TRPV1 Antagonist DWP05195 Induces ER Stress-Dependent Apoptosis through the ROS-p38-CHOP Pathway in Human Ovarian Cancer Cells


Abstract
:1. Introduction
2. Results
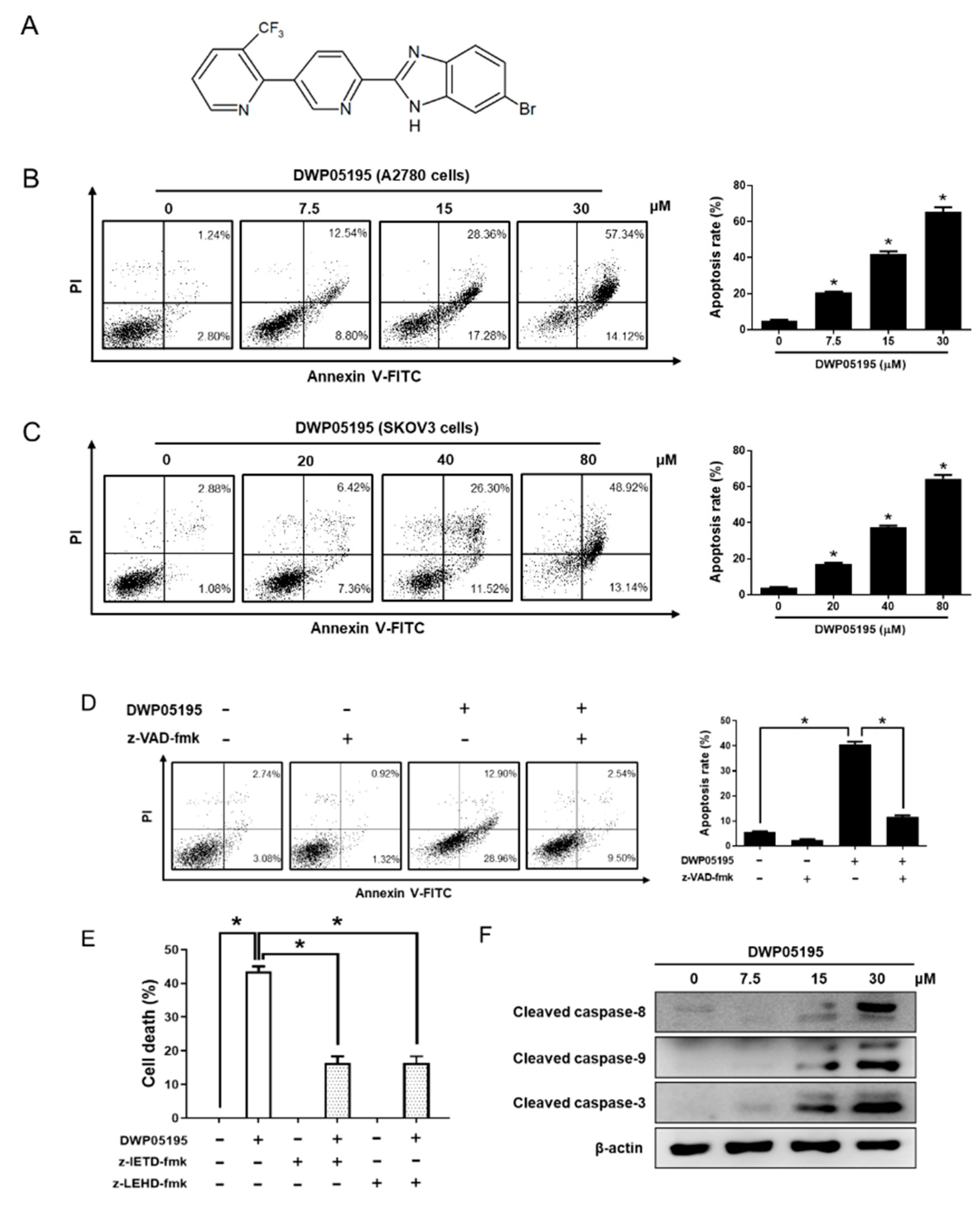
2.1. DWP05195 Stimulates Caspase-Dependent Apoptosis in Human Ovarian Cancer Cells
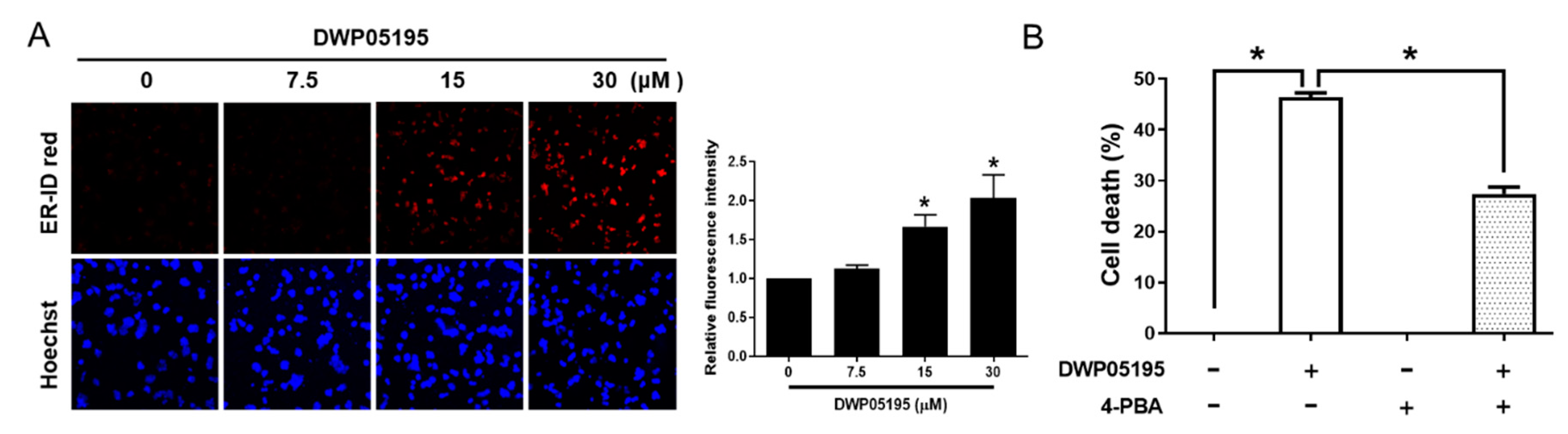
2.2. ER Stress is Involved in DWP05195-Induced Apoptosis
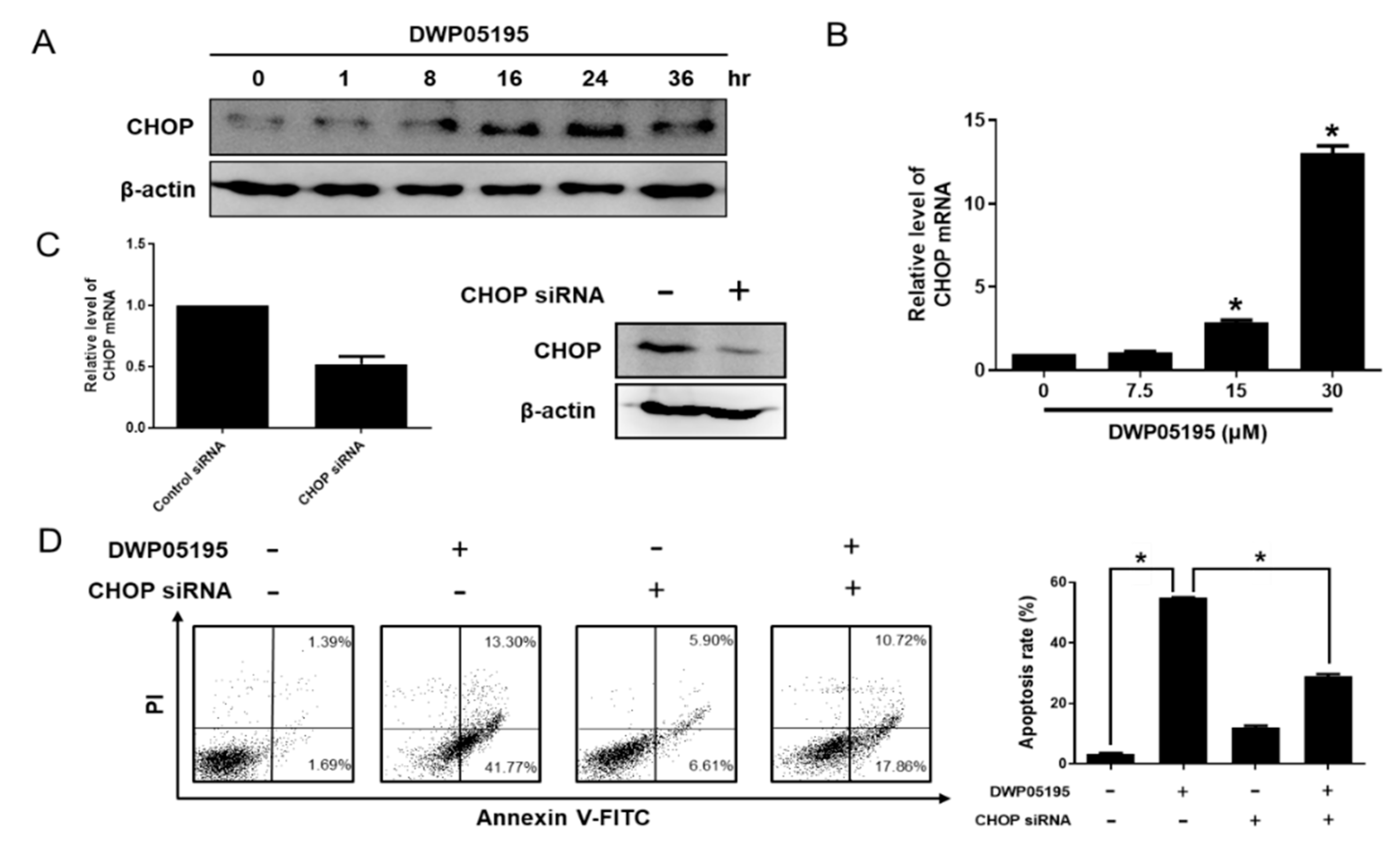
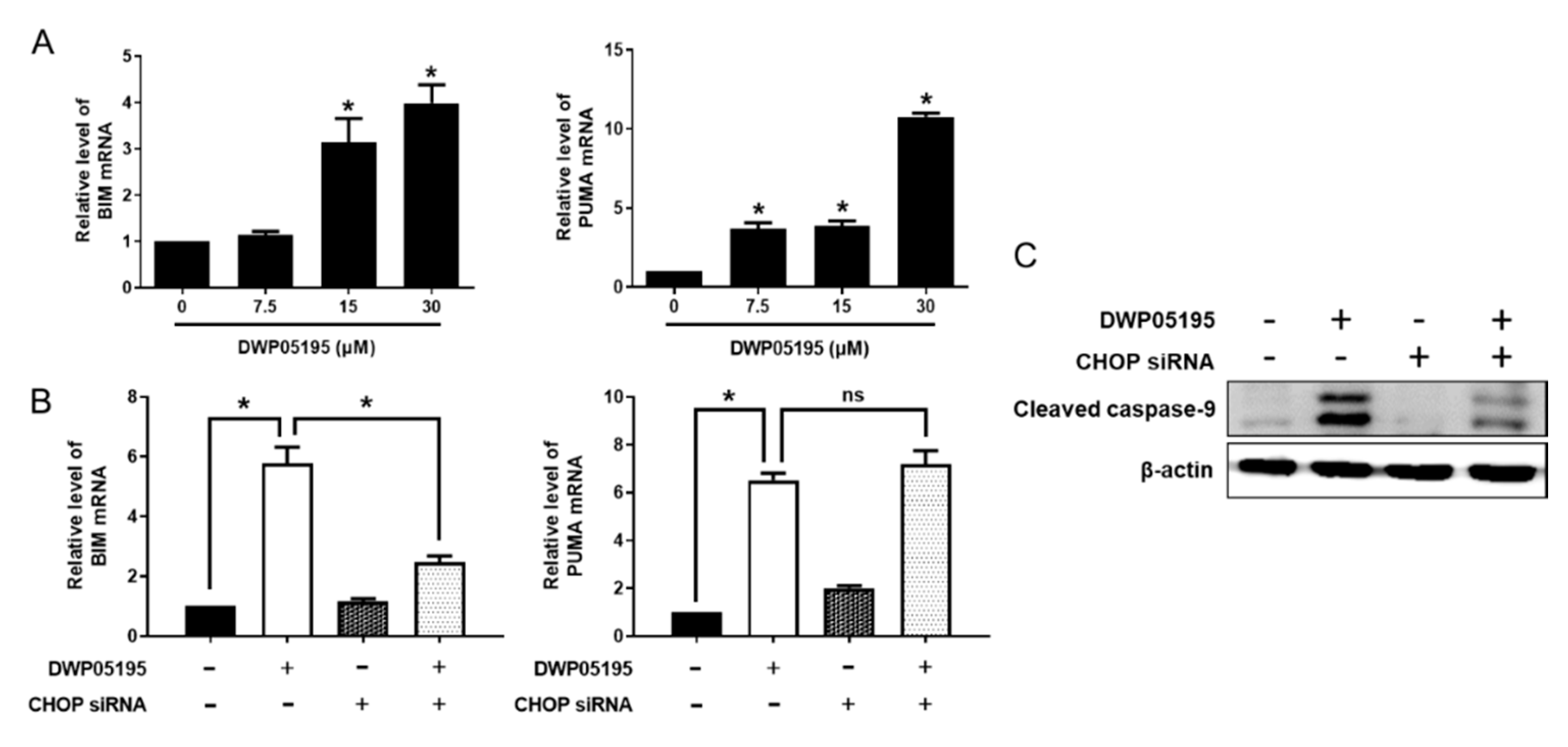
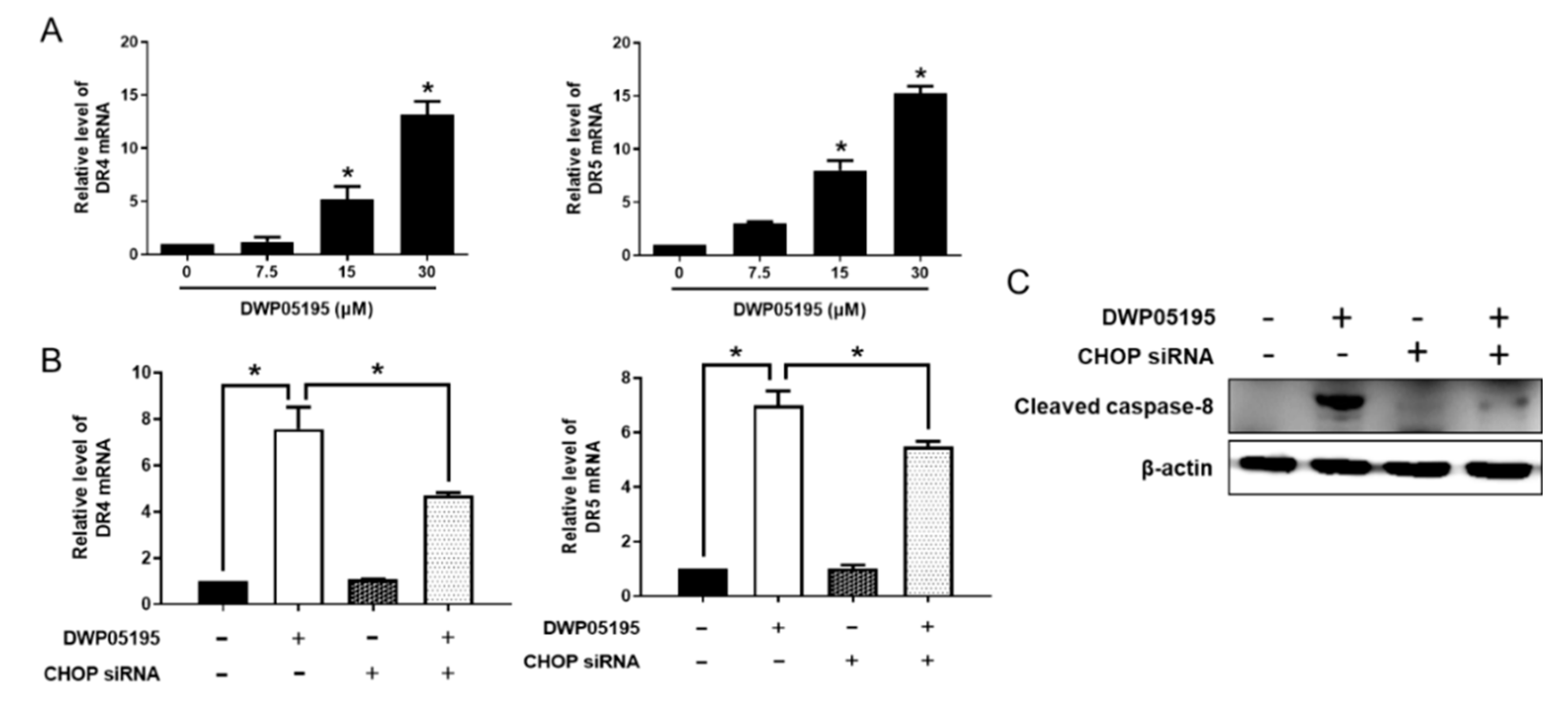
2.3. CHOP Upregulation by DWP05195 Stimulates Both Intrinsic and Extrinsic Apoptosis Pathways
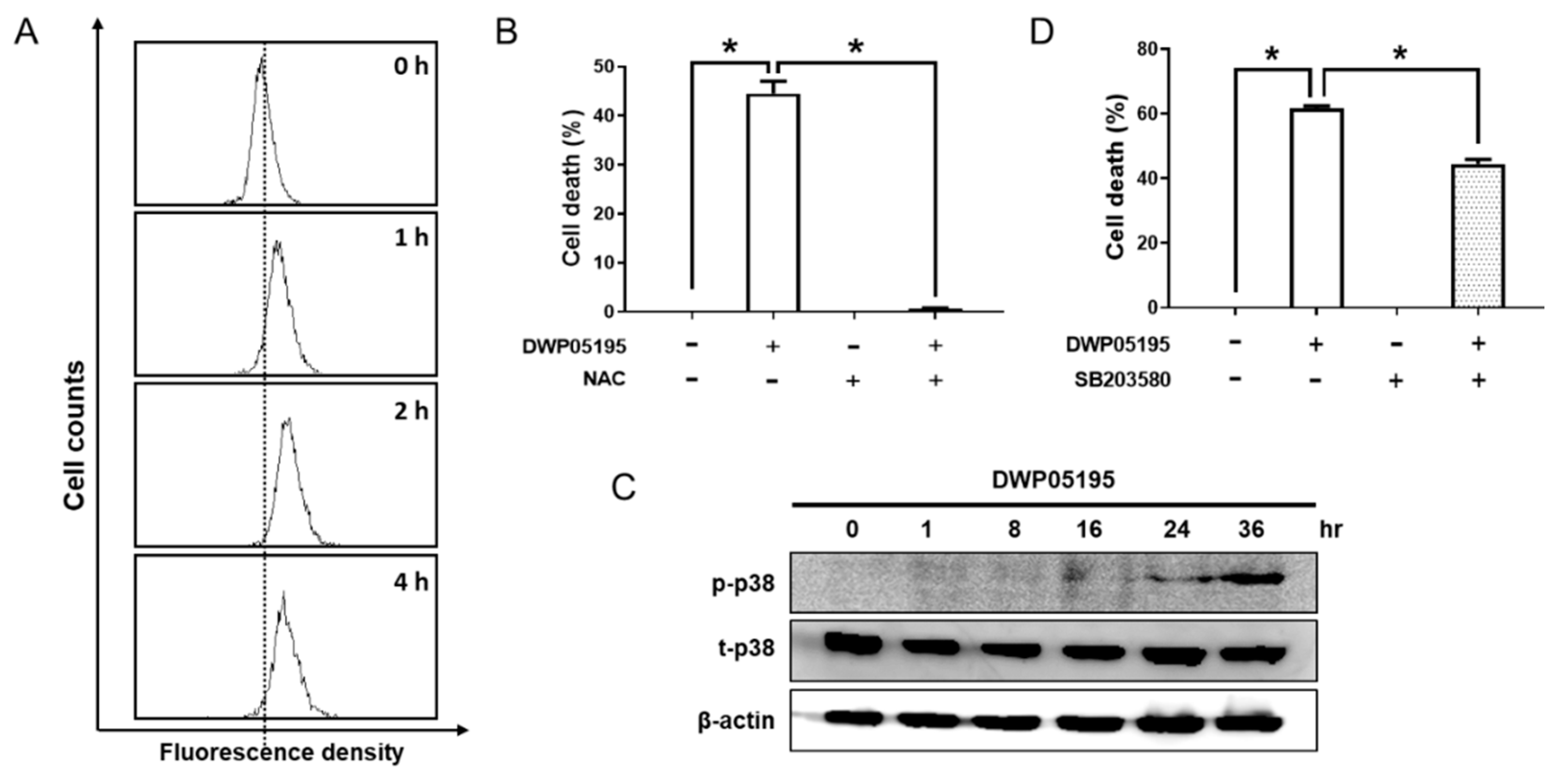
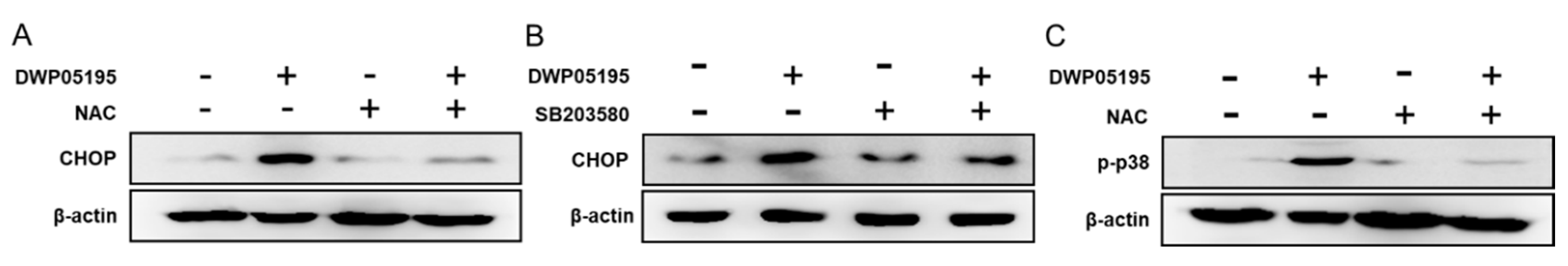
2.4. DWP05195 Induces ER Stress-Dependent Apoptosis by ROS Upregulation and p38 Activation
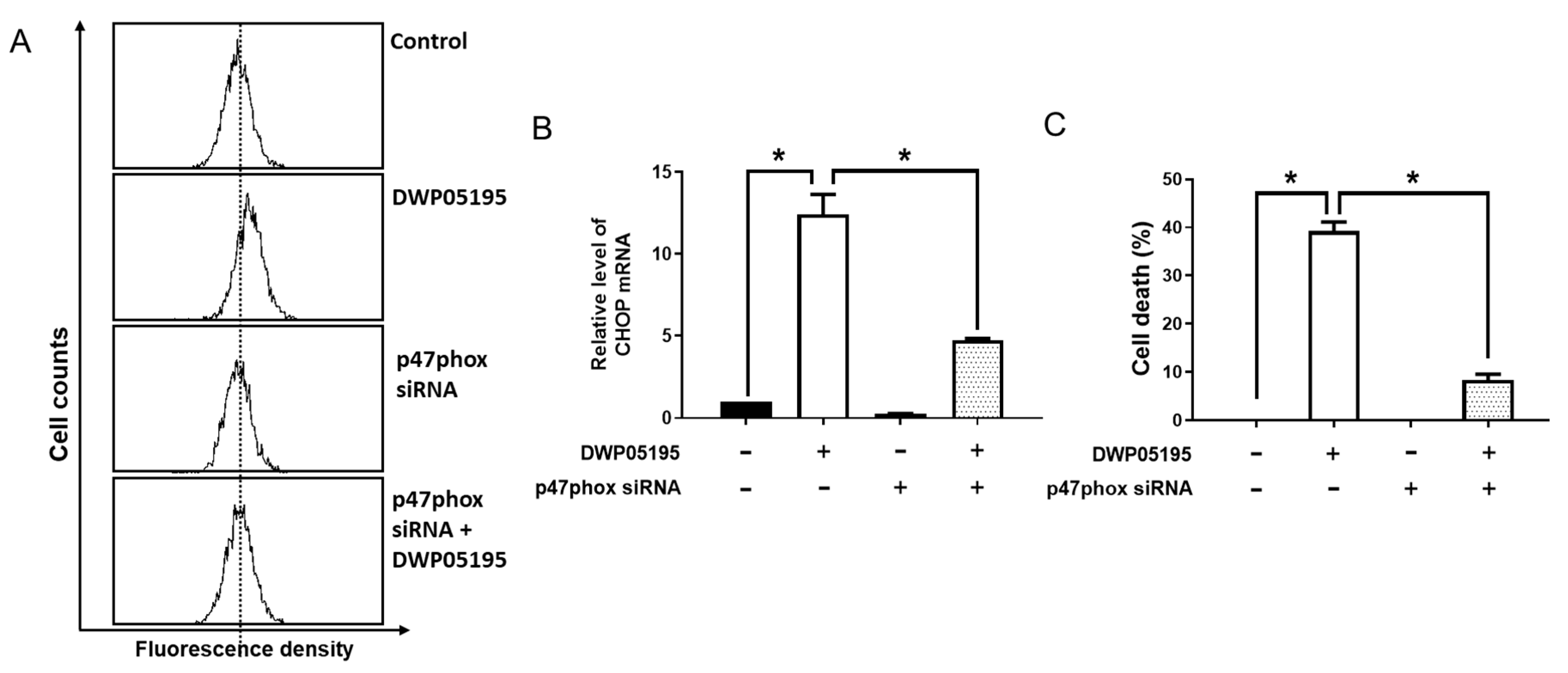
2.5. DWP05195 Regulates the Intracellular ROS Levels Through NADPH Oxidase
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Materials
4.2. MTT Assay
4.3. Annexin V and Propidium Iodide (PI) Double Staining for Apoptosis Analysis
4.4. Western Blot Analysis
4.5. Determination of Endoplasmic Reticulum Stress
4.6. Detction of Intracellular Reactive Oxygen Species
4.7. RNA Inteference for Gene Knockdown
4.8. Real-Time Reversed Transcription-PCR
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsh, S. Pharmacogenomics of taxane/platinum therapy in ovarian cancer. Int. J. Gynecol. Cancer 2009, 19, S30–S34. [Google Scholar] [CrossRef] [PubMed]
- Shapira, I.; Oswald, M.; Lovecchio, J.; Khalili, H.; Menzin, A.; Whyte, J.; Dos Santos, L.; Liang, S.; Bhuiya, T.; Keogh, M.; et al. Circulating biomarkers for detection of ovarian cancer and predicting cancer outcomes. Br. J. Cancer 2014, 110, 976–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jelovac, D.; Armstrong, D.K. Recent progress in the diagnosis and treatment of ovarian cancer. CA Cancer J. Clin. 2011, 61, 183–203. [Google Scholar] [CrossRef]
- Bhatt, P.; Vhora, I.; Patil, S.; Amrutiya, J.; Bhattacharya, C.; Misra, A.; Mashru, R. Role of antibodies in diagnosis and treatment of ovarian cancer: Basic approach and clinical status. J. Control. Release 2016, 226, 148–167. [Google Scholar] [CrossRef]
