1. Introduction
Despite recent progress in diagnosis and therapy, non-small cell lung cancer (NSCLC) remains a deadly disease with survival rates having only minimally improved in the last decades [
1]. Even though novel targeted therapies and immune checkpoint inhibitors (ICIs) have been introduced in daily clinical practice, platinum-based chemotherapy remains the mainstay of treatment in early as well as in metastatic disease [
2]. Chemotherapy efficacy though is significantly hampered by the development of primary or acquired resistance in most of the patients [
3].
The main mechanism of action of cisplatin and its analogues is the formation of DNA adducts followed by intra- and interstrand cross-links that block DNA replication and induce DNA damage [
4]. DNA damage then results in the activation of Ataxia telangiectasia mutated (ATM) and Ataxia telangiectasia and Rad 3-related (ATR) kinases that phosphorylate a wide range of downstream targets within a complex network of signaling pathways operating in cell-cycle arrest, DNA repair, and apoptosis [
5]. Activation of the apoptotic machinery results in cell death, whereas successful lesion repair leads to cell survival and drug resistance [
6]. Reported mechanisms of cisplatin resistance include decreased intracellular accumulation of cis-diamminedichloridoplatinum (II) (CDDP), increased detoxification systems, impaired apoptotic signalling after DNA damage and DNA repair, or tolerance of the damage [
7].
MicroRNAs (miRNAs), a class of small noncoding RNAs, regulate gene expression by posttranscriptional repression of their mRNA targets [
8]. Cancer is associated with global alteration of miRNA expression patterns, where miRNAs have been shown to exert either oncogenic or tumor suppressive effects depending on the context [
9]. Each miRNA is implicated in a wide array of pathological processes through the regulation of multiple gene targets, and inversely, each mRNA can be targeted by multiple miRNAs [
10]. Conceivably, the expression pattern of a small number of miRNAs may reflect extensive alterations of gene expression networks involving hundreds of messenger RNAs [
11]. Bioinformatics provide the opportunity for systemic analysis of pathways and biological processes that are specifically regulated by multiple dysregulated miRNAs, thus resulting in meaningful insights in the mechanisms underlying different steps of cancer cell function and tumor progression [
12].
Accumulating evidence suggests that critical genes involved in DNA repair are epigenetically regulated by miRNAs [
13]. Furthermore, DNA damage response results in the transcriptional or posttranscriptional modulation of miRNAs [
13] which regulate DNA damage response according to the nature and intensity of DNA damage [
14]. In addition, miRNAs have been associated with response or resistance to various types of cancer drugs including cisplatin [
15].
Thus, miR-21 modulates cell cycle progression and DNA damage checkpoint activation via cell division cycle 25 phosphatase (Cdc25A), which is involved in cell cycle arrest in response to DNA damage [
16]. MiR-21 has been also associated with resistance of NSCLC cells to cisplatin through phosphatase and tensin homolog (PTEN) targeting and inhibition of apoptosis [
17], whereas its deletion sensitized cells to DNA-damaging chemotherapy [
18]. MiR-128 represses E2F transcription factor 5 (E2F5), leading to the transcriptional induction of cyclin-dependent kinase inhibitor 1A (
p21waf1). In turn, p21 protein is localized to the cytoplasm, where it exerts antiapoptotic function in response to cisplatin [
19]. MiR-155 silencing enhances the sensitivity of A549 lung cancer cells to cisplatin by activating the initiator caspase-9 through apoptotic peptidase activating factor 1 (Apaf-1) [
20]. In addition, miR-155 has been shown to target tumor protein P53 (TP53) through a miR-155/TP53 negative feedback mechanism, ultimately leading to cisplatin resistance [
21]. Upregulation of miR-181a results in BCL2-associated apoptosis regulator (Bax) oligomerization and caspase activation, thus enhancing cisplatin cytotoxicity [
22]. In addition, miR-181a/b negatively regulates DNA damage response by impairing the expression and activity of the stress-sensor kinase ATM [
23].
MiRNAs are detected in the plasma and other biological fluids in various physiologic and pathologic conditions including cancer [
24]. There is evidence suggesting that circulating miRNAs promote tumorigenesis and participate in cancer invasion, metastasis, and angiogenesis by delivering signals in distant sites [
25]. In addition, few studies show their function in modulating chemosensitivity [
26]. Accordingly, it is considered that circulating miRNAs could serve as important biomarkers reflecting the dynamic processes occurring during tumor evolution [
27] and predicting therapeutic response in cancer [
28]. According to the above, we hypothesized that miRNAs involved in the regulation of cisplatin cytotoxicity may have a role as circulating biomarkers for the prediction of therapeutic efficacy of platinum-based chemotherapy in NSCLC. Thus, in the current report, we investigated the clinical significances of miR-21, miR-128, miR-155, and miR-181a expressions in the plasma of patients with NSCLC treated with first-line platinum-based doublets. As a great number of protein-coding mRNA targets have been reported for the four miRNAs under study, we performed a detailed bioinformatics analysis in order to narrow down and to prioritize the associated genes and pathways in the context of our study.
