Resveratrol’s Anti-Cancer Effects through the Modulation of Tumor Glucose Metabolism




 , ,
, ,  ,
,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Goal of the Review
3. Source of the Data
4. Glucose Metabolism (Glycolysis) in Tumors and the Warburg Effect
5. Glycolysis in Tumor Cells and the Pentose Phosphate Pathway (PPP)
6. The Influence of the Tumor Microenvironment on the Warburg Effect
7. Resveratrol: A Multi-Targeted Agent for the Prevention and Treatment of Chronic Diseases, Including Tumors
7.1. Resveratrol, Inflammation, and Tumors
7.2. Resveratrol and Tumors
7.2.1. Breast Cancer
7.2.2. Lung Cancer
7.2.3. Colorectal Cancer (CRC)
7.2.4. Prostate Cancer
8. Resveratrol: Its Impact on Intracellular Molecular Signaling Targets Related to Glucose Metabolism in Tumors
9. Sirtuins as Major Intracellular Targets for Resveratrol in Modulating Tumor Glucose Metabolism
10. Pharmacokinetics of Resveratrol: Challenges and Future Perspectives
10.1. Absorption
10.2. Metabolism
10.3. Bioavailability and Tissue Distribution
10.4. Improving the Biological Effectiveness of Resveratrol
11. Clinical Trials with Resveratrol
12. Conclusions and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 6PGDH | 6-phosphogluconate-dehydrogenase |
| Ach | Acetylcholine |
| AR | Androgen receptor |
| ATO | Arsenic trioxide |
| Bax | Bcl-2-associated X protein |
| Bcl-2 | B-cell lymphoma-2 |
| B-CLL | B-cell chronic lymphocytic leukemia |
| CamKKB/AMPK | Ca2+ Calmodulin kinase kinase B/AMP-activated kinase pathway |
| CHOP | CCAAT/enhancer-binding protein-homologous protein |
| CML | Chronic myelogenous leukemia |
| COX-2 | Cyclooxygenase-2 |
| CRC | Colorectal Cancer |
| CRPC | Castration-resistant prostate cancer |
| CYP1A1 | Cytochrome P450, family 1, subfamily A, polypeptide 1 |
| ECM | Extracellular Matrix |
| EMT | Epithelial-to-mesenchymal transition |
| FAK | Focal adhesion kinase |
| FOXO | Forkhead O box |
| G6PD | Glucose-6-phosphate-dehydrogenase |
| GBA1 | Glucosylceramidase beta 1 |
| GLUT1 | Glucose transporter 1 |
| GRP 78 | Glucose-regulated protein 78 |
| HBV | Hepatitis B virus |
| HCC | Hepatocellular carcinoma |
| HCL | Hairy cell leukemia |
| HER-2 | Human epidermal growth factor receptor-2 |
| HGF | Hepatocyte growth factor |
| HIF-1α | Hypoxia-inducible factor-1 alpha |
| HK2 | Hexokinase II |
| LDH | Lactate dehydrogenase A |
| mEH | Microsomal epoxide hydrolase |
| MMP | Matrix-metalloprotease |
| mTORC1 | Mechanistic target of rapamycin complex 1 |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B-cells |
| NQO-1 | NAD(P)H quinone oxidoreductase 1 |
| NSCLC | Non-small-cell lung cancer |
| OXPHOS | Oxidative phosphorylation |
| PCCs | Prostate cancer cells |
| PDAC | Pancreatic ductal adenocarcinoma |
| PDH | Pyruvate dehydrogenase |
| PDK1 | Pyruvate dehydrogenase kinase 1 |
| PDK4 | Pyruvate dehydrogenase lipoamide kinase 4 |
| PFK1 | 6-phosphofructo-1-kinase |
| PGAM | Phosphoglycerate mutase |
| PGC-1 | Proliferator-activated receptor-gamma-coactivator-1 |
| PI3K | Phosphoinositide 3-kinase signaling |
| PK | Pyruvate kinase |
| PKM2 | Pyruvate kinase M2 |
| PPP | Pentose phosphate pathway |
| pRb | Retinoblastoma protein |
| PSA | Prostate-specific antigen |
| PSCs | Pancreatic stellate cells |
| PTP1B | Tyrosine-protein phosphatase non-receptor type 1 |
| R5P | Ribose-5-phosphate |
| ROS | Reactive oxygen species |
| SREBP1 | Sterol regulatory element binding protein 1 |
| SULT | Human sulfotransferases |
| TKT | Transketolase |
| TCA | Tricarboxylic acid |
| UGT | UDP-glucuronosyltransferase |
| VDAC1 | Voltage-dependent anion channel 1 |
| VEGF | Vascular endothelial growth factor |
| WNT | Wnt signaling pathway |
| XBP1 | X-box binding protein 1 |
References
- Gerweck, L.E.; Seetharaman, K. Cellular pH gradient in tumor versus normal tissue: Potential exploitation for the treatment of cancer. Cancer Res. 1996, 56, 1194–1198. [Google Scholar]
- Warburg, O. On the Origin of Cancer Cells. Science 1956, 123, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Warburg, O.; Wind, F.; Negelein, E. The Metabolism of Tumors in the Body. J. Gen. Physiol. 1927, 8, 519–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolfe, D.F.; Brown, G.C. Cellular energy utilization and molecular origin of standard metabolic rate in mammals. Physiol. Rev. 1997, 77, 731–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vazquez, A.; Kamphorst, J.J.; Markert, E.K.; Schug, Z.T.; Tardito, S.; Gottlieb, E. Cancer metabolism briefly. J. Cell Sci. 2016, 129, 3367–3373. [Google Scholar] [CrossRef] [Green Version]
- Nonomura, S.; Kanagawa, H.; Makimoto, A. Chemical Constituents of Polygonaceous Plants. I. Studies on the Components of KO-J O-KON. (Polygonum cuspidatum Sieb. et zucc.). Yakugaku Zasshi J. Pharm. Soc. Jpn. 1963, 83, 988–990. [Google Scholar] [CrossRef] [Green Version]
- Frémont, L. Biological effects of resveratrol. Life Sci. 2000, 66, 663–673. [Google Scholar] [CrossRef]
- Jang, M.; Cai, L.; Udeani, G.O.; Slowing, K.V.; Thomas, C.F.; Beecher, C.W.W.; Fong, H.H.S.; Farnsworth, N.R.; Kinghorn, A.D.; Mehta, R.G.; et al. Cancer Chemopreventive Activity of Resveratrol, a Natural Product Derived from Grapes. Science 1997, 275, 218–220. [Google Scholar] [CrossRef] [Green Version]
- Baur, J.A.; Pearson, K.J.; Price, N.L.; Jamieson, H.A.; Lerin, C.; Kalra, A.; Prabhu, V.V.; Allard, J.S.; Lopez-Lluch, G.; Lewis, K.; et al. Resveratrol improves health and survival of mice on a high-calorie diet. Nature 2006, 444, 337–342. [Google Scholar] [CrossRef]
- Baur, J.A.; Sinclair, D.A. Therapeutic potential of resveratrol: The in vivo evidence. Nat. Rev. Drug Discov. 2006, 5, 493–506. [Google Scholar] [CrossRef]
- Cao, Z.; Li, Y. Potent induction of cellular antioxidants and phase 2 enzymes by resveratrol in cardiomyocytes: Protection against oxidative and electrophilic injury. Eur. J. Pharmacol. 2004, 489, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Saldanha, J.F.; Leal, V.D.O.; Stenvinkel, P.; Carraro-Eduardo, J.C.; Mafra, D. Resveratrol: Why Is It a Promising Therapy for Chronic Kidney Disease Patients? Oxid. Med. Cell. Longev. 2013, 2013, 963217. [Google Scholar] [CrossRef] [PubMed]
- Catalgol, B.; Batirel, S.; Taga, Y.; Ozer, N.K. Resveratrol: French Paradox Revisited. Front. Pharmacol. 2012, 3, 141. [Google Scholar] [CrossRef] [Green Version]
- Faber, A.C.; Dufort, F.J.; Blair, D.; Wagner, D.; Roberts, M.F.; Chiles, T.C. Inhibition of phosphatidylinositol 3-kinase-mediated glucose metabolism coincides with resveratrol-induced cell cycle arrest in human diffuse large B-cell lymphomas. Biochem. Pharmacol. 2006, 72, 1246–1256. [Google Scholar] [CrossRef] [PubMed]
- Gomez, L.S.; Zancan, P.; Marcondes, M.C.; Ramos-Santos, L.; Meyer-Fernandes, J.R.; Sola-Penna, M.; Dd Silva, D. Resveratrol decreases breast cancer cell viability and glucose metabolism by inhibiting 6-phosphofructo-1-kinase. Biochimie 2013, 95, 1336–1343. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, M.A.; Bamezai, R.N. Resveratrol Inhibits Cancer Cell Metabolism by Down Regulating Pyruvate Kinase M2 via Inhibition of Mammalian Target of Rapamycin. PLoS ONE 2012, 7, e36764. [Google Scholar] [CrossRef]
- Jung, K.-H.; Lee, J.H.; Quach, C.H.T.; Paik, J.-Y.; Oh, H.; Park, J.W.; Lee, E.J.; Moon, S.-H.; Lee, K.-H. Resveratrol Suppresses Cancer Cell Glucose Uptake by Targeting Reactive Oxygen Species-Mediated Hypoxia-Inducible Factor-1 Activation. J. Nucl. Med. 2013, 54, 2161–2167. [Google Scholar] [CrossRef] [Green Version]
- Kueck, A.; Opipari, A.W.; Griffith, K.A.; Tan, L.; Choi, M.; Huang, J.; Wahl, H.; Liu, J.R. Resveratrol inhibits glucose metabolism in human ovarian cancer cells. Gynecol. Oncol. 2007, 107, 450–457. [Google Scholar] [CrossRef]
- Almuhaideb, A.; Papathanasiou, N.; Bomanji, J. 18F-FDG PET/CT Imaging in Oncology. Ann. Saudi Med. 2011, 31, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Som, P.; Atkins, H.L.; Bandoypadhyay, D.; Fowler, J.S.; MacGregor, R.R.; Matsui, K.; Oster, Z.H.; Sacker, D.F.; Shiue, C.Y.; Turner, H.; et al. A fluorinated glucose analog, 2-fluoro-2-deoxy-D-glucose (F-18): Nontoxic tracer for rapid tumor detection. J. Nucl. Med. 1980, 21, 670–675. [Google Scholar] [CrossRef]
- Thompson, C.B. Rethinking the Regulation of Cellular Metabolism. Cold Spring Harb. Symp. Quant. Biol. 2011, 76, 23–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grassian, A.R.; Coloff, J.L.; Brugge, J.S. Extracellular Matrix Regulation of Metabolism and Implications for Tumorigenesis. Cold Spring Harb. Symp. Quant. Biol. 2011, 76, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Schafer, Z.T.; Grassian, A.R.; Song, L.; Jiang, Z.; Gerhart-Hines, Z.; Irie, H.Y.; Gao, S.; Puigserver, P.; Brugge, J.S. Antioxidant and oncogene rescue of metabolic defects caused by loss of matrix attachment. Nat. Cell Biol. 2009, 461, 109–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. The Hallmarks of Cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Vogelstein, B.; Kinzler, K.W. Cancer genes and the pathways they control. Nat. Med. 2004, 10, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Sreedhar, A.; Zhao, Y. Dysregulated metabolic enzymes and metabolicï ½reprogramming in cancer cells (Review). Biomed. Rep. 2017, 8, 3–10. [Google Scholar] [CrossRef]
- Eschbach, W. Ber den Stoffwechsel der Ektopie. Arch. Gynecol. Obstet. 1956, 188, 81–83. [Google Scholar] [CrossRef]
- Heiden, M.G.V.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg Effect: The Metabolic Requirements of Cell Proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liberti, M.V.; Locasale, J.W. The Warburg Effect: How Does it Benefit Cancer Cells? Trends Biochem. Sci. 2016, 41, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Fantin, V.R.; St-Pierre, J.; Leder, P. Attenuation of LDH-A expression uncovers a link between glycolysis, mitochondrial physiology, and tumor maintenance. Cancer Cell 2006, 9, 425–434. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Sánchez, R.; Rodríguez-Enríquez, S.; Marín-Hernández, A.; Saavedra, E. Energy metabolism in tumor cells. FEBS J. 2007, 274, 1393–1418. [Google Scholar] [CrossRef] [PubMed]
- Weinhouse, S. The Warburg hypothesis fifty years later. J. Cancer Res. Clin. Oncol. 1976, 87, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Zambrano, A.; Molt, M.; Uribe, E.; Salas, M. Glut 1 in Cancer Cells and the Inhibitory Action of Resveratrol as A Potential Therapeutic Strategy. Int. J. Mol. Sci. 2019, 20, 3374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niederacher, D.; Entian, K.-D. Characterization of Hex2 protein, a negative regulatory element necessary for glucose repression in yeast. JBIC J. Biol. Inorg. Chem. 1991, 200, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Herrero, P.; Galíndez, J.; Ruiz, N.; Martínez-Campa, C.; Moreno, F. Transcriptional regulation of the Saccharomyces cerevisiae HXK1, HXK2 andGLK1 genes. Yeast 1995, 11, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Christlieb, S.B.; Strandholdt, C.N.; Olsen, B.B.; Mylam, K.J.; Larsen, T.S.; Nielsen, A.L.; Rohde, M.; Gerke, O.; Olsen, K.E.; Moller, M.B.; et al. Dual time-point FDG PET/CT and FDG uptake and related enzymes in lymphadenopathies: Preliminary results. Eur. J. Nucl. Med. Mol. Imaging 2016, 43, 1824–1836. [Google Scholar] [CrossRef]
- Gu, J.J.; Singh, A.; Xue, K.; Mavis, C.; Barth, M.; Yanamadala, V.; Lenz, P.; Grau, M.; Lenz, G.; Czuczman, M.S.; et al. Up-regulation of hexokinase II contributes to rituximab-chemotherapy resistance and is a clinically relevant target for therapeutic development. Oncotarget 2017, 9, 4020–4033. [Google Scholar] [CrossRef] [Green Version]
- Rempel, A.; Mathupala, S.P.; Griffin, C.A.; Hawkins, A.L.; Pedersen, P.L. Glucose catabolism in cancer cells: Amplification of the gene encoding type II hexokinase. Cancer Res. 1996, 56, 2468–2471. [Google Scholar]
- Bustamante, E.; Pedersen, P.L. High aerobic glycolysis of rat hepatoma cells in culture: Role of mitochondrial hexokinase. Proc. Natl. Acad. Sci. USA 1977, 74, 3735–3739. [Google Scholar] [CrossRef] [Green Version]
- El-Bacha, T.; de Freitas, M.S.; Sola-Penna, M. Cellular distribution of phosphofructokinase activity and implications to metabolic regulation in human breast cancer. Mol. Genet. Metab. 2003, 79, 294–299. [Google Scholar] [CrossRef]
- Zancan, P.; Sola-Penna, M.; Furtado, C.M.; da Silva, D. Differential expression of phosphofructokinase-1 isoforms correlates with the glycolytic efficiency of breast cancer cells. Mol. Genet. Metab. 2010, 100, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Xiao, Z.; Chen, Z.; Zhang, X.; Li, J.; Wu, X.; Li, X.; Yi, H.; Li, M.; Zhu, G.; et al. Proteome analysis of human lung squamous carcinoma. Proteomics 2006, 6, 547–558. [Google Scholar] [CrossRef] [PubMed]
- Tokunaga, K.; Nakamura, Y.; Sakata, K.; Fujimori, K.; Ohkubo, M.; Sawada, K.; Sakiyama, S. Enhanced expression of a glyceraldehyde-3-phosphate dehydrogenase gene in human lung cancers. Cancer Res. 1987, 47, 5616–5619. [Google Scholar] [PubMed]
- Epner, D.E.; Partin, A.W.; Schalken, J.A.; Isaacs, J.T.; Coffey, D.S. Association of glyceraldehyde-3-phosphate dehydrogenase expression with cell motility and metastatic potential of rat prostatic adenocarcinoma. Cancer Res. 1993, 53, 1995–1997. [Google Scholar] [PubMed]
- Schek, N.; Hall, B.L.; Finn, O.J. Increased glyceraldehyde-3-phosphate dehydrogenase gene expression in human pancreatic adenocarcinoma. Cancer Res. 1988, 48, 6354–6359. [Google Scholar]
- Kudryavtseva, A.; Dmitriev, A.A.; Snezhkina, A.V.; Kudryavtseva, A.V. Deregulation of glycolysis in cancer: Glyceraldehyde-3-phosphate dehydrogenase as a therapeutic target. Expert Opin. Ther. Targets 2013, 17, 681–693. [Google Scholar] [CrossRef]
- Li, N.; Liu, X. Phosphoglycerate Mutase 1: Its Glycolytic and Non-Glycolytic Roles in Tumor Malignant Behaviors and Potential Therapeutic Significance. OncoTargets Ther. 2020, 13, 1787–1795. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Wu, C.; Yang, J.; Zhao, Y.; Jia, H.; Xue, M.; Xu, D.; Yang, F.; Fu, D.; Wang, C.; et al. Insulin-like growth factor 1-induced enolase 2 deacetylation by HDAC3 promotes metastasis of pancreatic cancer. Signal Transduct. Target. Ther. 2020, 5, 1–14. [Google Scholar] [CrossRef]
- Feng, C.; Gao, Y.; Wang, C.; Yu, X.; Zhang, W.; Guan, H.; Shan, Z.; Teng, W. Aberrant Overexpression of Pyruvate Kinase M2 Is Associated with Aggressive Tumor Features and the BRAF Mutation in Papillary Thyroid Cancer. J. Clin. Endocrinol. Metab. 2013, 98, 1524–1533. [Google Scholar] [CrossRef] [Green Version]
