
Metabolic Changes in Early-Stage Non–Small Cell Lung Cancer Patients after Surgical Resection

 , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patient Enrollment and Biofluid Sample Collection
2.2. Nuclear Magnetic Resonance (NMR)
2.2.1. Sample Preparation for NMR Analysis
2.2.2. NMR Analysis
2.3. Liquid Chromatography Quadrupole Time-of-Flight Mass Spectrometry (LC-QTOF-MS)
2.3.1. Sample Preparation for LC-QTOF-MS
2.3.2. LC-QTOF-MS Analysis
2.4. Statistical Analysis and Metabolite Identification
3. Results
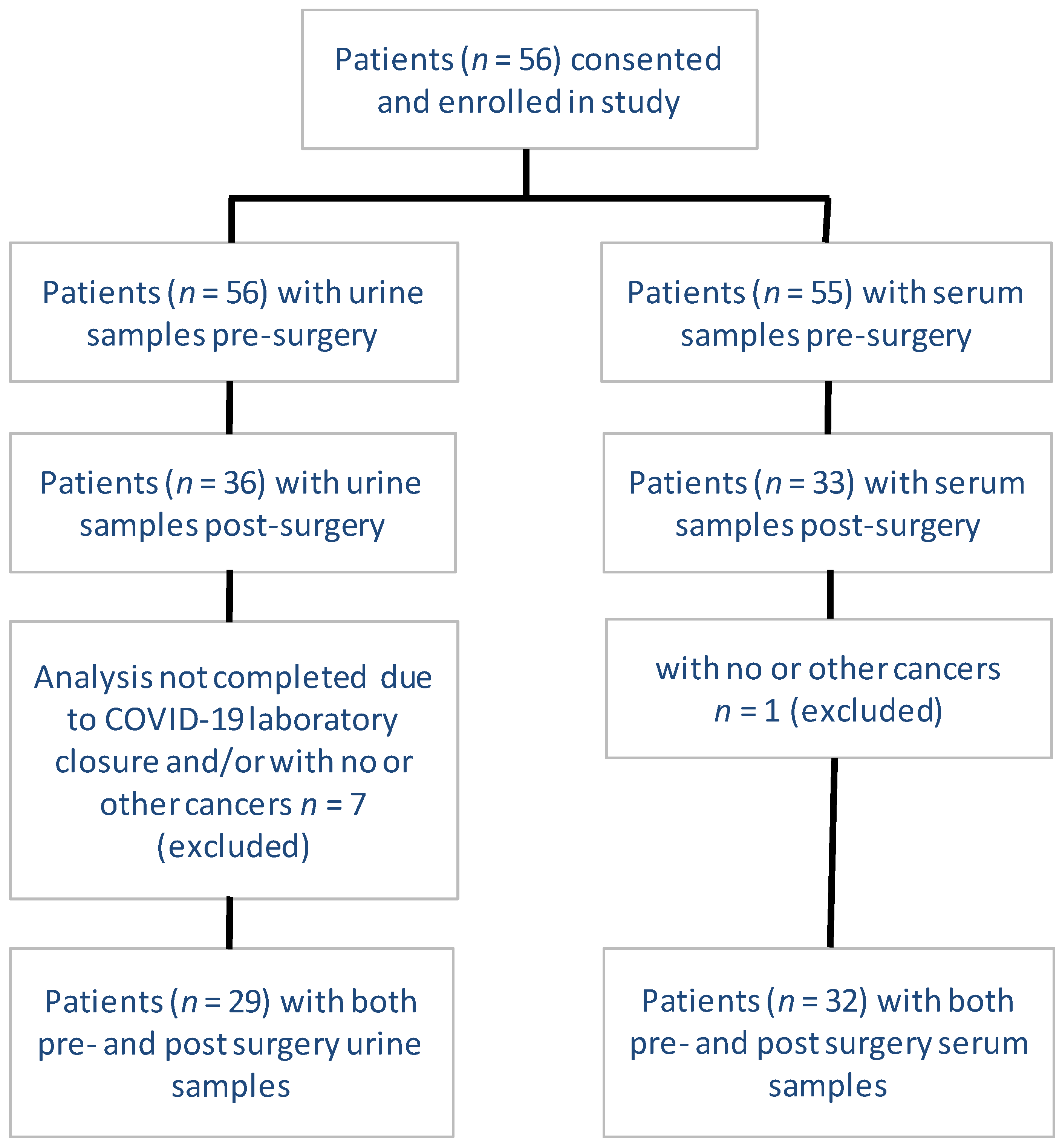
3.1. Patient Enrollment, Inclusion, and Exclusion
3.2. Inclusion, Exclusion, and Classification of Identified Metabolites
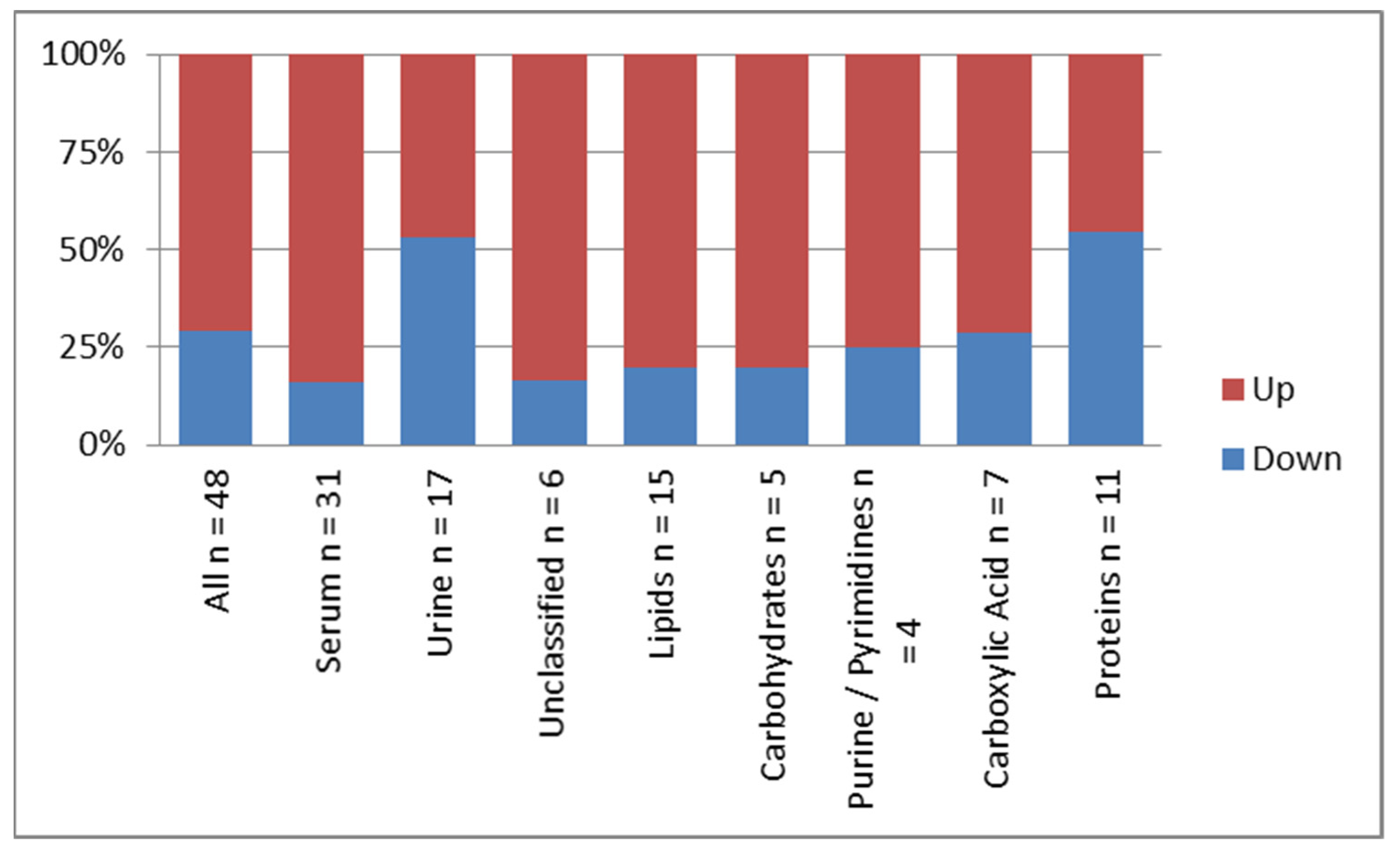
3.3. Pattern of Frequency and Distribution of the Identified Metabolites
3.4. Pattern of Dysregulation of the Identified Metabolites Post-Surgery
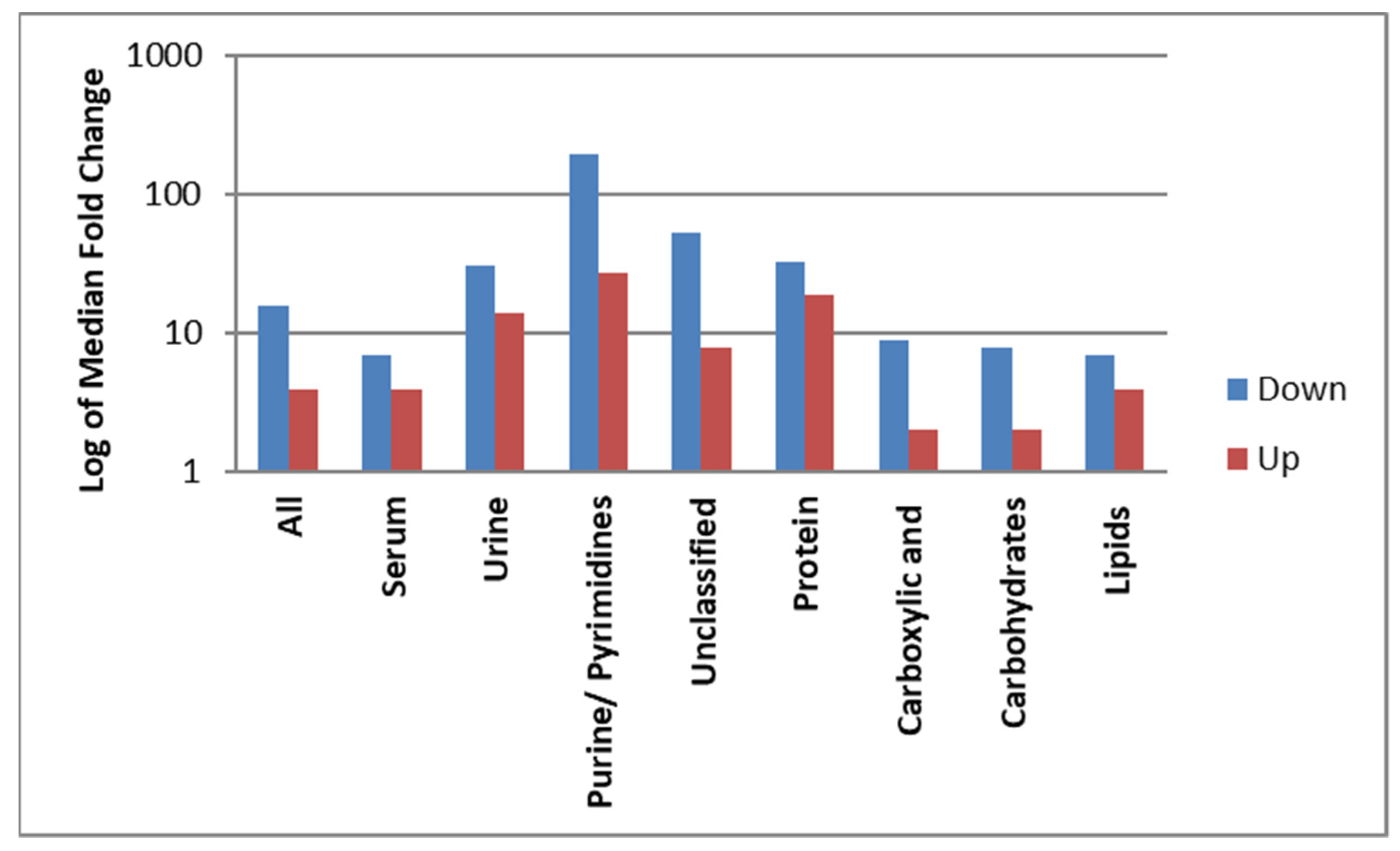
3.5. Magnitude of Dysregulation of the Identified Metabolites Post-Surgery
3.6. Metabolites of Significance with >10 FC Dysregulation and Their Association with Pathogenesis of Cancer
4. Discussion
4.1. Study Desgin and Metobolic Profile
4.2. Lipids and Derivatives
4.3. Proteins and Derivatives
4.4. Purines and Pyrimidines
4.5. Carboxylic Acid Derivatives and Carbohydrates
4.6. Endocrine Factors
4.7. Strengths and Limitations of the Study
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schvartsman, G.; Ferrarotto, R.; Massarelli, E. Checkpoint inhibitors in lung cancer: Latest developments and clinical potential. Ther. Adv. Med. Oncol. 2016, 8, 460–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barlesi, F.; Mazieres, J.; Merlio, J.-P.; Debieuvre, D.; Mosser, J.; Lena, H.; Ouafik, L.H.; Besse, B.; Rouquette, I.; Westeel, V. Routine molecular profiling of patients with advanced non-small-cell lung cancer: Results of a 1-year nationwide programme of the French Cooperative Thoracic Intergroup (IFCT). Lancet 2016, 387, 1415–1426. [Google Scholar] [CrossRef]
- Rosell, R.; Karachaliou, N. Large-scale screening for somatic mutations in lung cancer. Lancet 2016, 387, 1354–1356. [Google Scholar] [CrossRef]
- Kris, M.G.; Johnson, B.E.; Berry, L.D.; Kwiatkowski, D.J.; Iafrate, A.J.; Wistuba, I.I.; Varella-Garcia, M.; Franklin, W.A.; Aronson, S.L.; Su, P.-F. Using multiplexed assays of oncogenic drivers in lung cancers to select targeted drugs. JAMA J. Am. Med. Assoc. 2014, 311, 1998–2006. [Google Scholar] [CrossRef] [PubMed]
- Warburg, O. On the origin of cancer cells. Science 1956, 123, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg effect: The metabolic requirements of cell proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bamji-Stocke, S.; van Berkel, V.; Miller, D.M.; Frieboes, H.B. A review of metabolism-associated biomarkers in lung cancer diagnosis and treatment. Metabolomics 2018, 14, 81. [Google Scholar] [CrossRef]
- Yu, L.; Li, K.; Zhang, X. Next-generation metabolomics in lung cancer diagnosis, treatment and precision medicine: Mini review. Oncotarget 2017, 8, 115774–115786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seijo, L.M.; Peled, N.; Ajona, D.; Boeri, M.; Field, J.K.; Sozzi, G.; Pio, R.; Zulueta, J.J.; Spira, A.; Massion, P.P.; et al. Biomarkers in Lung Cancer Screening: Achievements, Promises, and Challenges. J. Thorac. Oncol. 2019, 14, 343–357. [Google Scholar] [CrossRef] [Green Version]