- Szallasi, A.; Cortright, D.N.; Blum, C.A.; Eid, S.R. The vanilloid receptor trpv1: 10 years from channel cloning to antagonist proof-of-concept. Nat. Rev. Drug Discov. 2007, 6, 357–372. [Google Scholar] [CrossRef]
- Brito, R.; Sheth, S.; Mukherjea, D.; Rybak, L.P.; Ramkumar, V. Trpv1: A potential drug target for treating various diseases. Cells 2014, 3, 517–545. [Google Scholar] [CrossRef] [Green Version]
- De La Chapa, J.J.; Singha, P.K.; Self, K.K.; Sallaway, M.L.; McHardy, S.F.; Hart, M.J.; McGuff, H.S.; Valdez, M.C.; Ruiz II, F.; Polusani, S.R.; et al. The novel capsazepine analog, cidd-99, significantly inhibits oral squamous cell carcinoma in vivo through a trpv1-independent induction of er stress, mitochondrial dysfunction, and apoptosis. J. Oral Pathol. Med. 2019, 48, 389–399. [Google Scholar] [CrossRef]
- Yang, M.H.; Jung, S.H.; Sethi, G.; Ahn, K.S. Pleiotropic pharmacological actions of capsazepine, a synthetic analogue of capsaicin, against various cancers and inflammatory diseases. Molecules 2019, 24, 995. [Google Scholar] [CrossRef] [Green Version]
- Sung, B.; Prasad, S.; Ravindran, J.; Yadav, V.R.; Aggarwal, B.B. Capsazepine, a trpv1 antagonist, sensitizes colorectal cancer cells to apoptosis by trail through ros-jnk-chop-mediated upregulation of death receptors. Free Radic. Biol. Med. 2012, 53, 1977–1987. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Kim, B.H.; Yu, K.S.; Kim, H.S.; Kim, J.D.; Cho, J.Y.; Lee, S.; Gu, N. A first-in-human, double-blind, placebo-controlled, randomized, dose escalation study of dwp05195, a novel trpv1 antagonist, in healthy volunteers. Drug Des. Devel. Ther. 2017, 11, 1301–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Liu, L.; Naik, I.; Braunstein, Z.; Zhong, J.; Ren, B. Transcription factor c/ebp homologous protein in health and diseases. Front. Immunol. 2017, 8, 1612. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Tian, M.; Ding, C.; Yu, S. The c/ebp homologous protein (chop) transcription factor functions in endoplasmic reticulum stress-induced apoptosis and microbial infection. Front. Immunol. 2019, 9, 3083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bujak, J.K.; Kosmala, D.; Szopa, I.M.; Majchrzak, K.; Bednarczyk, P. Inflammation, cancer and immunity-implication of trpv1 channel. Front. Oncol. 2019, 9, 1087. [Google Scholar] [CrossRef] [PubMed]
