USP47 Promotes Tumorigenesis by Negative Regulation of p53 through Deubiquitinating Ribosomal Protein S2

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
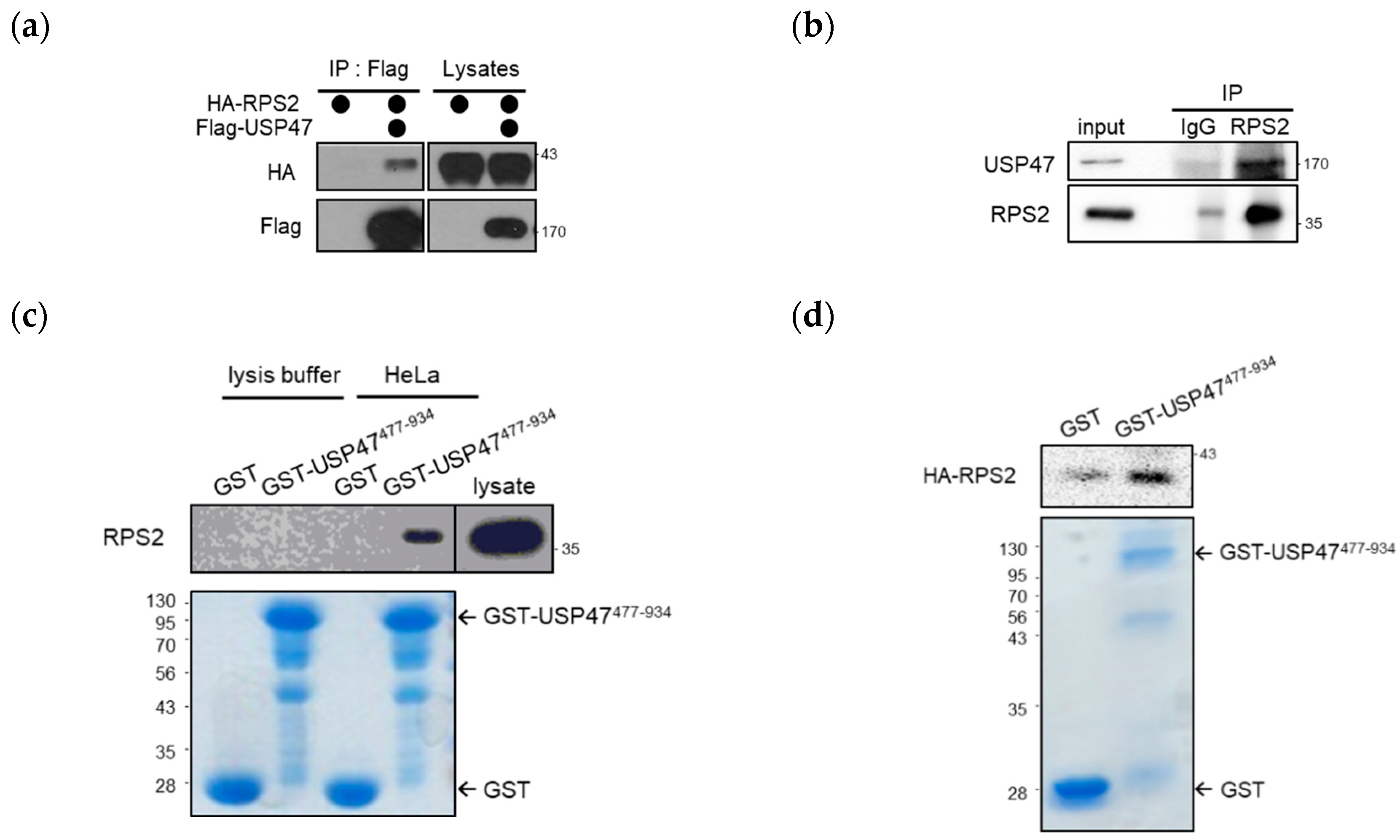
2.1. USP47 Interacts with RPS2
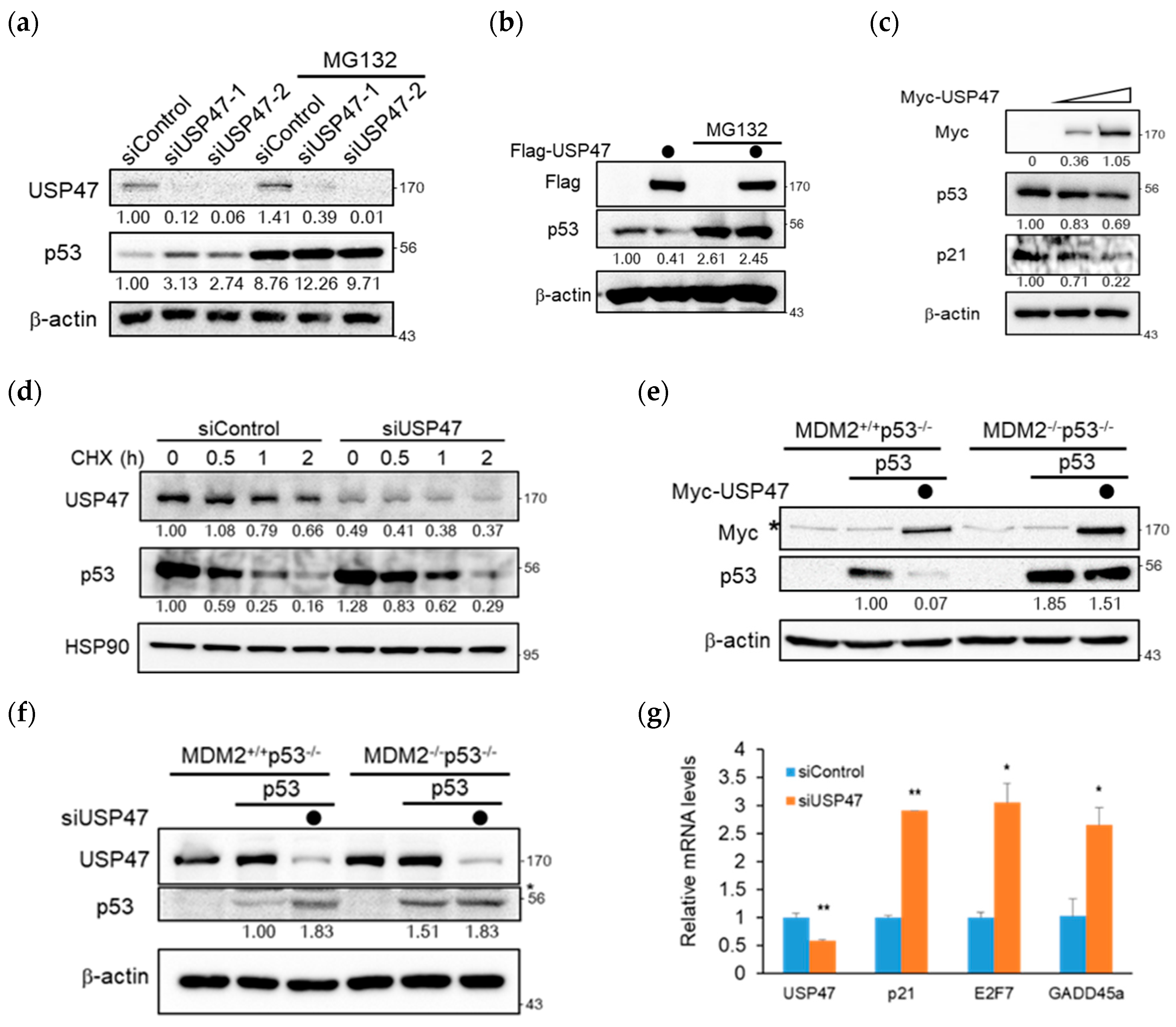
2.2. USP47 Regulates p53 in an MDM2-Dependent Manner
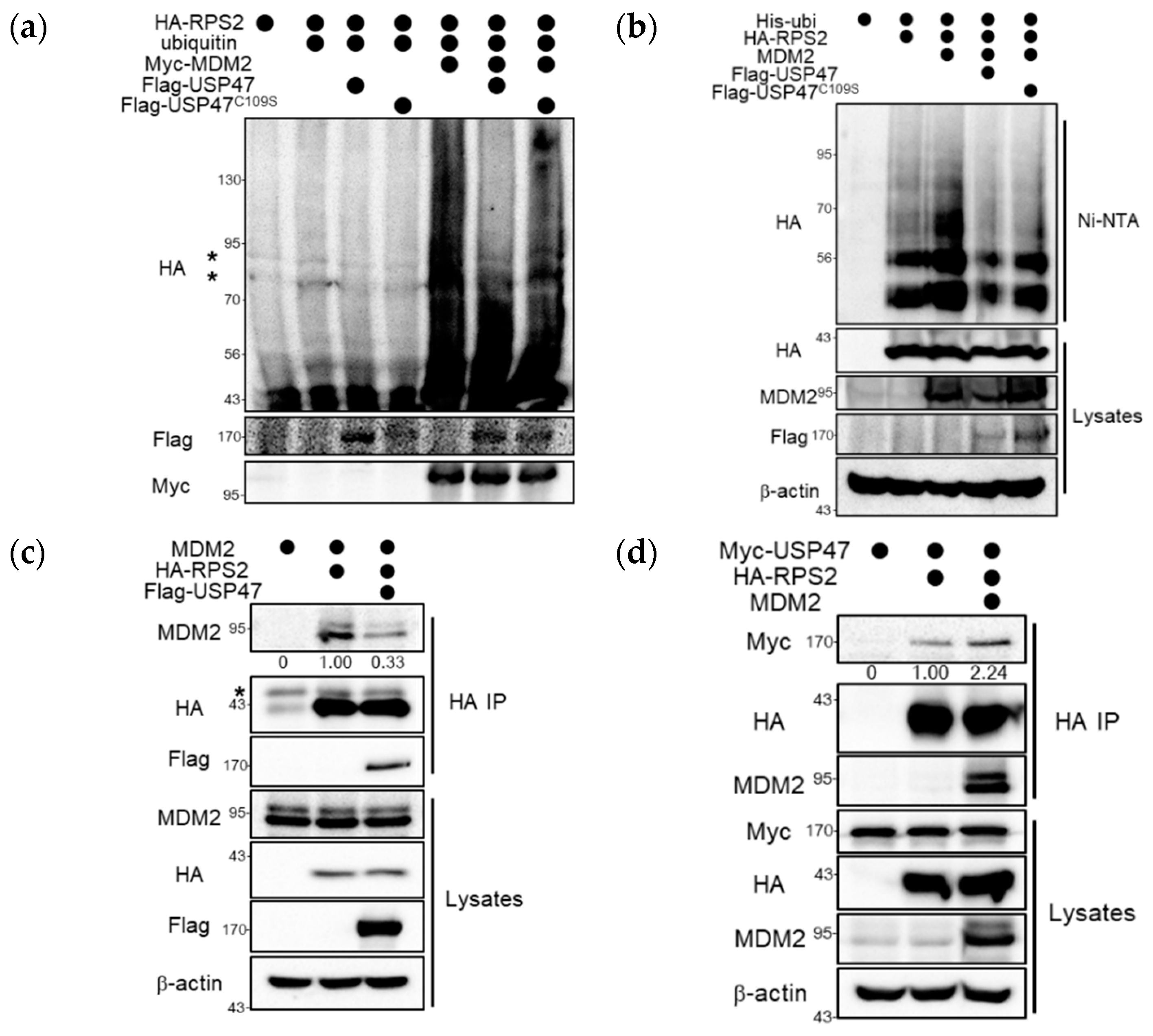
2.3. USP47 Deubiquitinates RPS2 and Regulates the Interaction between MDM2 and RPS2
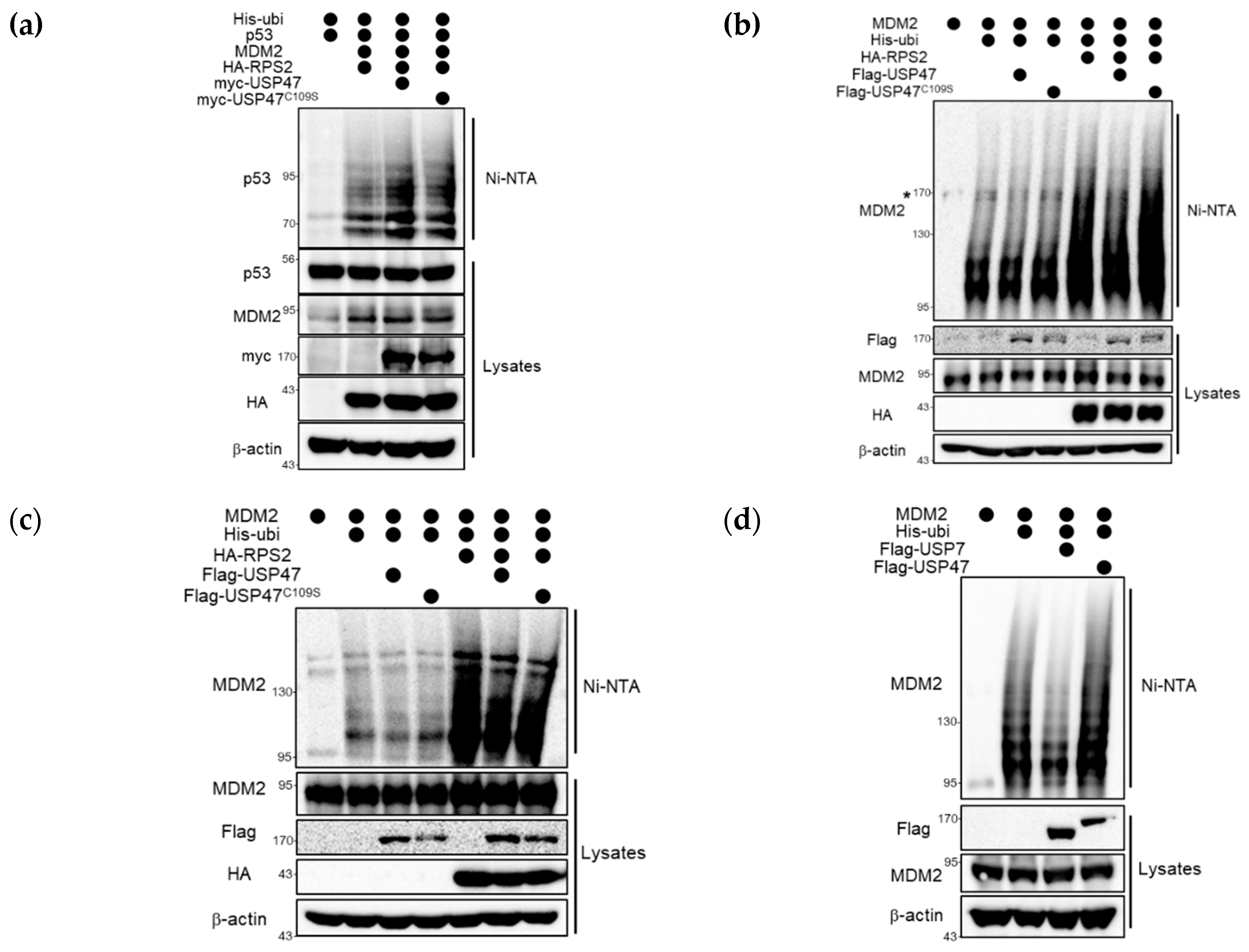
2.4. USP47 Alleviates RPS2-mediated Inhibition of MDM2 by Regulation of Ubiquitination Status of RPS2
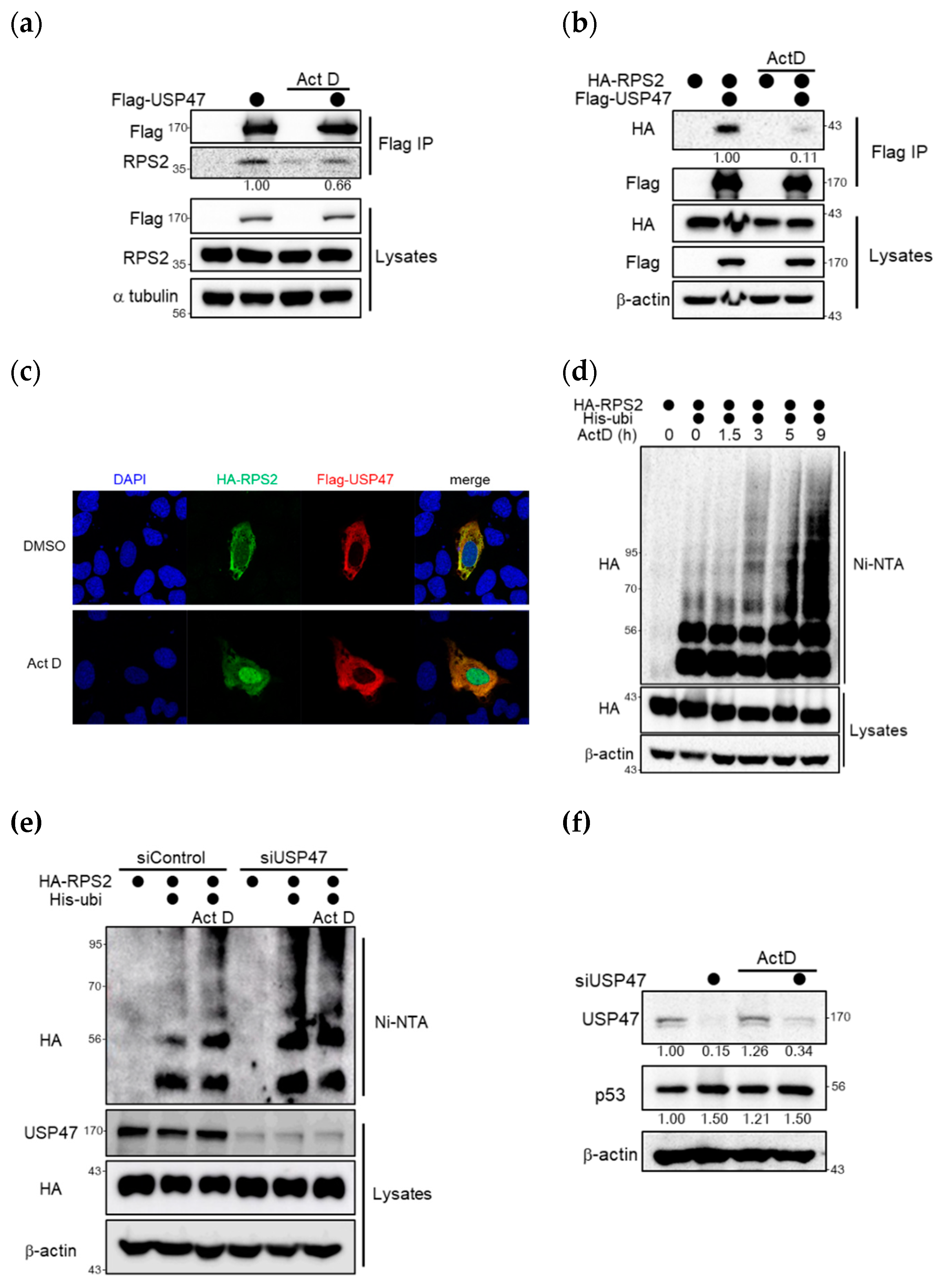
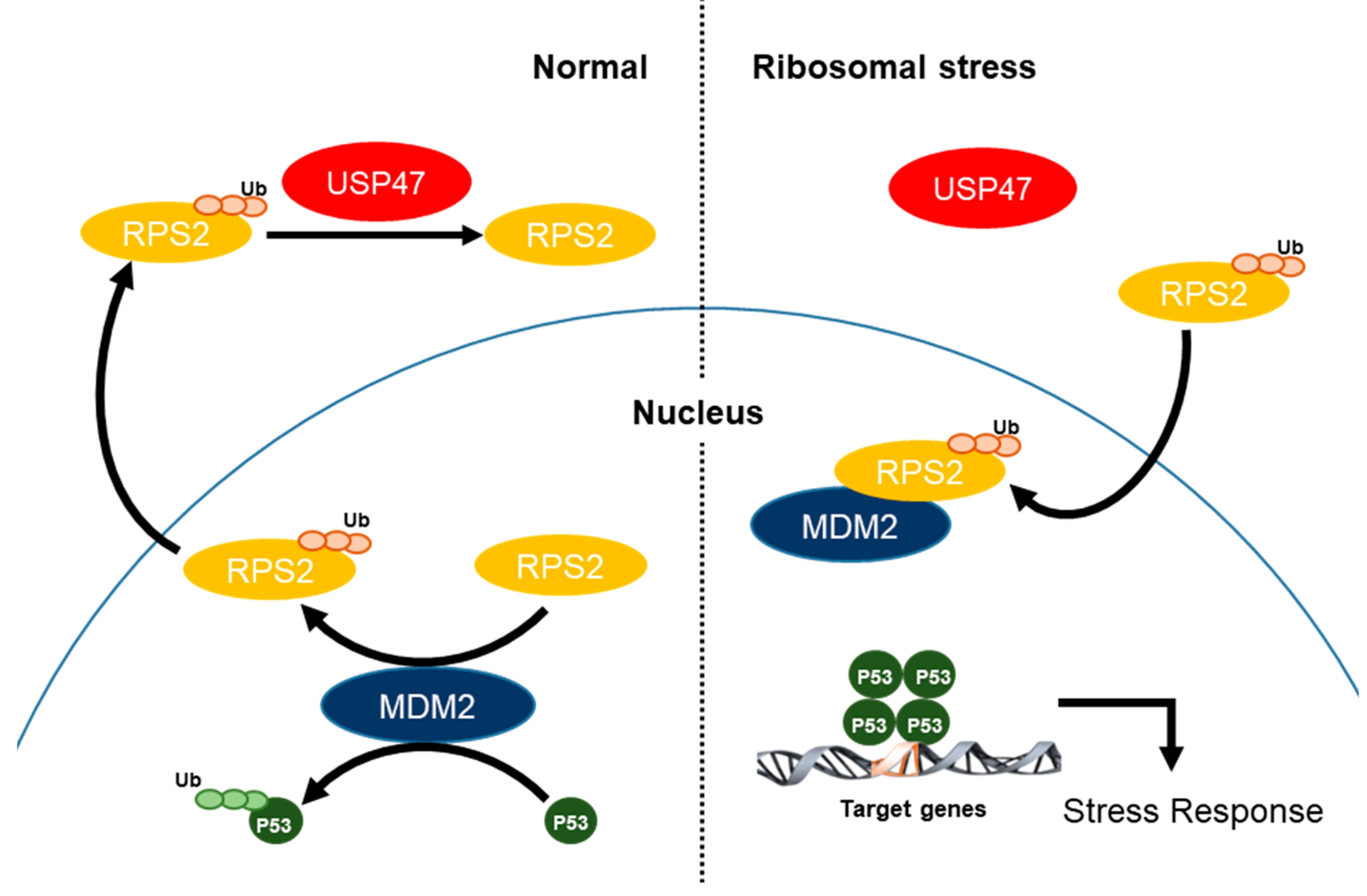
2.5. Ribosomal Stress Inhibits the Interaction between USP47 and RPS2 and thus Increases p53
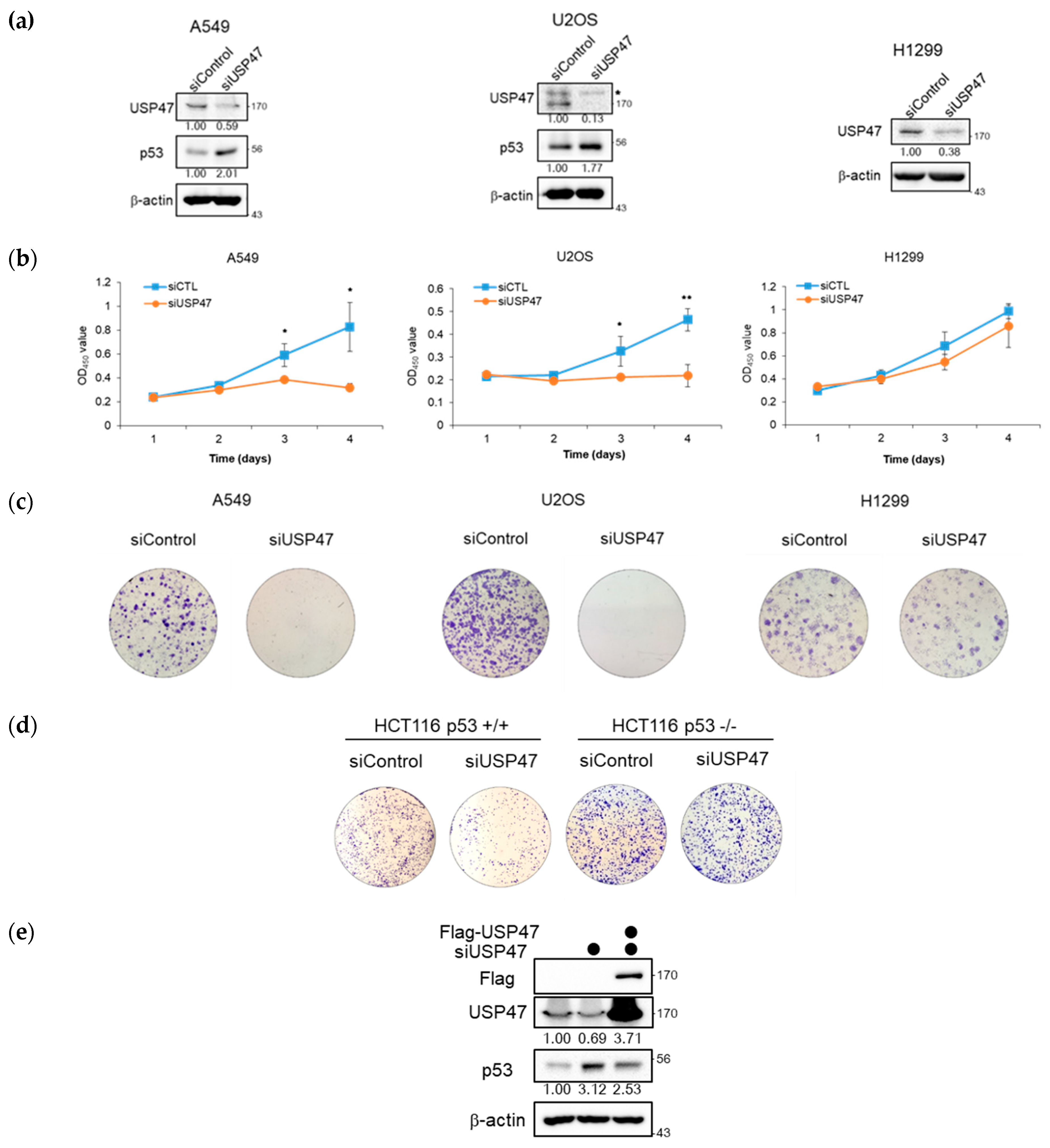
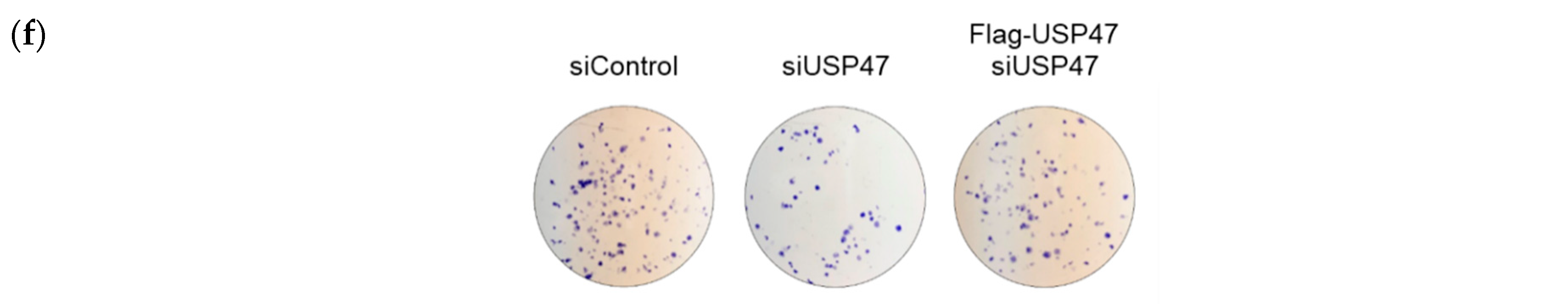
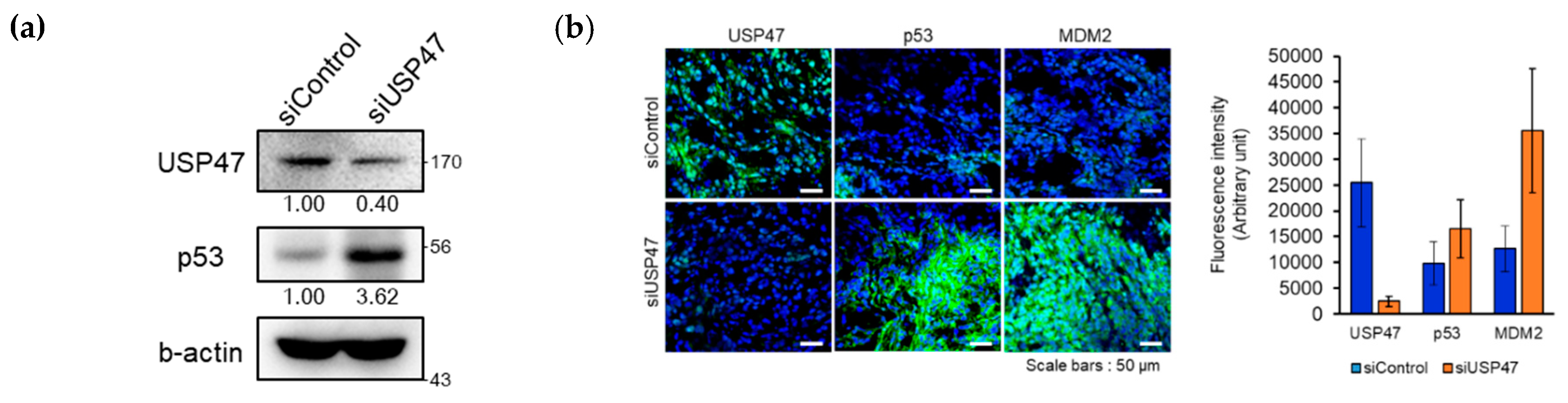
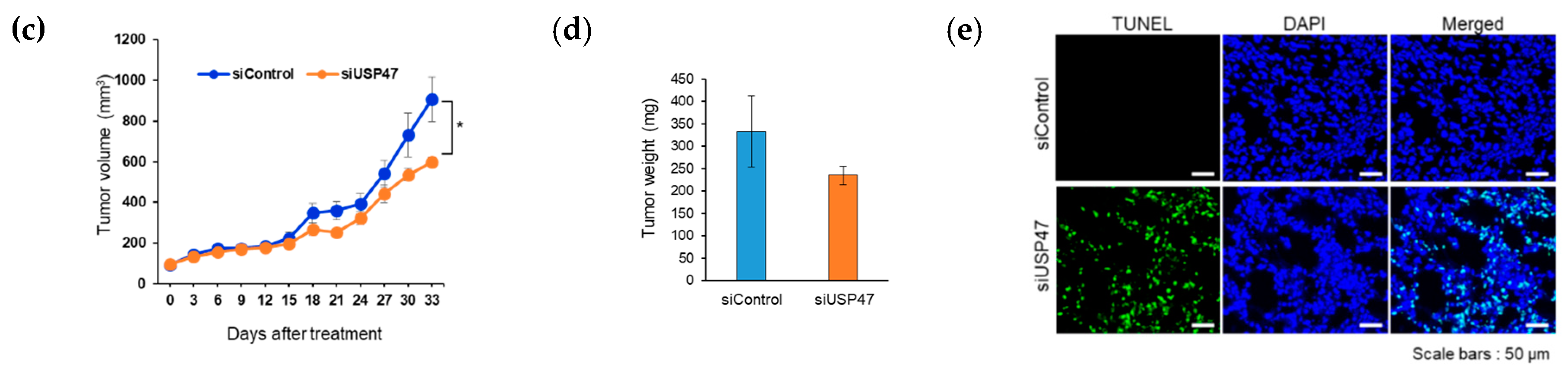
2.6. Suppression of USP47 Inhibits Cancer Cell Growth
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Transfection
4.2. Plasmids
4.3. Antibodies and Reagents
4.4. Immunoprecipitation
4.5. Ni-NTA Pulldown Assay
4.6. GST Pull-Down Assay
4.7. Quantitative Reverse Transcription-PCR Analysis
4.8. MTT Assay
4.9. Colony Formation Assay
4.10. A549 Tumor Xenografts
4.11. Tissue Immunofluorescence Staining and TUNEL Assay
4.12. Immunofluorescence
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Liu, D.; Xu, Y. P53, oxidative stress, and aging. Antioxid. Redox Signal. 2011, 15, 1669–1678. [Google Scholar] [CrossRef]
- Vousden, K.H.; Prives, C. Blinded by the light: The growing complexity of p53. Cell 2009, 137, 413–431. [Google Scholar] [CrossRef] [Green Version]