- Mathe, E.A.; Patterson, A.D.; Haznadar, M.; Manna, S.K.; Krausz, K.W.; Bowman, E.D.; Shields, P.G.; Idle, J.R.; Smith, P.B.; Anami, K.; et al. Noninvasive urinary metabolomic profiling identifies diagnostic and prognostic markers in lung cancer. Cancer Res. 2014, 74, 3259–3270. [Google Scholar] [CrossRef] [Green Version]
- Haznadar, M.; Cai, Q.; Krausz, K.W.; Bowman, E.D.; Margono, E.; Noro, R.; Thompson, M.D.; Mathe, E.A.; Munro, H.M.; Steinwandel, M.D.; et al. Urinary Metabolite Risk Biomarkers of Lung Cancer: A Prospective Cohort Study. Cancer Epidemiol. Prev. Biomark. 2016, 25, 978–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ros-Mazurczyk, M.; Wojakowska, A.; Marczak, L.; Polanski, K.; Pietrowska, M.; Polanska, J.; Dziadziuszko, R.; Jassem, J.; Rzyman, W.; Widlak, P. Panel of serum metabolites discriminates cancer patients and healthy participants of lung cancer screening—A pilot study. Acta Biochim. Pol. 2017, 64, 513–518. [Google Scholar] [CrossRef]
- Fahrmann, J.F.; Grapov, D.; DeFelice, B.C.; Taylor, S.; Kim, K.; Kelly, K.; Wikoff, W.R.; Pass, H.; Rom, W.N.; Fiehn, O.; et al. Serum phosphatidylethanolamine levels distinguish benign from malignant solitary pulmonary nodules and represent a potential diagnostic biomarker for lung cancer. Cancer Biomark. 2016, 16, 609–617. [Google Scholar] [CrossRef] [Green Version]
- Cameron, S.J.; Lewis, K.E.; Beckmann, M.; Allison, G.G.; Ghosal, R.; Lewis, P.D.; Mur, L.A. The metabolomic detection of lung cancer biomarkers in sputum. Lung Cancer 2016, 94, 88–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peralbo-Molina, A.; Calderon-Santiago, M.; Priego-Capote, F.; Jurado-Gamez, B.; Luque de Castro, M.D. Identification of metabolomics panels for potential lung cancer screening by analysis of exhaled breath condensate. J. Breath Res. 2016, 10, 026002. [Google Scholar] [CrossRef]
- Ahmed, N.; Bezabeh, T.; Ijare, O.B.; Myers, R.; Alomran, R.; Aliani, M.; Nugent, Z.; Banerji, S.; Kim, J.; Qing, G. Metabolic Signatures of Lung Cancer in Sputum and Exhaled Breath Condensate Detected by 1H Magnetic Resonance Spectroscopy: A Feasibility Study. Magn. Reson. Insights 2016, 9, 29. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Li, Z.; Lazar, L.; Fang, Z.; Tang, C.; Zhao, J. Metabolomics workflow for lung cancer: Discovery of biomarkers. Clin. Chim. Acta 2019, 495, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Tee, S.-S.; Keshari, K.R. Novel approaches to imaging tumor metabolism. Cancer J. 2015, 21, 165. [Google Scholar] [CrossRef] [Green Version]
- García-Figueiras, R.; Baleato-González, S.; Padhani, A.R.; Oleaga, L.; Vilanova, J.C.; Luna, A.; Gómez, J.C.C. Proton magnetic resonance spectroscopy in oncology: The fingerprints of cancer? Diagn. Interv. Radiol. 2016, 22, 75. [Google Scholar] [CrossRef]
- Ahmed, N.; Kidane, B.; Wang, L.; Qing, G.; Tan, L.; Buduhan, G.; Srinathan, S.; Aliani, M. Non-invasive exploration of metabolic profile of lung cancer with Magnetic Resonance Spectroscopy and Mass Spectrometry. Contemp. Clin. Trials Commun. 2019, 16, 100445. [Google Scholar] [CrossRef] [PubMed]
- Xi, B.; Gu, H.; Baniasadi, H.; Raftery, D. Statistical analysis and modeling of mass spectrometry-based metabolomics data. In Mass Spectrometry in Metabolomics; Springer: New York, NY, USA, 2014; pp. 333–353. [Google Scholar]