- Azoitei, N.; Becher, A.; Steinestel, K.; Rouhi, A.; Diepold, K.; Genze, F.; Simmet, T.; Seufferlein, T. PKM2 promotes tumor angiogenesis by regulating HIF-1α through NF-κB activation. Mol. Cancer 2016, 15, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Lu, W.; Cao, Y.; Zhang, Y.; Li, S.; Gao, J.; Wang, X.-A.; Mu, J.; Hu, Y.-P.; Jiang, L.; Dong, P.; et al. Up-regulation of PKM2 promote malignancy and related to adverse prognostic risk factor in human gallbladder cancer. Sci. Rep. 2016, 6, 26351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wittwer, J.A.; Robbins, D.; Wang, F.; Codarin, S.; Shen, X.; Kevil, C.G.; Huang, T.-T.; van Remmen, H.; Richardson, A.; Zhao, Y. Enhancing Mitochondrial Respiration Suppresses Tumor Promoter TPA-Induced PKM2 Expression and Cell Transformation in Skin Epidermal JB6 Cells. Cancer Prev. Res. 2011, 4, 1476–1484. [Google Scholar] [CrossRef] [Green Version]
- Le, A.; Cooper, C.R.; Gouw, A.M.; Dinavahi, R.; Maitra, A.; Deck, L.M.; Royer, R.E.; Jagt, D.L.V.; Semenza, G.L.; Dang, C.V. Inhibition of lactate dehydrogenase A induces oxidative stress and inhibits tumor progression. Proc. Natl. Acad. Sci. USA 2010, 107, 2037–2042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shim, H.; Chun, Y.S.; Lewis, B.C.; Dang, C.V. A unique glucose-dependent apoptotic pathway induced by c-Myc. Proc. Natl. Acad. Sci. USA 1998, 95, 1511–1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonas, S.K.; Benedetto, C.; Flatman, A.; Hammond, R.H.; Micheletti, L.; Riley, C.P.; Riley, P.A.; Spargo, D.J.; Zonca, M.; Slater, T.F. Increased activity of 6-phosphogluconate dehydrogenase and glucose-6-phosphate dehydrogenase in purified cell suspensions and single cells from the uterine cervix in cervical intraepithelial neoplasia. Br. J. Cancer 1992, 66, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Ahn, Y.S.; Chemeris, G.Y.; Turusov, V.S.; Bannasch, P. Enzymic Pattern of Preneoplastic and Neoplastic Lesions Induced in the Kidney of CBA Mice by 1,2-Dimethylhydrazine. Toxicol. Pathol. 1994, 22, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Lucarelli, G.; Galleggiante, V.; Rutigliano, M.; Sanguedolce, F.; Cagiano, S.; Bufo, P.; Lastilla, G.; Maiorano, E.; Ribatti, D.; Giglio, A.; et al. Metabolomic profile of glycolysis and the pentose phosphate pathway identifies the central role of glucose-6-phosphate dehydrogenase in clear cell-renal cell carcinoma. Oncotarget 2015, 6, 13371–13386. [Google Scholar] [CrossRef] [Green Version]
- D’Alessandro, A.; Amelio, I.; Berkers, C.R.; Antonov, A.; Vousden, K.H.; Melino, G.; Zolla, L. Metabolic effect of TAp63α: Enhanced glycolysis and pentose phosphate pathway, resulting in increased antioxidant defense. Oncotarget 2014, 5, 7722–7733. [Google Scholar] [CrossRef] [Green Version]
- Sukhatme, V.P.; Chan, B. Glycolytic cancer cells lacking 6-phosphogluconate dehydrogenase metabolize glucose to induce senescence. FEBS Lett. 2012, 586, 2389–2395. [Google Scholar] [CrossRef] [Green Version]
- Patra, K.C.; Hay, N. The pentose phosphate pathway and cancer. Trends Biochem. Sci. 2014, 39, 347–354. [Google Scholar] [CrossRef] [Green Version]
- Gandhi, N.; Das, G.M. Metabolic Reprogramming in Breast Cancer and Its Therapeutic Implications. Cells 2019, 8, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epstein, T.; Xu, L.; Gillies, R.J.; Gatenby, R. Separation of metabolic supply and demand: Aerobic glycolysis as a normal physiological response to fluctuating energetic demands in the membrane. Cancer Metab. 2014, 2, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shamim, U.; Hanif, S.; Albanyan, A.; Beck, F.W.J.; Bao, B.; Wang, Z.; Banerjee, S.; Sarkar, F.H.; Mohammad, R.M.; Hadi, S.M.; et al. Resveratrol-induced apoptosis is enhanced in low pH environments associated with cancer. J. Cell. Physiol. 2011, 227, 1493–1500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parkins, C.; Stratford, M.; Dennis, M.; Stubbs, M.; Chaplin, D. The relationship between extracellular lactate and tumour pH in a murine tumour model of ischaemia-reperfusion. Br. J. Cancer 1997, 75, 319–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerweck, L.E.; Vijayappa, S.; Kozin, S. Tumor pH controls the in vivo efficacy of weak acid and base chemotherapeutics. Mol. Cancer Ther. 2006, 5, 1275–1279. [Google Scholar] [CrossRef] [Green Version]
- Gerweck, L.E. The pH difference between tumor and normal tissue offers a tumor specific target for the treatment of cancer. Drug Resist. Updat. 2000, 3, 49–50. [Google Scholar] [CrossRef]
- Kozin, S.V.; Shkarin, P.; Gerweck, L.E. The cell transmembrane pH gradient in tumors enhances cytotoxicity of specific weak acid chemotherapeutics. Cancer Res. 2001, 61, 4740–4743. [Google Scholar]
- Gerweck, L.E. Tumor pH: Implications for treatment and novel drug design. Semin. Radiat. Oncol. 1998, 8, 176–182. [Google Scholar] [CrossRef]
- Barry, M.A.; Eastman, A. Endonuclease activation during apoptosis: The role of cytosolic Ca2+ and pH. Biochem. Biophys. Res. Commun. 1992, 186, 782–789. [Google Scholar] [CrossRef]
- Chen, R.; Lin, J.; Yan, W.; Chen, D. miR-522-3p Promotes Osteosarcoma Cell Growth by Regulating Glucose Uptake and GLUT1 Expression. OncoTargets Ther. 2019, 12, 9053–9058. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Song, Y.; Yu, Y.; Cheng, W.; Tong, X. miRNA-10a promotes cancer cell proliferation in oral squamous cell carcinoma by upregulating GLUT1 and promoting glucose metabolism. Oncol. Lett. 2019, 17, 5441–5446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, G.; Zhao, Y.; Wu, D.; Gao, C. Mir-150 Up-Regulates Glut1 and Increases Glycolysis in Osteosarcoma Cells. Asian Pac. J. Cancer Prev. 2017, 18, 1127–1131. [Google Scholar] [PubMed]
- Gregersen, L.H.; Skanderup, A.J.; Frankel, L.B.; Wen, J.; Krogh, A.; Lund, A.H. MicroRNA-143 down-regulates Hexokinase 2 in colon cancer cells. BMC Cancer 2012, 12, 232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, X.; Yao, L.; Zhang, J.; Han, P.; Li, C. Inhibition of microRNA-155 sensitizes lung cancer cells to irradiation via suppression of HK2-modulated glucose metabolism. Mol. Med. Rep. 2016, 14, 1332–1338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, G.; Sun, W.; Shen, Y.; Hu, Y.; Liu, H.; Li, W.; Wang, Y. PKM2 functions as a potential oncogene and is a crucial target of miR-148a and miR-326 in thyroid tumorigenesis. Am. J. Transl. Res. 2018, 10, 1793–1801. [Google Scholar]
- Guo, M.; Zhao, X.; Yuan, X.; Jiang, J.; Li, P. MiR-let-7a inhibits cell proliferation, migration, and invasion by down-regulating PKM2 in cervical cancer. Oncotarget 2017, 8, 28226–28236. [Google Scholar] [CrossRef] [Green Version]
- Belguendouz, L.; Frémont, L.; Gozzelino, M.-T. Interaction of Transresveratrol with Plasma Lipoproteins. Biochem. Pharmacol. 1998, 55, 811–816. [Google Scholar] [CrossRef]
- Fontecave, M.; Lepoivre, M.; Elleingand, E.; Gerez, C.; Guittet, O. Resveratrol, a remarkable inhibitor of ribonucleotide reductase. FEBS Lett. 1998, 421, 277–279. [Google Scholar] [CrossRef] [Green Version]
- Gehm, B.D.; McAndrews, J.M.; Chien, P.-Y.; Jameson, J.L. Resveratrol, a polyphenolic compound found in grapes and wine, is an agonist for the estrogen receptor. Proc. Natl. Acad. Sci. USA 1997, 94, 14138–14143. [Google Scholar] [CrossRef] [Green Version]
- Orsini, F.; Pelizzoni, F.; Verotta, L.; Aburjai, T.; Rogers, C.B. Isolation, Synthesis, and Antiplatelet Aggregation Activity of Resveratrol 3-O-β-d-Glucopyranoside and Related Compounds. J. Nat. Prod. 1997, 60, 1082–1087. [Google Scholar] [CrossRef]
- Uenobe, F.; Nakamura, S.-I.; Miyazawa, M. Antimutagenic effect of resveratrol against Trp-P-1. Mutat. Res. Mol. Mech. Mutagen. 1997, 373, 197–200. [Google Scholar] [CrossRef]
- Kolling, J.; Kolling, J.; de Franceschi, I.D.; Nishihira, V.S.; Baldissera, M.D.; Pinto, C.G.; Mezzomo, N.J.; Carmo, G.M.D.; Feksa, L.R.; Fernandes, L.S.; et al. Resveratrol and resveratrol-hydroxypropyl-β-cyclodextrin complex recovered the changes of creatine kinase and Na+, K+-ATPase activities found in the spleen from streptozotocin-induced diabetic rats. Anais Acad. Brasil. Ciênc. 2019, 91, e20181330. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, Y.; Mahmoudi, N.; Yousefi, B.; Karimian, A. The effects of statins with a high hepatoselectivity rank on the extra-hepatic tissues; New functions for statins. Pharmacol. Res. 2020, 152, 104621. [Google Scholar] [CrossRef] [PubMed]
- Hoseini, A.; Namazi, G.; Farrokhian, A.; Reiner, Ž.; Aghadavod, E.; Bahmani, F.; Asemi, Z. The effects of resveratrol on metabolic status in patients with type 2 diabetes mellitus and coronary heart disease. Food Funct. 2019, 10, 6042–6051. [Google Scholar] [CrossRef]
- Bertelli, A.A.; Giovannini, L.; Giannessi, D.; Migliori, M.; Bernini, W.; Fregoni, M. Antiplatelet activity of synthetic and natural resveratrol in red wine. Int. J. Tissue React. 1995, 17, 1–3. [Google Scholar]
- Pace-Asciak, C.R.; Hahn, S.; Diamandis, E.P.; Soleas, G.; Goldberg, D.M. The red wine phenolics trans-resveratrol and quercetin block human platelet aggregation and eicosanoid synthesis: Implications for protection against coronary heart disease. Clin. Chim. Acta 1995, 235, 207–219. [Google Scholar] [CrossRef]
- Wu, C.W.; Nakamoto, Y.; Hisatome, T.; Yoshida, S.; Miyazaki, H. Resveratrol and its dimers ε-viniferin and δ-viniferin in red wine protect vascular endothelial cells by a similar mechanism with different potency and efficacy. Kaohsiung J. Med Sci. 2020, 36, 535–542. [Google Scholar] [CrossRef] [Green Version]
- Paul, B.; Masih, I.; Deopujari, J.; Charpentier, C. Occurrence of resveratrol and pterostilbene in age-old darakchasava, an ayurvedic medicine from India. J. Ethnopharmacol. 1999, 68, 71–76. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Z.; Fang, A.; Wu, K.; Chen, X.; Wang, G.; Mao, F. Resveratrol Attenuates Inflammatory Bowel Disease in Mice by Regulating SUMO1. Biol. Pharm. Bull. 2020, 43, 450–457. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Wang, Z.; Qiu, Y.; Liu, Y.; Ding, M.; Zhang, Y. Anti-miRNA21 and resveratrol-loaded polysaccharide-based mesoporous silica nanoparticle for synergistic activity in gastric carcinoma. J. Drug Target. 2019, 27, 1135–1143. [Google Scholar] [CrossRef]
- Sareen, D.; Darjatmoko, S.R.; Albert, D.M.; Polans, A.S. Mitochondria, Calcium, and Calpain are Key Mediators of Resveratrol-Induced Apoptosis in Breast Cancer. Mol. Pharmacol. 2007, 72, 1466–1475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scarlatti, F.; Sala, G.; Somenzi, G.; Signorelli, P.; Sacchi, N.; Ghidoni, R. Resveratrol induces growth inhibition and apoptosis in metastatic breast cancer cells via de novo ceramide signaling. FASEB J. 2003, 17, 2339–2341. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Ma, X.; Li, N.; Liu, H.; Dong, Q.; Zhang, J.; Yang, C.; Liu, Y.; Liang, Q.; Zhang, S.; et al. Resveratrol inhibits Hexokinases II mediated glycolysis in non-small cell lung cancer via targeting Akt signaling pathway. Exp. Cell Res. 2016, 349, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Dasari, S.K.; Bialik, S.; Levin-Zaidman, S.; Levin-Salomon, V.; Futerman, A.H.; Kimchi, A. Signalome-wide RNAi screen identifies GBA1 as a positive mediator of autophagic cell death. Cell Death Differ. 2017, 24, 1288–1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mollerup, S.; Ovrebø, S.; Haugen, A. Lung carcinogenesis: Resveratrol modulates the expression of genes involved in the metabolism of PAH in human bronchial epithelial cells. Int. J. Cancer 2001, 92, 18–25. [Google Scholar] [CrossRef]
- Fouad, M.A.; Agha, A.M.; Al Merzabani, M.M.; Shouman, S.A. Resveratrol inhibits proliferation, angiogenesis and induces apoptosis in colon cancer cells. Hum. Exp. Toxicol. 2013, 32, 1067–1080. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.-H.; Lee, J.H.; Park, J.W.; Quach, C.H.T.; Moon, S.H.; Cho, Y.-S.; Lee, K.-H. Resveratrol-loaded polymeric nanoparticles suppress glucose metabolism and tumor growth in vitro and in vivo. Int. J. Pharm. 2015, 478, 251–257. [Google Scholar] [CrossRef]
- Saunier, E.; Antonio, S.; Regazzetti, A.; Auzeil, N.; Laprévote, O.; Shay, J.W.; Coumoul, X.; Barouki, R.; Benelli, C.; Huc-Lemarié, L.; et al. Resveratrol reverses the Warburg effect by targeting the pyruvate dehydrogenase complex in colon cancer cells. Sci. Rep. 2017, 7, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Vanamala, J.; Radhakrishnan, S.; Reddivari, L.; Bhat, V.B.; Ptitsyn, A. Resveratrol suppresses human colon cancer cell proliferation and induces apoptosis via targeting the pentose phosphate and the talin-FAK signaling pathways-A proteomic approach. Proteome Sci. 2011, 9, 49. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, J.; Moradi, F.; Maddalena, L.A.; Ferreira-Tollstadius, B.; Selim, S.; Stuart, J.A. Resveratrol integrates metabolic and growth effects in PC3 prostate cancer cells-involvement of prolyl hydroxylase and hypoxia inducible factor-1. Oncol. Lett. 2018, 17, 697–705. [Google Scholar] [CrossRef]
- Fonseca, J.; Moradi, F.; Valente, A.J.F.; Stuart, J.A. Oxygen and Glucose Levels in Cell Culture Media Determine Resveratrol’s Effects on Growth, Hydrogen Peroxide Production, and Mitochondrial Dynamics. Antioxidants 2018, 7, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gwak, H.; Haegeman, G.; Tsang, B.K.; Song, Y.S. Cancer-specific interruption of glucose metabolism by resveratrol is mediated through inhibition of Akt/GLUT1 axis in ovarian cancer cells. Mol. Carcinog. 2014, 54, 1529–1540. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tong, L.; Luo, Y.; Li, X.; Chen, G.; Wang, Y. Resveratrol inhibits the proliferation and induces the apoptosis in ovarian cancer cells via inhibiting glycolysis and targeting AMPK/mTOR signaling pathway. J. Cell. Biochem. 2018, 119, 6162–6172. [Google Scholar] [CrossRef]
- Yang, S.H.; Kim, J.S.; Oh, T.J.; Kim, M.S.; Lee, S.W.; Woo, S.K.; Cho, H.S.; Choi, Y.H.; Kim, Y.H.; Rha, S.Y.; et al. Genome-scale analysis of resveratrol-induced gene expression profile in human ovarian cancer cells using a cDNA microarray. Int. J. Oncol. 2003, 22, 741–750. [Google Scholar] [CrossRef]
- Billard, C.; Izard, J.-C.; Roman, V.; Kern, C.; Mathiot, C.; Mentz, F.; Kolb, J.-P. Comparative Antiproliferative and Apoptotic Effects of Resveratrol, ϵ-viniferin and Vine-shots Derived Polyphenols (Vineatrols) on Chronic B Lymphocytic Leukemia Cells and Normal Human Lymphocytes. Leuk. Lymphoma 2002, 43, 1991–2002. [Google Scholar] [CrossRef]
- Roman, V.; Billard, C.; Kern, C.; Ferry-Dumazet, H.; Izard, J.-C.; Mohammad, R.; Mossalayi, D.M.; Kolb, J.-P. Analysis of resveratrol-induced apoptosis in human B-cell chronic leukaemia. Br. J. Haematol. 2002, 117, 842–851. [Google Scholar] [CrossRef]
- Dörrie, J.; Gerauer, H.; Wachter, Y.; Zunino, S.J. Resveratrol induces extensive apoptosis by depolarizing mitochondrial membranes and activating caspase-9 in acute lymphoblastic leukemia cells. Cancer Res. 2001, 61, 4731–4739. [Google Scholar] [PubMed]
- Li, Y.; Zhu, W.; Li, J.; Liu, M.; Wei, M. Resveratrol suppresses the STAT3 signaling pathway and inhibits proliferation of high glucose-exposed HepG2 cells partly through SIRT1. Oncol. Rep. 2013, 30, 2820–2828. [Google Scholar] [CrossRef] [PubMed]
