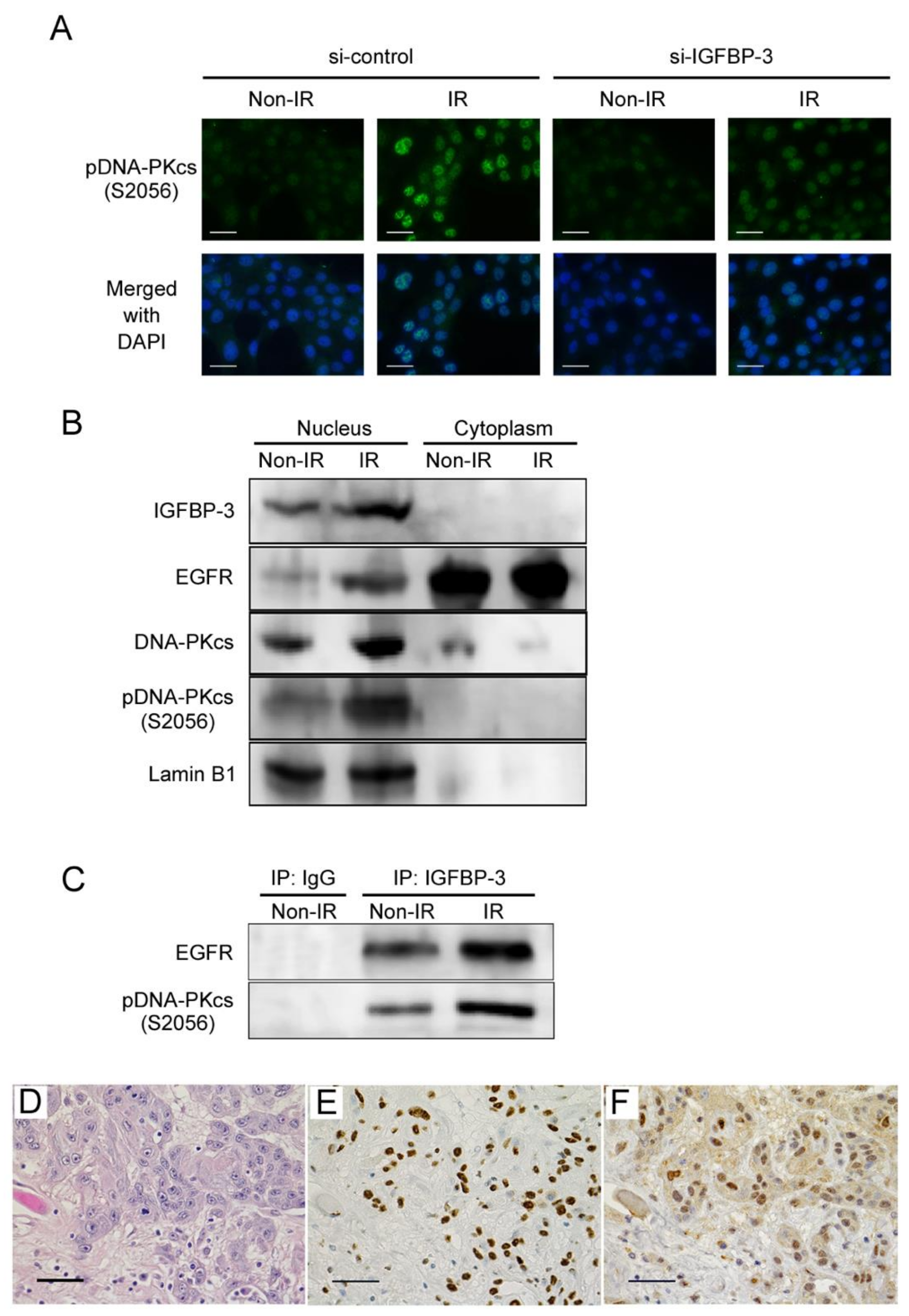
2.5. Functional Role of IGFBP-3 in the DNA Repair of Irradiated OSCC Cells
2.6. Interactions among IGFBP-3, EGFR, and Phosphorylated DNA-PKcs in Irradiated OSCC Cells
3. Discussion
4. Materials and Methods
4.1. Clinical Samples
4.2. Immunohistochemical Staining and Evaluation
4.3. Cell lines and Culture
4.4. Irradiation
4.5. siRNA Transfection
4.6. RNA Isolation and qRT-PCR
4.7. HDS Assays
4.8. Clonogenic Assays
4.9. Immunofluorescent Staining and Evaluation
4.10. Western Blot Analysis
4.11. Immunoprecipitation
4.12. Cell Proliferation Assays
4.13. Enzyme-Linked Immunosorbent Assay (ELISA) for Plasma IGFBP-3
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2016. CA Cancer J. Clin. 2016, 66, 7–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.; Kong, W.; Peng, Y.; Miao, Q.; Mackillop, W.J. Temporal trends in the incidence and survival of cancers of the upper aerodigestive tract in Ontario and the United States. Int. J. Cancer 2009, 125, 2159–2165. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Ye, W.; Shao, Q.; Zhang, M.; Liang, J. Nrf2 is a potential therapeutic target in radioresistance in human cancer. Crit. Rev. Oncol. Hematol. 2013, 88, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, V.N.; Thylur, D.S.; Bauschard, M.; Schmale, I.; Sinha, U.K. Overcoming radioresistance in head and neck squamous cell carcinoma. Oral Oncol. 2016, 63, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Daza, D.O.; Sundstrom, G.; Bergqvist, C.A.; Duan, C.; Larhammar, D. Evolution of the insulin-like growth factor binding protein (IGFBP) family. Endocrinology 2011, 152, 2278–2289. [Google Scholar] [CrossRef] [Green Version]
- Baxter, R.C. Nuclear actions of insulin-like growth factor binding protein-3. Gene 2015, 569, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Baxter, R.C. IGF binding proteins in cancer: Mechanistic and clinical insights. Nat. Rev. Cancer 2014, 14, 329–341. [Google Scholar] [CrossRef]
- Rohrmann, S.; Grote, V.A.; Becker, S.; Rinaldi, S.; Tjønneland, A.; Roswall, N.; Grønbæk, H.; Overvad, K.; Boutron-Ruault, M.C.; Clavel-Chapelon, F.; et al. Concentrations of IGF-I and IGFBP-3 and pancreatic cancer risk in the European Prospective Investigation into Cancer and Nutrition. Br. J. Cancer 2012, 106, 1004–1010. [Google Scholar] [CrossRef] [Green Version]
- Wolpin, B.M.; Meyerhardt, J.A.; Chan, A.T.; Ng, K.; Chan, J.A.; Wu, K.; Pollak, M.N.; Giovannucci, E.L.; Fuchs, C.S. Insulin, the insulin-like growth factor axis, and mortality in patients with nonmetastatic colorectal cancer. J. Clin. Oncol. 2009, 27, 176–185. [Google Scholar] [CrossRef]
- Rinaldi, S.; Cleveland, R.; Norat, T.; Biessy, C.; Rohrmann, S.; Linseisen, J.; Boeing, H.; Pischon, T.; Panico, S.; Agnoli, C.; et al. Serum levels of IGF-I, IGFBP-3 and colorectal cancer risk: Results from the EPIC cohort, plus a meta-analysis of prospective studies. Int. J. Cancer 2010, 126, 1702–1715. [Google Scholar] [CrossRef]