- Smith, C.A.; O’Maille, G.; Want, E.J.; Qin, C.; Trauger, S.A.; Brandon, T.R.; Custodio, D.E.; Abagyan, R.; Siuzdak, G. METLIN: A metabolite mass spectral database. Ther. Drug Monit. 2005, 27, 747–751. [Google Scholar] [CrossRef]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vázquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N. HMDB 4.0: The human metabolome database for 2018. Nucleic Acids Res. 2018, 46, D608–D617. [Google Scholar] [CrossRef]
- Sayers, E.W.; Barrett, T.; Benson, D.A.; Bolton, E.; Bryant, S.H.; Canese, K.; Chetvernin, V.; Church, D.M.; DiCuccio, M.; Federhen, S. Database resources of the national center for biotechnology information. Nucleic Acids Res. 2010, 39, D38–D51. [Google Scholar] [CrossRef] [Green Version]
- Hao, Y.; Li, D.; Xu, Y.; Ouyang, J.; Wang, Y.; Zhang, Y.; Li, B.; Xie, L.; Qin, G. Investigation of lipid metabolism dysregulation and the effects on immune microenvironments in pan-cancer using multiple omics data. BMC Bioinform. 2019, 20, 29–39. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Colby, J.K.; Zuo, X.; Jaoude, J.; Wei, D.; Shureiqi, I. The role of PPAR-δ in metabolism, inflammation, and cancer: Many characters of a critical transcription factor. Int. J. Mol. Sci. 2018, 19, 3339. [Google Scholar] [CrossRef] [Green Version]
- Röhrig, F.; Schulze, A. The multifaceted roles of fatty acid synthesis in cancer. Nat. Rev. Cancer 2016, 16, 732. [Google Scholar] [CrossRef] [PubMed]
- Salvador, M.M.; de Cedrón, M.G.; Rubio, J.M.; Martínez, S.F.; Martínez, R.S.; Casado, E.; de Molina, A.R.; Sereno, M. Lipid metabolism and lung cancer. Crit. Rev. Oncol. Hematol. 2017, 112, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Visca, P.; Sebastiani, V.; Botti, C.; Diodoro, M.G.; Lasagni, R.P.; Romagnoli, F.; Brenna, A.; De Joannon, B.C.; Donnorso, R.P.; Lombardi, G. Fatty acid synthase (FAS) is a marker of increased risk of recurrence in lung carcinoma. Anticancer Res. 2004, 24, 4169–4174. [Google Scholar] [PubMed]
- Wang, Y.; Zhang, X.; Tan, W.; Fu, J.; Zhang, W. Significance of fatty acid synthase expression in non-small cell lung cancer. Zhonghua Zhong Liu Za Zhi Chin. J. Oncol. 2002, 24, 271–273. [Google Scholar]
- Huang, J.; Fan, X.-X.; He, J.; Pan, H.; Li, R.-Z.; Huang, L.; Jiang, Z.; Yao, X.-J.; Liu, L.; Leung, E.L.-H. SCD1 is associated with tumor promotion, late stage and poor survival in lung adenocarcinoma. Oncotarget 2016, 7, 39970. [Google Scholar] [CrossRef] [Green Version]
- Yoh, K.; Ishii, G.; Yokose, T.; Minegishi, Y.; Tsuta, K.; Goto, K.; Nishiwaki, Y.; Kodama, T.; Suga, M.; Ochiai, A. Breast cancer resistance protein impacts clinical outcome in platinum-based chemotherapy for advanced non-small cell lung cancer. Clin. Cancer Res. 2004, 10, 1691–1697. [Google Scholar] [CrossRef] [Green Version]
- Butler, L.; Perone, Y.; Dehairs, J.; Lupien, L.E.; de Laat, V.; Talebi, A.; Loda, M.; Kinlaw, W.B.; Swinnen, J.V. Lipids and cancer: Emerging roles in pathogenesis, diagnosis and therapeutic intervention. Adv. Drug Deliv. Rev. 2020. [Google Scholar] [CrossRef]
- Thurnher, M.; Gruenbacher, G.; Nussbaumer, O. Regulation of mevalonate metabolism in cancer and immune cells. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2013, 1831, 1009–1015. [Google Scholar] [CrossRef]
- Fiala, O.; Pesek, M.; Finek, J.; Minarik, M.; Benesova, L.; Bortlicek, Z.; Topolcan, O. Statins augment efficacy of EGFR-TKIs in patients with advanced-stage non-small cell lung cancer harbouring KRAS mutation. Tumor Biol. 2015, 36, 5801–5805. [Google Scholar] [CrossRef] [PubMed]