- Chai, R.; Fu, H.; Zheng, Z.; Liu, T.; Ji, S.; Li, G. Resveratrol inhibits proliferation and migration through SIRT1 mediated post-translational modification of PI3K/AKT signaling in hepatocellular carcinoma cells. Mol. Med. Rep. 2017, 16, 8037–8044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, L.; Chen, X.; Xiao, X.; Ma, Q.; Li, W. Resveratrol inhibits hyperglycemia-driven ROS-induced invasion and migration of pancreatic cancer cells via suppression of the ERK and p38 MAPK signaling pathways. Int. J. Oncol. 2016, 49, 735–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivani, G.; Behera, S.K.; Dariya, B.; Aliya, S.; Alam, A.; Nagaraju, G.P. Resveratrol binds and inhibits transcription factor HIF-1α in pancreatic cancer. Exp. Cell Res. 2020, 394, 112126. [Google Scholar] [CrossRef] [PubMed]
- Yan, B.; Cheng, L.; Jiang, Z.; Chen, K.; Zhou, C.; Sun, L.; Cao, J.; Qian, W.; Li, J.; Shan, T.; et al. Resveratrol Inhibits ROS-Promoted Activation and Glycolysis of Pancreatic Stellate Cells via Suppression of miR-21. Oxidative Med. Cell. Longev. 2018, 2018, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Lowry, S.F.; Guarente, L.; Haimovich, B. Roles of SIRT1 in the Acute and Restorative Phases following Induction of Inflammation. J. Biol. Chem. 2010, 285, 41391–41401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, L.; Tan, S.; Zhou, Y.; Lin, J.; Wang, H.; Oyang, L.; Tian, Y.; Liu, L.; Su, M.; Wang, H.; et al. Role of the NFκB-signaling pathway in cancer. OncoTargets Ther. 2018, 11, 2063–2073. [Google Scholar] [CrossRef] [Green Version]
- Inoue, J.-I.; Gohda, J.; Akiyama, T.; Semba, K. NF-B activation in development and progression of cancer. Cancer Sci. 2007, 98, 268–274. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Ralhan, R. Nuclear factor-kappa B links carcinogenic and chemopreventive agents. Front. Biosci. 2009, 1, 45–60. [Google Scholar] [CrossRef]
- Buhrmann, C.; Yazdi, M.; Popper, B.; Shayan, P.; Goel, A.; Aggarwal, B.B.; Shakibaei, M. Resveratrol Chemosensitizes TNF-β-Induced Survival of 5-FU-Treated Colorectal Cancer Cells. Nutrients 2018, 10, 888. [Google Scholar] [CrossRef] [Green Version]
- Buhrmann, C.; Shayan, P.; Goel, A.; Shakibaei, M. Resveratrol Regulates Colorectal Cancer Cell Invasion by Modulation of Focal Adhesion Molecules. Nutrients 2017, 9, 1073. [Google Scholar] [CrossRef]
- Buhrmann, C.; Shayan, P.; Popper, B.; Goel, A.; Shakibaei, M. Sirt1 Is Required for Resveratrol-Mediated Chemopreventive Effects in Colorectal Cancer Cells. Nutrients 2016, 8, 145. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.-L.; Sun, Z.-J.; Yu, L.; Meng, K.-W.; Qin, X.-L.; Pan, C.-E. Effect of resveratrol and in combination with 5-FU on murine liver cancer. World J. Gastroenterol. 2004, 10, 3048–3052. [Google Scholar] [CrossRef] [PubMed]
- Kauppinen, A.; Suuronen, T.; Ojala, J.; Kaarniranta, K.; Salminen, A. Antagonistic crosstalk between NF-κB and SIRT1 in the regulation of inflammation and metabolic disorders. Cell. Signal. 2013, 25, 1939–1948. [Google Scholar] [CrossRef] [PubMed]
- Frazzi, R.; Valli, R.; Tamagnini, I.; Casali, B.; Latruffe, N.; Merli, F. Resveratrol-mediated apoptosis of hodgkin lymphoma cells involves SIRT1 inhibition and FOXO3a hyperacetylation. Int. J. Cancer 2012, 132, 1013–1021. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, S.; Loitsch, S.M.; Rau, O.; von Knethen, A.; Brüne, B.; Schubert-Zsilavecz, M.; Stein, J.M. Peroxisome Proliferator–Activated Receptor γ as a Molecular Target of Resveratrol-Induced Modulation of Polyamine Metabolism. Cancer Res. 2006, 66, 7348–7354. [Google Scholar] [CrossRef] [Green Version]
- Araújo, J.; Gonçalves, P.; Martel, F. Modulation of Glucose Uptake in a Human Choriocarcinoma Cell Line (BeWo) by Dietary Bioactive Compounds and Drugs of Abuse. J. Biochem. 2008, 144, 177–186. [Google Scholar] [CrossRef]
- Azevedo, C.; Correia-Branco, A.; Araújo, J.; Guimarães, J.T.; Keating, E.; Martel, F. The Chemopreventive Effect of the Dietary Compound Kaempferol on the MCF-7 Human Breast Cancer Cell Line Is Dependent on Inhibition of Glucose Cellular Uptake. Nutr. Cancer 2015, 67, 504–513. [Google Scholar] [CrossRef]
- George, J.; Nihal, M.; Singh, C.K.; Ahmad, N. 4′-Bromo-resveratrol, a dual Sirtuin-1 and Sirtuin-3 inhibitor, inhibits melanoma cell growth through mitochondrial metabolic reprogramming. Mol. Carcinog. 2019, 58, 1876–1885. [Google Scholar] [CrossRef]
- Graham, R.M.; Hernández, F.; Puerta, N.; de Angulo, G.; A Webster, K.; Vanni, S. Resveratrol augments ER stress and the cytotoxic effects of glycolytic inhibition in neuroblastoma by downregulating Akt in a mechanism independent of SIRT1. Exp. Mol. Med. 2016, 48, e210. [Google Scholar] [CrossRef] [Green Version]
- Gwak, H.; Kim, S.; Dhanasekaran, D.N.; Song, Y.S. Resveratrol triggers ER stress-mediated apoptosis by disrupting N-linked glycosylation of proteins in ovarian cancer cells. Cancer Lett. 2016, 371, 347–353. [Google Scholar] [CrossRef]
- Huang, T.-T.; Lin, H.-C.; Chen, C.-C.; Lu, C.-C.; Wei, C.-F.; Wu, T.-S.; Liu, F.-G.; Lai, H.-C. Resveratrol induces apoptosis of human nasopharyngeal carcinoma cells via activation of multiple apoptotic pathways. J. Cell. Physiol. 2010, 226, 720–728. [Google Scholar] [CrossRef]
- León, D.; Uribe, E.; Zambrano, A.; Salas, M. Implications of Resveratrol on Glucose Uptake and Metabolism. Molecules 2017, 22, 398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, C.E.; Kim, M.-J.; Lee, J.H.; Min, B.-I.; Bae, H.; Choe, W.; Kim, S.-S.; Ha, J. Resveratrol stimulates glucose transport in C2C12 myotubes by activating AMP-activated protein kinase. Exp. Mol. Med. 2007, 39, 222–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poulsen, M.M.; Jørgensen, J.O.L.; Jessen, N.; Richelsen, B.; Pedersen, S.B. Resveratrol in metabolic health: An overview of the current evidence and perspectives. Ann. New York Acad. Sci. 2013, 1290, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Salas, M.; Obando, P.; Ojeda, L.; Ojeda, P.; Pérez, A.; Vargas-Uribe, M.; Rivas, C.I.; Vera, J.C.; Reyes, A.M. Resolution of the direct interaction with and inhibition of the human GLUT1 hexose transporter by resveratrol from its effect on glucose accumulation. Am. J. Physiol. Physiol. 2013, 305, C90–C99. [Google Scholar] [CrossRef] [Green Version]
- Tan, L.; Wang, W.; He, G.; Kuick, R.D.; Gossner, G.; Kueck, A.S.; Wahl, H.; Opipari, A.W.; Liu, J.R. Resveratrol inhibits ovarian tumor growth in an in vivo mouse model. Cancer 2015, 122, 722–729. [Google Scholar] [CrossRef] [PubMed]
- Park, D.J.; Lenz, H.-J. Determinants of chemosensitivity in gastric cancer. Curr. Opin. Pharmacol. 2006, 6, 337–344. [Google Scholar] [CrossRef]
- Buhrmann, C.; Shayan, P.; Kraehe, P.; Popper, B.; Goel, A.; Shakibaei, M. Resveratrol induces chemosensitization to 5-fluorouracil through up-regulation of intercellular junctions, Epithelial-to-mesenchymal transition and apoptosis in colorectal cancer. Biochem. Pharmacol. 2015, 98, 51–68. [Google Scholar] [CrossRef]
- Cheng, Y.-J.; Chang, M.-Y.; Chang, W.-W.; Wang, W.-K.; Liu, C.-F.; Lin, S.-T.; Lee, C.-H. Resveratrol Enhances Chemosensitivity in Mouse Melanoma Model Through Connexin 43 Upregulation. Environ. Toxicol. 2015, 30, 877–886. [Google Scholar] [CrossRef]
- Jie, K.Y.; Wei, C.L.; Min, Z.; Ping, G.J.; Ying, W.; Dan, Z.; Sen, Z. Resveratrol enhances chemosensitivity of renal cell carcinoma to paclitaxel. Front. Biosci. 2019, 24, 1452–1461. [Google Scholar]
- Li, Y.; Yang, Y.; Liu, X.; Long, Y.; Zheng, Y. PRMT5 Promotes Human Lung Cancer Cell Apoptosis via Akt/Gsk3β Signaling Induced by Resveratrol. Cell Transplant. 2019, 28, 1664–1673. [Google Scholar] [CrossRef]
- Vinod, B.S.; Nair, H.H.; Vijayakurup, V.; Shabna, A.; Shah, S.; Krishna, A.; Pillai, K.S.; Thankachan, S.; Anto, R.J. Resveratrol chemosensitizes HER-2-overexpressing breast cancer cells to docetaxel chemoresistance by inhibiting docetaxel-mediated activation of HER-2–Akt axis. Cell Death Discov. 2015, 1, 15061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, C.; Qian, W.; Ma, J.; Cheng, L.; Jiang, Z.; Yan, B.; Li, J.; Duan, W.; Sun, L.; Cao, J.; et al. Resveratrol enhances the chemotherapeutic response and reverses the stemness induced by gemcitabine in pancreatic cancer cells via targeting SREBP1. Cell Prolif. 2019, 52, e12514. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Chen, C.; Jiang, X.; Zhang, Z. ROS-mediated endoplasmic reticulum stress and mitochondrial dysfunction underlie apoptosis induced by resveratrol and arsenic trioxide in A549 cells. Chem. Interact. 2016, 245, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-W.; Woo, K.J.; Lee, J.-T.; Lim, J.H.; Lee, T.; Kim, S.H.; Choi, Y.H.; Kwon, T.K. Resveratrol induces pro-apoptotic endoplasmic reticulum stress in human colon cancer cells. Oncol. Rep. 2007, 18, 1269–1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arafa, E.-S.A.; Abdelazeem, A.H.; Arab, H.H.; Omar, H.A. OSU-CG5, a novel energy restriction mimetic agent, targets human colorectal cancer cells in vitro. Acta Pharmacol. Sin. 2014, 35, 394–400. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Gao, Z.; Zhang, X. Resveratrol Induces Apoptosis in Murine Prostate Cancer Cells via Hypoxia-Inducible Factor 1-alpha (HIF-1α)/Reactive Oxygen Species (ROS)/P53 Signaling. Med. Sci. Monit. 2018, 24, 8970–8976. [Google Scholar] [CrossRef]
- Mitani, T.; Harada, N.; Tanimori, S.; Nakano, Y.; Inui, H.; Yamaji, R. Resveratrol inhibits hypoxia-inducible factor-1α-mediated androgen receptor signaling and represses tumor progression in castration-resistant prostate cancer. J. Nutr. Sci. Vitaminol. 2014, 60, 276–282. [Google Scholar] [CrossRef] [Green Version]
- Varghese, E.; Samuel, S.M.; Liskova, A.; Samec, M.; Kubatka, P.; Büsselberg, D. Targeting Glucose Metabolism to Overcome Resistance to Anticancer Chemotherapy in Breast Cancer. Cancers 2020, 12, 2252. [Google Scholar] [CrossRef]
- Barron, C.C.; Bilan, P.J.; Tsakiridis, T.; Tsiani, E. Facilitative glucose transporters: Implications for cancer detection, prognosis and treatment. Metabolism 2016, 65, 124–139. [Google Scholar] [CrossRef]
- Labak, C.M.; Wang, P.Y.; Arora, R.; Guda, M.R.; Asuthkar, S.; Tsung, A.J.; Velpula, K.K. Glucose transport: Meeting the metabolic demands of cancer, and applications in glioblastoma treatment. Am. J. Cancer Res. 2016, 6, 1599–1608. [Google Scholar]
- Massari, F.; Ciccarese, C.; Santoni, M.; Iacovelli, R.; Mazzucchelli, R.; Piva, F.; Scarpelli, M.; Berardi, R.; Tortora, G.; Lopez-Beltran, A.; et al. Metabolic phenotype of bladder cancer. Cancer Treat. Rev. 2016, 45, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Szablewski, L. Expression of glucose transporters in cancers. Biochim. Biophys. Acta Bioenerg. 2013, 1835, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Demarco, V.G.; Nicholl, M.B. Resveratrol enhances radiation sensitivity in prostate cancer by inhibiting cell proliferation and promoting cell senescence and apoptosis. Cancer Sci. 2012, 103, 1090–1098. [Google Scholar] [CrossRef] [PubMed]
- Riles, W.L.; Erickson, J.; Nayyar, S.; Atten, M.J.; Attar, B.M.; Holian, O. Resveratrol engages selective apoptotic signals in gastric adenocarcinoma cells. World J. Gastroenterol. 2006, 12, 5628–5634. [Google Scholar] [CrossRef]
- Opipari, A.W.; Tan, L.; Boitano, A.E.; Sorenson, D.R.; Aurora, A.; Liu, J.R. Resveratrol-induced Autophagocytosis in Ovarian Cancer Cells. Cancer Res. 2004, 64, 696–703. [Google Scholar] [CrossRef] [Green Version]
- Varoni, E.M.; lo Faro, A.F.; Sharifi-Rad, J.; Iriti, M. Anticancer Molecular Mechanisms of Resveratrol. Front. Nutr. 2016, 3, 8. [Google Scholar] [CrossRef] [Green Version]
- Hoxhaj, G.; Manning, B.D. The PI3K–AKT network at the interface of oncogenic signalling and cancer metabolism. Nat. Rev. Cancer 2019, 20, 74–88. [Google Scholar] [CrossRef]
- Li, M.; Jin, R.; Wang, W.; Zhang, T.; Sang, J.; Li, N.; Han, Q.; Zhao, W.; Li, C.; Liu, Z. STAT3 regulates glycolysis via targeting hexokinase 2 in hepatocellular carcinoma cells. Oncotarget 2017, 8, 24777–24784. [Google Scholar] [CrossRef] [Green Version]
- Sexton, É.; van Themsche, C.; Leblanc, K.; Parent, S.; Lemoine, P.; Asselin, E. Resveratrol interferes with AKT activity and triggers apoptosis in human uterine cancer cells. Mol. Cancer 2006, 5, 45. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.-N.; Lin, V.C.-H.; Rau, K.-M.; Shieh, P.-C.; Kuo, D.-H.; Shieh, J.-C.; Chen, W.J.; Tsai, S.-C.; Way, T.-D. Resveratrol Modulates Tumor Cell Proliferation and Protein Translation via SIRT1-Dependent AMPK Activation. J. Agric. Food Chem. 2010, 58, 1584–1592. [Google Scholar] [CrossRef]
- Dai, W.; Wang, F.; Lu, J.; Xia, Y.; He, L.; Chen, K.; Li, J.; Li, S.; Liu, T.; Zheng, Y.; et al. By reducing hexokinase 2, resveratrol induces apoptosis in HCC cells addicted to aerobic glycolysis and inhibits tumor growth in mice. Oncotarget 2015, 6, 13703–13717. [Google Scholar] [CrossRef] [Green Version]
- Narayanan, N.K.; Narayanan, B.A.; Nixon, D.W. Resveratrol-induced cell growth inhibition and apoptosis is associated with modulation of phosphoglycerate mutase B in human prostate cancer cells: Two-dimensional sodium dodecyl sulfate-polyacrylamide gel electrophoresis and mass spectrometry evaluation. Cancer Detect. Prev. 2004, 28, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Sicard, F.; Gayral, M.; Lulka, H.; Buscail, L.; Cordelier, P. Targeting miR-21 for the Therapy of Pancreatic Cancer. Mol. Ther. 2013, 21, 986–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peralta-Zaragoza, O.; Deas, J.; Meneses-Acosta, A.; de la O-Gómez, F.; Fernández-Tilapa, G.; Gómez-Cerón, C.; Benítez-Boijseauneau, O.; Burguete-García, A.; Torres-Poveda, K.; Bermúdez-Morales, V.H.; et al. Relevance of miR-21 in regulation of tumor suppressor gene PTEN in human cervical cancer cells. BMC Cancer 2016, 16, 215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Wang, Y.; Wu, C.; Yang, P.; Li, H.; Li, Z. Resveratrol Induces Cancer Cell Apoptosis through MiR-326/PKM2-Mediated ER Stress and Mitochondrial Fission. J. Agric. Food Chem. 2016, 64, 9356–9367. [Google Scholar] [CrossRef] [PubMed]
- Thiel, G.; Rössler, O.G. Resveratrol regulates gene transcription via activation of stimulus-responsive transcription factors. Pharmacol. Res. 2017, 117, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Suh, D.H.; Kim, M.K.; Kim, H.S.; Chung, H.H.; Song, Y.S. Cancer-specific Therapeutic Potential of Resveratrol: Metabolic Approach against Hallmarks of Cancer. Funct. Foods Health Dis. 2013, 3, 332. [Google Scholar] [CrossRef]
- Song, J.; Yang, B.; Jia, X.; Li, M.; Tan, W.; Ma, S.; Shi, X.; Feng, L. Distinctive Roles of Sirtuins on Diabetes, Protective or Detrimental? Front. Endocrinol. 2018, 9, 724. [Google Scholar] [CrossRef] [PubMed]