- Kruse, J.P.; Gu, W. Modes of p53 regulation. Cell 2009, 137, 609–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooks, C.L.; Gu, W. P53 ubiquitination: Mdm2 and beyond. Mol. Cell 2006, 21, 307–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Chen, D.; Shiloh, A.; Luo, J.; Nikolaev, A.Y.; Qin, J.; Gu, W. Deubiquitination of p53 by hausp is an important pathway for p53 stabilization. Nature 2002, 416, 648–653. [Google Scholar] [CrossRef] [PubMed]
- Khoronenkova, S.V.; Dianova, I.I.; Ternette, N.; Kessler, B.M.; Parsons, J.L.; Dianov, G.L. Atm-dependent downregulation of usp7/hausp by ppm1g activates p53 response to DNA damage. Mol. Cell 2012, 45, 801–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, J.; Luo, K.; Zhang, L.; Cheville, J.C.; Lou, Z. Usp10 regulates p53 localization and stability by deubiquitinating p53. Cell 2010, 140, 384–396. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Nemzow, L.; Chen, H.; Lubin, A.; Rong, X.; Sun, Z.; Harris, T.K.; Gong, F. The deubiquitinating enzyme usp24 is a regulator of the uv damage response. Cell Rep. 2015, 10, 140–147. [Google Scholar] [CrossRef] [Green Version]
- Hock, A.K.; Vigneron, A.M.; Carter, S.; Ludwig, R.L.; Vousden, K.H. Regulation of p53 stability and function by the deubiquitinating enzyme usp42. EMBO J. 2011, 30, 4921–4930. [Google Scholar] [CrossRef]
- Stevenson, L.F.; Sparks, A.; Allende-Vega, N.; Xirodimas, D.P.; Lane, D.P.; Saville, M.K. The deubiquitinating enzyme usp2a regulates the p53 pathway by targeting mdm2. EMBO J. 2007, 26, 976–986. [Google Scholar] [CrossRef] [Green Version]
- Palazon-Riquelme, P.; Worboys, J.D.; Green, J.; Valera, A.; Martin-Sanchez, F.; Pellegrini, C.; Brough, D.; Lopez-Castejon, G. Usp7 and usp47 deubiquitinases regulate nlrp3 inflammasome activation. EMBO Rep. 2018, 19. [Google Scholar] [CrossRef]