- Göbel, A.; Rauner, M.; Hofbauer, L.C.; Rachner, T.D. Cholesterol and beyond-the role of the mevalonate pathway in cancer biology. Biochim. Biophys. Acta BBA Rev. Cancer 2020, 1873, 188351. [Google Scholar] [CrossRef] [PubMed]
- Hassanabad, A.F.; Mina, F. Targeting the Mevalonate Pathway for Treating Lung Cancer. Am. J. Clin. Oncol. 2020, 43, 69–70. [Google Scholar] [CrossRef] [PubMed]
- Debruyne, P.R.; Bruyneel, E.A.; Li, X.; Zimber, A.; Gespach, C.; Mareel, M.M. The role of bile acids in carcinogenesis. Mutat. Res. Fundam. Mol. Mech. Mutagenesis 2001, 480, 359–369. [Google Scholar] [CrossRef]
- Di Ciaula, A.; Wang, D.Q.-H.; Molina-Molina, E.; Baccetto, R.L.; Calamita, G.; Palmieri, V.O.; Portincasa, P. Bile acids and cancer: Direct and environmental-dependent effects. Ann. Hepatol. 2017, 16, S87–S105. [Google Scholar] [CrossRef]
- Muhammad, N.; Lee, H.M.; Kim, J. Oncology therapeutics targeting the metabolism of amino acids. Cells 2020, 9, 1904. [Google Scholar] [CrossRef]
- Vettore, L.; Westbrook, R.L.; Tennant, D.A. New aspects of amino acid metabolism in cancer. Br. J. Cancer 2020, 122, 150–156. [Google Scholar] [CrossRef]
- Lieu, E.L.; Nguyen, T.; Rhyne, S.; Kim, J. Amino acids in cancer. Exp. Mol. Med. 2020, 52, 15–30. [Google Scholar] [CrossRef]
- Phang, J.M. Proline metabolism in cell regulation and cancer biology: Recent advances and hypotheses. Antioxid. Redox Signal. 2019, 30, 635–649. [Google Scholar] [CrossRef] [Green Version]
- D’aniello, C.; Patriarca, E.J.; Phang, J.M.; Minchiotti, G. Proline metabolism in tumor growth and metastatic progression. Front. Oncol. 2020, 10, 776. [Google Scholar] [CrossRef]
- Bayraktutan, Z.; Kiziltunc, A.; Bakan, E.; Alp, H.H. Determination of Endothelial Nitric Oxide Synthase Gene Polymorphism and Plasma Asymmetric Dimethyl Arginine Concentrations in Patients with Lung Cancer. Eurasian J. Med. 2020, 52, 185. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Nilsson, R.; Sharma, S.; Madhusudhan, N.; Kitami, T.; Souza, A.L.; Kafri, R.; Kirschner, M.W.; Clish, C.B.; Mootha, V.K. Metabolite profiling identifies a key role for glycine in rapid cancer cell proliferation. Science 2012, 336, 1040–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stepulak, A.; Rola, R.; Polberg, K.; Ikonomidou, C. Glutamate and its receptors in cancer. J. Neural Transm. 2014, 121, 933–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedley, A.M.; Benkovic, S.J. A new view into the regulation of purine metabolism: The purinosome. Trends Biochem. Sci. 2017, 42, 141–154. [Google Scholar] [CrossRef] [Green Version]
- Yin, J.; Ren, W.; Huang, X.; Deng, J.; Li, T.; Yin, Y. Potential mechanisms connecting purine metabolism and cancer therapy. Front. Immunol. 2018, 9, 1697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, X.; Su, R.; Feng, X.; Wei, M.; Chen, J. Role of N6-methyladenosine modification in cancer. Curr. Opin. Genet. Dev. 2018, 48, 1–7. [Google Scholar] [CrossRef]
- He, L.; Li, H.; Wu, A.; Peng, Y.; Shu, G.; Yin, G. Functions of N6-methyladenosine and its role in cancer. Mol. Cancer 2019, 18, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Qian, Q.; Zhao, X.; Ma, L.; Chen, P. N6-methyladenosine ALKBH5 promotes non-small cell lung cancer progress by regulating TIMP3 stability. Gene 2020, 731, 144348. [Google Scholar] [CrossRef]