- Howitz, K.T.; Bitterman, K.J.; Cohen, H.Y.; Lamming, D.W.; Lavu, S.; Wood, J.G.; Zipkin, R.E.; Chung, P.; Kisielewski, A.; Zhang, L.-L.; et al. Small molecule activators of sirtuins extend Saccharomyces cerevisiae lifespan. Nature 2003, 425, 191–196. [Google Scholar] [CrossRef]
- Wood, J.G.; Rogina, B.; Lavu, S.; Howitz, K.; Helfand, S.L.; Tatar, M.; Sinclair, D. Sirtuin activators mimic caloric restriction and delay ageing in metazoans. Nat. Cell Biol. 2004, 430, 686–689. [Google Scholar] [CrossRef]
- Gaál, Z.; Csernoch, L. Impact of Sirtuin Enzymes on the Altered Metabolic Phenotype of Malignantly Transformed Cells. Front. Oncol. 2020, 10, 45. [Google Scholar] [CrossRef] [PubMed]
- Kaeberlein, M.; McVey, M.; Guarente, L. The SIR2/3/4 complex and SIR2 alone promote longevity in Saccharomyces cerevisiae by two different mechanisms. Genes Dev. 1999, 13, 2570–2580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imai, S.-I.; Armstrong, C.M.; Kaeberlein, M.; Guarente, L. Transcriptional silencing and longevity protein Sir2 is an NAD-dependent histone deacetylase. Nat. Cell Biol. 2000, 403, 795–800. [Google Scholar] [CrossRef] [PubMed]
- Carafa, V.; Altucci, L.; Nebbioso, A. Dual Tumor Suppressor and Tumor Promoter Action of Sirtuins in Determining Malignant Phenotype. Front. Pharmacol. 2019, 10, 38. [Google Scholar] [CrossRef] [Green Version]
- Lagouge, M.; Argmann, C.; Gerhart-Hines, Z.; Meziane, H.; Lerin, C.; Daussin, F.; Messadeq, N.; Milne, J.; Lambert, P.; Elliott, P.; et al. Resveratrol Improves Mitochondrial Function and Protects against Metabolic Disease by Activating SIRT1 and PGC-1α. Cell 2006, 127, 1109–1122. [Google Scholar] [CrossRef]
- Dembic, M.; Andersen, H.S.; Bastin, J.; Doktor, T.K.; Corydon, T.J.; Sass, J.O.; Costa, A.L.; Djouadi, F.; Andresen, B.S. Next generation sequencing of RNA reveals novel targets of resveratrol with possible implications for Canavan disease. Mol. Genet. Metab. 2019, 126, 64–76. [Google Scholar] [CrossRef] [Green Version]
- Song, R.-H.; Xu, W.; Chen, Y.; Li, Z.; Zeng, Y.; Fu, Y. The expression of Sirtuins 1 and 4 in peripheral blood leukocytes from patients with type 2 diabetes. Eur. J. Histochem. 2011, 55, e10. [Google Scholar] [CrossRef] [Green Version]
- Kolthur-Seetharam, U.; Dantzer, F.; McBurney, M.W.; de Murcia, G.; Sassone-Corsi, P. Control of AIF-mediated Cell Death by the Functional Interplay of SIRT1 and PARP-1 in Response to DNA Damage. Cell Cycle 2006, 5, 873–877. [Google Scholar] [CrossRef] [Green Version]
- Gomes, B.A.Q.; Silva, J.P.B.; Romeiro, C.F.R.; dos Santos, S.M.; Rodrigues, C.A.; Gonçalves, P.R.; Sakai, J.T.; Mendes, P.F.S.; Varela, E.L.P.; Monteiro, M.C. Neuroprotective Mechanisms of Resveratrol in Alzheimer’s Disease: Role of SIRT1. Oxidative Med. Cell. Longev. 2018, 2018, 1–15. [Google Scholar] [CrossRef]
- Banks, A.S.; Kon, N.; Knight, C.; Matsumoto, M.; Gutiérrez-Juárez, R.; Rossetti, L.; Gu, W.; Accili, D. SirT1 Gain of Function Increases Energy Efficiency and Prevents Diabetes in Mice. Cell Metab. 2008, 8, 333–341. [Google Scholar] [CrossRef] [Green Version]
- Bordone, L.; Cohen, D.; Robinson, A.; Motta, M.C.; van Veen, E.; Czopik, A.; Steele, A.D.; Crowe, H.; Marmor, S.; Luo, J.; et al. SIRT1 transgenic mice show phenotypes resembling calorie restriction. Aging Cell 2007, 6, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Simmons, J.G.E.; Pruitt, W.M.; Pruitt, K. Diverse Roles of SIRT1 in Cancer Biology and Lipid Metabolism. Int. J. Mol. Sci. 2015, 16, 950–965. [Google Scholar] [CrossRef] [PubMed]
- Cantó, C.; Auwerx, J. AMP-activated protein kinase and its downstream transcriptional pathways. Cell. Mol. Life Sci. 2010, 67, 3407–3423. [Google Scholar] [CrossRef] [Green Version]
- Rodgers, J.T.; Lerin, C.; Haas, W.; Gygi, S.P.; Spiegelman, B.M.; Puigserver, P. Nutrient control of glucose homeostasis through a complex of PGC-1α and SIRT1. Nat. Cell Biol. 2005, 434, 113–118. [Google Scholar] [CrossRef]
- Sun, C.; Zhang, F.; Ge, X.; Yan, T.; Chen, X.; Shi, X.; Zhai, Q. SIRT1 Improves Insulin Sensitivity under Insulin-Resistant Conditions by Repressing PTP1B. Cell Metab. 2007, 6, 307–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulkarni, S.S.; Canto, C. The molecular targets of resveratrol. Biochim. Biophys. Acta Mol. Basis Dis. 2015, 1852, 1114–1123. [Google Scholar] [CrossRef] [Green Version]
- Sin, T.K.; Yung, B.Y.; Siu, P.M. Modulation of SIRT1-Foxo1 Signaling axis by Resveratrol: Implications in Skeletal Muscle Aging and Insulin Resistance. Cell. Physiol. Biochem. 2015, 35, 541–552. [Google Scholar] [CrossRef]
- Tang, Y.; Xu, J.; Qu, W.; Peng, X.; Xin, P.; Yang, X.; Ying, C.; Sun, X.; Hao, L. Resveratrol reduces vascular cell senescence through attenuation of oxidative stress by SIRT1/NADPH oxidase-dependent mechanisms. J. Nutr. Biochem. 2012, 23, 1410–1416. [Google Scholar] [CrossRef]
- Saunders, L.R.; Verdin, E. Sirtuins: Critical regulators at the crossroads between cancer and aging. Oncogene 2007, 26, 5489–5504. [Google Scholar] [CrossRef] [Green Version]
- Chung, J.H.; Manganiello, V.; Dyck, J.R. Resveratrol as a calorie restriction mimetic: Therapeutic implications. Trends Cell Biol. 2012, 22, 546–554. [Google Scholar] [CrossRef] [Green Version]
- Yeung, F.; E Hoberg, J.; Ramsey, C.S.; Keller, M.D.; Jones, D.R.; A Frye, R.; Mayo, M.W. Modulation of NF-κB-dependent transcription and cell survival by the SIRT1 deacetylase. EMBO J. 2004, 23, 2369–2380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Zuo, S.; Xin, W. miR-27b overexpression improves mitochondrial function in a Sirt1-dependent manner. J. Physiol. Biochem. 2015, 71, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Kleszcz, R.; Paluszczak, J.; Krajka-Kuźniak, V.; Baer-Dubowska, W. The inhibition of c-MYC transcription factor modulates the expression of glycolytic and glutaminolytic enzymes in FaDu hypopharyngeal carcinoma cells. Adv. Clin. Exp. Med. 2018, 27, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Mostoslavsky, R. SIRT6. Transcription 2010, 1, 17–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Yuan, H.; Roth, M.; Stark, J.M.; Bhatia, R.; Chen, W.Y. SIRT1 deacetylase promotes acquisition of genetic mutations for drug resistance in CML cells. Oncogene 2013, 32, 589–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, E.; Hou, J.; Ke, X.; Abbas, M.N.; Kausar, S.; Zhang, L.; Cui, H. The Roles of Sirtuin Family Proteins in Cancer Progression. Cancers 2019, 11, 1949. [Google Scholar] [CrossRef] [Green Version]
- Matsushima, S.; Sadoshima, J. The role of sirtuins in cardiac disease. Am. J. Physiol. Circ. Physiol. 2015, 309, H1375–H1389. [Google Scholar] [CrossRef] [Green Version]
- Wątroba, M.; Szukiewicz, D. The role of sirtuins in aging and age-related diseases. Adv. Med Sci. 2016, 61, 52–62. [Google Scholar] [CrossRef]
- Haigis, M.C.; Sinclair, D.A. Chapter 11—Sirtuins in Aging and Age-Related Diseases. In Handbook of the Biology of Aging, 7th ed.; Masoro, E.J., Austad, S.N., Eds.; Academic Press: San Diego, CA, USA, 2011; pp. 243–274. [Google Scholar] [CrossRef]
- Bosch-Presegué, L.; Vaquero, A. The Dual Role of Sirtuins in Cancer. Genes Cancer 2011, 2, 648–662. [Google Scholar] [CrossRef]
- German, N.J.; Haigis, M.C. Sirtuins and the Metabolic Hurdles in Cancer. Curr. Biol. 2015, 25, R569–R583. [Google Scholar] [CrossRef] [Green Version]
- Chao, S.-C.; Chen, Y.-J.; Huang, K.-Y.; Kuo, K.-L.; Yang, T.-H.; Wang, C.-C.; Tang, C.-H.; Yang, R.-S.; Liu, S.H. Induction of sirtuin-1 signaling by resveratrol induces human chondrosarcoma cell apoptosis and exhibits antitumor activity. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Liarte, S.; Alonso-Romero, J.L.; Nicolás, F.J. SIRT1 and Estrogen Signaling Cooperation for Breast Cancer Onset and Progression. Front. Endocrinol. 2018, 9, 552. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Wei, Y.; Xu, F.; Zhao, M.; Dai, K.; Shen, R.; Yang, S.; Zhang, N. SIRT1 promotes formation of breast cancer through modulating Akt activity. J. Cancer 2018, 9, 2012–2023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, S.; Sharma, S.; Vora, J.; Shrivastava, N. Emerging role of sirtuins in breast cancer metastasis and multidrug resistance: Implication for novel therapeutic strategies targeting sirtuins. Pharmacol. Res. 2020, 158, 104880. [Google Scholar] [CrossRef]
- Shaito, A.; Posadino, A.M.; Younes, N.; Hasan, H.; Halabi, S.; Alhababi, D.; Al-Mohannadi, A.; Abdel-Rahman, W.M.; Eid, A.H.; Nasrallah, G.K.; et al. Potential Adverse Effects of Resveratrol: A Literature Review. Int. J. Mol. Sci. 2020, 21, 2084. [Google Scholar] [CrossRef] [Green Version]
- Herranz, D.; Maraver, A.; Cañamero, M.; Gómez-López, G.; Inglada-Pérez, L.; Robledo, M.; Castelblanco, E.; Matias-Guiu, X.; Serrano, M. SIRT1 promotes thyroid carcinogenesis driven by PTEN deficiency. Oncogene 2012, 32, 4052–4056. [Google Scholar] [CrossRef]
- Wu, W.; Zhang, L.; Lin, J.; Huang, H.; Shi, B.; Lin, X.; Huang, Z.; Wang, C.; Qiu, J.; Wei, X. Hypermethylation of the HIC1 promoter and aberrant expression of HIC1/SIRT1 contribute to the development of thyroid papillary carcinoma. Oncotarget 2016, 7, 84416–84427. [Google Scholar] [CrossRef] [Green Version]
- Herranz, D.; Muñoz-Martin, M.; Cañamero, M.; Mulero, F.; Martinez-Pastor, B.; Fernandez-Capetillo, O.; Serrano, M. Sirt1 improves healthy ageing and protects from metabolic syndrome-associated cancer. Nat. Commun. 2010, 1, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Menssen, A.; Hydbring, P.; Kapelle, K.; Vervoorts, J.; Diebold, J.; Lüscher, B.; Larsson, L.-G.; Hermeking, H. The c-MYC oncoprotein, the NAMPT enzyme, the SIRT1-inhibitor DBC1, and the SIRT1 deacetylase form a positive feedback loop. Proc. Natl. Acad. Sci. USA 2011, 109, E187–E196. [Google Scholar] [CrossRef] [Green Version]
- Kriegl, L.; Vieth, M.; Kirchner, T.; Menssen, A. Up-regulation of c-MYC and SIRT1 expression correlates with malignant transformation in the serrated route to colorectal cancer. Oncotarget 2012, 3, 1182–1193. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.; Wang, Z.; Li, L.; Zhang, H.; Modi, H.; Horne, D.; Bhatia, R.; Chen, W. Activation of stress response gene SIRT1 by BCR-ABL promotes leukemogenesis. Blood 2012, 119, 1904–1914. [Google Scholar] [CrossRef] [Green Version]
- Sasca, D.; Hähnel, P.S.; Szybinski, J.; Khawaja, K.; Kriege, O.; Pante, S.V.; Bullinger, L.; Strand, S.; Strand, D.; Theobald, M.; et al. SIRT1 prevents genotoxic stress-induced p53 activation in acute myeloid leukemia. Blood 2014, 124, 121–133. [Google Scholar] [CrossRef] [Green Version]
- Wong, S.; Weber, J.D. Deacetylation of the retinoblastoma tumour suppressor protein by SIRT1. Biochem. J. 2007, 407, 451–460. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.-X.; Li, H.; Chen, X.-M.; Yang, X.-H.; Wang, Q.; Wu, M.-L.; Kong, Q.-Y.; Li, Z.-X.; Liu, J. Expression patterns and potential roles of SIRT1 in human medulloblastoma cellsin vivoandin vitro. Neuropathology 2013, 33, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, X.; Cui, Y.; Wei, Q.; Chen, S.-Y.; Wang, X. Effects of SIRT1 silencing on viability, invasion and metastasis of human glioma cell lines. Oncol. Lett. 2019, 17, 3701–3708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Yang, W.; Dong, F.; Guo, Y.; Tan, J.; Ruan, S.; Huang, T. The prognostic role of Sirt1 expression in solid malignancies: A meta-analysis. Oncotarget 2017, 8, 66343–66351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiraike, H.; Wada-Hiraike, O.; Nakagawa, S.; Saji, S.; Maeda, D.; Miyamoto, Y.; Sone, K.; Tanikawa, M.; Oda, K.; Nakagawa, K.; et al. Expression of DBC1 is associated with nuclear grade and HER2 expression in breast cancer. Exp. Ther. Med. 2011, 2, 1105–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.; Wei, W.; Xiao, X.; Guo, J.; Xie, X.; Li, L.; Kong, Y.; Lv, N.; Jia, W.; Zhang, Y.; et al. Expression of SIRT1 is associated with lymph node metastasis and poor prognosis in both operable triple-negative and non-triple-negative breast cancer. Med. Oncol. 2012, 29, 3240–3249. [Google Scholar] [CrossRef]
- Lee, H.; Kim, K.R.; Noh, S.J.; Park, H.S.; Kwon, K.S.; Park, B.-H.; Jung, S.H.; Youn, H.J.; Kil Lee, B.; Chung, M.J.; et al. Expression of DBC1 and SIRT1 is associated with poor prognosis for breast carcinoma. Hum. Pathol. 2011, 42, 204–213. [Google Scholar] [CrossRef]
- Jung-Hynes, B.; Nihal, M.; Zhong, W.; Ahmad, N. Role of Sirtuin Histone Deacetylase SIRT1 in Prostate Cancer. J. Biol. Chem. 2008, 284, 3823–3832. [Google Scholar] [CrossRef] [Green Version]
- Huffman, D.M.; Grizzle, W.E.; Bamman, M.M.; Kim, J.-S.; Eltoum, I.A.; Elgavish, A.; Nagy, T.R. SIRT1 Is Significantly Elevated in Mouse and Human Prostate Cancer. Cancer Res. 2007, 67, 6612–6618. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhang, M.; Dong, H.; Yong, S.; Li, X.; E Olashaw, N.; A Kruk, P.; Cheng, J.Q.; Bai, W.; Chen, J.; et al. Deacetylation of cortactin by SIRT1 promotes cell migration. Oncogene 2008, 28, 445–460. [Google Scholar] [CrossRef] [Green Version]
- Tseng, R.-C.; Lee, C.-C.; Hsu, H.-S.; Tzao, C.; Wang, Y.-C. Distinct HIC1-SIRT1-p53 Loop Deregulation in Lung Squamous Carcinoma and Adenocarcinoma Patients. Neoplasia 2009, 11, 763–W11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, A.N.; Zhang, L.H.; Fan, X.S.; Huang, Q.; Ye, Q.; Wu, H.Y.; Yang, J. Expression of SIRT1 in Gastric Cardiac Cancer and Its Clinicopathologic Significance. Int. J. Surg. Pathol. 2011, 19, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Cha, E.J.; Noh, S.J.; Kwon, K.S.; Kim, C.Y.; Park, B.-H.; Park, H.S.; Lee, H.; Chung, M.J.; Kang, M.J.; Lee, D.-G.; et al. Expression of DBC1 and SIRT1 Is Associated with Poor Prognosis of Gastric Carcinoma. Clin. Cancer Res. 2009, 15, 4453–4459. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Zhang, B.; Wong, N.; Lo, A.W.; To, K.-F.; Chan, A.W.; Ng, M.H.; Ho, C.Y.; Cheng, S.-H.; Lai, P.B.; et al. Sirtuin 1 Is Upregulated in a Subset of Hepatocellular Carcinomas where It Is Essential for Telomere Maintenance and Tumor Cell Growth. Cancer Res. 2011, 71, 4138–4149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, H.J.; Noh, J.H.; Kim, J.K.; Eun, J.W.; Jung, K.H.; Kim, M.G.; Chang, Y.G.; Shen, Q.; Kim, S.-J.; Park, W.S.; et al. MicroRNA-29c functions as a tumor suppressor by direct targeting oncogenic SIRT1 in hepatocellular carcinoma. Oncogene 2013, 33, 2557–2567. [Google Scholar] [CrossRef] [Green Version]
- Jang, K.Y.; Noh, S.J.; Lehwald, N.; Tao, G.-Z.; Bellovin, D.I.; Park, H.S.; Moon, W.S.; Felsher, D.W.; Sylvester, K.G. SIRT1 and c-Myc Promote Liver Tumor Cell Survival and Predict Poor Survival of Human Hepatocellular Carcinomas. PLoS ONE 2012, 7, e45119. [Google Scholar] [CrossRef]
- Chen, H.-C.; Jeng, Y.-M.; Yuan, R.-H.; Hsu, H.-C.; Chen, Y.-L. SIRT1 Promotes Tumorigenesis and Resistance to Chemotherapy in Hepatocellular Carcinoma and its Expression Predicts Poor Prognosis. Ann. Surg. Oncol. 2011, 19, 2011–2019. [Google Scholar] [CrossRef]