- Peeters, P.H.; Lukanova, A.; Allen, N.; Berrino, F.; Key, T.; Dossus, L.; Rinaldi, S.; van Gils, C.H.; Bueno-de-Mesquita, H.B.; Boeing, H.; et al. Serum IGF-I, its major binding protein (IGFBP-3) and epithelial ovarian cancer risk: The European Prospective Investigation into Cancer and Nutrition (EPIC). Endocr. Relat. Cancer 2007, 14, 81–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vatten, L.J.; Holly, J.M.; Gunnell, D.; Tretli, S. Nested case-control study of the association of circulating levels of serum insulin-like growth factor I and insulin-like growth factor binding protein 3 with breast cancer in young women in Norway. Cancer Epidemiol. Biomark. Prev. 2008, 17, 2097–2100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.; Liu, S.; Xu, W.; Wang, X.; Zhao, W.; Wu, J. IGF-I and IGFBP-3 and the risk of lung cancer: A meta-analysis based on nested case-control studies. J. Exp. Clin. Cancer Res. 2009, 28, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansson, M.; McKay, J.D.; Rinaldi, S.; Wiklund, F.; Adami, H.O.; Grönberg, H.; Kaaks, R.; Stattin, P. Genetic and plasma variation of insulin-like growth factor binding proteins in relation to prostate cancer incidence and survival. Prostate 2009, 69, 1281–1291. [Google Scholar] [CrossRef] [PubMed]
- Roddam, A.W.; Allen, N.E.; Appleby, P.; Key, T.J.; Ferrucci, L.; Carter, H.B.; Metter, E.J.; Chen, C.; Weiss, N.S.; Fitzpatrick, A.; et al. Insulin-like growth factors, their binding proteins, and prostate cancer risk: Analysis of individual patient data from 12 prospective studies. Ann. Int. Med. 2008, 149, 461–471. [Google Scholar] [PubMed] [Green Version]
- Sheen-Chen, S.M.; Zhang, H.; Huang, C.C.; Tang, R.P. Insulin-like growth factor-binding protein-3 in breast cancer: Analysis with tissue microarray. Anticancer. Res. 2009, 29, 1131–1135. [Google Scholar]
- Santosh, V.; Arivazhagan, A.; Sreekanthreddy, P.; Srinivasan, H.; Thota, B.; Srividya, M.R.; Vrinda, M.; Sridevi, S.; Shailaja, B.C.; Samuel, C.; et al. Grade-specific expression of insulin-like growth factor-binding proteins-2, -3, and -5 in astrocytomas: IGFBP-3 emerges as a strong predictor of survival in patients with newly diagnosed glioblastoma. Cancer Epidemiol. Biomark. Prev. 2010, 19, 1399–1408. [Google Scholar] [CrossRef] [Green Version]
- Kuwahara, Y.; Roudkenar, M.H.; Urushihara, Y.; Saito, Y.; Tomita, K.; Roushandeh, A.M.; Sato, T.; Kurimasa, A.; Fukumoto, M. Clinically relevant radioresistant cell line: A simple model to understand cancer radioresistance. Med. Mol. Morphol. 2017, 50, 195–204. [Google Scholar] [CrossRef]
- Jackson, S.P.; Bartek, J. The DNA-damage response in human biology and disease. Nature 2009, 461, 1071–1078. [Google Scholar] [CrossRef] [Green Version]
- Kuwahara, Y.; Li, L.; Baba, T.; Nakagawa, H.; Shimura, T.; Yamamoto, Y.; Ohkubo, Y.; Fukumoto, M. Clinically relevant radioresistant cells efficiently repair DNA double-strand breaks induced by X-rays. Cancer Sci. 2009, 100, 747–752. [Google Scholar] [CrossRef]