3. Discussion
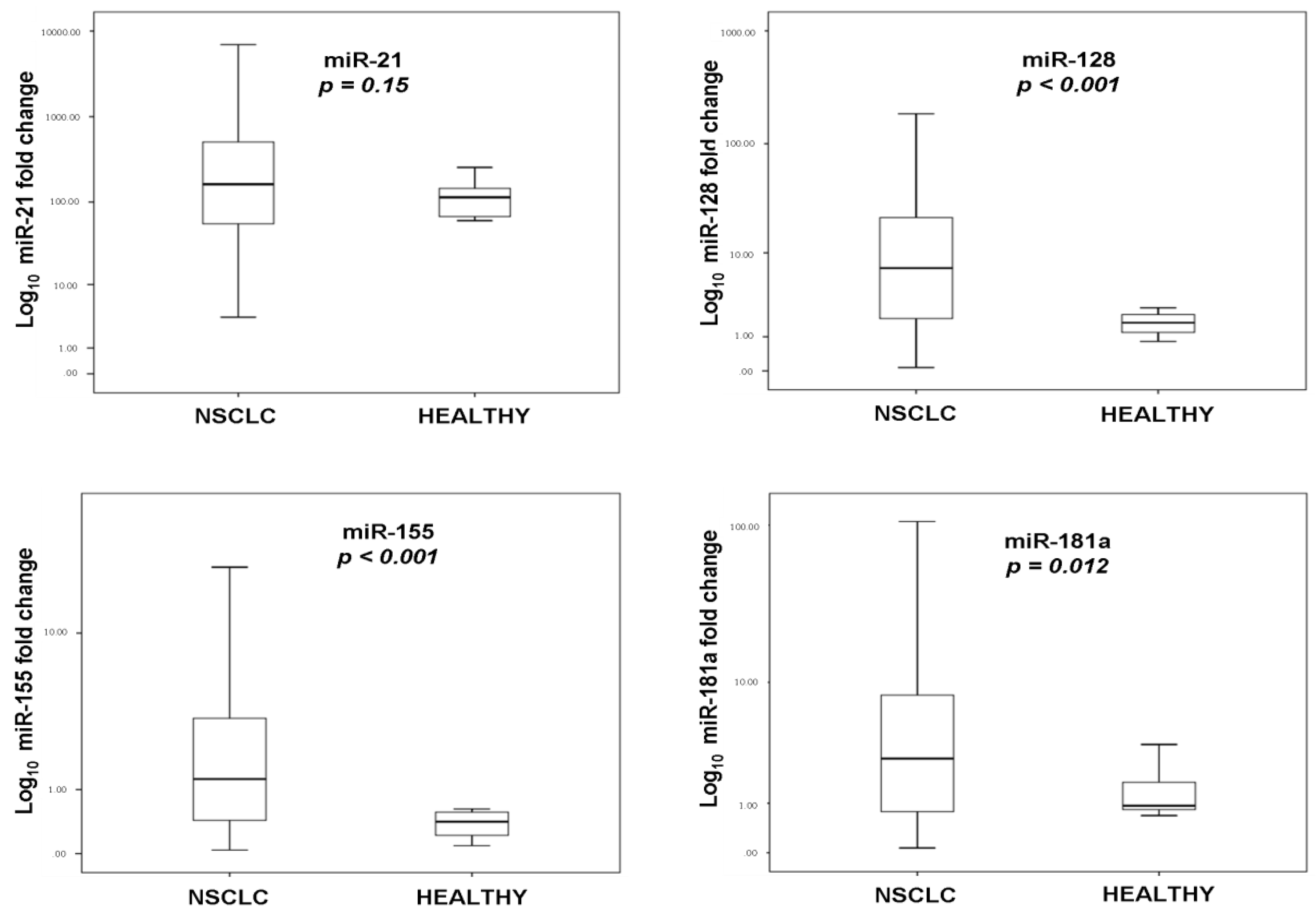
Over the last few years, circulating miRNAs have been recognized as potential diagnostic and prognostic biomarkers in patients with cancer [
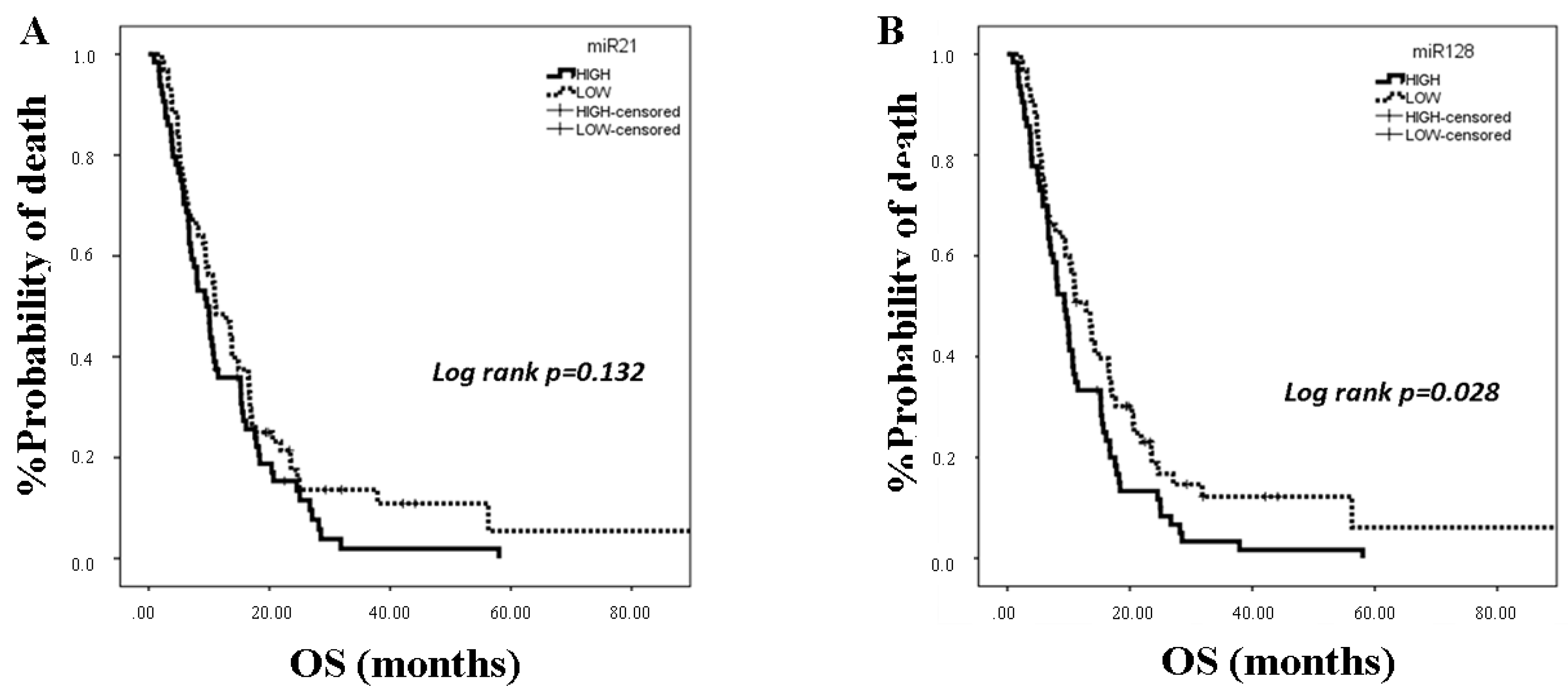
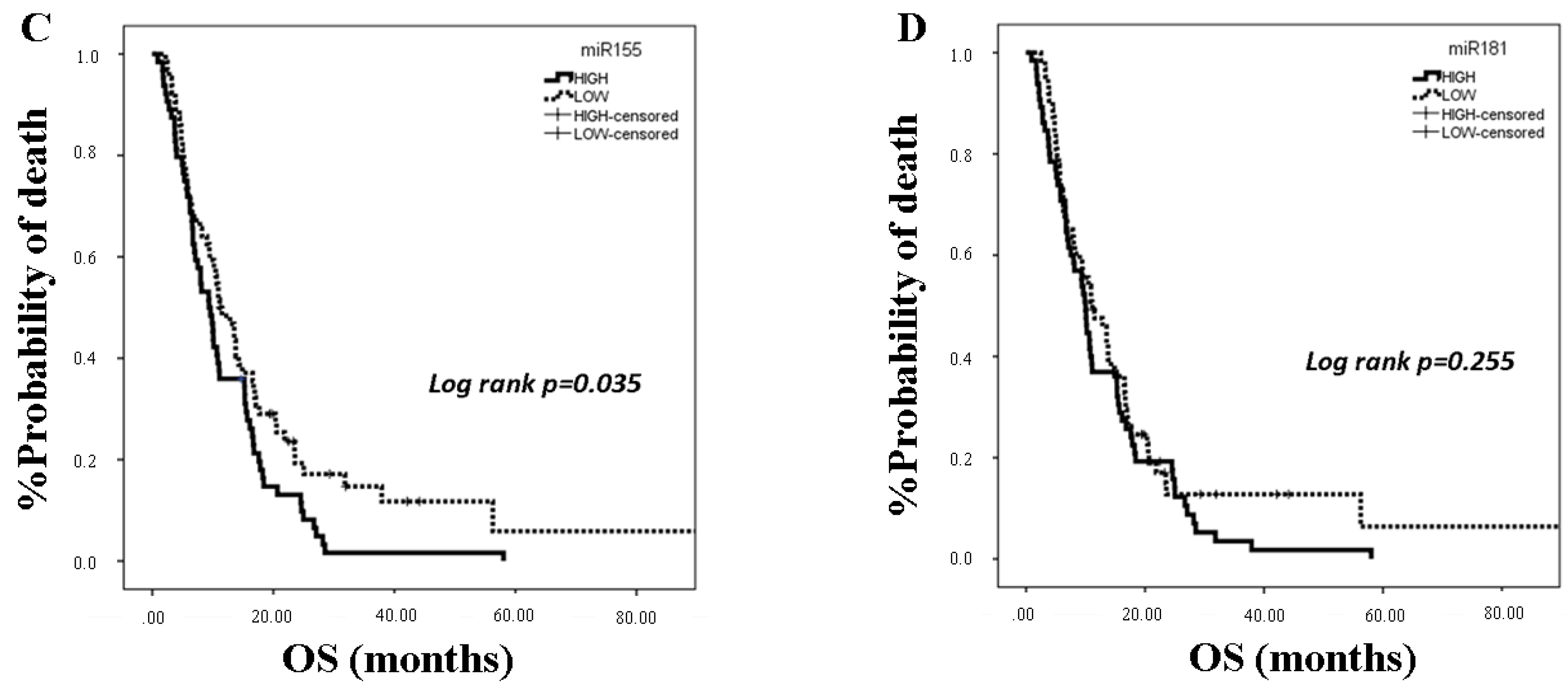
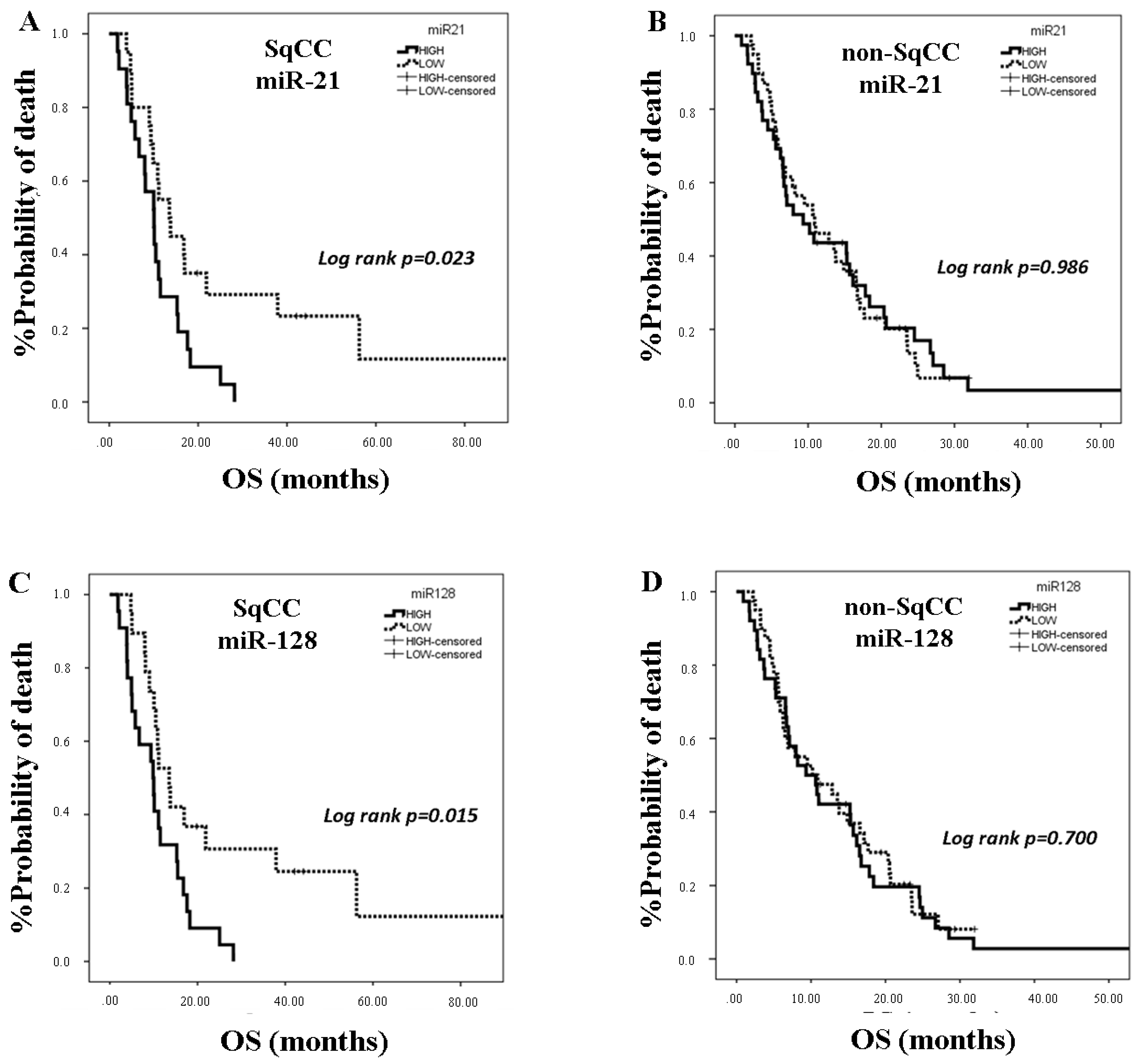
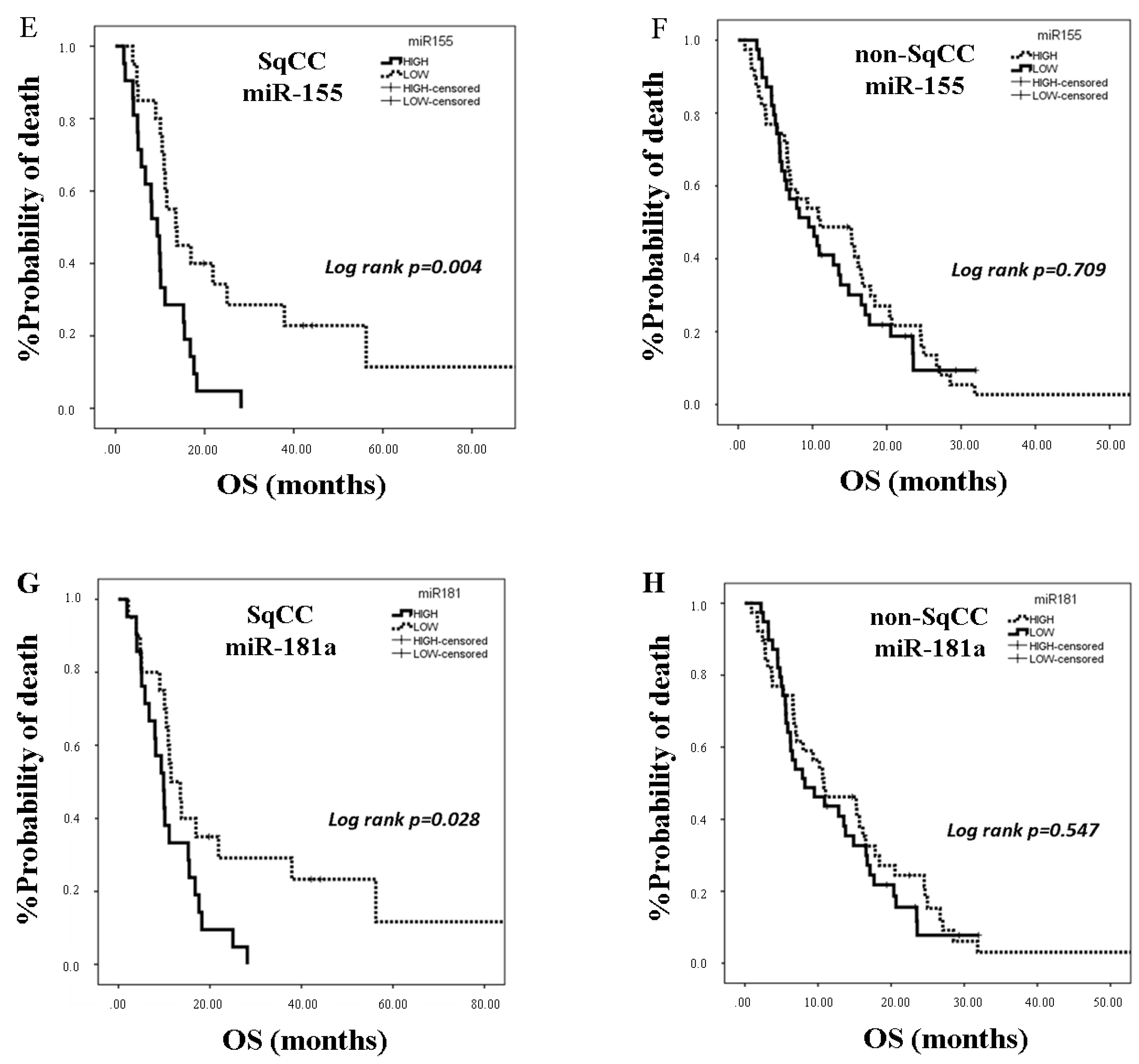
25]. In the present study, we investigated the clinical relevance of plasma miR-21, miR-128, miR-155, and miR-181a in NSCLC patients treated with first-line platinum-based doublets and