- Jang, K.Y.; Kim, K.S.; Hwang, S.H.; Kwon, K.S.; Kim, K.R.; Park, H.S.; Park, B.-H.; Chung, M.J.; Kang, M.J.; Lee, D.-G.; et al. Expression and prognostic significance of SIRT1 in ovarian epithelial tumours. Pathology 2009, 41, 366–371. [Google Scholar] [CrossRef]
- Mvunta, D.H.; Miyamoto, T.; Asaka, R.; Yamada, Y.; Ando, H.; Higuchi, S.; Ida, K.; Kashima, H.; Shiozawa, T. Overexpression of SIRT1 is Associated with Poor Outcomes in Patients with Ovarian Carcinoma. Appl. Immunohistochem. Mol. Morphol. 2017, 25, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Chu, Z.; Ma, P.; Meng, Y.; Yang, Y. SIRT1 promotes the proliferation and metastasis of human pancreatic cancer cells. Tumor Biol. 2017, 39, 1010428317691180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, G.; Cui, J.; Zhang, J.-G.; Qin, Q.; Chen, Q.; Yin, T.; Deng, S.-C.; Liu, Y.; Liu, L.; Wang, B.; et al. SIRT1 RNAi knockdown induces apoptosis and senescence, inhibits invasion and enhances chemosensitivity in pancreatic cancer cells. Gene Ther. 2011, 18, 920–928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benavente, C.A.; Schnell, S.A.; Jacobson, E.L. Effects of Niacin Restriction on Sirtuin and PARP Responses to Photodamage in Human Skin. PLoS ONE 2012, 7, e42276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilking, M.J.; Singh, C.K.; Nihal, M.; Zhong, W.; Ahmad, N. SIRT1 deacetylase is overexpressed in human melanoma and its small molecule inhibition imparts anti-proliferative response via p53 activation. Arch. Biochem. Biophys. 2014, 563, 94–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.-H.; Sengupta, K.; Li, C.; Kim, H.-S.; Cao, L.; Xiao, C.; Kim, S.; Xu, X.; Zheng, Y.; Chilton, B.; et al. Impaired DNA Damage Response, Genome Instability, and Tumorigenesis in SIRT1 Mutant Mice. Cancer Cell 2008, 14, 312–323. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.-H.; Zheng, Y.; Kim, H.-S.; Xu, X.; Cao, L.; Lahusen, T.; Lee, M.-H.; Xiao, C.; Vassilopoulos, A.; Chen, W.; et al. Interplay among BRCA1, SIRT1, and Survivin during BRCA1-Associated Tumorigenesis. Mol. Cell 2008, 32, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Costa-Machado, L.F.; Martín-Hernández, R.; Sanchez-Luengo, M. Ángel; Hess, K.; Vales-Villamarín, C.; Barradas, M.; Lynch, C.; de la Nava, D.; Diaz-Ruiz, A.; de Cabo, R.; et al. Sirt1 protects from K-Ras-driven lung carcinogenesis. EMBO Rep. 2018, 19, e43879. [Google Scholar] [CrossRef]
- Yang, Q.; Wang, B.; Gao, W.; Huang, S.; Liu, Z.; Li, W.; Jia, J. SIRT1 Is Downregulated in Gastric Cancer and Leads to G1-phase Arrest via NF-κB/Cyclin D1 Signaling. Mol. Cancer Res. 2013, 11, 1497–1507. [Google Scholar] [CrossRef] [Green Version]
- Firestein, R.; Blander, G.; Michan, S.; Oberdoerffer, P.; Ogino, S.; Campbell, J.; Bhimavarapu, A.; Luikenhuis, S.; de Cabo, R.; Fuchs, C.; et al. The SIRT1 Deacetylase Suppresses Intestinal Tumorigenesis and Colon Cancer Growth. PLoS ONE 2008, 3, e2020. [Google Scholar] [CrossRef]
- Kabra, N.; Li, Z.; Chen, L.; Li, B.; Zhang, X.; Wang, C.; Yeatman, T.; Coppola, D.; Chen, J. SirT1 Is an Inhibitor of Proliferation and Tumor Formation in Colon Cancer. J. Biol. Chem. 2009, 284, 18210–18217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oon, C.E.; Strell, C.; Yeong, K.Y.; Östman, A.; Prakash, J. SIRT1 inhibition in pancreatic cancer models: Contrasting effects in vitro and in vivo. Eur. J. Pharmacol. 2015, 757, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chan, A.W.; To, K.-F.; Chen, W.; Zhang, Z.; Ren, J.; Song, C.; Cheung, Y.-S.; Lai, P.B.; Cheng, S.-H.; et al. SIRT2 overexpression in hepatocellular carcinoma mediates epithelial to mesenchymal transition by protein kinase B/glycogen synthase kinase-3β/β-catenin signaling. Hepatology 2013, 57, 2287–2298. [Google Scholar] [CrossRef]
- Huang, S.; Zhao, Z.; Tang, D.; Zhou, Q.; Li, Y.; Zhou, L.; Yin, Y.; Wang, Y.; Pan, Y.; Dorfman, R.G.; et al. Downregulation of SIRT2 Inhibits Invasion of Hepatocellular Carcinoma by Inhibiting Energy Metabolism. Transl. Oncol. 2017, 10, 917–927. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.Y.; Xu, N.; Malyukova, A.; Scarlett, C.J.; Sun, Y.T.; Zhang, X.D.; Ling, D.; Su, S.-P.; Nelson, C.; Chang, D.K.; et al. The histone deacetylase SIRT2 stabilizes Myc oncoproteins. Cell Death Differ. 2012, 20, 503–514. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Ni, T.K.; Wronski, A.; Glass, B.; Skibinski, A.; Beck, A.; Kuperwasser, C. The SIRT2 Deacetylase Stabilizes Slug to Control Malignancy of Basal-like Breast Cancer. Cell Rep. 2016, 17, 1302–1317. [Google Scholar] [CrossRef] [Green Version]
- Jing, H.; Hu, J.; He, B.; Abril, Y.L.N.; Stupinski, J.; Weiser, K.; Carbonaro, M.; Chiang, Y.-L.; Southard, T.; Giannakakou, P.; et al. A SIRT2-Selective Inhibitor Promotes c-Myc Oncoprotein Degradation and Exhibits Broad Anticancer Activity. Cancer Cell 2016, 29, 607. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Bao, Y.-C.; Ji, X.-X.; Chen, B.; Deng, Q.-F.; Zhou, S.-W. SPOP promotes SIRT2 degradation and suppresses non-small cell lung cancer cell growth. Biochem. Biophys. Res. Commun. 2017, 483, 880–884. [Google Scholar] [CrossRef]
- Wei, R.; He, D.; Zhang, X. Role of SIRT2 in Regulation of Stemness of Cancer Stem-Like Cells in Renal Cell Carcinoma. Cell. Physiol. Biochem. 2018, 49, 2348–2357. [Google Scholar] [CrossRef]
- Funato, K.; Hayashi, T.; Echizen, K.; Negishi, L.; Shimizu, N.; Koyama-Nasu, R.; Nasu-Nishimura, Y.; Morishita, Y.; Tabar, V.; Todo, T.; et al. SIRT 2-mediated inactivation of p73 is required for glioblastoma tumorigenicity. EMBO Rep. 2018, 19, e45587. [Google Scholar] [CrossRef]
- Wilking-Busch, M.J.; Ndiaye, M.A.; Huang, W.; Ahmad, N. Expression profile of SIRT2 in human melanoma and implications for sirtuin-based chemotherapy. Cell Cycle 2017, 16, 574–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Y.; Wu, J.; Zhang, H.; Li, S.; Sun, H. Reduced expression of SIRT2 in serous ovarian carcinoma promotes cell proliferation through disinhibition of CDK4 expression. Mol. Med. Rep. 2017, 15, 1638–1646. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-S.; Vassilopoulos, A.; Wang, R.-H.; Lahusen, T.; Xiao, Z.; Xu, X.; Li, C.; Veenstra, T.D.; Li, B.; Yu, H.; et al. SIRT2 Maintains Genome Integrity and Suppresses Tumorigenesis through Regulating APC/C Activity. Cancer Cell 2011, 20, 487–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.; Jiang, K.; Shen, M.; Qian, Y.; Peng, Y. SIRT2 suppresses non-small cell lung cancer growth by targeting JMJD2A. Biol. Chem. 2015, 396, 929–936. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Huang, J.; Yuan, H.; Chen, Z.; Luo, Q.; Lu, S. SIRT2 inhibits non-small cell lung cancer cell growth through impairing Skp2-mediated p27 degradation. Oncotarget 2016, 7, 18927–18939. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Xie, Q.R.; Chen, Z.; Lu, S.; Xia, W. Regulation of SIRT2 levels for human non-small cell lung cancer therapy. Lung Cancer 2013, 82, 9–15. [Google Scholar] [CrossRef]
- Song, H.Y.; Biancucci, M.; Kang, H.-J.; O’Callaghan, C.; Park, S.-H.; Principe, D.R.; Jiang, H.; Yan, Y.; Satchell, K.F.; Raparia, K.; et al. SIRT2 deletion enhances KRAS-induced tumorigenesis in vivo by regulating K147 acetylation status. Oncotarget 2016, 7, 80336–80349. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.H.; Laurent, G.; Bause, A.S.; Spang, R.; German, N.; Haigis, M.C.; Haigis, K.M. HDAC6 and SIRT2 Regulate the Acetylation State and Oncogenic Activity of Mutant K-RAS. Mol. Cancer Res. 2013, 11, 1072–1077. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Wang, L.; Zhou, L.; Dorfman, R.G.; Pan, Y.; Tang, D.; Wang, Y.; Yin, Y.; Jiang, C.; Xu, G.; et al. The SIRT2/cMYC Pathway Inhibits Peroxidation-Related Apoptosis in Cholangiocarcinoma through Metabolic Reprogramming. Neoplasia 2019, 21, 429–441. [Google Scholar] [CrossRef]
- Damodaran, S.; Damaschke, N.; Gawdzik, J.; Yang, B.; Shi, C.; Allen, G.O.; Huang, W.; Denu, J.M.; Jarrard, D.F. Dysregulation of Sirtuin 2 (SIRT2) and histone H3K18 acetylation pathways associates with adverse prostate cancer outcomes. BMC Cancer 2017, 17, 874. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.-L.; Zhan, L.; Jin, Y.-D.; Min, Z.-L.; Wei, C.; Wang, Q.; Chen, Y.-J.; Wu, Q.-M.; Hu, X.-M.; Yuan, Q. SIRT2 mediated antitumor effects of shikonin on metastatic colorectal cancer. Eur. J. Pharmacol. 2017, 797, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Quan, S.; Principe, D.R.; Dean, A.E.; Park, S.-H.; Grippo, P.J.; Gius, D.; Horikoshi, N. Loss of Sirt2 increases and prolongs a caerulein-induced pancreatitis permissive phenotype and induces spontaneous oncogenic Kras mutations in mice. Sci. Rep. 2018, 8, 16501. [Google Scholar] [CrossRef] [PubMed]
- Ming, M.; Qiang, L.; Zhao, B.; He, Y.-Y. Mammalian SIRT2 inhibits keratin 19 expression and is a tumor suppressor in skin. Exp. Dermatol. 2014, 23, 207–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Temel, M.; Koç, M.N.; Ulutaş, S.; Göğebakan, B. The expression levels of the sirtuins in patients with BCC. Tumor Biol. 2015, 37, 6429–6435. [Google Scholar] [CrossRef]
- Li, S.; Banck, M.; Mujtaba, S.; Zhou, M.-M.; Sugrue, M.M.; Walsh, M.J. p53-Induced Growth Arrest Is Regulated by the Mitochondrial SirT3 Deacetylase. PLoS ONE 2010, 5, e10486. [Google Scholar] [CrossRef]
- Liu, C.; Huang, Z.; Jiang, H.; Shi, F. The Sirtuin 3 Expression Profile Is Associated with Pathological and Clinical Outcomes in Colon Cancer Patients. BioMed Res. Int. 2014, 2014, 1–9. [Google Scholar] [CrossRef]
- Wei, Z.; Song, J.; Wang, G.; Cui, X.; Zheng, J.; Tang, Y.; Chen, X.; Li, J.; Cui, L.; Liu, C.-Y.; et al. Deacetylation of serine hydroxymethyl-transferase 2 by SIRT3 promotes colorectal carcinogenesis. Nat. Commun. 2018, 9, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.; Li, X.; Zhang, Z.; Jia, L.; Wang, M.; Zhao, J.; Han, Y. SIRT3 is correlated with the malignancy of non-small cell lung cancer. Int. J. Oncol. 2017, 50, 903–910. [Google Scholar] [CrossRef] [Green Version]
- Marfe, G.; Tafani, M.; Indelicato, M.; Sinibaldi-Salimei, P.; Reali, V.; Pucci, B.; Fini, M.; Russo, M.A. Kaempferol induces apoptosis in two different cell lines via Akt inactivation, Bax and SIRT3 activation, and mitochondrial dysfunction. J. Cell. Biochem. 2009, 106, 643–650. [Google Scholar] [CrossRef]
- Dong, X.-C.; Jing, L.-M.; Wang, W.-X.; Gao, Y.-X. Down-regulation of SIRT3 promotes ovarian carcinoma metastasis. Biochem. Biophys. Res. Commun. 2016, 475, 245–250. [Google Scholar] [CrossRef]
- George, J.; Nihal, M.; Singh, C.K.; Zhong, W.; Liu, X.; Ahmad, N. Pro-Proliferative Function of Mitochondrial Sirtuin Deacetylase SIRT3 in Human Melanoma. J. Investig. Dermatol. 2016, 136, 809–818. [Google Scholar] [CrossRef] [Green Version]
- He, S.; He, C.; Yuan, H.; Xiong, S.; Xiao, Z.; Chen, L. The SIRT 3 Expression Profile is Associated with Pathological and Clinical Outcomes in Human Breast Cancer Patients. Cell. Physiol. Biochem. 2014, 34, 2061–2069. [Google Scholar] [CrossRef]
- Quan, Y.; Wang, N.; Chen, Q.; Xu, J.; Cheng, W.; Di, M.; Xia, W.; Gao, W.-Q. SIRT3 inhibits prostate cancer by destabilizing oncoprotein c-MYC through regulation of the PI3K/Akt pathway. Oncotarget 2015, 6, 26494–26507. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Quan, Y.; Xia, W. SIRT3 inhibits prostate cancer metastasis through regulation of FOXO3A by suppressing Wnt/β-catenin pathway. Exp. Cell Res. 2018, 364, 143–151. [Google Scholar] [CrossRef]
- Zeng, X.; Wang, N.; Wang, R.; Zhai, H.; Wu, J.; Pu, W. SIRT3 functions as a tumor suppressor in hepatocellular carcinoma. Tumor Biol. 2017, 39, 1010428317691178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Cheng, W.; Yin, X.-M.; Jiang, B. The expression of SIRT3 in primary hepatocellular carcinoma and the mechanism of its tumor suppressing effects. Eur. Rev. Med Pharmacol. Sci. 2017, 21, 978–998. [Google Scholar] [PubMed]
- Bo, Z.; Qin, L.; Zhou, C.-J.; Liu, Y.-L.; Qian, H.; He, S.-B. SIRT3 expression in hepatocellular carcinoma and its impact on proliferation and invasion of hepatoma cells. Asian Pac. J. Trop. Med. 2013, 6, 649–652. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.Z.; Liu, L.; Cai, M.; Pan, Y.; Fu, J.; Cao, Y.; Yun, J. Low SIRT3 Expression Correlates with Poor Differentiation and Unfavorable Prognosis in Primary Hepatocellular Carcinoma. PLoS ONE 2012, 7, e51703. [Google Scholar] [CrossRef]
- Huang, S.; Guo, H.; Cao, Y.; Xiong, J. MiR-708-5p inhibits the progression of pancreatic ductal adenocarcinoma by targeting Sirt3. Pathol. Res. Pract. 2019, 215, 794–800. [Google Scholar] [CrossRef]
- Kamarajan, P.; Dds, T.Y.A.; Danciu, T.; Silva, N.J.D.; Verdin, E.; Kapila, Y.L. Receptor-interacting protein (RIP) and Sirtuin-3 (SIRT3) are on opposite sides of anoikis and tumorigenesis. Cancer 2012, 118, 5800–5810. [Google Scholar] [CrossRef]
- Shackelford, R.; Hirsh, S.; Henry, K.; Abdel-Mageed, A.; Kandil, E.; Coppola, D. Nicotinamide phosphoribosyltransferase and SIRT3 expression are increased in well-differentiated thyroid carcinomas. Anticancer Res. 2013, 33, 3047–3052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Wang, W.-Y.; Cao, L. SIRT3 inhibits cell proliferation in human gastric cancer through down-regulation of Notch-1. Int. J. Clin. Exp. Med. 2015, 8, 5263–5271. [Google Scholar] [PubMed]
- Huang, K.H.; Hsu, C.-C.; Fang, W.-L.; Chi, C.-W.; Sung, M.-T.; Kao, H.-L.; Li, A.F.-Y.; Yin, P.-H.; Yang, M.; Lee, H.-C. SIRT3 Expression as a Biomarker for Better Prognosis in Gastric Cancer. World J. Surg. 2013, 38, 910–917. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Fu, X.; Shao, L.; Ding, Y.; Zeng, D. Aberrant expression of SIRT3 is conversely correlated with the progression and prognosis of human gastric cancer. Biochem. Biophys. Res. Commun. 2014, 443, 156–160. [Google Scholar] [CrossRef]
- Huang, S.; Chen, X.; Zheng, J.; Huang, Y.; Song, L.; Yin, Y.; Xiong, J. Low SIRT3 expression contributes to tumor progression, development and poor prognosis in human pancreatic carcinoma. Pathol. Res. Pract. 2017, 213, 1419–1423. [Google Scholar] [CrossRef]
- Finley, L.W.; Carracedo, A.; Lee, J.; Souza, A.; Egia, A.; Zhang, J.; Teruya-Feldstein, J.; Moreira, P.I.; Cardoso, S.M.; Clish, C.B.; et al. SIRT3 Opposes Reprogramming of Cancer Cell Metabolism through HIF1α Destabilization. Cancer Cell 2011, 19, 416–428. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-S.; Patel, K.; Muldoon-Jacobs, K.; Bisht, K.S.; Aykin-Burns, N.; Pennington, J.D.; van der Meer, R.; Nguyen, P.; Savage, J.; Owens, K.M.; et al. SIRT3 Is a Mitochondria-Localized Tumor Suppressor Required for Maintenance of Mitochondrial Integrity and Metabolism during Stress. Cancer Cell 2010, 17, 41–52. [Google Scholar] [CrossRef] [Green Version]
- Desouki, M.M.; Doubinskaia, I.; Gius, D.; Abdulkadir, S.A. Decreased mitochondrial SIRT3 expression is a potential molecular biomarker associated with poor outcome in breast cancer. Hum. Pathol. 2014, 45, 1071–1077. [Google Scholar] [CrossRef] [Green Version]