- Liou, G.Y.; Storz, P. Reactive oxygen species in cancer. Free Radic. Res. 2010, 44, 479–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial ros-induced ros release: An update and review. Biochim. Biophys. Acta. Bioenerg. 2006, 1757, 509–517. [Google Scholar] [CrossRef] [Green Version]
- Magnani, F.; Mattevi, A. Structure and mechanisms of ros generation by nadph oxidases. Curr. Opin. Struct. Biol. 2019, 59, 91–97. [Google Scholar] [CrossRef]
- Mukherjea, D.; Jajoo, S.; Sheehan, K.; Kaur, T.; Sheth, S.; Bunch, J.; Perro, C.; Rybak, L.P.; Ramkumar, V. Nox3 nadph oxidase couples transient receptor potential vanilloid 1 to signal transducer and activator of transcription 1-mediated inflammation and hearing loss. Antioxid. Redox. Sign. 2011, 14, 999–1010. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.S.; Lee, S.H.; Huang, H.S.; Chen, Y.S.; Ma, M.C. H2o2 generated by nadph oxidase 4 contributes to transient receptor potential vanilloid 1 channel-mediated mechanosensation in the rat kidney. Am. J. Physiol. Ren. Physiol. 2015, 309, F369–F376. [Google Scholar] [CrossRef] [Green Version]
- Taylor-Clark, T.E. Role of reactive oxygen species and trp channels in the cough reflex. Cell Calcium 2016, 60, 155–162. [Google Scholar] [CrossRef] [Green Version]
- Domotor, A.; Peidl, Z.; Vincze, A.; Hunyady, B.; Szolcsanyi, J.; Kereskay, L.; Szekeres, G.; Mozsik, G. Immunohistochemical distribution of vanilloid receptor, calcitonin-gene related peptide and substance p in gastrointestinal mucosa of patients with different gastrointestinal disorders. Inflammopharmacology 2005, 13, 161–177. [Google Scholar] [CrossRef] [PubMed]
- Lazzeri, M.; Vannucchi, M.G.; Spinelli, M.; Bizzoco, E.; Beneforti, P.; Turini, D.; Faussone-Pellegrini, M.S. Transient receptor potential vanilloid type 1 (trpv1) expression changes from normal urothelium to transitional cell carcinoma of human bladder. Eur. Urol. 2005, 48, 691–698. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, M.G.; Sanchez, A.M.; Collado, B.; Malagarie-Cazenave, S.; Olea, N.; Carmena, M.J.; Prieto, J.C.; Diaz-Laviada, I. Expression of the transient receptor potential vanilloid 1 (trpv1) in lncap and pc-3 prostate cancer cells and in human prostate tissue. Eur. J. Pharmacol. 2005, 515, 20–27. [Google Scholar] [CrossRef]