demonstrated that high expressions of circulating miR-128 and miR-155 were associated with shorter OS. In the SqCC subgroup, patients with high miR-21, miR-128, miR-155, and miR-181 had decreased OS compared to those with low expression values. Importantly, high miR-128 independently predicted for worse OS in the whole population whereas high miR-155 emerged as an independent predictor for worse OS in squamous NSCLC. Bioinformatics analysis of mRNA targets revealed interesting associations related to potential molecular mechanisms of response or resistance to treatment regulated by these miRNAs.
MiR-128, via Drosha ribonuclease III (Drosha) and Dicer ribonuclease III (Dicer) targeting, is a key regulator of the malignant phenotype in lung cancer cells by promoting epithelial to mesenchymal transition (EMT) and cell migration [
30]. In addition, miR-128-3p has been reported as the only miRNA significantly upregulated in NSCLC as compared with normal tissue in The Cancer Genome Atlas (TCGA) lung cancer data sets [
31]. Similarly, in a large-scale analysis of miRNA profiles, miR-128-3p emerged amongst the most commonly upregulated miRs in lung cancer tissue [
32]. Importantly, in a cohort of NSCLC patients treated with cisplatin-based chemotherapy, high tumor miR-128 independently predicted for worse outcome [
31]. Furthermore, miR-128 expression in the whole blood could distinguish early lung cancer patients from healthy donors [
33]. We here demonstrate for the first time that high plasma miR-128 is an independent prognostic factor for worse survival in NSCLC patients treated with first-line platinum doublets.