- Huang, G.; Lin, Y.; Zhu, G. SIRT4 is upregulated in breast cancer and promotes the proliferation, migration and invasion of breast cancer cells. Int. J. Clin. Exp. Pathol. 2017, 10, 11849–11856. [Google Scholar]
- Jeong, S.M.; Xiao, C.; Finley, L.W.; Lahusen, T.; Souza, A.L.; Pierce, K.; Li, Y.-H.; Wang, X.; Laurent, G.; German, N.J.; et al. SIRT4 Has Tumor-Suppressive Activity and Regulates the Cellular Metabolic Response to DNA Damage by Inhibiting Mitochondrial Glutamine Metabolism. Cancer Cell 2013, 23, 450–463. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.; Dong, Q.; He, J.; Wang, X.; Xing, J.; Wang, E.; Qiu, X.; Li, Q. SIRT4 inhibits malignancy progression of NSCLCs, through mitochondrial dynamics mediated by the ERK-Drp1 pathway. Oncogene 2017, 36, 2724–2736. [Google Scholar] [CrossRef]
- Gong, J.; Wang, H.; Lou, W.; Wang, G.; Tao, H.; Wen, H.; Liu, Y.; Xie, Q. Associations of sirtuins with clinicopathological parameters and prognosis in non–small cell lung cancer. Cancer Manag. Res. 2018, 10, 3341–3356. [Google Scholar] [CrossRef] [Green Version]
- Nakahara, Y.; Yamasaki, M.; Sawada, G.; Miyazaki, Y.; Makino, T.; Kurokawa, Y.; Takiguchi, S.; Mimori, K.; Doki, Y.; Takahashi, T.; et al. Downregulation of SIRT4 Expression Is Associated with Poor Prognosis in Esophageal Squamous Cell Carcinoma. Oncology 2016, 90, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Csibi, A.; Fendt, S.-M.; Li, C.; Poulogiannis, G.; Choo, A.Y.; Chapski, D.J.; Jeong, S.M.; Dempsey, J.M.; Parkhitko, A.; Morrison, T.; et al. The mTORC1 Pathway Stimulates Glutamine Metabolism and Cell Proliferation by Repressing SIRT4. Cell 2013, 153, 840–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, G.; Cheng, J.; Yu, F.; Liu, X.; Yuan, C.; Liu, C.; Chen, X.; Peng, Z. Clinical and therapeutic significance of sirtuin-4 expression in colorectal cancer. Oncol. Rep. 2016, 35, 2801–2810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyo, M.; Yamamoto, H.; Konno, M.; Colvin, H.; Nishida, N.; Koseki, J.; Kawamoto, K.; Ogawa, H.; Hamabe, A.; Uemura, M.; et al. Tumour-suppressive function of SIRT4 in human colorectal cancer. Br. J. Cancer 2015, 113, 492–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.; Huang, D.; Liu, G.; Jian, F.; Zhu, J.; Zhang, L. SIRT4 acts as a tumor suppressor in gastric cancer by inhibiting cell proliferation, migration, and invasion. OncoTargets Ther. 2018, 11, 3959–3968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Du, L.; Liang, X.; Meng, P.; Bi, L.; Wang, Y.; Wang, C.; Tang, B. Sirtuin 4 Depletion Promotes Hepatocellular Carcinoma Tumorigenesis Through Regulating Adenosine-Monophosphate–Activated Protein Kinase Alpha/Mammalian Target of Rapamycin Axis in Mice. Hepatology 2019, 69, 1614–1631. [Google Scholar] [CrossRef]
- Wang, C.; Piao, C.; Liu, J.; Zhang, Z.; Zhu, Y.; Kong, C. Mammalian SIRT4 is a tumor suppressor of clear cell renal cell carcinoma by inhibiting cancer proliferation, migration and invasion. Cancer Biomark. 2020, 29, 453–462. [Google Scholar] [CrossRef]
- Wang, Y.; Guo, Y.; Gao, J.; Yuan, X. Tumor-suppressive function of SIRT4 in neuroblastoma through mitochondrial damage. Cancer Manag. Res. 2018, 10, 5591–5603. [Google Scholar] [CrossRef] [Green Version]
- Shi, Q.; Liu, T.; Zhang, X.; Geng, J.; He, X.; Nu, M.; Pang, D. Decreased sirtuin 4 expression is associated with poor prognosis in patients with invasive breast cancer. Oncol. Lett. 2016, 12, 2606–2612. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-Q.; Wang, H.-L.; Xu, J.; Tan, J.; Fu, L.-N.; Wang, J.-L.; Zou, T.-H.; Sun, D.-F.; Gao, Q.-Y.; Chen, Y.-X.; et al. Sirtuin5 contributes to colorectal carcinogenesis by enhancing glutaminolysis in a deglutarylation-dependent manner. Nat. Commun. 2018, 9, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Z.; Liu, X.; Chen, T.; Gao, W.; Wu, Z.; Hu, Z.; Wei, D.; Gao, C.; Li, Q. Targeting a Sirt5-Positive Subpopulation Overcomes Multidrug Resistance in Wild-Type Kras Colorectal Carcinomas. Cell Rep. 2018, 22, 2677–2689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Wang, Z.; Li, X.; Liu, B.; Liu, M.; Liu, L.; Chen, S.; Ren, M.; Wang, Y.; Yu, M.; et al. SHMT2 Desuccinylation by SIRT5 Drives Cancer Cell Proliferation. Cancer Res. 2018, 78, 372–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, X.-B.; Liu, L.; Cheng, C.; Yu, B.; Xiong, L.; Hu, K.; Tang, J.; Zeng, L.; Sang, Y. SUN2 exerts tumor suppressor functions by suppressing the Warburg effect in lung cancer. Sci. Rep. 2015, 5, 17940. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Zuo, Y.; Feng, Y.; Zhang, M. SIRT5 facilitates cancer cell growth and drug resistance in non-small cell lung cancer. Tumor Biol. 2014, 35, 10699–10705. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Xi, L.; Liu, Y.; Liu, R.; Wu, Z.; Jian, Z. SIRT5 promotes cell proliferation and invasion in hepatocellular carcinoma by targeting E2F1. Mol. Med. Rep. 2017, 17, 342–349. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Qi, Y.; Wang, L.; Zheng, Z.; Zhang, Y.; Zheng, J. SIRT5-mediated SDHA desuccinylation promotes clear cell renal cell carcinoma tumorigenesis. Free. Radic. Biol. Med. 2019, 134, 458–467. [Google Scholar] [CrossRef]
- Park, J.; Chen, K.; Park, J.; Pak, M.; Verhaegen, M.; Fullen, D.; Scott, D.; Osterman, A.; Wang, M.; Andea, A.; et al. Human Melanoma Cell Need SIRT5 to Survive. Free. Radic. Biol. Med. 2016, 100, S128. [Google Scholar] [CrossRef]
- Xu, L.; Che, X.; Wu, Y.; Song, N.; Shi, S.; Wang, S.; Li, C.; Zhang, L.; Zhang, X.; Qu, X.; et al. SIRT5 as a biomarker for response to anthracycline-taxane-based neoadjuvant chemotherapy in triple-negative breast cancer. Oncol. Rep. 2018, 39, 2315–2323. [Google Scholar] [CrossRef]
- Li, F.; He, X.; Ye, D.; Lin, Y.; Yu, H.; Yao, C.; Huang, L.; Zhang, J.; Wang, F.; Xu, S.; et al. NADP+-IDH Mutations Promote Hypersuccinylation that Impairs Mitochondria Respiration and Induces Apoptosis Resistance. Mol. Cell 2015, 60, 661–675. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Tian, M.; Sun, R.; Zhang, M.; Zhou, L.; Jin, L.; Chen, L.; Zhou, W.; Duan, K.; Chen, Y.; et al. SIRT 5 inhibits peroxisomal ACOX 1 to prevent oxidative damage and is downregulated in liver cancer. EMBO Rep. 2018, 19, e45124. [Google Scholar] [CrossRef] [PubMed]
- Ran, L.-K.; Chen, Y.; Zhang, Z.-Z.; Tao, N.-N.; Ren, J.-H.; Zhou, L.; Tang, H.; Chen, X.; Chen, K.; Li, W.-Y.; et al. SIRT6 Overexpression Potentiates Apoptosis Evasion in Hepatocellular Carcinoma via BCL2-Associated X Protein–Dependent Apoptotic Pathway. Clin. Cancer Res. 2016, 22, 3372–3382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ning, S.; Ma, S.; Saleh, A.Q.; Guo, L.; Zhao, Z.; Chen, Y. SHMT2 Overexpression Predicts Poor Prognosis in Intrahepatic Cholangiocarcinoma. Gastroenterol. Res. Pract. 2018, 2018, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefort, K.; Brooks, Y.; Ostano, P.; Cario-André, M.; Calpini, V.; Guinea-Viniegra, J.; Albinger-Hegyi, A.; Hoetzenecker, W.; Kolfschoten, I.; Wagner, E.F.; et al. A miR-34a-SIRT6 axis in the squamous cell differentiation network. EMBO J. 2013, 32, 2248–2263. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Peterson, L.M.; Ndiaye, M.A.; Singh, C.K.; Chhabra, G.; Huang, W.; Ahmad, N. SIRT6 histone deacetylase functions as a potential oncogene in human melanoma. Genes Cancer 2017, 8, 701–712. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Guo, W.; Ma, J.; Dai, W.; Liu, L.; Guo, S.; Chen, J.; Wang, H.; Yang, Y.; Chunying, L.; et al. Aberrant SIRT6 expression contributes to melanoma growth: Role of the autophagy paradox and IGF-AKT signaling. Autophagy 2017, 14, 518–533. [Google Scholar] [CrossRef] [Green Version]
- Azuma, Y.; Yokobori, T.; Mogi, A.; Altan, B.; Yajima, T.; Kosaka, T.; Onozato, R.; Yamaki, E.; Asao, T.; Nishiyama, M.; et al. SIRT6 expression is associated with poor prognosis and chemosensitivity in patients with non-small cell lung cancer. J. Surg. Oncol. 2015, 112, 231–237. [Google Scholar] [CrossRef]
- Qu, N.; Hu, J.; Zhang, T.; Sun, G.; Shi, R.-L.; Ji, Q.; Liu, L. SIRT6 is upregulated and associated with cancer aggressiveness in papillary thyroid cancer via BRAF/ERK/Mcl-1 pathway. Int. J. Oncol. 2017, 50, 1683–1692. [Google Scholar] [CrossRef] [Green Version]
- Bae, J.S.; Park, S.-H.; Jamiyandorj, U.; Kim, K.M.; Noh, S.J.; Kim, J.R.; Park, H.J.; Kwon, K.S.; Jung, S.H.; Park, H.S.; et al. CK2α/CSNK2A1 Phosphorylates SIRT6 and Is Involved in the Progression of Breast Carcinoma and Predicts Shorter Survival of Diagnosed Patients. Am. J. Pathol. 2016, 186, 3297–3315. [Google Scholar] [CrossRef] [Green Version]
- Guan, D.; Lim, J.H.; Peng, L.; Liu, Y.; Lam, M.; Seto, E.; Kao, H.-Y. Deacetylation of the tumor suppressor protein PML regulates hydrogen peroxide-induced cell death. Cell Death Dis. 2014, 5, e1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, L.; Ji, Y.; Bakiri, L.; Qiu, Z.; Cen, J.; Chen, X.; Chen, L.; Scheuch, H.; Zheng, H.; Qin, L.; et al. Liver cancer initiation is controlled by AP-1 through SIRT6-dependent inhibition of survivin. Nat. Cell Biol. 2012, 14, 1203–1211. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, A.; Das, S. SIRT6 deacetylates PKM2 to suppress its nuclear localization and oncogenic functions. Proc. Natl. Acad. Sci. USA 2016, 113, E538–E547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.-G.; Qin, C.-Y. Sirt6 suppresses hepatocellular carcinoma cell growth via inhibiting the extracellular signal-regulated kinase signaling pathway. Mol. Med. Rep. 2013, 9, 882–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohli, S.; Bhardwaj, A.; Kumari, R.; Das, S. SIRT6 Is a Target of Regulation by UBE3A That Contributes to Liver Tumorigenesis in an ANXA2-Dependent Manner. Cancer Res. 2017, 78, 645. [Google Scholar] [CrossRef] [Green Version]
- Sebastián, C.; Zwaans, B.M.M.; Silberman, D.M.; Gymrek, M.; Goren, A.; Zhong, L.; Ram, O.; Truelove, J.; Guimaraes, A.R.; Toiber, D.; et al. The Histone Deacetylase SIRT6 Is a Tumor Suppressor that Controls Cancer Metabolism. Cell 2012, 151, 1185–1199. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.; Yang, H.; Tan, C.; Li, J.; Liu, Z.; Quan, Q.; Kong, S.; Ye, J.; Gao, B.; Fang, D. USP10 Antagonizes c-Myc Transcriptional Activation through SIRT6 Stabilization to Suppress Tumor Formation. Cell Rep. 2013, 5, 1639–1649. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Tian, H.; Xue, L.; Wang, L. SIRT6 abrogation promotes adrenocortical carcinoma through activation of NF-κB signaling. Mol. Cell. Biochem. 2019, 458, 1–10. [Google Scholar] [CrossRef]
- Feng, J.; Yan, P.-F.; Zhao, H.-Y.; Zhang, F.-C.; Zhao, W.-H.; Feng, M. SIRT6 suppresses glioma cell growth via induction of apoptosis, inhibition of oxidative stress and suppression of JAK2/STAT3 signaling pathway activation. Oncol. Rep. 2015, 35, 1395–1402. [Google Scholar] [CrossRef] [Green Version]
- Han, Z.; Liu, L.; Liu, Y.; Li, S. Sirtuin SIRT6 suppresses cell proliferation through inhibition of Twist1 expression in non-small cell lung cancer. Int. J. Clin. Exp. Pathol. 2014, 7, 4774–4781. [Google Scholar]
- Zhang, J.; Yin, X.-J.; Xu, C.-J.; Ning, Y.-X.; Chen, M.; Zhang, H.; Chen, S.-F.; Yao, L.-Q. The histone deacetylase SIRT6 inhibits ovarian cancer cell proliferation via down-regulation of Notch 3 expression. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 818–824. [Google Scholar] [PubMed]
- Kugel, S.; Sebastián, C.; Fitamant, J.; Ross, K.N.; Saha, S.K.; Jain, E.; Gladden, A.; Arora, K.S.; Kato, Y.; Rivera, M.N.; et al. SIRT6 Suppresses Pancreatic Cancer through Control of Lin28b. Cell 2016, 165, 1401–1415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strub, T.; Ghiraldini, F.G.; Carcamo, S.; Li, M.; Wroblewska, A.; Singh, R.; Goldberg, M.S.; Hasson, D.; Wang, Z.; Gallagher, S.J.; et al. SIRT6 haploinsufficiency induces BRAFV600E melanoma cell resistance to MAPK inhibitors via IGF signalling. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Choe, M.; Brusgard, J.L.; Chumsri, S.; Bhandary, L.; Zhao, X.F.; Lu, S.; Goloubeva, O.G.; Polster, B.M.; Fiskum, G.; Girnun, G.D.; et al. The RUNX2 Transcription Factor Negatively Regulates SIRT6 Expression to Alter Glucose Metabolism in Breast Cancer Cells. J. Cell. Biochem. 2015, 116, 2210–2226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, W.; Jing, Z.X.; Ke, Z.; Yi, P. Sirtuin 7 plays an oncogenic role in human osteosarcoma via downregulating CDC4 expression. Am. J. Cancer Res. 2017, 7, 1788–1803. [Google Scholar] [PubMed]
- Wang, H.-L.; Lu, R.-Q.; Xie, S.-H.; Zheng, H.; Wen, X.-M.; Gao, X.; Guo, L. SIRT7 Exhibits Oncogenic Potential in Human Ovarian Cancer Cells. Asian Pac. J. Cancer Prev. 2015, 16, 3573–3577. [Google Scholar] [CrossRef] [Green Version]
- Aljada, A.; Saleh, A.M.; Alkathiri, M.; Shamsa, H.B.; Al-Bawab, A.; Nasr, A. Altered Sirtuin 7 Expression is Associated with Early-Stage Breast Cancer. Breast Cancer Basic Clin. Res. 2015, 9, 3–8. [Google Scholar] [CrossRef]
- Zhang, S.; Chen, P.; Huang, Z.; Hu, X.; Chen, M.; Hu, S.; Hu, Y.; Cai, T. Sirt7 promotes gastric cancer growth and inhibits apoptosis by epigenetically inhibiting miR-34a. Sci. Rep. 2015, 5, 9787. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.K.; Noh, J.H.; Jung, K.H.; Eun, J.W.; Bae, H.J.; Kim, M.G.; Chang, Y.G.; Shen, Q.; Park, W.S.; Lee, J.Y.; et al. Sirtuin7 oncogenic potential in human hepatocellular carcinoma and its regulation by the tumor suppressors MiR-125a-5p and MiR-125b. Hepatology 2013, 57, 1055–1067. [Google Scholar] [CrossRef]
- Malik, S.; Villanova, L.; Tanaka, S.; Aonuma, M.; Roy, N.; Berber, E.; Pollack, J.R.; Michishita-Kioi, E.; Chua, K.F. SIRT7 inactivation reverses metastatic phenotypes in epithelial and mesenchymal tumors. Sci. Rep. 2015, 5, 9841. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Zhu, D.; Qin, S. SIRT7 suppresses the epithelial-to-mesenchymal transition in oral squamous cell carcinoma metastasis by promoting SMAD4 deacetylation. J. Exp. Clin. Cancer Res. 2018, 37, 148. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Shi, L.; Xie, N.; Liu, Z.; Qian, M.; Meng, F.; Xu, Q.; Zhou, M.; Cao, X.; Zhu, W.-G.; et al. SIRT7 antagonizes TGF-β signaling and inhibits breast cancer metastasis. Nat. Commun. 2017, 8, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa-Machado, L.F.; Fernández-Marcos, P.J. The sirtuin family in cancer. Cell Cycle 2019, 18, 2164–2196. [Google Scholar] [CrossRef] [PubMed]
- Intagliata, S.; Modica, M.N.; Santagati, L.M.; Montenegro, L. Strategies to Improve Resveratrol Systemic and Topical Bioavailability: An Update. Antioxidants 2019, 8, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sergides, C.; Chirilă, M.; Silvestro, L.; Pitta, D.; Pittas, A. Bioavailability and safety study of resveratrol 500 mg tablets in healthy male and female volunteers. Exp. Ther. Med. 2016, 11, 164–170. [Google Scholar] [CrossRef] [Green Version]