- Nilius, B. Trp channels in disease. Biochim. Biophys. Acta Mol. Basis Dis. 2007, 1772, 805–812. [Google Scholar] [CrossRef] [Green Version]
- Czifra, G.; Varga, A.; Nyeste, K.; Marincsak, R.; Toth, B.I.; Kovacs, I.; Kovacs, L.; Biro, T. Increased expressions of cannabinoid receptor-1 and transient receptor potential vanilloid-1 in human prostate carcinoma. J. Cancer Res. Clin. Oncol. 2009, 135, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Santoni, G.; Caprodossi, S.; Farfariello, V.; Liberati, S.; Gismondi, A.; Amantini, C. Antioncogenic effects of transient receptor potential vanilloid 1 in the progression of transitional urothelial cancer of human bladder. ISRN Urol. 2012, 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozano, C.; Cordova, C.; Marchant, I.; Zuniga, R.; Ochova, P.; Ramirez-Barrantes, R.; Gonzalez-Arriagada, W.A.; Rodriguez, B.; Olivero, P. Intracellular aggregated trpv1 is associated with lower survival in breast cancer patients. Breast Cancer (Dove Med Press) 2018, 10, 161–168. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.C.; Hsieh, P.F.; Hsieh, M.K.; Zeng, Z.M.; Cheng, H.L.; Liao, J.W.; Chueh, P.J. Capsaicin-mediated tnox (enox2) up-regulation enhances cell proliferation and migration in vitro and in vivo. J. Agric. Food Chem. 2012, 60, 2758–2765. [Google Scholar] [CrossRef]
- Zhang, R.; Humphreys, I.; Sahu, R.P.; Shi, Y.; Srivastava, S.K. In vitro and in vivo induction of apoptosis by capsaicin in pancreatic cancer cells is mediated through ros generation and mitochondrial death pathway. Apoptosis 2008, 13, 1465–1478. [Google Scholar] [CrossRef]