- Zhang, X.; Berger, F.G.; Yang, J.; Lu, X. Usp4 inhibits p53 through deubiquitinating and stabilizing arf-bp1. EMBO J. 2011, 30, 2177–2189. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Yang, H.; Kong, Q.; Li, J.; Lee, S.M.; Gao, B.; Dong, H.; Wei, J.; Song, J.; Zhang, D.D.; et al. Usp22 antagonizes p53 transcriptional activation by deubiquitinating sirt1 to suppress cell apoptosis and is required for mouse embryonic development. Mol. Cell 2012, 46, 484–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.X.; Challagundla, K.B.; Dai, M.S. Positive regulation of p53 stability and activity by the deubiquitinating enzyme otubain 1. EMBO J. 2012, 31, 576–592. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Liao, W.J.; Liao, J.M.; Liao, P.; Lu, H. Ribosomal proteins: Functions beyond the ribosome. J. Mol. Cell Biol. 2015, 7, 92–104. [Google Scholar] [CrossRef] [Green Version]
- Bursac, S.; Brdovcak, M.C.; Pfannkuchen, M.; Orsolic, I.; Golomb, L.; Zhu, Y.; Katz, C.; Daftuar, L.; Grabusic, K.; Vukelic, I.; et al. Mutual protection of ribosomal proteins l5 and l11 from degradation is essential for p53 activation upon ribosomal biogenesis stress. Proc. Natl. Acad. Sci. USA 2012, 109, 20467–20472. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Poyurovsky, M.V.; Li, Y.; Biderman, L.; Stahl, J.; Jacq, X.; Prives, C. Ribosomal protein s7 is both a regulator and a substrate of mdm2. Mol. Cell 2009, 35, 316–326. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.X.; DeVine, T.; Challagundla, K.B.; Dai, M.S. Interplay between ribosomal protein s27a and mdm2 protein in p53 activation in response to ribosomal stress. J. Biol. Chem. 2011, 286, 22730–22741. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.; Park, J.; Shin, S.C.; Kim, J.H.; Kim, E.E.; Song, E.J. Ribosomal protein s2 interplays with mdm2 to induce p53. Biochem. Biophys. Res. Commun. 2020, 523, 542–547. [Google Scholar] [CrossRef]
- Turnbull, A.P.; Ioannidis, S.; Krajewski, W.W.; Pinto-Fernandez, A.; Heride, C.; Martin, A.C.L.; Tonkin, L.M.; Townsend, E.C.; Buker, S.M.; Lancia, D.R.; et al. Molecular basis of usp7 inhibition by selective small-molecule inhibitors. Nature 2017, 550, 481–486. [Google Scholar] [CrossRef]
- Kategaya, L.; Di Lello, P.; Rouge, L.; Pastor, R.; Clark, K.R.; Drummond, J.; Kleinheinz, T.; Lin, E.; Upton, J.P.; Prakash, S.; et al. Usp7 small-molecule inhibitors interfere with ubiquitin binding. Nature 2017, 550, 534–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamberto, I.; Liu, X.; Seo, H.S.; Schauer, N.J.; Iacob, R.E.; Hu, W.; Das, D.; Mikhailova, T.; Weisberg, E.L.; Engen, J.R.; et al. Structure-guided development of a potent and selective non-covalent active-site inhibitor of usp7. Cell Chem. Biol. 2017, 24, 1490–1500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poondla, N.; Chandrasekaran, A.P.; Kim, K.S.; Ramakrishna, S. Deubiquitinating enzymes as cancer biomarkers: New therapeutic opportunities? BMB Rep. 2019, 52, 181–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faesen, A.C.; Luna-Vargas, M.P.; Sixma, T.K. The role of ubl domains in ubiquitin-specific proteases. Biochem. Soc. Trans. 