- Gambini, J.; Inglés, M.; Olaso, G.; Lopezgrueso, R.; Bonetcosta, V.; Gimenomallench, L.; Mas-Bargues, C.; Abdelaziz, K.M.; Gomezcabrera, M.C.; Vina, J.; et al. Properties of Resveratrol: In Vitro and in Vivo Studies about Metabolism, Bioavailability, and Biological Effects in Animal Models and Humans. Oxidative Med. Cell. Longev. 2015, 2015, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Springer, M.; Moco, S. Resveratrol and Its Human Metabolites—Effects on Metabolic Health and Obesity. Nutrients 2019, 11, 143. [Google Scholar] [CrossRef] [Green Version]
- Leifer, A.; Barberio, D.M. Direct ingestion method for enhancing production and bioavailability of resveratrol and other phytoalexins in Vitis vinifera. Med. Hypotheses 2016, 88, 1–5. [Google Scholar] [CrossRef]
- Tsai, H.-Y.; Ho, C.-T.; Chen, Y.K. Biological actions and molecular effects of resveratrol, pterostilbene, and 3′-hydroxypterostilbene. J. Food Drug Anal. 2017, 25, 134–147. [Google Scholar] [CrossRef] [Green Version]
- Draijer, R.; van Dorsten, F.A.; Zebregs, Y.E.; Hollebrands, B.; Peters, S.; Duchateau, G.; Grün, C.H. Impact of Proteins on the Uptake, Distribution, and Excretion of Phenolics in the Human Body. Nutrients 2016, 8, 814. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Sang, S. Metabolism and pharmacokinetics of resveratrol and pterostilbene. BioFactors 2018, 44, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Garza, S.L.; Laveriano-Santos, E.P.; Marhuenda-Muñoz, M.; Storniolo, C.E.; Tresserra-Rimbau, A.; Vallverdú-Queralt, A.; Lamuela-Raventós, R.M. Health Effects of Resveratrol: Results from Human Intervention Trials. Nutrients 2018, 10, 1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Zhang, G.; Li, C.; Wang, S.; Zhu, M.; Wang, J.; Yue, H.; Ma, X.; Zhen, Y.; Shu, X. Metabolic profile and structure–activity relationship of resveratrol and its analogs in human bladder cancer cells. Cancer Manag. Res. 2019, 11, 4631–4642. [Google Scholar] [CrossRef] [Green Version]
- Aires, V.; Limagne, E.; Cotte, A.K.; Latruffe, N.; Ghiringhelli, F.; Delmas, D. Resveratrol metabolites inhibit human metastatic colon cancer cells progression and synergize with chemotherapeutic drugs to induce cell death. Mol. Nutr. Food Res. 2013, 57, 1170–1181. [Google Scholar] [CrossRef] [PubMed]
- Guthrie, A.R.; Chow, H.-H.S.; Miller, J.A. Effects of resveratrol on drug- and carcinogen-metabolizing enzymes, implications for cancer prevention. Pharmacol. Res. Perspect. 2017, 5, e00294. [Google Scholar] [CrossRef]
- Pannu, N.; Bhatnagar, A. Resveratrol: From enhanced biosynthesis and bioavailability to multitargeting chronic diseases. Biomed. Pharmacother. 2019, 109, 2237–2251. [Google Scholar] [CrossRef]
- Patel, K.R.; Brown, V.A.; Jones, D.J.L.; Britton, R.G.; Hemingway, D.; Miller, A.S.; West, K.P.; Booth, T.D.; Perloff, M.; Crowell, J.A.; et al. Clinical Pharmacology of Resveratrol and Its Metabolites in Colorectal Cancer Patients. Cancer Res. 2010, 70, 7392–7399. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-J.; Yim, D.-G.; Hur, S.J. Changes in the Content and Bioavailability of Onion Quercetin and Grape Resveratrol during In Vitro Human Digestion. Foods 2020, 9, 694. [Google Scholar] [CrossRef]
- Brown, V.A.; Patel, K.R.; Viskaduraki, M.; Crowell, J.A.; Perloff, M.; Booth, T.D.; Vasilinin, G.; Sen, A.; Schinas, A.M.; Piccirilli, G.; et al. Repeat Dose Study of the Cancer Chemopreventive Agent Resveratrol in Healthy Volunteers: Safety, Pharmacokinetics, and Effect on the Insulin-like Growth Factor Axis. Cancer Res. 2010, 70, 9003–9011. [Google Scholar] [CrossRef] [Green Version]
- Almeida, L.; Vaz-Da-Silva, M.; Falcão, A.; Soares, E.; Costa, R.; Loureiro, A.I.; Fernandes-Lopes, C.; Rocha, J.-F.; Nunes, T.; Wright, L.; et al. Pharmacokinetic and safety profile of trans-resveratrol in a rising multiple-dose study in healthy volunteers. Mol. Nutr. Food Res. 2009, 53, S7–S15. [Google Scholar] [CrossRef]
- Vasconcelos, T.; Araújo, F.; Lopes, C.; Loureiro, A.; Das Neves, J.; Marques, S.; Sarmento, B. Multicomponent self-nano emulsifying delivery systems of resveratrol with enhanced pharmacokinetics profile. Eur. J. Pharm. Sci. 2019, 137, 105011. [Google Scholar] [CrossRef]
- Calvo-Castro, L.A.; Schiborr, C.; David, F.; Ehrt, H.; Voggel, J.; Sus, N.; Behnam, D.; Bosy-Westphal, A.; Frank, J. The Oral Bioavailability of Trans-Resveratrol from a Grapevine-Shoot Extract in Healthy Humans is Significantly Increased by Micellar Solubilization. Mol. Nutr. Food Res. 2018, 62, e1701057. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.C.; Veiga, F.; Sequeira, J.A.D.; Fortuna, A.; Falcão, A.; Souto, E.B.; Pattekari, P.; Ribeiro, C.F.; Ribeiro, A.J. First-time oral administration of resveratrol-loaded layer-by-layer nanoparticles to rats—A pharmacokinetics study. Analyst 2019, 144, 2062–2079. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Wang, Y.; Xie, Y.; Liu, G.; Lu, Y.; Wu, W.; Chen, L. Oat protein-shellac nanoparticles as a delivery vehicle for resveratrol to improve bioavailability in vitro and in vivo. Nanomedicine 2019, 14, 2853–2871. [Google Scholar] [CrossRef] [PubMed]
- Peñalva, R.; Morales, J.; González-Navarro, C.J.; Larrañeta, E.; Quincooces, G.; Peñuelas, I.; Irache, J. (Juan M. Increased Oral Bioavailability of Resveratrol by Its Encapsulation in Casein Nanoparticles. Int. J. Mol. Sci. 2018, 19, 2816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.K.; Makadia, V.; Sharma, S.; Rashid, M.; Shahi, S.; Mishra, P.R.; Wahajuddin, M.; Gayen, J.R. Preparation and in-vitro/in-vivo characterization of trans-resveratrol nanocrystals for oral administration. Drug Deliv. Transl. Res. 2017, 7, 395–407. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Zhong, C.; Deng, Y.; Zhang, Q.; Zhang, X.; Zhao, X. Resveratrol loaded glycyrrhizic acid-conjugated human serum albumin nanoparticles for tail vein injection II: Pharmacokinetics, tissue distribution and bioavailability. Drug Deliv. 2020, 27, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Katekar, R.; Thombre, G.; Riyazuddin, M.; Husain, A.; Rani, H.; Praveena, K.S.; Gayen, J.R. Pharmacokinetics and brain targeting of trans-resveratrol loaded mixed micelles in rats following intravenous administration. Pharm. Dev. Technol. 2019, 25, 300–307. [Google Scholar] [CrossRef]
- Guo, L.; Peng, Y.; Yao, J.; Sui, L.; Gu, A.; Wang, J. Anticancer Activity and Molecular Mechanism of Resveratrol–Bovine Serum Albumin Nanoparticles on Subcutaneously Implanted Human Primary Ovarian Carcinoma Cells in Nude Mice. Cancer Biother. Radiopharm. 2010, 25, 471–477. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.J.; Nihal, M.; Siddiqui, I.A.; Scarlett, C.O.; Bailey, H.H.; Mukhtar, H.; Ahmad, N. Enhancing the bioavailability of resveratrol by combining it with piperine. Mol. Nutr. Food Res. 2011, 55, 1169–1176. [Google Scholar] [CrossRef] [Green Version]
- de Santi, C.; Pietrabissa, A.; Spisni, R.; Mosca, F.; Pacifici, G.M. Sulphation of resveratrol, a natural compound present in wine, and its inhibition by natural flavonoids. Xenobiotica 2000, 30, 857–866. [Google Scholar] [CrossRef] [PubMed]
- Chimento, A.; de Amicis, F.; Sirianni, R.; Sinicropi, M.S.; Puoci, F.; Casaburi, I.; Saturnino, C.; Pezzi, V. Progress to Improve Oral Bioavailability and Beneficial Effects of Resveratrol. Int. J. Mol. Sci. 2019, 20, 1381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salehi, B.; Mishra, A.P.; Nigam, M.; Sener, B.; Kilic, M.; Sharifi-Rad, M.; Fokou, P.V.T.; Martins, N.; Sharifi-Rad, J. Resveratrol: A Double-Edged Sword in Health Benefits. Biomedicines 2018, 6, 91. [Google Scholar] [CrossRef] [Green Version]
- Berman, A.Y.; Motechin, R.A.; Wiesenfeld, M.Y.; Holz, M.K. The therapeutic potential of resveratrol: A review of clinical trials. NPJ Precis. Oncol. 2017, 1, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, H.-H.S.; Garland, L.L.; Hsu, C.-H.; Vining, D.R.; Chew, W.M.; Miller, J.A.; Perloff, M.; Crowell, J.A.; Alberts, D.S. Resveratrol Modulates Drug- and Carcinogen-Metabolizing Enzymes in a Healthy Volunteer Study. Cancer Prev. Res. 2010, 3, 1168–1175. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.; Qin, W.; Zhang, K.; Rottinghaus, G.E.; Chen, Y.-C.; Kliethermes, B.; Sauter, E.R. Trans-Resveratrol Alters Mammary Promoter Hypermethylation in Women at Increased Risk for Breast Cancer. Nutr. Cancer 2012, 64, 393–400. [Google Scholar] [CrossRef] [Green Version]
- Chow, H.-H.S.; Garland, L.L.; Heckman-Stoddard, B.M.; Hsu, C.-H.; Butler, V.D.; A Cordova, C.; Chew, W.M.; Cornelison, T.L. A pilot clinical study of resveratrol in postmenopausal women with high body mass index: Effects on systemic sex steroid hormones. J. Transl. Med. 2014, 12, 223. [Google Scholar] [CrossRef] [Green Version]
- Klement, R.J.; Fink, M.K. Dietary and pharmacological modification of the insulin/IGF-1 system: Exploiting the full repertoire against cancer. Oncogenesis 2016, 5, e193. [Google Scholar] [CrossRef] [Green Version]
- Ter-Braak, B.; Siezen, C.L.; Lee, J.S.; Rao, P.; Voorhoeve, C.; Ruppin, E.; van der Laan, J.W.; van de Water, B. Insulin-like growth factor 1 receptor activation promotes mammary gland tumor development by increasing glycolysis and promoting biomass production. Breast Cancer Res. 2017, 19, 14. [Google Scholar] [CrossRef] [Green Version]
- Banaszewska, B.; Wrotyńska-Barczyńska, J.; Spaczynski, R.Z.; Pawelczyk, L.; Duleba, A.J. Effects of Resveratrol on Polycystic Ovary Syndrome: A Double-blind, Randomized, Placebo-controlled Trial. J. Clin. Endocrinol. Metab. 2016, 101, 4322–4328. [Google Scholar] [CrossRef]
- Mo, Y.; Wang, Y.; Zhang, L.; Yang, L.; Zhou, M.; Li, X.; Li, Y.; Li, G.; Zeng, Z.; Xiong, W.; et al. The role of Wnt signaling pathway in tumor metabolic reprogramming. J. Cancer 2019, 10, 3789–3797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashrafizadeh, M.; Ahmadi, Z.; Farkhondeh, T.; Samarghandian, S. Resveratrol targeting the Wnt signaling pathway: A focus on therapeutic activities. J. Cell. Physiol. 2020, 235, 4135–4145. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, A.V.; Martinez, M.; Stamos, M.J.; Moyer, M.P.; Planutis, K.; Hope, C.; Holcombe, R.F. Results of a phase I pilot clinical trial examining the effect of plant-derived resveratrol and grape powder on Wnt pathway target gene expression in colonic mucosa and colon cancer. Cancer Manag. Res. 2009, 1, 25–37. [Google Scholar] [PubMed]
- Turner, R.S.; Thomas, R.G.; Craft, S.; van Dyck, C.H.; Mintzer, J.; Reynolds, B.A.; Brewer, J.B.; Rissman, R.A.; Raman, R.; Aisen, P.S.; et al. A randomized, double-blind, placebo-controlled trial of resveratrol for Alzheimer disease. Neurology 2015, 85, 1383–1391. [Google Scholar] [CrossRef] [PubMed]
- la Porte, C.J.L.; Voduc, N.; Zhang, G.; Seguin, I.; Tardiff, D.; Singhal, N.; Cameron, D.W. Steady-State Pharmacokinetics and Tolerability of Trans-Resveratrol 2000 mg Twice Daily with Food, Quercetin and Alcohol (Ethanol) in Healthy Human Subjects. Clin. Pharmacokinet. 2010, 49, 449–454. [Google Scholar] [CrossRef]
- Samec, M.; Liskova, A.; Koklesova, L.; Samuel, S.M.; Zhai, K.; Buhrmann, C.; Varghese, E.; Abotaleb, M.; Qaradakhi, T.; Zulli, A.; et al. Flavonoids against the Warburg phenotype—Concepts of predictive, preventive and personalised medicine to cut the Gordian knot of cancer cell metabolism. EPMA J. 2020, 11, 377–398. [Google Scholar] [CrossRef]
- Ashrafizadeh, M.; Ahmadi, Z.; Farkhondeh, T.; Samarghandian, S. Autophagy as a molecular target of quercetin underlying its protective effects in human diseases. Arch. Physiol. Biochem. 2019, 10, 1–9. [Google Scholar] [CrossRef]
- Ashrafizadeh, M.; Fekri, H.S.; Ahmadi, Z.; Farkhondeh, T.; Samarghandian, S. Therapeutic and biological activities of berberine: The involvement of Nrf2 signaling pathway. J. Cell. Biochem. 2020, 121, 1575–1585. [Google Scholar] [CrossRef]
- Abotaleb, M.; Samuel, S.M.; Varghese, E.; Varghese, S.; Kubatka, P.; Líšková, A.; Büsselberg, D. Flavonoids in Cancer and Apoptosis. Cancers 2018, 11, 28. [Google Scholar] [CrossRef] [Green Version]
- Liskova, A.; Kubatka, P.; Samec, M.; Zubor, P.; Mlyncek, M.; Bielik, T.; Samuel, S.M.; Zulli, A.; Kwon, T.K.; Büsselberg, D. Dietary Phytochemicals Targeting Cancer Stem Cells. Molecules 2019, 24, 899. [Google Scholar] [CrossRef] [Green Version]
- Kubatka, P.; Uramova, S.; Kello, M.; Kajo, K.; Samec, M.; Jašek, K.; Vybohova, D.; Liskova, A.; Mojzis, J.; Adamkov, M.; et al. Anticancer Activities of Thymus vulgaris L. in Experimental Breast Carcinoma in Vivo and in Vitro. Int. J. Mol. Sci. 2019, 20, 1749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liskova, A.; Stefanicka, P.; Samec, M.; Smejkal, K.; Zubor, P.; Bielik, T.; Biskupska-Bodova, K.; Kwon, T.K.; Danko, J.; Büsselberg, D.; et al. Dietary phytochemicals as the potential protectors against carcinogenesis and their role in cancer chemoprevention. Clin. Exp. Med. 2020, 20, 173–190. [Google Scholar] [CrossRef] [PubMed]
- Liskova, A.; Koklesova, L.; Samec, M.; Smejkal, K.; Samuel, S.M.; Varghese, E.; Abotaleb, M.; Biringer, K.; Kudela, E.; Danko, J.; et al. Flavonoids in Cancer Metastasis. Cancers 2020, 12, 1498. [Google Scholar] [CrossRef] [PubMed]
- Buhrmann, C.; Popper, B.; Aggarwal, B.B.; Shakibaei, M. Resveratrol downregulates inflammatory pathway activated by lymphotoxin α (TNF-β) in particular chondrocytes: Comparison with TNF-α. PLoS ONE 2017, 12, e0186993. [Google Scholar] [CrossRef] [PubMed]
- Buhrmann, C.; Shayan, P.; Banik, K.; Kunnumakkara, A.B.; Kubatka, P.; Koklesova, L.; Shakibaei, M. Targeting NF-kB Signaling by Calebin A, a Compound of Turmeric, in Multicellular Tumor Microenvironment: Potential Role of Apoptosis Induction in CRC Cells. Biomedicines 2020, 8, 236. [Google Scholar] [CrossRef] [PubMed]
- Buhrmann, C.; Yazdi, M.; Popper, B.; Shayan, P.; Goel, A.; Aggarwal, B.B.; Shakibaei, M. Evidence that TNF-β induces proliferation in colorectal cancer cells and resveratrol can down-modulate it. Exp. Biol. Med. 2019, 244, 1–12. [Google Scholar] [CrossRef]
- Buhrmann, C.; Kraehe, P.; Lueders, C.; Shayan, P.; Goel, A.; Shakibaei, M. Curcumin Suppresses Crosstalk between Colon Cancer Stem Cells and Stromal Fibroblasts in the Tumor Microenvironment: Potential Role of EMT. PLoS ONE 2014, 9, e107514. [Google Scholar] [CrossRef] [Green Version]
- Shakibaei, M.; Buhrmann, C.; Kraehe, P.; Shayan, P.; Lueders, C.; Goel, A. Curcumin Chemosensitizes 5-Fluorouracil Resistant MMR-Deficient Human Colon Cancer Cells in High Density Cultures. PLoS ONE 2014, 9, e85397. [Google Scholar] [CrossRef] [Green Version]
- Shakibaei, M.; Kraehe, P.; Popper, B.; Shayan, P.; Goel, A.; Buhrmann, C. Curcumin potentiates antitumor activity of 5-fluorouracil in a 3D alginate tumor microenvironment of colorectal cancer. BMC Cancer 2015, 15, 250. [Google Scholar] [CrossRef] [Green Version]