- Amantini, C.; Ballarini, P.; Caprodossi, S.; Nabissi, M.; Morelli, M.B.; Lucciarini, R.; Cardarelli, M.A.; Mammana, G.; Santoni, G. Triggering of transient receptor potential vanilloid type 1 (trpv1) by capsaicin induces fas/cd95-mediated apoptosis of urothelial cancer cells in an atm-dependent manner. Carcinogenesis 2009, 30, 1320–1329. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Wang, G.; Tao, H.; Yang, Z.; Wang, Y.; Meng, Z.; Cao, R.; Xiao, Y.; Wang, X.; Zhou, J. Capsaicin mediates caspases activation and induces apoptosis through p38 and jnk mapk pathways in human renal carcinoma. BMC Cancer 2016, 16, 790. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Zhang, L.; Cheng, X.; Yu, H.; Bao, J.; Lu, R. Capsaicin inhibits the metastasis of human papillary thyroid carcinoma bcpap cells through the modulation of the trpv1 channel. Food Funct. 2018, 9, 344–354. [Google Scholar] [CrossRef]
- Chien, C.S.; Ma, K.H.; Lee, H.S.; Liu, P.S.; Li, Y.H.; Huang, Y.S.; Chueh, S.H. Dual effect of capsaicin on cell death in human osteosarcoma g292 cells. Eur. J. Pharmacol. 2013, 718, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Amantini, C.; Mosca, M.; Nabissi, M.; Lucciarini, R.; Caprodossi, S.; Arcella, A.; Giangaspero, F.; Santoni, G. Capsaicin-induced apoptosis of glioma cells is mediated by trpv1 vanilloid receptor and requires p38 mapk activation. J. Neurochem. 2007, 102, 977–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, L.; Zhuang, Y.X.; Zhang, H.W.; Tian, N.N.; Dang, W.Z.; Wu, S.Y. Capsaicin-loaded folic acid-conjugated lipid nanoparticles for enhanced therapeutic efficacy in ovarian cancers. Biomed. Pharmacother. 2017, 91, 999–1005. [Google Scholar] [CrossRef] [PubMed]
- Hartel, M.; Di Mola, F.F.; Selvaggi, F.; Mascetta, G.; Wente, M.N.; Felix, K.; Giese, N.A.; Hinz, U.; Di Sebastiano, P.; Buchler, M.W.; et al. Vanilloids in pancreatic cancer: Potential for chemotherapy and pain management. Gut 2006, 55, 519–528. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Guo, W.; Ma, J.; Xu, P.; Zhang, W.; Guo, S.; Liu, L.; Ma, J.; Shi, Q.; Jian, Z.; et al. Downregulated trpv1 expression contributes to melanoma growth via the calcineurin-atf3-p53 pathway. J. Invest. Dermatol. 2018, 138, 2205–2215. [Google Scholar] [CrossRef] [Green Version]
- Teng, H.P.; Huang, C.J.; Yeh, J.H.; Hsu, S.S.; Lo, Y.K.; Cheng, J.S.; Cheng, H.H.; Chen, J.S.; Jiann, B.P.; Chang, H.T.; et al. Capsazepine elevates intracellular ca2+ in human osteosarcoma cells, questioning its selectivity as a vanilloid receptor antagonist. Life Sci. 2004, 75, 2515–2526. [Google Scholar] [CrossRef]
