
SPTAN1 Expression Predicts Treatment and Survival Outcomes in Colorectal Cancer

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Ethical Approval
2.2. Protein Analysis
2.3. Gene Expression Data
2.4. Single-Sample Gene Set Enrichment Analysis
2.5. Gene Set Enrichment Analysis
2.6. Treatment Response Prediction
2.7. Cell Lines
2.8. Generation of Lentiviral Particles and shRNA Knockdown
2.9. Western Blotting
2.10. Cell Viability Assay
2.11. Statistical Analysis
3. Results
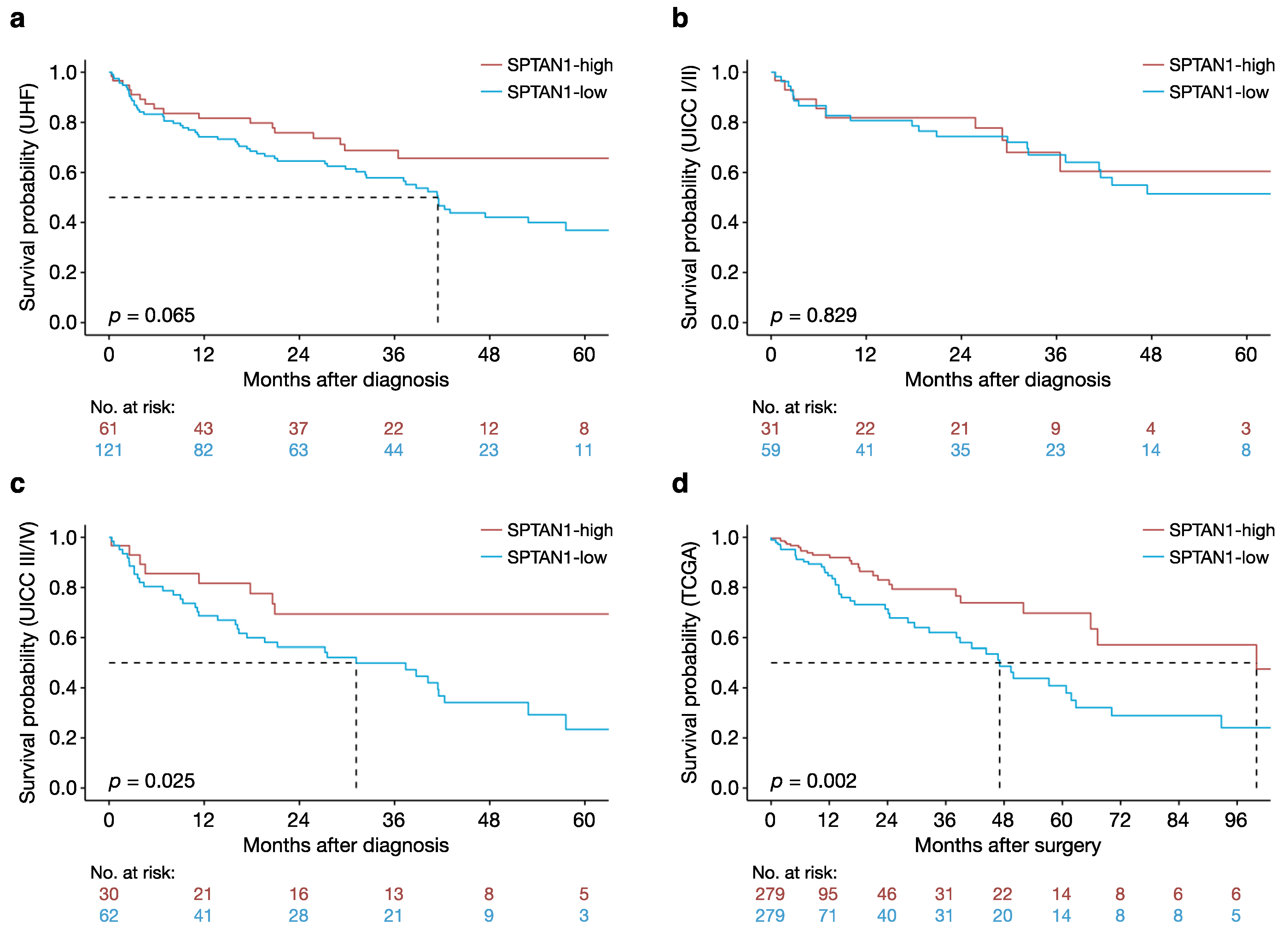
3.1. High Intratumoural SPTAN1 Protein Levels Predict Better Overall Survival in Patients with CRC
3.2. High Intratumoural SPTAN1 mRNA Levels Predict Better Overall Survival in Patients with CRC
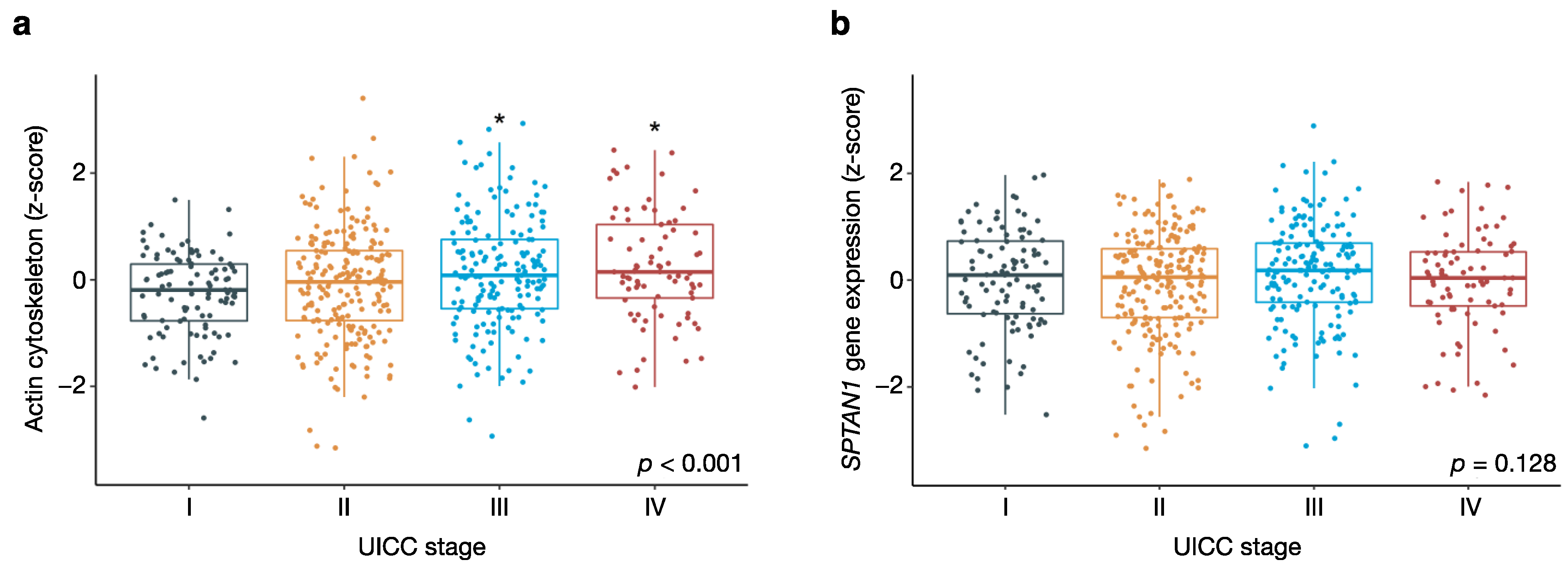
3.3. Actin Cytoskeletal Genes Are Upregulated in Aggressive Tumours with Distant Metastasis
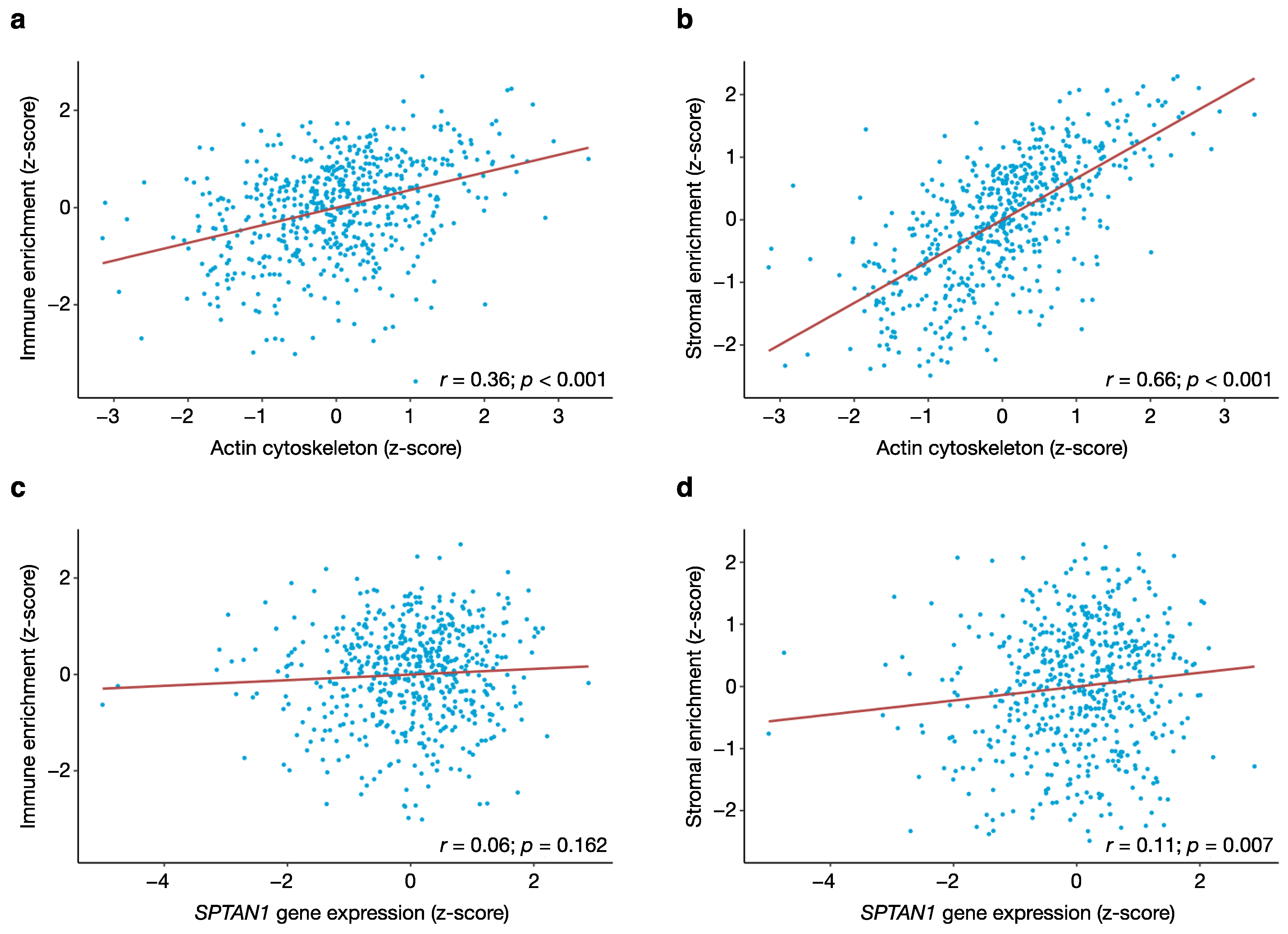
3.4. Tumour Purity Affects the Expression of Actin Cytoskeletal Genes But Not SPTAN1 Gene Expression