- Matsuoka, Y.; Nakayama, H.; Yoshida, R.; Hirosue, A.; Nagata, M.; Tanaka, T.; Kawahara, K.; Sakata, J.; Arita, H.; Nakashima, H.; et al. IL-6 controls resistance to radiation by suppressing oxidative stress via the Nrf2-antioxidant pathway in oral squamous cell carcinoma. Br. J. Cancer 2016, 115, 1234–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Gent, D.C.; Hoeijmakers, J.H.; Kanaar, R. Chromosomal stability and the DNA double-stranded break connection. Nat. Rev. Genet. 2001, 2, 196–206. [Google Scholar] [CrossRef]
- Ferguson, D.O.; Alt, F.W. DNA double strand break repair and chromosomal translocation: Lessons from animal models. Oncogene 2001, 20, 5572–5579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, J.S.; Um, J.H.; Kang, C.D.; Bae, J.H.; Kim, D.U.; Lee, Y.J.; Kim, D.W.; Chung, B.S.; Kim, S.H. Fractionated irradiation leads to restoration of drug sensitivity in MDR cells that correlates with down-regulation of P-gp and DNA-dependent protein kinase activity. Radiat. Res. 2004, 162, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Wanner, G.; Mayer, C.; Kehlbach, R.; Rodemann, H.P.; Dittmann, K. Activation of protein kinase Cepsilon stimulates DNA-repair via epidermal growth factor receptor nuclear accumulation. Radiother. Oncol. 2008, 86, 383–390. [Google Scholar] [CrossRef]
- Luo, L.L.; Zhao, L.; Wang, Y.X.; Tian, X.P.; Xi, M.; Shen, J.X.; He, L.R.; Li, Q.Q.; Liu, S.L.; Zhang, P.; et al. Insulin-like growth factor binding protein-3 is a new predictor of radiosensitivity on esophageal squamous cell carcinoma. Sci. Rep. 2015, 5, 17336. [Google Scholar] [CrossRef] [Green Version]
- Yoshino, K.; Motoyama, S.; Koyota, S.; Shibuya, K.; Usami, S.; Maruyama, K.; Saito, H.; Minamiya, Y.; Sugiyama, T.; Ogawa, J.I. IGFBP3 and BAG1 enhance radiation-induced apoptosis in squamous esophageal cancer cells. Biochem. Biophys. Res. Commun. 2011, 404, 1070–1075. [Google Scholar] [CrossRef]
- Sun, Y.; Zheng, S.; Torossian, A.; Speirs, C.K.; Schleicher, S.; Giacalone, N.J.; Carbone, D.P.; Zhao, Z.; Lu, B. Role of insulin-like growth factor-1 signaling pathway in cisplatin-resistant lung cancer cells. Int. J. Radiat. Oncol. Biol. Phys. 2012, 82, e563–e572. [Google Scholar] [CrossRef] [Green Version]
- Zhong, L.P.; Yang, X.; Zhang, L.; Wei, K.J.; Pan, H.Y.; Zhou, X.J.; Li, J.; Chen, W.T.; Zhang, Z.Y. Overexpression of insulin-like growth factor binding protein 3 in oral squamous cell carcinoma. Oncol. Rep. 2008, 20, 1441–1447. [Google Scholar] [CrossRef] [Green Version]
- Yen, Y.C.; Hsiao, J.R.; Jiang, S.S.; Chang, J.S.; Wang, S.H.; Shen, Y.Y.; Chen, C.H.; Chang, I.S.; Chang, J.Y.; Chen, Y.W. Insulin-like growth factor-independent insulin-like growth factor binding protein 3 promotes cell migration and lymph node metastasis of oral squamous cell carcinoma cells by requirement of integrin β1. Oncotarget 2015, 6, 41837–41855. [Google Scholar] [CrossRef] [Green Version]