- Huang, J.K.; Cheng, H.H.; Huang, C.J.; Kuo, C.C.; Chen, W.C.; Liu, S.I.; Hsu, S.S.; Chang, H.T.; Lu, Y.C.; Tseng, L.L.; et al. Effect of capsazepine on cytosolic ca(2+) levels and proliferation of human prostate cancer cells. Toxicol In Vitro 2006, 20, 567–574. [Google Scholar] [CrossRef]
- Gonzales, C.B.; Kirma, N.B.; De La Chapa, J.J.; Chen, R.; Henry, M.A.; Luo, S.; Hargreaves, K.M. Vanilloids induce oral cancer apoptosis independent of trpv1. Oral Oncol. 2014, 50, 437–447. [Google Scholar] [CrossRef] [Green Version]
- Cao, S.S.; Kaufman, R.J. Endoplasmic reticulum stress and oxidative stress in cell fate decision and human disease. Antioxid. Redox. Sign. 2014, 21, 396–413. [Google Scholar] [CrossRef] [PubMed]
- Yadav, R.K.; Chae, S.W.; Kim, H.R.; Chae, H.J. Endoplasmic reticulum stress and cancer. J. Cancer Prev. 2014, 19, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Tabas, I.; Ron, D. Integrating the mechanisms of apoptosis induced by endoplasmic reticulum stress. Nat. Cell Biol. 2011, 13, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Gardner, B.M.; Pincus, D.; Gotthardt, K.; Gallagher, C.M.; Walter, P. Endoplasmic reticulum stress sensing in the unfolded protein response. Cold Spring Harb. Perspect. Biol. 2013, 5. [Google Scholar] [CrossRef] [Green Version]
- Iurlaro, R.; Munoz-Pinedo, C. Cell death induced by endoplasmic reticulum stress. FEBS J. 2016, 283, 2640–2652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Indran, I.R.; Tufo, G.; Pervaiz, S.; Brenner, C. Recent advances in apoptosis, mitochondria and drug resistance in cancer cells. Biochim. Biophys. Acta. Bioenerg. 2011, 1807, 735–745. [Google Scholar] [CrossRef] [Green Version]
- Puthalakath, H.; O’Reilly, L.A.; Gunn, P.; Lee, L.; Kelly, P.N.; Huntington, N.D.; Hughes, P.D.; Michalak, E.M.; McKimm-Breschkin, J.; Motoyama, N.; et al. Er stress triggers apoptosis by activating bh3-only protein bim. Cell 2007, 129, 1337–1349. [Google Scholar] [CrossRef] [Green Version]
- Miller, W.H., Jr.; Schipper, H.M.; Lee, J.S.; Singer, J.; Waxman, S. Mechanisms of action of arsenic trioxide. Cancer Res. 2002, 62, 3893–3903. [Google Scholar]