3.5. SPTAN1 Is Related to Cell Adhesion and Polarity, Cytoskeletal Organisation, Motility and Invasion
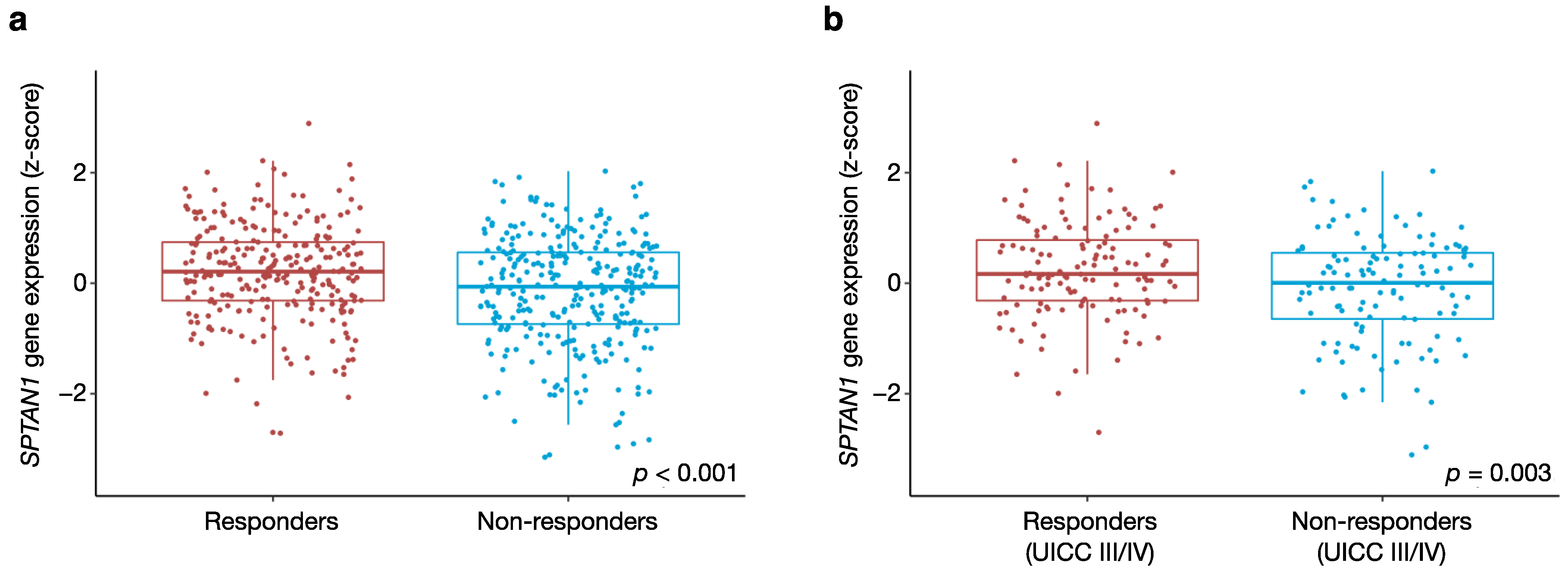
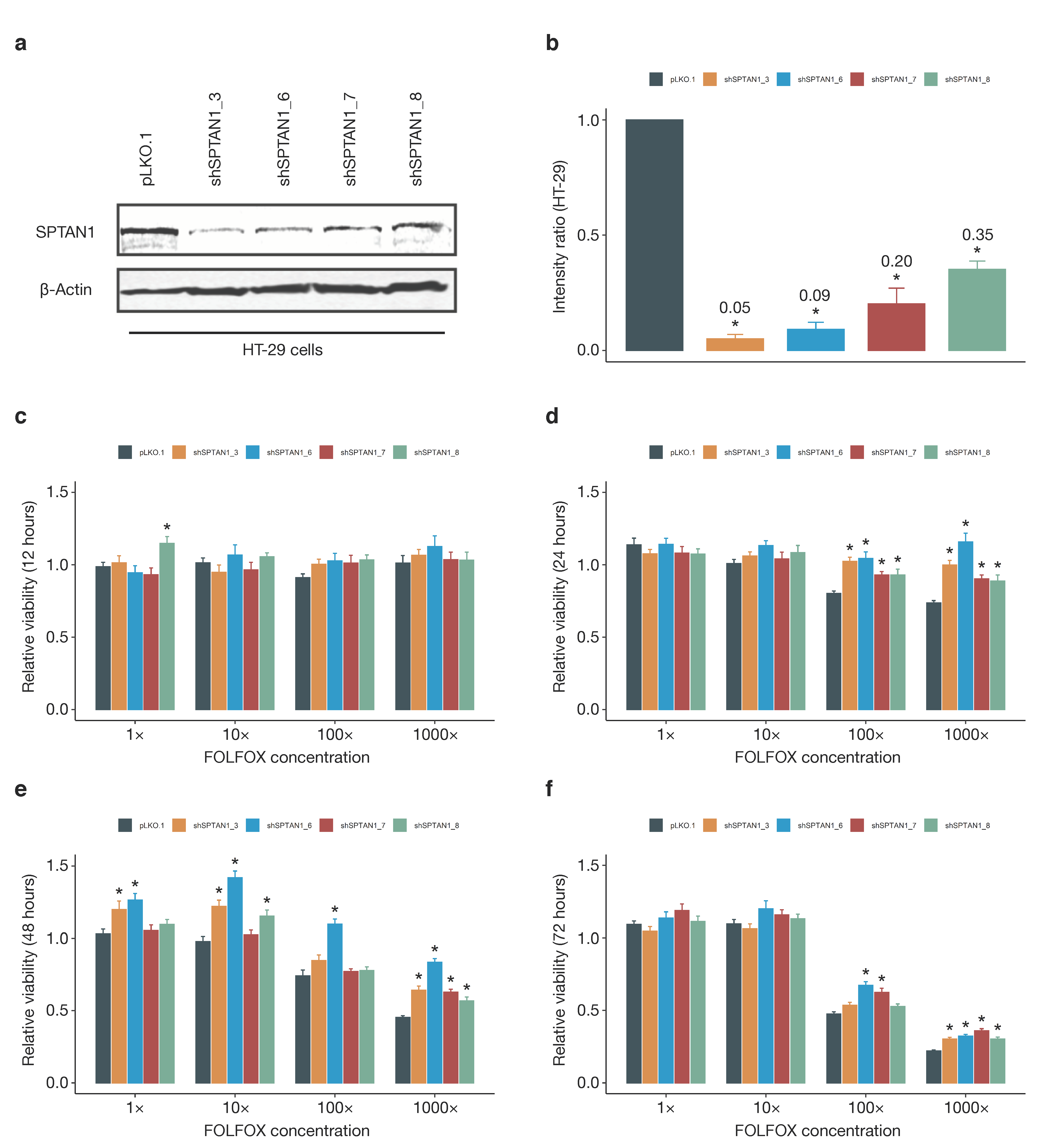
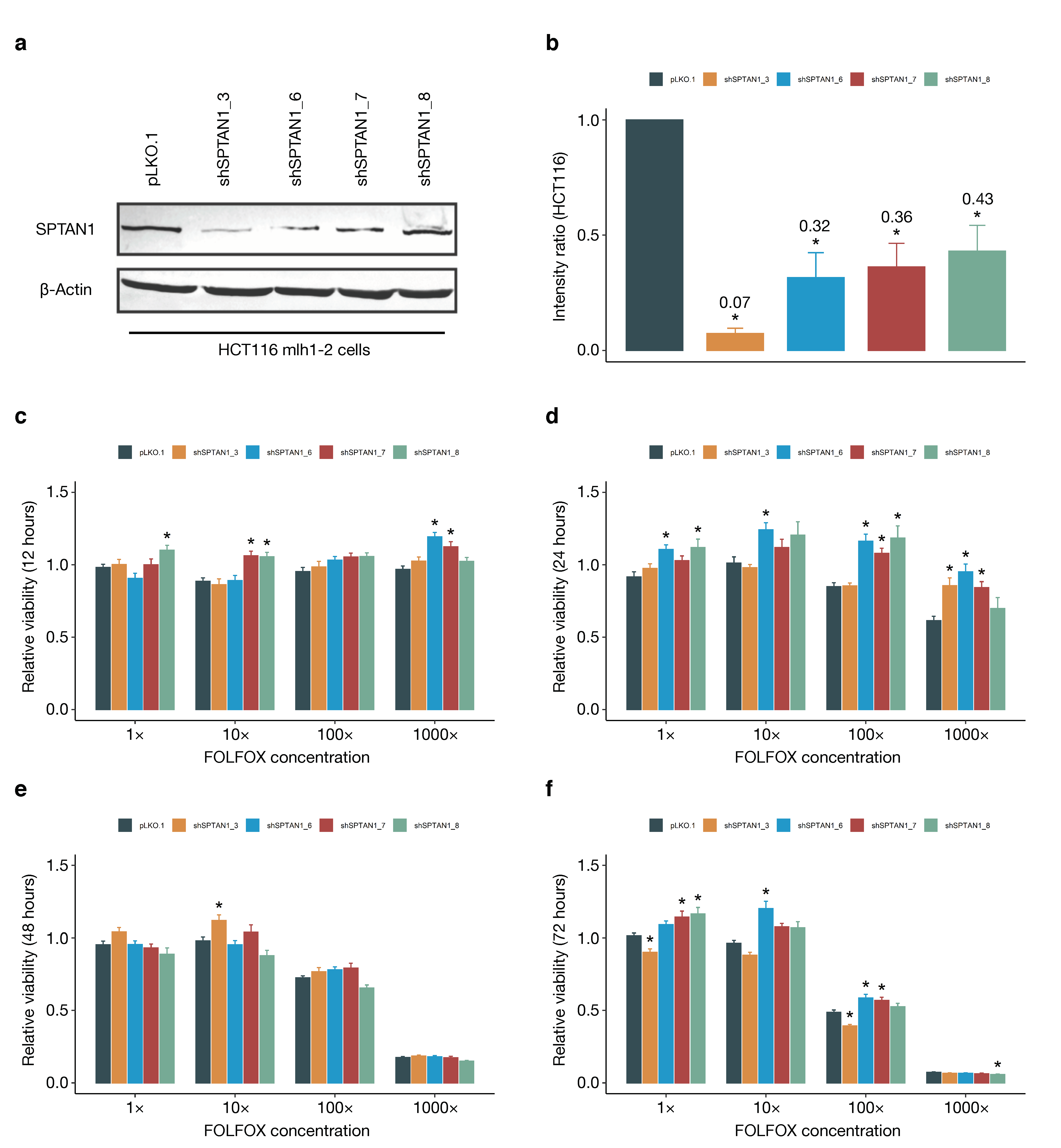
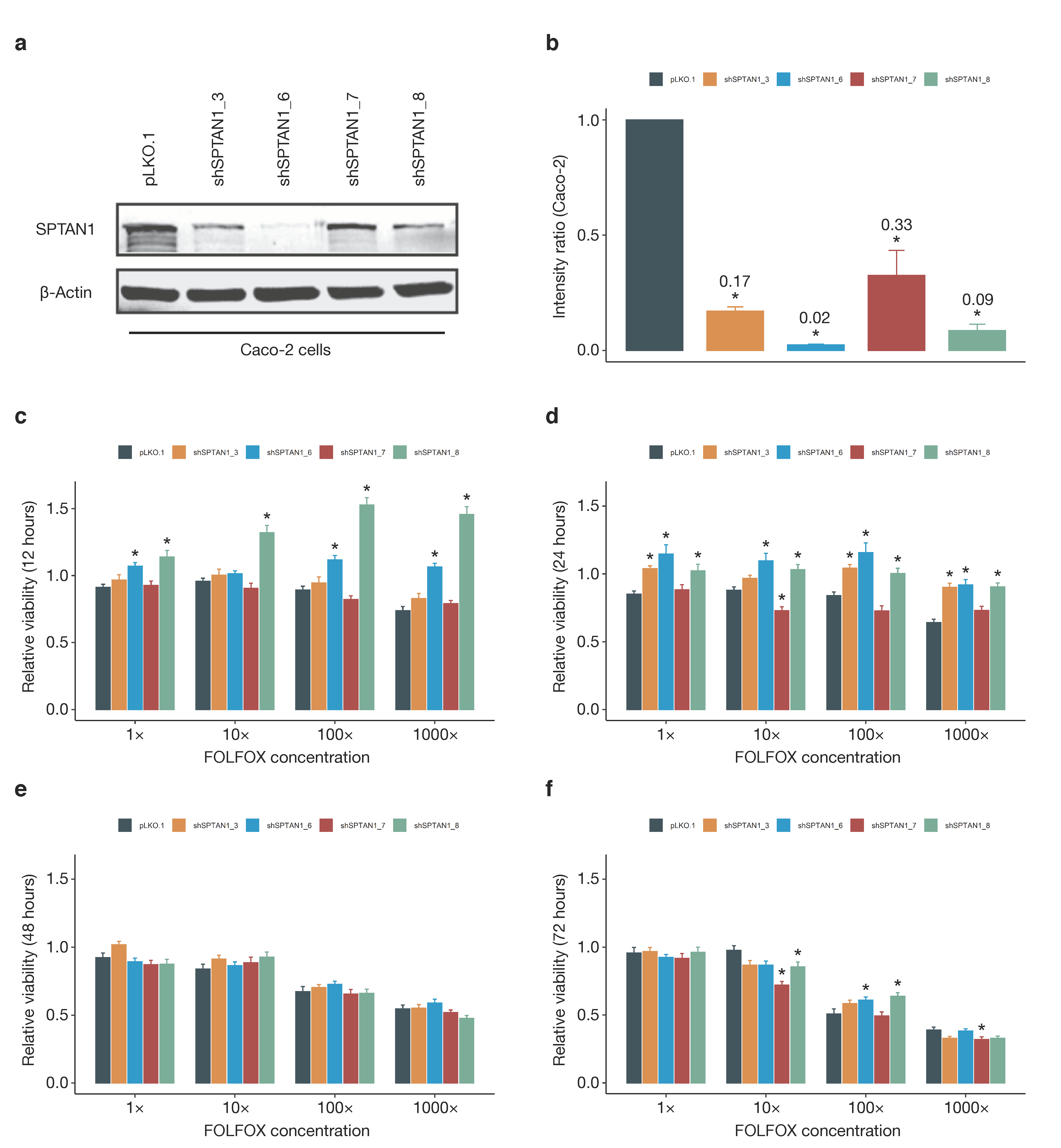
3.6. SPTAN1 Gene Expression Predicts Response to FOLFOX Chemotherapy
4. Discussion
4.1. SPTAN1 as a Prognostic Biomarker
4.2. Biological Roles of SPTAN1 in Cancer
4.3. SPTAN1 as a Predictor of Therapy Response
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Ralston, S.; Penman, I.; Strachan, M.; Hobson, R. Davidson’s Principles and Practice of Medicine, 23rd ed.; Churchill Livingstone: Edinburgh, UK, 2018. [Google Scholar]
- Boland, C.R.; Goel, A. Microsatellite instability in colorectal cancer. Gastroenterology 2010, 138, 2073–2087.e3. [Google Scholar] [CrossRef] [PubMed]