- Xu, R.; Pang, G.; Zhao, Q.; Yang, L.; Chen, S.; Jiang, L.; Shen, Y.; Shao, W. The momentous role of N6-methyladenosine in lung cancer. J. Cell. Physiol. 2020, 236, 3244–3256. [Google Scholar] [CrossRef]
- Yiu, A.; Van Hemelrijck, M.; Garmo, H.; Holmberg, L.; Malmström, H.; Lambe, M.; Hammar, N.; Walldius, G.; Jungner, I.; Wulaningsih, W. Circulating uric acid levels and subsequent development of cancer in 493,281 individuals: Findings from the AMORIS Study. Oncotarget 2017, 8, 42332. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Shin, Y.; Kim, T.H.; Kim, D.-H.; Lee, A. Urinary Metabolites as Biomarkers for Diagnosis of Breast Cancer: A Preliminary Study. J. Breast Dis. 2019, 7, 44–51. [Google Scholar] [CrossRef]
- Zhang, L.; Zheng, J.; Ahmed, R.; Huang, G.; Reid, J.; Mandal, R.; Maksymuik, A.; Sitar, D.S.; Tappia, P.S.; Ramjiawan, B. A high-performing plasma metabolite panel for early-stage lung cancer detection. Cancers 2020, 12, 622. [Google Scholar] [CrossRef] [Green Version]
- Hori, S.; Nishiumi, S.; Kobayashi, K.; Shinohara, M.; Hatakeyama, Y.; Kotani, Y.; Hatano, N.; Maniwa, Y.; Nishio, W.; Bamba, T. A metabolomic approach to lung cancer. Lung Cancer 2011, 74, 284–292. [Google Scholar] [CrossRef]
- Bose, S.; Le, A. Glucose metabolism in cancer. Heterog. Cancer Metab. 2018, 1063, 3–12. [Google Scholar]
- Werk, E.E., Jr.; MacGee, J.; Sholiton, L.J. Altered cortisol metabolism in advanced cancer and other terminal illnesses: Excretion of 6-hydroxycortisol. Metabolism 1964, 13, 1425–1438. [Google Scholar] [CrossRef]
- Milosevic, I. Immunofluorometrical Exploring of FSH Levels in the Serum of Patients with Histologically Verified Macrocellular Lung Cancer. Clin. Immunol. 2007, S50–S51. [Google Scholar] [CrossRef]
- Harlos, C.; Musto, G.; Lambert, P.; Ahmed, R.; Pitz, M.W. Androgen pathway manipulation and survival in patients with lung cancer. Horm. Cancer 2015, 6, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Chakroborty, D.; Sarkar, C.; Basu, B.; Dasgupta, P.S.; Basu, S. Catecholamines regulate tumor angiogenesis. Cancer Res. 2009, 69, 3727–3730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabender, C.; Alam, A.; Waters, M.; Mikkelsen, R. Biopterin metabolism drives tumor progression. Cancer Res. 2015, 75. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Characteristics | Total (n = 35) | Urine (n = 29) | Serum (n = 32) |
|---|---|---|---|
| Age Mean (SD) in years | 64.7 (7.4) (n = 34) | 63.8 (7.0) (n = 28) | 64.6 (7.5) (n = 31) |
| Females | 63% (22) | 62% (18) | 62% (20) |
| Males | 37% (13) | 38% (11) | 38% (12) |
| Smoker | 34% (10) | 36% (10) | 35% (9) |
| Ex-Smoker | 52% (15) | 50% (14) | 50% (13) |
| Never Smoked | 14% (4) | 14% (4) | 15% (4) |
| Diabetes | 17% (6) | 10% (3) | 19% (6) |
| COPD | 49% (17) | 48% (14) | 50% (16) |
| Previous cancers | 29% (10) | 31% (9) | 28% (9) |
| On steroids oral/inhalers | 0% | 0% | 0% |
| Squamous cell carcinoma | 14% (5) | 14% (4) | 16% (5) |
| Adenocarcinoma | 80% (28) | 79% (23) | 81% (26) |
| Other | 6% (2) | 7% (2) | 3% (1) |
| Right Upper Lobe | 29% (10) | 31% (9) | 25% (8) |
| Right Lower Lobe | 20% (7) | 17% (5) | 22% (7) |
| Left Upper Lobe | 31% (11) | 34% (10) | 31% (10) |
| Left Lower Lobe | 19% (7) | 17% (5) | 22% (7) |
| PET before surgery | 86% (30) | 86% (25) | 84% (27) |
| Type of surgery: | |||
| Wedge Resection/Segmentectomy | 40% (14) | 38% (11) | 41% (13) |
| Lobectomy | 51% (18) | 55% (16) | 50% (16) |
| Pneumonectomy | 3% (1) | 3% (1) | 3% (1) |