- Parkinson, E.I.; Hergenrother, P.J. Runaway ros as a selective anticancer strategy. ChemMedChem 2011, 6, 1957–1959. [Google Scholar] [CrossRef]
- Taha, M.M.E.; Sheikh, B.Y.; Salim, L.Z.A.; Mohan, S.; Khan, A.; Kamalidehghan, B.; Ahmadipour, F.; Abdelwahab, S.I. Thymoquinone induces apoptosis and increase ros in ovarian cancer cell line. Cell. Mol. Biol. 2016, 62, 97–101. [Google Scholar]
- Li, Z.; Huang, L.; Wei, L.; Hou, Z.; Ye, W.; Huang, S. Chaetocin induces caspase-dependent apoptosis in ovarian cancer cells via the generation of reactive oxygen species. Oncol. Lett. 2019, 18, 1915–1921. [Google Scholar] [CrossRef] [PubMed]
- Juhasz, A.; Ge, Y.; Markel, S.; Chiu, A.; Matsumoto, L.; van Balgooy, J.; Roy, K.; Doroshow, J.H. Expression of nadph oxidase homologues and accessory genes in human cancer cell lines, tumours and adjacent normal tissues. Free Radic. Res. 2009, 43, 523–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, E.C.; Jiang, F.; Peshavariya, H.M.; Dusting, G.J. Regulation of cell proliferation by nadph oxidase-mediated signaling: Potential roles in tissue repair, regenerative medicine and tissue engineering. Pharmacol. Therapeut. 2009, 122, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Cuenca, I.; Roncero, C.; Sancho, P.; Caja, L.; Fausto, N.; Fernandez, M.; Fabregat, I. Upregulation of the nadph oxidase nox4 by tgf-beta in hepatocytes is required for its pro-apoptotic activity. J. Hepatol. 2008, 49, 965–976. [Google Scholar] [CrossRef] [PubMed]
- Morre, D.J.; Chueh, P.J.; Morre, D.M. Capsaicin inhibits preferentially the nadh oxidase and growth of transformed cells in culture. Proc. Natl. Acad. Sci. USA 1995, 92, 1831–1835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, M.; Forman, H.J. Redox signaling and the map kinase pathways. Biofactors 2003, 17, 287–296. [Google Scholar] [CrossRef]
- Dhillon, A.S.; Hagan, S.; Rath, O.; Kolch, W. Map kinase signalling pathways in cancer. Oncogene 2007, 26, 3279–3290. [Google Scholar] [CrossRef] [Green Version]
- Haagenson, K.K.; Wu, G.S. Mitogen activated protein kinase phosphatases and cancer. Cancer Biol. Ther. 2010, 9, 337–340. [Google Scholar] [CrossRef] [Green Version]