- Major, J.M.; Pollak, M.N.; Snyder, K.; Virtamo, J.; Albanes, D. Insulin-like growth factors and risk of kidney cancer in men. Br. J. Cancer 2010, 103, 132–135. [Google Scholar] [CrossRef] [PubMed]
- Wraight, C.J.; Liepe, I.J.; White, P.J.; Hibbs, A.R.; Werther, G.A. Intranuclear localization of insulin-like growth factor binding protein-3 (IGFBP-3) during cell division in human keratinocytes. J. Investig. Dermatol. 1998, 111, 239–242. [Google Scholar] [CrossRef] [Green Version]
- Seligson, D.B.; Yu, H.; Tze, S.; Said, J.; Pantuck, A.J.; Cohen, P.; Lee, K.W. IGFBP-3 nuclear localization predicts human prostate cancer recurrence. Horm. Cancer 2013, 4, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Schedlich, L.J.; Young, T.F.; Firth, S.M.; Baxter, R.C. Insulin-like growth factor-binding protein (IGFBP)-3 and IGFBP-5 share a common nuclear transport pathway in T47D human breast carcinoma cells. J. Biol. Chem. 1998, 273, 18347–18352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liccardi, G.; Hartley, J.A.; Hochhauser, D. EGFR nuclear translocation modulates DNA repair following cisplatin and ionizing radiation treatment. Cancer Res. 2011, 71, 1103–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, M.Z.; Marzec, K.A.; Martin, J.L.; Baxter, R.C. The role of insulin-like growth factor binding protein-3 in the breast cancer cell response to DNA-damaging agents. Oncogene 2014, 33, 85–96. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.; Lee, S.; Seo, D.; Kim, D.; Kim, K.; Kim, E.; Kang, J.; Seong, K.M.; Youn, H.; Youn, B. Cellular stress responses in radiotherapy. Cells 2019, 8, 1105. [Google Scholar] [CrossRef] [Green Version]
- Nagata, M.; Nakayama, H.; Tanaka, T.; Yoshida, R.; Yoshitake, Y.; Fukuma, D.; Kawahara, K.; Nakagawa, Y.; Ota, K.; Hiraki, A.; et al. Overexpression of cIAP2 contributes to 5-FU resistance and a poor prognosis in oral squamous cell carcinoma. Br. J. Cancer 2011, 105, 1322–1330. [Google Scholar] [CrossRef]
- Yoshida, R.; Nakayama, H.; Nagata, M.; Hirosue, A.; Tanaka, T.; Kawahara, K.; Nakagawa, Y.; Matsuoka, Y.; Sakata, J.; Arita, H.; et al. Overexpression of nucleostemin contributes to an advanced malignant phenotype and a poor prognosis in oral squamous cell carcinoma. Br. J. Cancer 2014, 111, 2308–2315. [Google Scholar] [CrossRef] [Green Version]
- Kuwahara, Y.; Mori, M.; Oikawa, T.; Shimura, T.; Ohtake, Y.; Mori, S.; Ohkubo, Y.; Fukumoto, M. The modified high-density survival assay is the useful tool to predict the effectiveness of fractionated radiation exposure. J. Radiat. Res. 2010, 51, 297–302. [Google Scholar] [CrossRef] [Green Version]
- Taneja, N.; Davis, M.; Choy, J.S.; Beckett, M.A.; Singh, R.; Kron, S.J.; Weichselbaum, R.R. Histone H2AX phosphorylation as a predictor of radiosensitivity and target for radiotherapy. J. Biol. Chem. 2004, 279, 2273–2280. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Total | IGFBP-3 Status | p-Value | |