- Shakibaei, M.; Mobasheri, A.; Lueders, C.; Busch, F.; Shayan, P.; Goel, A. Curcumin Enhances the Effect of Chemotherapy against Colorectal Cancer Cells by Inhibition of NF-κB and Src Protein Kinase Signaling Pathways. PLoS ONE 2013, 8, e57218. [Google Scholar] [CrossRef] [Green Version]
- Toden, S.; Okugawa, Y.; Jascur, T.; Wodarz, D.; Komarova, N.L.; Buhrmann, C.; Shakibaei, M.; Boland, C.R.; Goel, A. Curcumin mediates chemosensitization to 5-fluorouracil through miRNA-induced suppression of epithelial-to-mesenchymal transition in chemoresistant colorectal cancer. Carcinogenesis 2015, 36, 355–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]




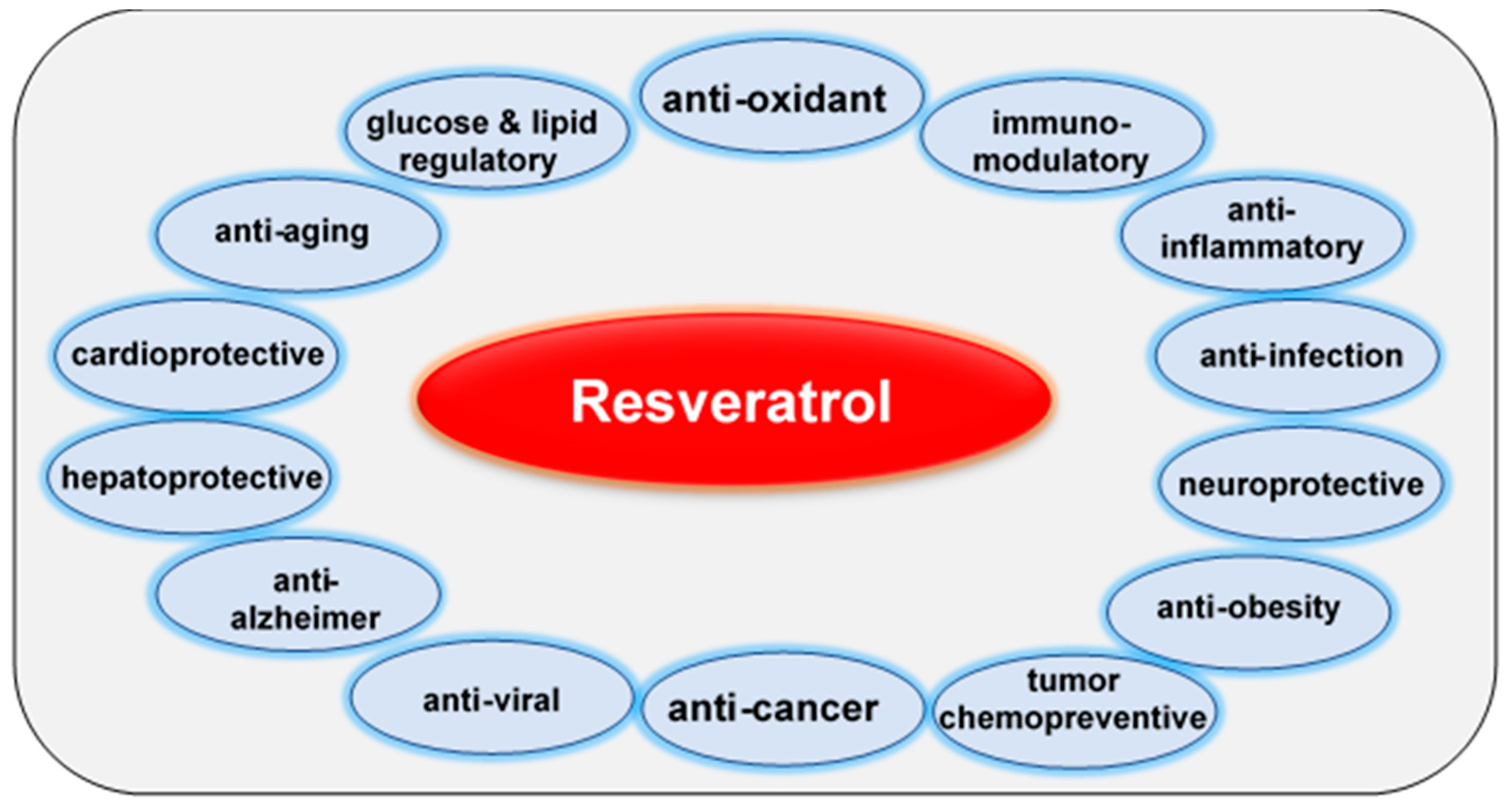{kind=link}
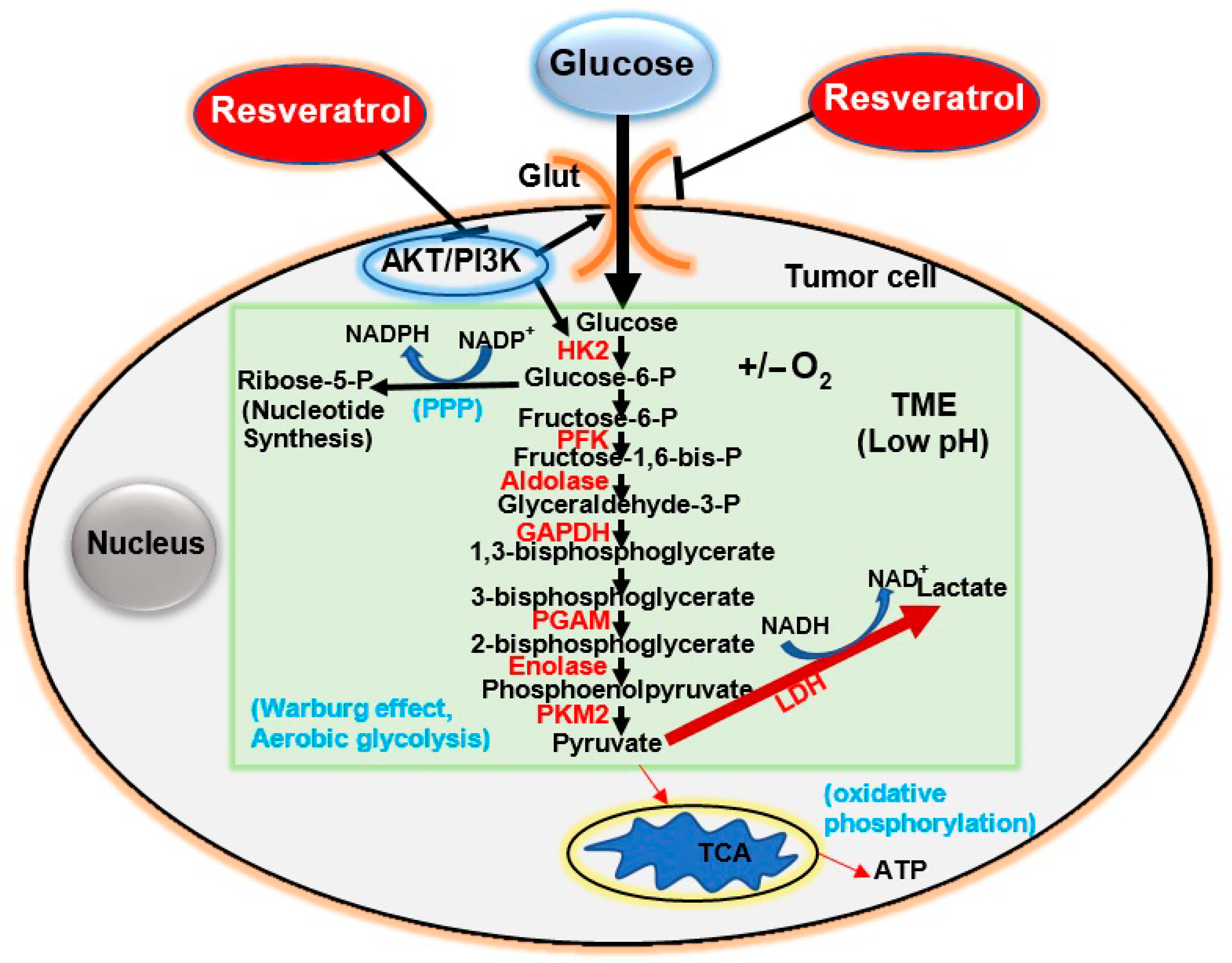{kind=link}
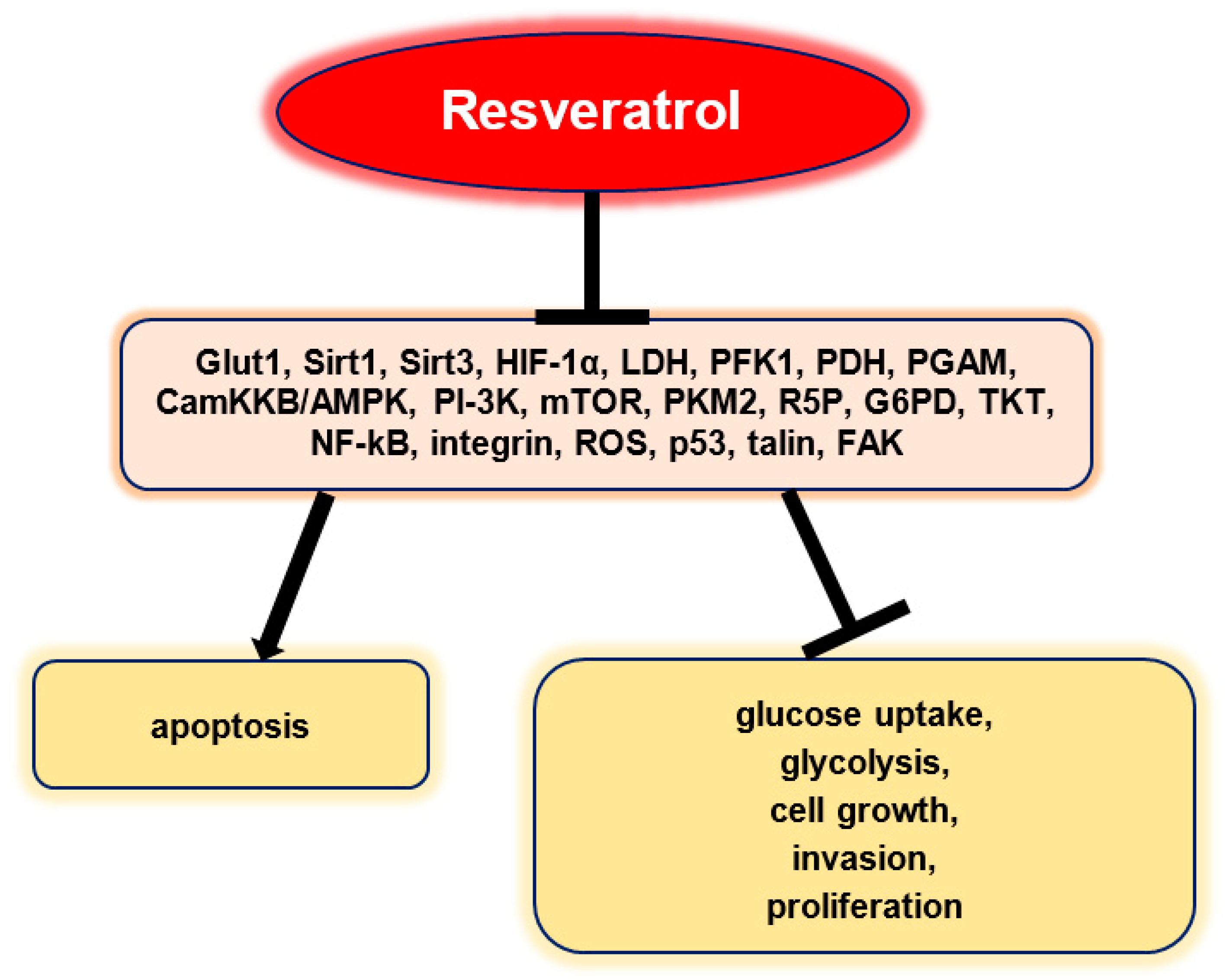{kind=link}
| Type of Tumor | Tumor Cells | Mode of Action | Reference |
|---|---|---|---|
| Breast cancer | MCF-7 breast cancer cells. | Suppresses PFK activity, disrupting glucose metabolism and reducing viability in cancer cells. | [15] |
| T47D cells, BALB/c-ν mice. | Suppresses cancer cell 18F-FDG uptake and glycolytic metabolism, reduces intracellular ROS, and downregulates HIF-1α accumulation. | [17] | |
| MDA-MB-231 breast cancer cells. | Induces apoptosis and inhibits growth by activating the de novo ceramide synthesis pathway. | [91] | |
| MCF-7 breast cancer cells. | Antiproliferative and cytotoxic effects by decreasing Glut1-mediated glucose uptake. | [92] | |
| Lung cancer | Lewis lung carcinoma cells, BALB/c-ν mice. | Suppresses glucose uptake by targeting ROS-mediated HIF-1α activation. | [17] |
| NSCLC, xenograft mouse model. | Impairs HK2-mediated glycolysis and inhibits anchorage-dependent and -independent growth of cells. | [93] | |
| Human lung carcinoma A549 cells. | Increases glucocerebrosidase expression, intracellular ceramide levels, and apoptosis. | [94] | |
| Human bronchial epithelial cell line BEP2D. | Inhibits the induced expression of CYP1A1 and CYP1B1; changes the formation and metabolism of carcinogenic benzo[a]pyrene metabolites. | [95] | |
| Colon cancer (CRC) | HT-29 cells, BALB/c-ν mice. | Suppresses glucose uptake by targeting ROS-mediated HIF-1α activation. | [17] |
| HCT116 and Caco2 cells. | Downregulates glucose uptake, glycolytic enzymes (PK, LDH), and VEGF; induces apoptosis. | [96] | |
| CT26 cells, CT26 tumor bearing mice. | Suppresses tumor growth and glucose uptake; increases apoptosis. | [97] | |
| Caco2 and HTC116 colon cancer cells. | Induces oxidative capacities through the CamKKB/AMPK pathway; increases PDH activity. | [98] | |
| HT-29 colon cancer cells. | Suppresses proliferation and induces apoptosis by targeting PPP and the talin-pFAK. | [99] | |
| Prostate cancer | PC3 prostate cancer cells. | Inhibits growth via a metabolic shift from glucose fermentation to mitochondrial respiration. | [100] |
| PC3 prostate cancer cells. | Suppresses tumor growth by interfering with glucose fermentation and thereby promoting oxidative respiration. | [101] | |
| Ovarian cancer | SKOV3 and CaOV3 Ovarian cancer cells. | Reduces glucose uptake and lactate production via suppression of the Akt/mTOR pathway. | [18] |
| Ovarian cancer cells. | Inhibits glucose uptake and induces apoptosis by impairing the Akt/Glut1 axis. | [102] | |
| A2780 and SKOV3 ovarian cancer cells. | Anti-proliferative, pro-apoptotic effects through the inhibition of glycolysis and targeting of the AMPK/mTOR pathway. | [103] | |
| Preclinical mouse model of ovarian cancer. | Inhibits glucose uptake with antineoplastic effects; suppresses tumor regrowth after therapy with cisplatin. | [104] | |
| Leukemia | B-CLL and HCL lymphocytic leukemia. | Decreases the mitochondrial transmembrane potential, inhibits proliferation and growth, and induces apoptosis. | [105,106] |
| CD95-sensitive leukemia cells, B-leukemic cells. | Promotes apoptosis by depolarizing mitochondrial membranes and activating caspase-9. | [107] | |
| Liver cancer | HepG2 cells. | Suppresses glucose uptake via the Sirt1-dependent p-STAT3 signaling pathway. | [108] |
| HCC, HepG2, Bel-7402, and SMMC-7721 cells. | Suppresses proliferation and migration via Sirt1-mediated post-translational modifications of the PI3K/Akt pathway. | [109] | |
| Pancreatic cancer | Panc-1 human pancreatic cancer cells. | Suppresses ROS-induced hyperglycemia; inhibits the ERK and p38-MAPK signaling pathways. | [110] |
| Pancreatic cancer cells. | Suppresses proliferation and glucose uptake by targeting HIF-1α. | [111] | |
| Pancreatic cancer cells. | Suppresses migration by downregulating ROS/miR-21-mediated activation and glycolysis in PSCs. | [112] |
| Experimental Model | Study Type | Finding | Reference |
|---|---|---|---|
| Nuclear magnetic resonance spectroscopy identified glycolysis as the primary glucose catabolic pathway. | In vitro/LY18 human diffuse large B-cell lymphoma cells. | Inhibits PI-3K signaling and glucose metabolism; induces cell cycle arrest. | [14] |
| Effects of resveratrol on PKM2 expression, and effects on cancer metabolism. | In vitro/HeLa, HepG2, and MCF-7 cells. | Downregulates PKM2 by inhibiting mTOR signaling, reduces glucose uptake, lactate production, and R5P. | [16] |
| Glycolytic metabolism, lactate production assay, hexokinase activity assay, intracellular RO assay, and 18F-FDG uptake. | In vitro/in vivo LLC, HT-29, T47D cells, BALB/c-v mice. | Reduces 18F-FDG uptake, glycolytic metabolism, intracellular ROS, and HIF-1α accumulation. | [17] |
| Growth inhibition assay; [3H]-2-deoxyglucose uptake and lactate assays. | In vitro/multiple human ovarian carcinoma cells. | Induces autophagy; inhibits glycolysis. | [18] |
| Glucose metabolism regulation via Glut1 modulation. | In vitro/human ovarian cancer cells. | Promotes apoptosis by impairing glucose uptake, involving Akt-regulated membrane Glut1 trafficking. | [102] |
| Metabolic and anti-tumor effects of resveratrol. | In vitro/in vivo CT26 colon cancer cells; tumor bearing mice. | Resveratrol-nanoparticles (NP) increase apoptosis and reduce 18F FDG uptake and ROS. | [97] |
| The effects of resveratrol on glucose metabolism. | In vitro/in vivo human ovarian cancer cells; murine xenograft model. | Inhibits glycolysis and glucose uptake by activating AMPK/mTOR; inhibits growth and metastasis. | [103] |
| The effects of resveratrol on glucose uptake and accumulation, and Glut1. | In vitro/HL-60 and U-937 leukemic cell lines. | Resveratrol blocks Glut1-mediated hexose uptake. | [134] |
| Mice were treated with cisplatin, resveratrol, or vehicle alone. The effect of resveratrol on glucose uptake was determined using micro-positron emission. | In vivo/murine xenograft model of ovarian cancer. | Inhibits glucose uptake, with antineoplastic effects; suppresses tumor regrowth after cisplatin therapy. | [135] |
| Molecular Signaling Target | Study Type | Finding | Reference |
|---|---|---|---|
| PI3K signaling | In vitro/B cell lymphoma. | Suppresses glucose catabolism; induces growth arrest. | [14] |
| PFK1 | In vitro/human breast cancer cells. | Suppresses glucose uptake and tumor viability. | [15] |
| mTOR, PKM2, R5P | In vitro/HeLa, HepG2, and MCF-7 cells. | Suppresses glucose uptake and growth. | [16] |
| HIF-1α/ROS signaling | In vitro/in vivo HT-29 cells; BALB/c-ν mice. | Suppresses glycolytic metabolism. | [17] |
| Glut1 | In vitro/in vivo HL-60, U-937, and HT-29 cells; BALB/c-ν mice. | Suppresses glucose uptake. | [17,33,134] |
| FAK, NF-κB, Integrin | In vitro/human CRC cells. | Induces apoptosis. | [119] |
| Sirt1, PDH, Glut1, NF-κB | In vitro/human CRC cells. | Suppresses glucose uptake and invasion; induces apoptosis. | [120] |
| Glut1, LDH, Sirt1, Sirt3 | In vitro/human melanoma cells. | Suppresses proliferation; induces cell cycle arrest and apoptosis. | [127] |
| PDH, CamKKB/AMPK | In vitro/human CRC cells. | Induces oxidative capacities; inhibits glycolysis. | [98] |
| G6PD, TKT, Talin | In vitro/human CRC cells. | Suppresses proliferation and invasion. | [99] |
| HIF-1α/ROS/p53 signaling | In vitro/prostate cancer cells. | Induces apoptosis. | [146] |
| HK2 | In vitro/in vivo. | Inhibits apoptosis. | [161] |
| PGAM | In vitro/human prostate cancer cells. | Inhibits tumor cell growth. | [162] |
| Sirtuin | Metabolic Pathways Affected |
|---|---|
| SIRT1 | Gluconeogenesis; Glycolysis; Insulin Synthesis and Secretion; Cholesterol and Fatty Acid Synthesis |
| SIRT2 | Gluconeogenesis; Triglyceride Synthesis |
| SIRT3 | Glutamine Metabolism; Ketone Body Formation; Urea Cycle; β-Oxidation of Fatty Acids |
| SIRT4 | Glutamine, Leucine, and Carbohydrate Metabolism; β-Oxidation of Fatty Acids |
| SIRT5 | Glycolysis; Tri-Carboxylic Acid Cycle; Ketone Body Formation |
| SIRT6 | Gluconeogenesis; Glycolysis; β-Oxidation of Fatty Acids |
| SIRT7 | Lipid Metabolism |
| Sirtuin | Cellular Location | Histone Targets | Enzymatic Activity | Oncogenic in Cancers | Genes/Targets as Promoter | Suppressor in Cancers | Genes/Targets as Suppressor |
|---|---|---|---|---|---|---|---|
| SIRT1 | Nucleus | H1-K26Ac H3-K9Ac H4-K16Ac H4-K20Ac H3-K9Ac | Deacetylase | TC [207,208,209], CRC [210,211], Leukemia [212,213], RB [214], Glioma [215,216], BC [217,218,219,220], PC [209,221,222], NSCLC [217,223,224], GC [225,226], HCC [227,228,229,230], OC [231,232], PAC [233,234], SSCC [235], Melanoma [236] | c-MYC, Oct4, ZEB1, Nanog, Cripto, Tert, Lin28, STAT5, EGF, FOXO1, p53, Ku70, Rb, EMT pathway, Akt, KRas, PI3K, BRCA1, Survivin, DBC1 | BC [237,238], NSCLC [239], HCC [209,237], OC [231,237], GC [240], PC [237], CRC [241,242], PAC [243], GB [237] | BRCA1, KRas, PI3K, Smad4/β-catenin, RCA1, Survivin |
| SIRT2 | Cytoplasm | H3-K18Ac H3-K56Ac H4-K16Ac | Deacetylase, Demyristoylase | HCC [244,245], NB [246], PAC [246], GC, BC [247,248], NSCLC [249], RCC [250], GB [251], SSCC [235], Melanoma [252] | Slug, α-tubulin, c-MYC, Akt/GSK3β/β-catenin axis | OC [253], BC [254], NSCLC [255,256,257,258,259], CCA [260], HCC [254], PC [261], CRC [262], PAC [258,263], SSCC [264], SBCC [265] | CDK4, APC, CDC20, p53, c-MYC, PKM2, KRas |
| SIRT3 | Mitochondria | Deacetylase | BLC [266], CRC [267,268], NSCLC [269], Leukemia [270], OSCC [270], OC [271], SSCC [235], Melanoma [272], BC [273] | p53, Akt/PTEN, Bax/Bcl2, Akt, RIP, SHMT2, GDH | PC [274,275], HCC [276,277,278,279], PDC [280], OSCC [281], OC [271], TC [282], GC [283,284,285], PAC [286], SBCC [265], BC [287,288,289] | c-MYC, PI3K/Akt, mTOR, Bax/Bcl2, p53, RIP, FOXO3A, Wnt/β-catenin, Twist, HIF-1α, SOD, IDH2, Notch1 | |
| SIRT4 | Mitochondria | ADP-ribosylase | SSCC [235], BC [290] | NSCLC [291,292,293], ESCC [294], CRC [295,296,297], GC [291,295,298], HCC [299], RCC [300], OC [291,295], NB [301], BC [291,295,302] | GDH, ERK/Drp1 axis, E-cadherin, LKB1/AMPKα/mTOR Axis | ||
| SIRT5 | Mitochondria | Deacetylase, Desuccinylase Deglutarylase Demalonylase | CRC [303,304], OSA [305], NSCLC [306,307], HCC [308], RCC [309], SSCC [235], Melanoma [310], BC [311] | Glut1, SHMT2, PKM2, E2F1, SDHA, Vimentin, GDH | NB [312], HCC [313], BC [311] | SOD | |
| SIRT6 | Nucleus | H3-K9Ac H3-K56Ac H3-K18Ac | Deacetylase ADP-ribosylase Demyristoylase Depalmitoylase | HCC [314], SSCC [315,316], Melanoma [317,318], NSCLC [319], TC [320], BC [321] | Bax, COX2, Akt, AMPK, Akt/PTEN, JAK2/STAT3, ERK1/2 | HCC [322,323,324,325,326], CRC [327,328], ACC [329], GBM [330], NSCLC [331], OC [332], PAC [327,333], Melanoma [334], BC [335] | PKM2, PTEN/Akt, NF-κB, JAK2/STAT3, Bax, HIF-1α, c-MYC, Twist (EMT), Notch3 |
| SIRT7 | Nucleus | H3-K18Ac | Deacetylase Desuccinylase | OSA [336], OC [337], BC [338], GC [339], HCC [340], PC [341], SSCC [235] | CDC4, NF-κB, p38-MAPK, Bax/Bcl2 | OSCC [342], BC [343] | SMAD4, NF-κB, p38-MAPK, TGFβ, EMT signaling |
| Delivery System | Study Details | Results | Reference |
|---|---|---|---|
| Self-emulsifying drug delivery systems of resveratrol. | In vitro, in vivo (rats). | Increased solubility, reduced metabolism, and improved its pharmacokinetic profile. | [362] |
| Micellar solubilization of resveratrol. | Twelve healthy volunteers (oral administration). | Increased oral bioavailability. | [363] |
| Layer-by-layer nanoparticles, resveratrol nanocores, and free resveratrol suspension. | In vivo (Wistar rats, oral administration, 20 mg/kg). | Layer-by-layer nanoparticles and resveratrol nanocores enhanced systemic exposure compared to free resveratrol. | [364] |
| Oat protein-shellac nanoparticle delivery system. | In vitro, in vivo (rat model). | Protected resveratrol in gastric fluid and controlled its release into the small intestine. Improved cell uptake and transport compared to free resveratrol. Increased bioavailability. | [365] |
| Encapsulation of resveratrol in casein nanoparticles. | In vitro, in vivo (rats). | Oral administration in rats: remained in the gut and reached intestinal epithelium. Produced high plasma levels of resveratrol (sustained for at least 8 h) and similar results for its metabolites. Oral bioavailability was 10 times higher compared to an oral solution of resveratrol. | [366] |
| Trans-resveratrol nanocrystals. | In vitro, in vivo (rats). | Increased oral bioavailability. | [367] |
| Resveratrol-loaded glycyrrhizic acid-conjugated human serum albumin nanoparticles. | In vivo (rats; single-dose tail vein injection). | Improved bioavailability of resveratrol. Elevated concentrations in the main organs of rats compared to pure resveratrol. Highest concentrations were observed in the liver (promising liver-targeted delivery system). | [368] |
| Trans-resveratrol-loaded mixed micelles. | In vivo (rats; intravenous administration). | Enhanced pharmacokinetic parameters. Brain targeting. | [369] |
| Resveratrol-bovine serum albumin nanoparticles (RES-BSANP). | In vivo (nude mice; intraperitoneal injection). | Improved dispersal and water solubility. Inhibited carcinoma growth in nude mice bearing human primary ovarian tumors. | [370] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brockmueller, A.; Sameri, S.; Liskova, A.; Zhai, K.; Varghese, E.; Samuel, S.M.; Büsselberg, D.; Kubatka, P.; Shakibaei, M. Resveratrol’s Anti-Cancer Effects through the Modulation of Tumor Glucose Metabolism. Cancers 2021, 13, 188. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13020188
Brockmueller A, Sameri S, Liskova A, Zhai K, Varghese E, Samuel SM, Büsselberg D, Kubatka P, Shakibaei M. Resveratrol’s Anti-Cancer Effects through the Modulation of Tumor Glucose Metabolism. Cancers. 2021; 13(2):188. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13020188
Chicago/Turabian StyleBrockmueller, Aranka, Saba Sameri, Alena Liskova, Kevin Zhai, Elizabeth Varghese, Samson Mathews Samuel, Dietrich Büsselberg, Peter Kubatka, and Mehdi Shakibaei. 2021. "Resveratrol’s Anti-Cancer Effects through the Modulation of Tumor Glucose Metabolism" Cancers 13, no. 2: 188. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13020188