2012, 40, 539–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kayama, K.; Watanabe, S.; Takafuji, T.; Tsuji, T.; Hironaka, K.; Matsumoto, M.; Nakayama, K.I.; Enari, M.; Kohno, T.; Shiraishi, K.; et al. Grwd1 negatively regulates p53 via the rpl11-mdm2 pathway and promotes tumorigenesis. EMBO Rep. 2017, 18, 123–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, M.; Kawahara, K.; Nishio, M.; Mimori, K.; Kogo, R.; Hamada, K.; Itoh, B.; Wang, J.; Komatsu, Y.; Yang, Y.R.; et al. Regulation of the mdm2-p53 pathway and tumor growth by pict1 via nucleolar rpl11. Nat. Med. 2011, 17, 944–951. [Google Scholar] [CrossRef] [Green Version]
- Fang, Z.; Cao, B.; Liao, J.M.; Deng, J.; Plummer, K.D.; Liao, P.; Liu, T.; Zhang, W.; Zhang, K.; Li, L.; et al. Spin1 promotes tumorigenesis by blocking the ul18 (universal large ribosomal subunit protein 18)-mdm2-p53 pathway in human cancer. eLife 2018, 7, e31275. [Google Scholar] [CrossRef]
- Holmberg Olausson, K.; Nister, M.; Lindstrom, M.S. P53 -dependent and -independent nucleolar stress responses. Cells 2012, 1, 774–798. [Google Scholar] [CrossRef] [Green Version]
- Ashcroft, M.; Taya, Y.; Vousden, K.H. Stress signals utilize multiple pathways to stabilize p53. Mol. Cell. Biol. 2000, 20, 3224–3233. [Google Scholar] [CrossRef] [Green Version]
- Choi, B.J.; Park, S.A.; Lee, S.Y.; Cha, Y.N.; Surh, Y.J. Hypoxia induces epithelial-mesenchymal transition in colorectal cancer cells through ubiquitin-specific protease 47-mediated stabilization of snail: A potential role of sox9. Sci. Rep. 2017, 7, 15918. [Google Scholar] [CrossRef] [Green Version]
- Parsons, J.L.; Dianova, I.I.; Khoronenkova, S.V.; Edelmann, M.J.; Kessler, B.M.; Dianov, G.L. Usp47 is a deubiquitylating enzyme that regulates base excision repair by controlling steady-state levels of DNA polymerase beta. Mol. Cell 2011, 41, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Peschiaroli, A.; Skaar, J.R.; Pagano, M.; Melino, G. The ubiquitin-specific protease usp47 is a novel beta-trcp interactor regulating cell survival. Oncogene 2010, 29, 1384–1393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macias, E.; Jin, A.; Deisenroth, C.; Bhat, K.; Mao, H.; Lindstrom, M.S.; Zhang, Y. An arf-independent c-myc-activated tumor suppression pathway mediated by ribosomal protein-mdm2 interaction. Cancer Cell 2010, 18, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Mahata, B.; Sundqvist, A.; Xirodimas, D.P. Recruitment of rpl11 at promoter sites of p53-regulated genes upon nucleolar stress through nedd8 and in an mdm2-dependent manner. Oncogene 2012, 31, 3060–3071. [Google Scholar] [CrossRef] [Green Version]
- Ofir-Rosenfeld, Y.; Boggs, K.; Michael, D.; Kastan, M.B.; Oren, M. Mdm2 regulates p53 mrna translation through inhibitory interactions with ribosomal protein l26. Mol. Cell 2008, 32, 180–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donati, G.; Peddigari, S.; Mercer, C.A.; Thomas, G. 5s ribosomal rna is an essential component of a nascent ribosomal precursor complex that regulates the hdm2-p53 checkpoint. Cell Rep. 2013, 4, 87–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sloan, K.E.; Bohnsack, M.T.; Watkins, N.J. The 5s rnp couples p53 homeostasis to ribosome biogenesis and nucleolar stress. Cell Rep. 2013, 5, 237–247. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Wang, J.; Chen, C.; Yuan, H.; Wen, X.; Sun, H. Usp7: Target validation and drug discovery for cancer therapy. Med. Chem. 2018, 14, 3–18. [Google Scholar] [CrossRef]