MiR-155 is an oncomir with regulatory roles in tumor growth, EMT, metastasis, apoptosis, and response to chemotherapy [
21,
34]. MiR-155, along with miR-21, is among the most frequently amplified miRNA genes in NSCLC [
35]. Unique miRNA profiling studies demonstrated that lung adenocarcinoma or SqCC presented higher miR-155 expression levels compared to normal tissues and suggested that its oncogenic role [
34] is possibly related to direct inhibition of tumor suppressors, PTEN, and the suppressors of cytokine signalling 1 and 6 (SOCS1 and SOCS6) [
36]. High tumor miR-155 has been demonstrated as an independent prognostic factor for poor overall survival in patients with lung adenocarcinoma [
34] or SqCC [
37]. Moreover, miR-155 was previously demonstrated to be differentially expressed in the plasma of patients with early NSCLC compared to healthy donors and was included in the plasma signature of 3 miRNAs associated with increased risk for progression in resected lung adenocarcinoma [
38]. Furthermore, in a small cohort of NSCLC patients, Gao et al. showed that serum levels of miR-155 in combination with the tumor markers carcinoembryonic antigen (CEA) and CA-125 increased the efficiency for the early diagnosis of lung adenocarcinoma [
39]. Our results further support the association of miR-155 with poor prognosis in NSCLC, and they show for the first time that plasma miR-155 represents an independent poor prognostic indicator in the squamous subtype.
MiR-21 and miR-155 share nearly 30% of their predicted targets [
36]. In a previous report, both miRNAs expressed in tumor tissue, promoted NSCLC progression, and predicted recurrence and unfavorable survival of patients with stage I-III NSCLC [
36,
40]. Moreover, plasma miR-21 and miR-155 were amongst the miRNAs with the highest performance to discriminate patients with SqCC among early stage NSCLC patients [
41]. In addition, high serum miR-21 expression was correlated with shorter survival in patients with early stage NSCLC [
42]. We here show for the first time that high plasma miR-21 was associated with shorter OS in the SqCC subtype in patients with advanced stages of the disease and that circulating miR-21 and miR-155 expression levels were strongly correlated, potentially explaining the independent prognostic role for miR-155 only.
MiR-181 family members operate as tumor suppressors in lung cancer in contrast to their role in other tumor types [
43]. Low miR-181a expression in NSCLC tissue was significantly correlated with poor patient survival [
43]. We here demonstrate for the first time that plasma miR-181a expression levels are prognostic in NSCLC patients treated with first-line chemotherapy. Our findings regarding the association of high plasma miR-181a with shorter OS in the squamous patient subgroup potentially suggest a differential biological role of circulating miR-181a in SqCC NSCLC.
It should be noted here that, although the investigated miRNAs were selected based on their implications in DNA damage response and/or the modulation of response to the cytotoxic effects of cisplatin, no associations were found between their plasma levels and response rates or PFS with platinum doublets. These observations imply that they may have a role in determining prognosis in patients treated with platinum-based doublets. Additional studies are required to further define their prognostic relevance in NSCLC patients treated with other modalities.
Tumor-associated circulating miRNAs are derived from tumor cell death and lyses or from active secretion by the tumor [
44]. However, a significant proportion originates from immune cells in the blood and the tumor microenvironment or from other organs, potentially representing the host’s response to the presence of the tumor [
44]. In NSCLC, distinct deregulated expression profiles of circulating miRNAs have been previously associated with disease progression, prognosis, or drug resistance, thus reflecting distinct genes and pathways [
27]. It is however debated whether circulating miRNAs represent simple disease by-products indicative of these processes or whether they actively participate in their regulation [
25,
45,
46]. Circulating miRNAs have indeed been suggested to modulate tumor biology by altering the cellular transcriptome of recipient tumor cells in a paracrine fashion or by delivering signals in distant sites [
25,
45]. Considering the high tissue specificity of miRNAs [
46], the possibility of circulating cancer-associated miRNAs being secreted from blood immune cells to promote or inhibit cancer cell proliferation, invasion, apoptosis, and antitumor immune response by delivering signals to recipient cells cannot be excluded [
44]. Thus, miR-21, miR-155, and miR-181a have been involved in both immune response and cancer [
47,
48,
49], indicating that deranged expression of these miRNAs may represent an important link between antitumor immunity and cancer progression [
44]. Interestingly, we have previously shown that high circulating miR-21 was associated with disease relapse and shorter disease-free and overall survival in patients with early breast cancer [
50].