| Wedge and Lobectomy | 6% (2) | 3% (1) | 6% (2) |
| Pathological Stage (n = 34) | n = 28 | n = 31 | |
| T1-T2, N0 M0 | 79% (27) | 75% (21) | 77% (24) |
| T3-T4, N0 M0 | 9% (3) | 11% (3) | 10% (3) |
| T1-T4, N1-2 M0 | 12% (4) | 14% (4) | 13% (4) |
| Mean Tumor size based on CT scan before surgical resection Mean (SD) in cm | 2.4 (1.6) (n = 35) | 2.5 (1.7) (n = 29) | 2.5 (1.7) (n = 32) |
| Mean Tumor size base on surgical pathology Mean (SD) in cm | 2.7 (1.9) (n = 34) | 2.8 (2.0) (n = 28) | 2.8 (1.9) (n = 31) |
| Mean Maximum PET_SUV Mean (SD) | 8.2 (6.5) (n = 30) | 8.2 (6.9) (n = 25) | 8.1 (6.7) (n = 27) |
| PDL1: <1% (10/23) | 43% | 47% (9/19) | 45% (10/22) |
| PDL1: 1–49% (9/23) | 39% | 42% (8/19) | 41% (9/22) |
| PDL1: >50% (4/23) | 17% | 11% (2/19) | 14% (3/22) |
| ALK: Negative (n = 23) | 100% | 100% (n = 20) | 100% (n = 22) |
| Compound | Formula | m/z | Polarity | FC | p Value | Reg | Class | Biofluid | Platform |
|---|---|---|---|---|---|---|---|---|---|
| Lipids and Derivatives | |||||||||
| 2-Propylpent-3-enoic acid | C8H14O2 | 160.1328 | + | 50 | <0.0001 | up | Fatty Acid | Serum | QTOF |
| 13,14-Dihydro PGE1/Prostaglandin F1a | C20H36O5 | 713.493 | + | 5 | 0.047 | up | Prostaglandins | Serum | QTOF |
| 2-Hexenoylcarnitine | C13H23NO4 | 258.1654 | + | 16 | 0.013 | up | Acyl Carnitines | Serum | QTOF |
| 2-Octenoylcarnitine | C15H27NO4 | 286.2013 | + | 3 | 0.026 | up | Acyl Carnitines | Serum | QTOF |
| Chenodeoxycholic/ Deoxycholic acid glycine conjugate/ Glycoursodeoxycholic acid | C26H43NO5 | 450.3208 | + | 28 | 0.0005 | up | Bile Acid | Serum | QTOF |
| Cholic acid | C24H40O5 | 426.3246 ^ | + | 4 | 0.026 | down | Bile Acid | Serum | QTOF |
| cis-5-Tetradecenoylcarnitine | C21H39NO4 | 370.2948 | + | 4 | 0.033 | up | Acyl Carnitines | Serum | QTOF |
| Decanoylcarnitine | C17H33NO4 | 3,162,481 | + | 2 | 0.0001 | up | Acyl Carnitines | Serum | QTOF |
| Dodecanoylcarnitine | C19H38NO4 | 344.2792 | + | 6 | 0.0009 | up | Acyl Carnitines | Serum | QTOF |
| Isopentenyladenine | C16H23N5O5 | 204.1242 | + | 31 | 0.0001 | down | Mevalonate Pathway | Urine | QTOF |
| L-Carnitine | C7H15NO3 | 162.1124 | + | 3 | <0.0001 | up | Carnitines (Lipid Metabolism) | Serum | QTOF |
| L-Octanoylcarnitine | C15H29NO4 | 288.2166 | + | 2 | 0.0008 | up | acyl carnitines | Serum | QTOF |
| LysoPC(P-18:1) | C26H52NO7P | 522.3552 | + | 3 | 0.026 | up | Fatty Acid | Serum | QTOF |
| PG(18:1/18:2) | C42H77O10P | 773.536 | + | 6 | 0.0087 | up | phosphatidyl glycerols | Serum | QTOF |
| PI(16:0/18:1) | C43H81O13P | 854.5691 ^ | + | 7 | 0.0002 | down | phosphatidy linositols | Serum | QTOF |
| Proteins and Derivatives | |||||||||
| 4-Guanidinobutanoic acid | C5H11N3O2 | 163.1156 ^ | + | 4 | 0.029 | up | amino acid (Gamma) | Serum | QTOF |
| Aspartyl glycine | C8H13N3O6 | 248.0938 | + | 50 | 0.0006 | down | dipeptide | Urine | QTOF |
| Asymmetric dimethylarginine (ADMA) | C8H18N4O2 | 203.1505 | + | 16 | 0.044 | down | amino acid | Urine | QTOF |
| Hypoglycin | C7H11NO2 | 142.0875 | + | 442 | <0.0001 | up | amino acid | Urine | QTOF |
| Isodesmosine | C24H40N5O8 | 527.296 | + | 19 | 0.043 | up | amino acid | Serum | QTOF |
| L-Glutamic acid n-butyl ester | C9H17NO4 | 204.1233 | + | 65 | 0.0009 | up | amino acid | Urine | QTOF |
| L-Isoleucyl-L-proline | C11H20N2O3 | 229.152 | + | 2 | 0.0005 | up | dipeptide | Serum | QTOF |
| N(alpha)-t-Butoxycarbonyl-L-leucine | C11H21NO4 | 232.1547 | + | 161 | <0.0001 | down | amino acid | Urine | QTOF |