- Marshall, C.J. Specificity of receptor tyrosine kinase signaling: Transient versus sustained extracellular signal-regulated kinase activation. Cell 1995, 80, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Kyriakis, J.M.; Avruch, J. Protein kinase cascades activated by stress and inflammatory cytokines. Bioessays 1996, 18, 567–577. [Google Scholar] [CrossRef]
- Hill, C.S.; Treisman, R. Transcriptional regulation by extracellular signals: Mechanisms and specificity. Cell 1995, 80, 199–211. [Google Scholar] [CrossRef] [Green Version]
- Xia, Z.; Dickens, M.; Raingeaud, J.; Davis, R.J.; Greenberg, M.E. Opposing effects of erk and jnk-p38 map kinases on apoptosis. Science 1995, 270, 1326–1331. [Google Scholar] [CrossRef] [PubMed]
- Johnson, N.L.; Gardner, A.M.; Diener, K.M.; Lange-Carter, C.A.; Gleavy, J.; Jarpe, M.B.; Minden, A.; Karin, M.; Zon, L.I.; Johnson, G.L. Signal transduction pathways regulated by mitogen-activated/extracellular response kinase kinase kinase induce cell death. J. Biol. Chem. 1996, 271, 3229–3237. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Lines | A2780 | SKOV3 | OVCAR3 | TOV-21G | Hey8A |
|---|---|---|---|---|---|
| aIC50 (μM) | 17.61 ± 1.12 | 43.87 ± 5.59 | 78.86 ± 27.05 | 35.92 ± 7.34 | 40.83 ± 4.02 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.-Y.; Lee, K.-T.; Lim, M.C.; Choi, J.-H. TRPV1 Antagonist DWP05195 Induces ER Stress-Dependent Apoptosis through the ROS-p38-CHOP Pathway in Human Ovarian Cancer Cells. Cancers 2020, 12, 1702. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12061702
Wang Y-Y, Lee K-T, Lim MC, Choi J-H. TRPV1 Antagonist DWP05195 Induces ER Stress-Dependent Apoptosis through the ROS-p38-CHOP Pathway in Human Ovarian Cancer Cells. Cancers. 2020; 12(6):1702. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12061702
Chicago/Turabian StyleWang, Yi-Yue, Kyung-Tae Lee, Myong Cheol Lim, and Jung-Hye Choi. 2020. "TRPV1 Antagonist DWP05195 Induces ER Stress-Dependent Apoptosis through the ROS-p38-CHOP Pathway in Human Ovarian Cancer Cells" Cancers 12, no. 6: 1702. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12061702