- Gryfe, R.; Kim, H.; Hsieh, E.T.; Aronson, M.D.; Holowaty, E.J.; Bull, S.B.; Redston, M.; Gallinger, S. Tumor microsatellite instability and clinical outcome in young patients with colorectal cancer. N. Engl. J. Med. 2000, 342, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Malesci, A.; Laghi, L.; Bianchi, P.; Delconte, G.; Randolph, A.; Torri, V.; Carnaghi, C.; Doci, R.; Rosati, R.; Montorsi, M.; et al. Reduced likelihood of metastases in patients with microsatellite-unstable colorectal cancer. Clin. Cancer Res. 2007, 13, 3831–3839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brieger, A.; Adryan, B.; Wolpert, F.; Passmann, S.; Zeuzem, S.; Trojan, J. Cytoskeletal scaffolding proteins interact with Lynch-Syndrome associated mismatch repair protein MLH1. Proteomics 2010, 10, 3343–3355. [Google Scholar] [CrossRef] [PubMed]
- Hinrichsen, I.; Ernst, B.P.; Nuber, F.; Passmann, S.; Schäfer, D.; Steinke, V.; Friedrichs, N.; Plotz, G.; Zeuzem, S.; Brieger, A. Reduced migration of MLH1 deficient colon cancer cells depends on SPTAN1. Mol. Cancer 2014, 13, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ackermann, A.; Schrecker, C.; Bon, D.; Friedrichs, N.; Bankov, K.; Wild, P.; Plotz, G.; Zeuzem, S.; Herrmann, E.; Hansmann, M.L.; et al. Downregulation of SPTAN1 is related to MLH1 deficiency and metastasis in colorectal cancer. PLoS ONE 2019, 14, e0213411. [Google Scholar] [CrossRef] [Green Version]
- Machnicka, B.; Grochowalska, R.; Bogusławska, D.M.; Sikorski, A.F.; Lecomte, M.C. Spectrin-based skeleton as an actor in cell signaling. Cell Mol. Life Sci. 2012, 69, 191–201. [Google Scholar] [CrossRef] [Green Version]
- An, X.; Mohandas, N. Disorders of red cell membrane. Br. J. Haematol. 2008, 141, 367–375. [Google Scholar] [CrossRef]
- Ackermann, A.; Brieger, A. The Role of Nonerythroid Spectrin αII in Cancer. J. Oncol. 2019, 2019, 7079604. [Google Scholar] [CrossRef] [Green Version]
- Guinney, J.; Dienstmann, R.; Wang, X.; de Reyniès, A.; Schlicker, A.; Soneson, C.; Marisa, L.; Roepman, P.; Nyamundanda, G.; Angelino, P.; et al. The consensus molecular subtypes of colorectal cancer. Nat. Med. 2015, 21, 1350–1356. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Barbie, D.A.; Tamayo, P.; Boehm, J.S.; Kim, S.Y.; Moody, S.E.; Dunn, I.F.; Schinzel, A.C.; Sandy, P.; Meylan, E.; Scholl, C.; et al. Systematic RNA interference reveals that oncogenic KRAS-driven cancers require TBK1. Nature 2009, 462, 108–112. [Google Scholar] [CrossRef]
- Yoshihara, K.; Shahmoradgoli, M.; Martínez, E.; Vegesna, R.; Kim, H.; Torres-Garcia, W.; Treviño, V.; Shen, H.; Laird, P.W.; Levine, D.A.; et al. Inferring tumour purity and stromal and immune cell admixture from expression data. Nat. Commun. 2013, 4, 2612. [Google Scholar] [CrossRef]
- Reich, M.; Liefeld, T.; Gould, J.; Lerner, J.; Tamayo, P.; Mesirov, J.P. GenePattern 2.0. Nat. Genet. 2006, 38, 500–501. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Liberzon, A.; Birger, C.; Thorvaldsdóttir, H.; Ghandi, M.; Mesirov, J.P.; Tamayo, P. The Molecular Signatures Database (MSigDB) hallmark gene set collection. Cell Syst. 2015, 1, 417–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Cheng, J.; Guan, Q.; Yan, H.; Li, Y.; Zhao, W.; Guo, Z.; Wang, X. Qualitative transcriptional signature for predicting pathological response of colorectal cancer to FOLFOX therapy. Cancer Sci. 2020, 111, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Dirks, W.G.; Faehnrich, S.; Estella, I.A.; Drexler, H.G. Short tandem repeat DNA typing provides an international reference standard for authentication of human cell lines. Altex 2005, 22, 103–109. [Google Scholar]
- Berg, K.C.G.; Eide, P.W.; Eilertsen, I.A.; Johannessen, B.; Bruun, J.; Danielsen, S.A.; Bjørnslett, M.; Meza-Zepeda, L.A.; Eknæs, M.; Lind, G.E.; et al. Multi-omics of 34 colorectal cancer cell lines—A resource for biomedical studies. Mol. Cancer 2017, 16, 116. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 30 April 2020).