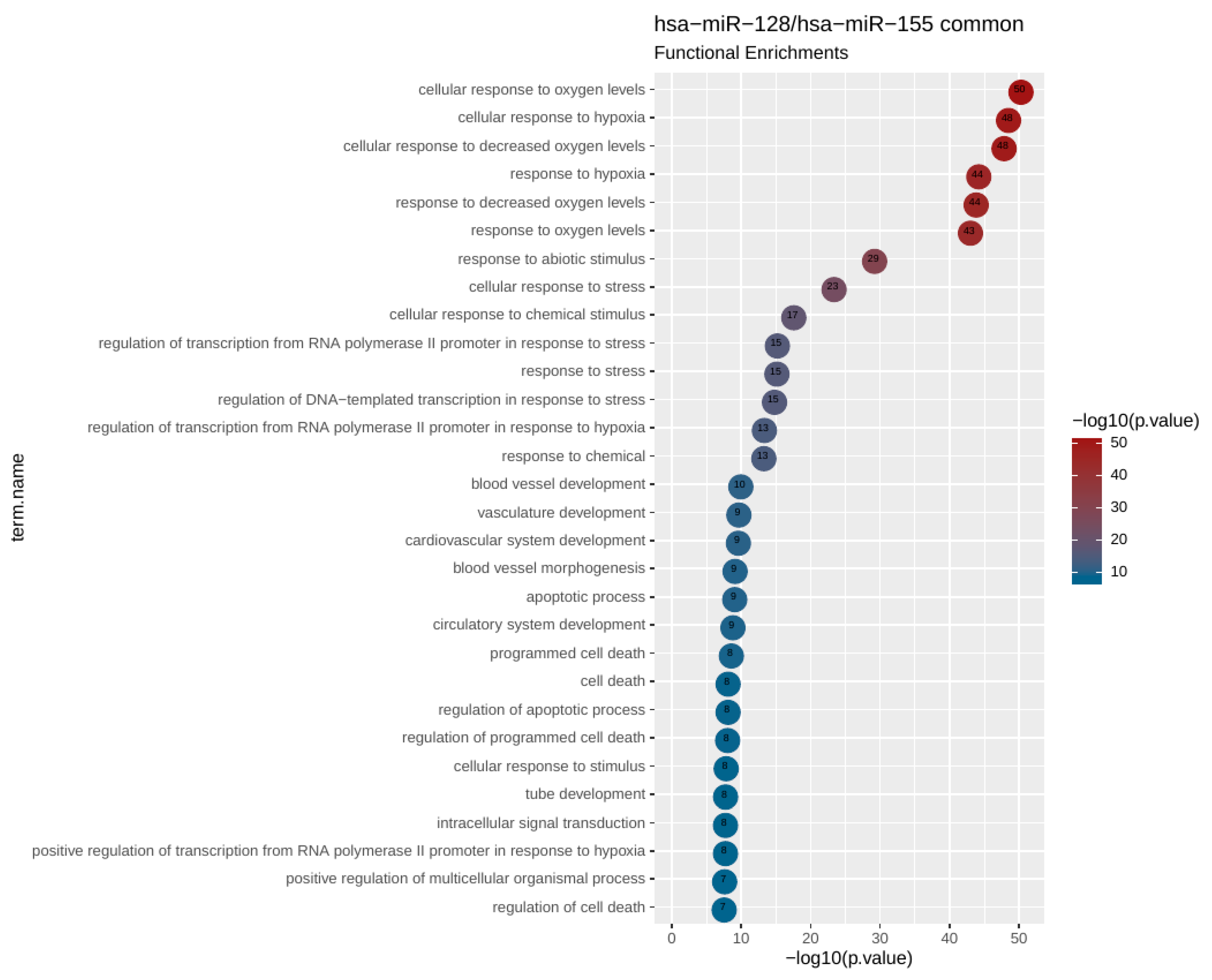
To uncover potential genes and pathways that could be regulated by the investigated circulating miRNAs, we performed bioinformatics analysis revealing that miR-128 and miR-155 shared 26 target genes in pathways related to response to hypoxia. Further focused analysis of a 26 hypoxia-related gene set revealed additional pathways associated with vasculature development, angiogenesis, apoptosis, and cell death.
Tumor hypoxia has been correlated with tumor aggressiveness and metastasis as well as with adverse outcome in several tumor types [
51], suggesting that these miRNAs may promote tumor progression through the regulation of lung cancer cell response to hypoxic states. Interestingly, significant preclinical evidence exists on the role of hypoxia in enhancing DNA damage and mutagenesis as well as in the functional impairment of key genes in DNA repair pathways, ultimately resulting in genomic instability and tumor progression [
52]. Moreover, alterations in cell type or tissue-specific miRNA profiles have been reported in response to hypoxia, which then modulates the expression of key components of DNA repair pathways [
53,
54]. Thus, hypoxia-induced miR-155 has been shown to promote radioresistance [
55], whereas in another work, it was implicated in DNA damage response by targeting mutL homolog 1 (MLH1) and mutS homolog 2 (MSH2) involved in DNA mismatch repair [
56].