| Pro Leu | C11H20N2O3 | 229.1548 | + | 625 | <0.0001 | down | dipeptide | Urine | QTOF |
| Serine | C3H7NO3 | 1 | 0.030 | down | amino acid | Serum | NMR | ||
| Carbohydrates | |||||||||
| Myoinositol | C6H12O6 | 203.0524 * | – | 2 | 0.0002 | up | Carbohydrate | Serum | QTOF |
| Glyceraldehyde | C3H6O3 | 203.0524 * | – | 2 | 0.0001 | up | Carbohydrate | Serum | QTOF |
| Glucose | C6H12O6 | 2 | 0.0499 | up | Carbohydrate | Urine | NMR | ||
| Lactate | C3H5O3 | 203.0524 * | – | 2 | 0.013 | up | Glycolysis Product | Urine | NMR |
| Beta-Cortol | 8 | 0.013 | down | Carbohydrate | Serum | QTOF | |||
| Purine/Pyrimidines | |||||||||
| 1-Methyladenine | C6H7N5 | 321.1307 * | + | 59 | <0.0001 | up | Purine | Urine | QTOF |
| 3-Methyluric acid | C6H6N4O3 | 183.0515 | + | 198 | <0.0001 | down | Purine | Urine | QTOF |
| 5-Acetylamino-6-formylamino-3-methyluracil | C8H10N4O4 | 249.0608 * | + | 3 | 0.029 | up | Hydroxypyrimidine | Serum | QTOF |
| N6-Methyladenosine | C11H15N5O4 | 282.1199 | + | 27 | 0.0085 | up | purine nucleoside | Urine | QTOF |
| Carboxylic acid and Derivatives | |||||||||
| cis-Aconitate | C6H6O6 | 1 | 0.035 | up | carboxylic acid | Urine | NMR | ||
| Malonate | C3H3O4 | 2 | 0.014 | up | carboxylic acid | Urine | NMR | ||
| 4-Hydroxycyclohexylcarboxylic acid | C7H12O3 | 162.1126 ^ | + | 3 | <0.0001 | up | carboxylic acid | Serum | QTOF |
| Fumaric acid | C4H4O4 | 139.0026 | + | 17 | 0.011 | down | carboxylic acid | Urine | QTOF |
| Guanidinosuccinic acid | C5H9N3O4 | 176.0654 | + | 3 | 0.023 | up | carboxylic acid (aspartic acid) | Serum | QTOF |
| Proline betaine | C7H14NO2 | 144.1014 | + | 2 | 0.024 | up | carboxylic acid (proline derivative) | Serum | QTOF |
| Succinate | C4H6O4 | 2 | 0.046 | down | carboxylic acid | Urine | NMR | ||
| Unclassified | |||||||||
| Androstanediol | C19H32O2 | 623.4382 $ | + | 54 | <0.0001 | down | Androgens | Serum | QTOF |
| Dopamine | C8H11NO2 | 154.0823 | + | 5 | <0.0001 | up | catecholamine | Serum | QTOF |
| Epinephrine | C9H13NO3 | 184.0944 | + | 134 | <0.0001 | up | catecholamine | Serum | QTOF |
| Androstenedione | C19H26O2 | 287.2041 | + | 8 | 0.023 | up | Androgens | Serum | QTOF |
| Tetrahydrobiopterin/Sapropterin (BH4, THB) | C9H15N5O3 | 500.2767 ǂ | + | 8 | 0.0031 | up | Biopterin | Serum | QTOF |
| N-Desmethylaminopyrine | C12H15N3O | 218.1378 | + | 10 | 0.0003 | up | phenylpyrazoles | Serum | QTOF |
| Source or Class of Metabolites | Frequency n (%) |
|---|---|
| All | 48 (100) |
| Serum | 31 (65) |
| Urine | 17 (35) |
| Lipids and Derivatives | 15 (31) |
| Protein and Derivatives | 11(23) |
| Carboxylic Acid and Derivatives | 7 (15) |
| Unclassified | 6 (13) |
| Carbohydrates | 5 (10) |
| Purine/Pyrimidines | 4 (8) |
| Class of Metabolites | Upregulated | Downregulated | p Value | ||||||
|---|---|---|---|---|---|---|---|---|---|
| N | Med FC | Min FC | Max FC | n | Med FC | Min FC | Max FC | ||
| All | 34 | 4 | 1 | 442 | 14 | 16 | 1 | 625 | 0.043 |
| Biofluid | |||||||||
| Serum | 26 | 4 | 2 | 134 | 5 | 7 | 1 | 54 | 0.69 |
| Urine | 8 | 14 | 1 | 442 | 9 | 31 | 2 | 625 | 0.28 |
| Class | |||||||||
| Lipids and derivatives | 12 | 4 | 2 | 50 | 3 | 7 | 4 | 31 | |
| Proteins and derivatives | 5 | 19 | 2 | 442 | 6 | 33 | 1 | 625 | |
| Carbohydrates | 4 | 2 | 2 | 2 | 1 | 8 | |||
| Purine/Pyrimidines | 3 | 27 | 3 | 59 | 1 | 198 | |||