- Therneau, T. A Package for Survival Analysis in R. R Package Version 3.2–7. 2020. Available online: https://CRAN.R-project.org/package=survival (accessed on 30 April 2020).
- Kassambara, A. Survminer: Drawing Survival Curves Using ‘ggplot2’. 2020. Available online: https://CRAN.R-project.org/package=survminer (accessed on 30 April 2020).
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yilmaz, M.; Christofori, G. EMT, the cytoskeleton, and cancer cell invasion. Cancer Metastasis Rev. 2009, 28, 15–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, H.T.; Machesky, L.M. Actin cytoskeletal control during epithelial to mesenchymal transition: Focus on the pancreas and intestinal tract. Br. J. Cancer 2015, 112, 613–620. [Google Scholar] [CrossRef] [Green Version]
- Chakravarthy, A.; Khan, L.; Bensler, N.P.; Bose, P.; De Carvalho, D.D. TGF-β-associated extracellular matrix genes link cancer-associated fibroblasts to immune evasion and immunotherapy failure. Nat. Commun. 2018, 9, 4692. [Google Scholar] [CrossRef] [PubMed]
- Klingler-Hoffmann, M.; Mittal, P.; Hoffmann, P. The Emerging Role of Cytoskeletal Proteins as Reliable Biomarkers. Proteomics 2019, 19, e1800483. [Google Scholar] [CrossRef] [Green Version]
- Younes, M.; Harris, A.S.; Morrow, J.S. Fodrin as a differentiation marker. Redistributions in colonic neoplasia. Am. J. Pathol. 1989, 135, 1197–1212. [Google Scholar] [PubMed]
- Simpson, J.F.; Page, D.L. Altered expression of a structural protein (fodrin) within epithelial proliferative disease of the breast. Am. J. Pathol. 1992, 141, 285–289. [Google Scholar]
- Sormunen, R.; Pääkkö, P.; Palovuori, R.; Soini, Y.; Lehto, V.P. Fodrin and actin in the normal, metaplastic, and dysplastic respiratory epithelium and in lung carcinoma. Am. J. Respir. Cell Mol. Biol. 1994, 11, 75–84. [Google Scholar] [CrossRef]
- Tuominen, H.; Sormunen, R.; Kallioinen, M. Non-erythroid spectrin (fodrin) in cutaneous tumours: Diminished in cell membranes, increased in the cytoplasm. Br. J. Dermatol. 1996, 135, 576–580. [Google Scholar] [CrossRef]
- Lee, S.; Baek, M.; Yang, H.; Bang, Y.J.; Kim, W.H.; Ha, J.H.; Kim, D.K.; Jeoung, D.I. Identification of genes differentially expressed between gastric cancers and normal gastric mucosa with cDNA microarrays. Cancer Lett. 2002, 184, 197–206. [Google Scholar] [CrossRef]
- Sun, R.; Meng, X.; Wang, W.; Liu, B.; Lv, X.; Yuan, J.; Zeng, L.; Chen, Y.; Yuan, B.; Yang, S. Five genes may predict metastasis in non-small cell lung cancer using bioinformatics analysis. Oncol. Lett. 2019, 18, 1723–1732. [Google Scholar] [CrossRef]
- Cancer Genome Atlas Network. Comprehensive molecular characterization of human colon and rectal cancer. Nature 2012, 487, 330–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Nicolas, G.; Fournier, C.M.; Galand, C.; Malbert-Colas, L.; Bournier, O.; Kroviarski, Y.; Bourgeois, M.; Camonis, J.H.; Dhermy, D.; Grandchamp, B.; et al. Tyrosine phosphorylation regulates alpha II spectrin cleavage by calpain. Mol. Cell Biol. 2002, 22, 3527–3536. [Google Scholar] [CrossRef] [Green Version]
- Nedrelow, J.H.; Cianci, C.D.; Morrow, J.S. c-Src binds alpha II spectrin’s Src homology 3 (SH3) domain and blocks calpain susceptibility by phosphorylating Tyr1176. J. Biol. Chem. 2003, 278, 7735–7741. [Google Scholar] [CrossRef] [Green Version]
- Weigand, J.E.; Boeckel, J.N.; Gellert, P.; Dimmeler, S. Hypoxia-induced alternative splicing in endothelial cells. PLoS ONE 2012, 7, e42697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, S.J.; O’Brien, G.A.; Nishioka, W.K.; McGahon, A.J.; Mahboubi, A.; Saido, T.C.; Green, D.R. Proteolysis of fodrin (non-erythroid spectrin) during apoptosis. J. Biol. Chem. 1995, 270, 6425–6428. [Google Scholar] [CrossRef] [Green Version]