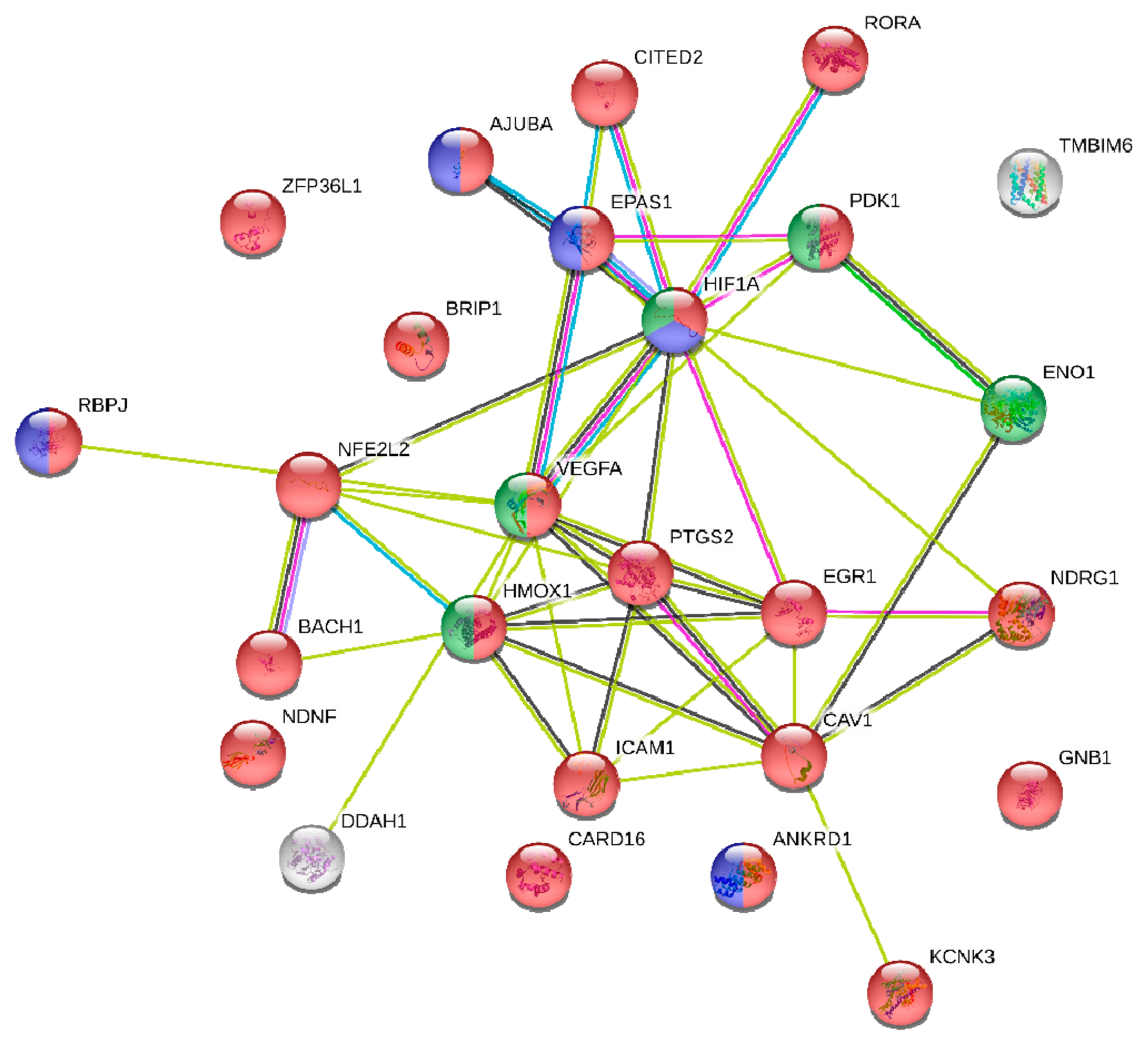
Bioinformatics analysis revealed that hypoxia-inducible factor-1a (HIF-1a), a central regulator of the transcriptional response to hypoxia, was amongst the 26 hypoxia-related gene set of common targets of miR-128 and miR-155. HIF-1a induces miR-155 expression, which in turn targets HIF-1a mRNA, suggesting that miR-155 is a component of a HIF-1α regulatory network during hypoxia [
57]. Accordingly, in another study, hypoxia led to vascular endothelial growth factor (VEGF)-induced miR-155 expression that regulated HIF-1a expression and enforced endothelial cell maturation and angiogenesis [
58]. In accordance to the above studies, integrated function analysis implicated miR-155 in the regulation of HIF-1a expression.
In the current study, we carefully considered pre-analytical and analytical variables that may influence miRNA detection and quantification; however, methodological approaches for circulating biomarkers need to be validated and standardized across laboratories [
59,
60]. It should be also noted that our findings regarding the prognostic role of the investigated miRNAs are derived from a relatively small group of patients and lack validation in an independent patient cohort. Identification and further analyses of miRNA protein-coding gene targets relied exclusively on prior knowledge as documented in a reference database [
12]. Nevertheless, one should take note of the limitations of such an analysis due to the large numbers of reported targets (even when restricting to cancerous datasets) that are highly likely to reflect the miRNA regulatory potential in variable environments [
61]. On the other hand, the significant gene and functional overlaps that we found among the targets of miR-128 and miR-155 suggest that our findings are rather robust and reflect a genuine functional link between the two miRNAs, a link that appears to be quite strong in functions related to hypoxia. Last but not least, out of the 26 common genes targets identified with highly stringent criteria, 23 (>88%) were shown to carry gene regulatory function. This abundance of transcriptional regulators, revealed in our network analysis, is suggestive of a possible regulatory circuitry, in which mir-155 and mir-128 are likely to be key players.
In summary, our results suggest for the first time that pretreatment levels of circulating miR-128 and miR-155 could serve as promising prognostic biomarkers in NSCLC patients treated with first-line platinum-based doublets. Furthermore, using a bioinformatics approach, we showed that these miRNAs are involved in major biological processes in cancer and we highlighted a link between hypoxia, DNA repair, and chemoresistance. Since extracellular miRNAs represent a means of cell–cell communication [
25], miR-128 and miR-155 may promote these effects in recipient cells. In addition, although hypoxia-targeted therapy trials in NSCLC have not as yet been translated into patient benefit, if the investigated miRNAs are indeed functional, they could be further exploited therapeutically, taking into account potential histology-driven differences.

 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}