- Fan, Y.H.; Cheng, J.; Vasudevan, S.A.; Dou, J.; Zhang, H.; Patel, R.H.; Ma, I.T.; Rojas, Y.; Zhao, Y.; Yu, Y.; et al. Usp7 inhibitor p22077 inhibits neuroblastoma growth via inducing p53-mediated apoptosis. Cell Death Dis. 2013, 4, e867. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Zhang, Y.; Wang, T.; Zhang, J.; Zhou, Z.; Sun, Y.; Wang, S.; Shi, Y.; Luan, X.; Zhang, Y.; et al. The usp7 inhibitor p5091 induces cell death in ovarian cancers with different p53 status. Cell. Physiol. Biochem. 2017, 43, 1755–1766. [Google Scholar] [CrossRef]
- Weinstock, J.; Wu, J.; Cao, P.; Kingsbury, W.D.; McDermott, J.L.; Kodrasov, M.P.; McKelvey, D.M.; Suresh Kumar, K.G.; Goldenberg, S.J.; Mattern, M.R.; et al. Selective dual inhibitors of the cancer-related deubiquitylating proteases usp7 and usp47. ACS Med. Chem. Lett. 2012, 3, 789–792. [Google Scholar] [CrossRef] [Green Version]
- Barboza, J.A.; Iwakuma, T.; Terzian, T.; El-Naggar, A.K.; Lozano, G. Mdm2 and mdm4 loss regulates distinct p53 activities. Mol. Cancer Res. 2008, 6, 947–954. [Google Scholar] [CrossRef] [Green Version]
- Kingston, R.E.; Chen, C.A.; Okayama, H. Calcium phosphate transfection. Curr. Protoc. Cell Biol. 2003, 63, 9.1.1–9.1.11. [Google Scholar]
- Yang, S.W.; Oh, K.H.; Park, E.; Chang, H.M.; Park, J.M.; Seong, M.W.; Ka, S.H.; Song, W.K.; Park, D.E.; Baas, P.W.; et al. Usp47 and c terminus of hsp70-interacting protein (chip) antagonistically regulate katanin-p60-mediated axonal growth. J. Neurosci. 2013, 33, 12728–12738. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Lee, J.M.; Lee, G.; Bhin, J.; Oh, S.K.; Kim, K.; Pyo, K.E.; Lee, J.S.; Yim, H.Y.; Kim, K.I.; et al. DNA damage-induced roralpha is crucial for p53 stabilization and increased apoptosis. Mol. Cell 2011, 44, 