- Rotter, B.; Kroviarski, Y.; Nicolas, G.; Dhermy, D.; Lecomte, M.C. AlphaII-spectrin is an in vitro target for caspase-2, and its cleavage is regulated by calmodulin binding. Biochem. J. 2004, 378, 161–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubielecka, P.M.; Grzybek, M.; Kolondra, A.; Jaźwiec, B.; Draga, A.; Aleksandrowicz, P.; Kołodziejczyk, M.; Serwotka, A.; Dolińska-Krajewska, B.; Warchoł, J.; et al. Aggregation of spectrin and PKCtheta is an early hallmark of fludarabine/mitoxantrone/dexamethasone-induced apoptosis in Jurkat T and HL60 cells. Mol. Cell Biochem. 2010, 339, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Ribic, C.M.; Sargent, D.J.; Moore, M.J.; Thibodeau, S.N.; French, A.J.; Goldberg, R.M.; Hamilton, S.R.; Laurent-Puig, P.; Gryfe, R.; Shepherd, L.E.; et al. Tumor microsatellite-instability status as a predictor of benefit from fluorouracil-based adjuvant chemotherapy for colon cancer. N. Engl. J. Med. 2003, 349, 247–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sargent, D.J.; Marsoni, S.; Monges, G.; Thibodeau, S.N.; Labianca, R.; Hamilton, S.R.; French, A.J.; Kabat, B.; Foster, N.R.; Torri, V.; et al. Defective mismatch repair as a predictive marker for lack of efficacy of fluorouracil-based adjuvant therapy in colon cancer. J. Clin. Oncol. 2010, 28, 3219–3226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- L’Espérance, S.; Popa, I.; Bachvarova, M.; Plante, M.; Patten, N.; Wu, L.; Têtu, B.; Bachvarov, D. Gene expression profiling of paired ovarian tumors obtained prior to and following adjuvant chemotherapy: Molecular signatures of chemoresistant tumors. Int. J. Oncol. 2006, 29, 5–24. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Liu, C.; Niu, Y.; Jing, Y.; Zhang, H.; Wang, J.; Yang, J.; Zen, K.; Zhang, J.; Zhang, C.Y.; et al. MicroRNA-128-3p regulates mitomycin C-induced DNA damage response in lung cancer cells through repressing SPTAN1. Oncotarget 2017, 8, 58098–58107. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Hazard Ratio (95% CI) | Significance Level (p-Value) |
|---|---|---|
| SPTAN1 Protein (high vs. low) | 0.59 (0.35–1.01) | 0.054 |
| Age at Diagnosis (Years) | 1.03 * (1.01–1.06) | 0.001 |
| UICC Stage II (vs. Stage I) | 1.14 (0.54–2.41) | 0.737 |
| UICC Stage III (vs. Stage I) | 1.01 (0.47–2.18) | 0.982 |
| UICC Stage IV (vs. Stage I) | 3.45 * (1.72–6.92) | 5.0 × 10−4 |
| Variables | Hazard Ratio (95% CI) | Significance Level (p-Value) |
|---|---|---|
| SPTAN1 mRNA (z-Score) | 0.59 * (0.45–0.76) | 7.4 × 10−5 |
| Age at Surgery (Years) | 1.03 * (1.01–1.05) | 0.003 |
| UICC Stage II (vs. Stage I) | 1.11 (0.41–3.01) | 0.830 |
| UICC Stage III (vs. Stage I) | 1.37 (0.50–3.71) | 0.538 |
| UICC Stage IV (vs. Stage I) | 4.34 * (1.58–11.87) | 0.004 |
| Cytoskeletal Genes (z-Score) | 2.58 * (1.56–4.26) | 2.0 × 10−4 |
| Adhesion Genes (z-Score) | 0.54 * (0.34–0.86) | 0.009 |
| Gene Set | NES | p-Value | FDR |
|---|---|---|---|
| Hallmark Apical Junction | 1.82 | <0.001 | 0.044 |
| Hallmark Apical Surface | 1.66 | 0.010 | 0.098 |
| KEGG Focal Adhesion | 1.94 | 0.002 | 0.033 |
| KEGG Gap Junction | 1.81 | 0.002 | 0.048 |
| KEGG Adherens Junction | 1.74 | 0.006 | 0.047 |
| KEGG Tight Junction | 1.74 | 0.004 | 0.046 |
| GO Actin Cytoskeleton | 1.82 | <0.001 | 0.062 |
| GO Actin Filament Organisation | 1.85 | <0.001 | 0.057 |
| GO Actin Filament-Based Movement | 1.75 | 0.010 | 0.078 |
| GO Lamellipodium | 1.75 | 0.004 | 0.078 |
| GO Invadopodium | 1.64 | 0.022 | 0.105 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schrecker, C.; Behrens, S.; Schönherr, R.; Ackermann, A.; Pauli, D.; Plotz, G.; Zeuzem, S.; Brieger, A. SPTAN1 Expression Predicts Treatment and Survival Outcomes in Colorectal Cancer. Cancers 2021, 13, 3638. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13143638
Schrecker C, Behrens S, Schönherr R, Ackermann A, Pauli D, Plotz G, Zeuzem S, Brieger A. SPTAN1 Expression Predicts Treatment and Survival Outcomes in Colorectal Cancer. Cancers. 2021; 13(14):3638. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13143638
Chicago/Turabian StyleSchrecker, Christopher, Sophia Behrens, Rebecca Schönherr, Anne Ackermann, Daniel Pauli, Guido Plotz, Stefan Zeuzem, and Angela Brieger. 2021. "SPTAN1 Expression Predicts Treatment and Survival Outcomes in Colorectal Cancer" Cancers 13, no. 14: 3638. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13143638