|---|---|---|---|---|
| High Expression | Low Expression | |||
| n (%) | n (%) | |||
| 52 | 27 (51.9) | 25 (48.1) | ||
| Age, years | ||||
| Median | 66.5 | 67.3 | 65.6 | |
| Range | 30–85 | 30–85 | 47–84 | |
| ≤65 | 23 | 9 (39.1) | 14 (60.9) | 0.100 |
| >65 | 29 | 18 (62.1) | 11 (37.9) | |
| Sex | ||||
| Male | 31 | 13 (41.9) | 18 (58.1) | 0.080 |
| Female | 21 | 14 (66.7) | 7 (33.3) | |
| Primary site | ||||
| Tongue | 16 | 9 (56.3) | 7 (43.7) | 0.570 |
| Mandible gingiva | 15 | 8 (53.3) | 7 (46.7) | |
| Maxilla gingiva | 7 | 5 (71.4) | 2 (28.6) | |
| Buccal mucosa | 10 | 4 (40.0) | 6 (60.0) | |
| Oral floor | 4 | 1 (25.0) | 3 (75.0) | |
| Clinical stage | ||||
| III | 18 | 10 (55.6) | 8 (44.4) | |
| IV | 34 | 17 (50.0) | 17 (50.0) | |
| pT-stage | ||||
| T2 | 10 | 4 (40.4) | 6 (60.0) | 0.545 |
| T3 | 24 | 12 (50.0) | 12 (50.0) | |
| T4 | 18 | 11 (61.1) | 7 (38.9) | |
| pN-stage | ||||
| N0 | 31 | 18 (58.1) | 13 (41.9) | 0.282 |
| ≥N1 | 21 | 9 (42.9) | 12 (57.1) | |
| Differentiation | ||||
| Well | 40 | 21 (55.3) | 19 (44.7) | 0.879 |
| Moderate, poor | 12 | 6 (50.0) | 6 (50.0) | |
| Pathological response | ||||
| Grade 0, I, II | 29 | 19 (65.5) | 10 (34.5) | 0.028 * |
| Grade III, IV | 23 | 8 (34.8) | 15 (65.2) | |
| Characteristic | Assigned Score | OS | DFS | ||
|---|---|---|---|---|---|
| Hazard Ratio (95% CI) | p-Value | Hazard Ratio (95% CI) | p-Value | ||
| IGFBP-3 expression | |||||
| Low | 0 | 3.783 (1.148–17.11) | 0.028 * | 3.878 (1.294–14.35) | 0.014 * |
| High | 1 | ||||
| pN-stage | |||||
| N0 | 0 | 2.951 (1.033–8.598) | 0.043 * | 3.107 (1.136–8.641) | 0.028 * |
| ≥N1 | 1 | ||||
| Pathological response | |||||
| Grade 0, I, II | 0 | 0.087 (0.005–0.446) | 0.001 ** | 0.135 (0.021–0.495) | 0.001 ** |
| Grade III, IV | 1 | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakata, J.; Hirosue, A.; Yoshida, R.; Matsuoka, Y.; Kawahara, K.; Arita, H.; Nakashima, H.; Yamamoto, T.; Nagata, M.; Kawaguchi, S.; et al. Enhanced Expression of IGFBP-3 Reduces Radiosensitivity and Is Associated with Poor Prognosis in Oral Squamous Cell Carcinoma. Cancers 2020, 12, 494. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12020494
Sakata J, Hirosue A, Yoshida R, Matsuoka Y, Kawahara K, Arita H, Nakashima H, Yamamoto T, Nagata M, Kawaguchi S, et al. Enhanced Expression of IGFBP-3 Reduces Radiosensitivity and Is Associated with Poor Prognosis in Oral Squamous Cell Carcinoma. Cancers. 2020; 12(2):494. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12020494
Chicago/Turabian StyleSakata, Junki, Akiyuki Hirosue, Ryoji Yoshida, Yuichiro Matsuoka, Kenta Kawahara, Hidetaka Arita, Hikaru Nakashima, Tatsuro Yamamoto, Masashi Nagata, Sho Kawaguchi, and et al. 2020. "Enhanced Expression of IGFBP-3 Reduces Radiosensitivity and Is Associated with Poor Prognosis in Oral Squamous Cell Carcinoma" Cancers 12, no. 2: 494. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12020494