797–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, E.J.; Werner, S.L.; Neubauer, J.; Stegmeier, F.; Aspden, J.; Rio, D.; Harper, J.W.; Elledge, S.J.; Kirschner, M.W.; Rape, M. The prp19 complex and the usp4sart3 deubiquitinating enzyme control reversible ubiquitination at the spliceosome. Genes Dev. 2010, 24, 1434–1447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.; Kwon, M.S.; Kim, E.E.; Lee, H.; Song, E.J. Usp35 regulates mitotic progression by modulating the stability of aurora b. Nat. Commun. 2018, 9, 688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, H.; Li, D.; Wu, T.; Xie, D.; Hua, K.; Hu, J.; Deng, X.; Ji, C.; Deng, Y.; Fang, L. Microrna-301b promotes cell proliferation and apoptosis resistance in triple-negative breast cancer by targeting cyld. BMB Rep. 2018, 51, 602–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]










© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, J.; Park, J.; Shin, S.C.; Jang, M.; Kim, J.-H.; Kim, E.E.; Song, E.J. USP47 Promotes Tumorigenesis by Negative Regulation of p53 through Deubiquitinating Ribosomal Protein S2. Cancers 2020, 12, 1137. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12051137
Cho J, Park J, Shin SC, Jang M, Kim J-H, Kim EE, Song EJ. USP47 Promotes Tumorigenesis by Negative Regulation of p53 through Deubiquitinating Ribosomal Protein S2. Cancers. 2020; 12(5):1137. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12051137
Chicago/Turabian StyleCho, Jinhong, Jinyoung Park, Sang Chul Shin, Mihue Jang, Jae-Hong Kim, Eunice EunKyeong Kim, and Eun Joo Song. 2020. "USP47 Promotes Tumorigenesis by Negative Regulation of p53 through Deubiquitinating Ribosomal Protein S2" Cancers 12, no. 5: 1137. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12051137