| Carboxylic acid and derivatives | 5 | 2 | 1 | 3 | 2 | 9 | 2 | 17 | |
| Unclassified | 5 | 8 | 5 | 134 | 1 | 54 | |||
| Class | Biofluid | Metabolite | Regulation | Fold Change | Previously Known Association to Cancer Pathogenesis |
|---|---|---|---|---|---|
| Protein and Derivatives | Urine | Pro Leu | down | 625 | Yes |
| Purine/Pyrimidines | Urine | 3-Methyluric acid | down | 198 | Yes |
| Protein and Derivatives | Urine | N(alpha)-t-Butoxycarbonyl-L-leucine | down | 161 | No |
| Androgens | Serum | Androstanediol | down | 54 | Yes |
| Protein and Derivatives | Urine | Aspartyl glycine | down | 50 | Yes |
| Lipid and Derivatives | Urine | Isopentenyladenine | down | 31 | Yes |
| Carboxylic Acid and Derivatives | Urine | Fumaric acid | down | 17 | Yes |
| Protein and Derivatives | Urine | Asymmetric dimethylarginine (ADMA) | down | 16 | Yes |
| Protein and Derivatives | Urine | Hypoglycin | up | 442 | No |
| Unclassified | Serum | Epinephrine | up | 134 | Yes |
| Protein and Derivatives | Urine | L-Glutamic acid n-butyl ester | up | 65 | Yes |
| Purine/Pyrimidines | Urine | 1-Methyladenine | up | 59 | No |
| Lipid and Derivatives | Serum | 2-Propylpent-3-enoic acid | up | 50 | No |
| Lipid and Derivative | Serum | Chenodeoxycholic/Deoxycholic/ Glycoursodeoxycholic acid | up | 28 | Yes |
| Purine/Pyrimidines | Urine | N6-Methyladenosine | up | 27 | Yes |
| Protein and Derivative | Serum | Isodesmosine | up | 19 | No |
| Lipid and Derivative | Serum | 2-Hexenoylcarnitine | up | 16 | Possible |
| Unclassified | Serum | N-Desmethylaminopyrine | up | 10.3 | No |
| Metabolite | Regulation | Metabolic Pathway |
|---|---|---|
| Pro Leu | down | Proline metabolism facilitated by PRODH/POX |
| 3-Methyluric acid | down | Purine Metabolism |
| Androstanediol | down | Androgen pathway through AR in NSCLC |
| Aspartyl glycine | down | Mitochondrial glycine biosynthetic pathway |
| Isopentenyladenine | down | Mevalonate pathway |
| Fumaric acid | down | Krebs cycle |
| Asymmetric dimethylarginine (ADMA) | down | Overexpression of protein arginine methyl transferase |
| Epinephrine | up | Angiogenesis |
| 2-Propylpent-3-enoic acid | up | Fatty acid metabolism |
| L-Glutamic acid n-butyl ester | up | Growth factor, metabotropic/ionotropic glutamate receptors |
| Chenodeoxycholic/Deoxycholic/ Glycoursodeoxycholic acid | up | Over expressed FXR |
| N6-Methyladenosine | up | Degradation of tRNA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, N.; Kidane, B.; Wang, L.; Nugent, Z.; Moldovan, N.; McElrea, A.; Shariati-Ievari, S.; Qing, G.; Tan, L.; Buduhan, G.; et al. Metabolic Changes in Early-Stage Non–Small Cell Lung Cancer Patients after Surgical Resection. Cancers 2021, 13, 3012. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13123012
Ahmed N, Kidane B, Wang L, Nugent Z, Moldovan N, McElrea A, Shariati-Ievari S, Qing G, Tan L, Buduhan G, et al. Metabolic Changes in Early-Stage Non–Small Cell Lung Cancer Patients after Surgical Resection. Cancers. 2021; 13(12):3012. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13123012
Chicago/Turabian StyleAhmed, Naseer, Biniam Kidane, Le Wang, Zoann Nugent, Nataliya Moldovan, April McElrea, Shiva Shariati-Ievari, Gefei Qing, Lawrence Tan, Gordon Buduhan, and et al. 2021. "Metabolic Changes in Early-Stage Non–Small Cell Lung Cancer Patients after Surgical Resection" Cancers 13, no. 12: 3012. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13123012