
Prognostic Significance of SLFN11 Methylation in Plasma Cell-Free DNA in Advanced High-Grade Serous Ovarian Cancer

 , ,
, ,  , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
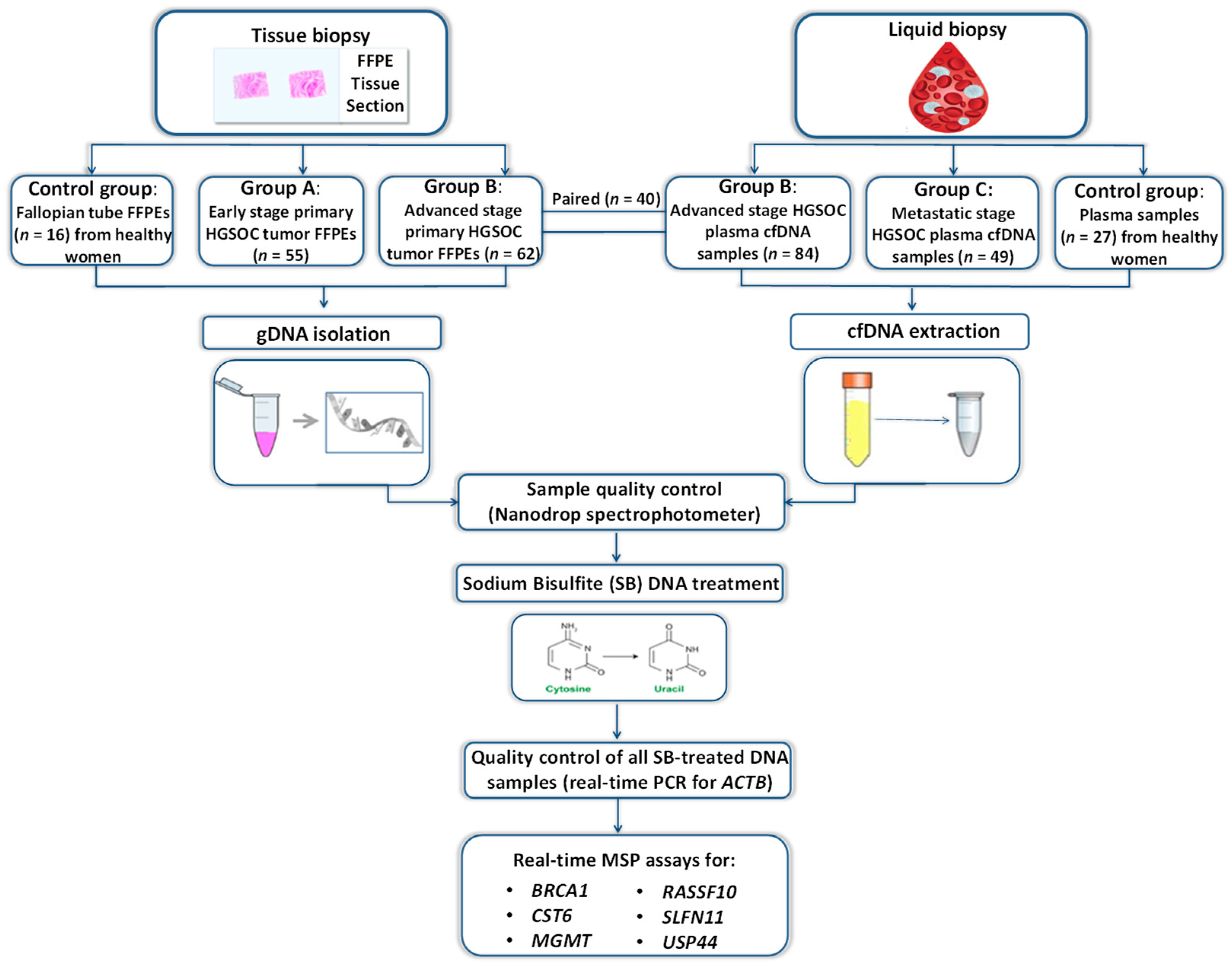
2.1. Clinical Samples
2.2. DNA Isolation from FFPEs and Plasma
2.3. Sodium Bisulfite Conversion
2.4. Real-Time Methylation-Specific PCR (Real-Time MSP)
2.5. Statistical Analysis
3. Results
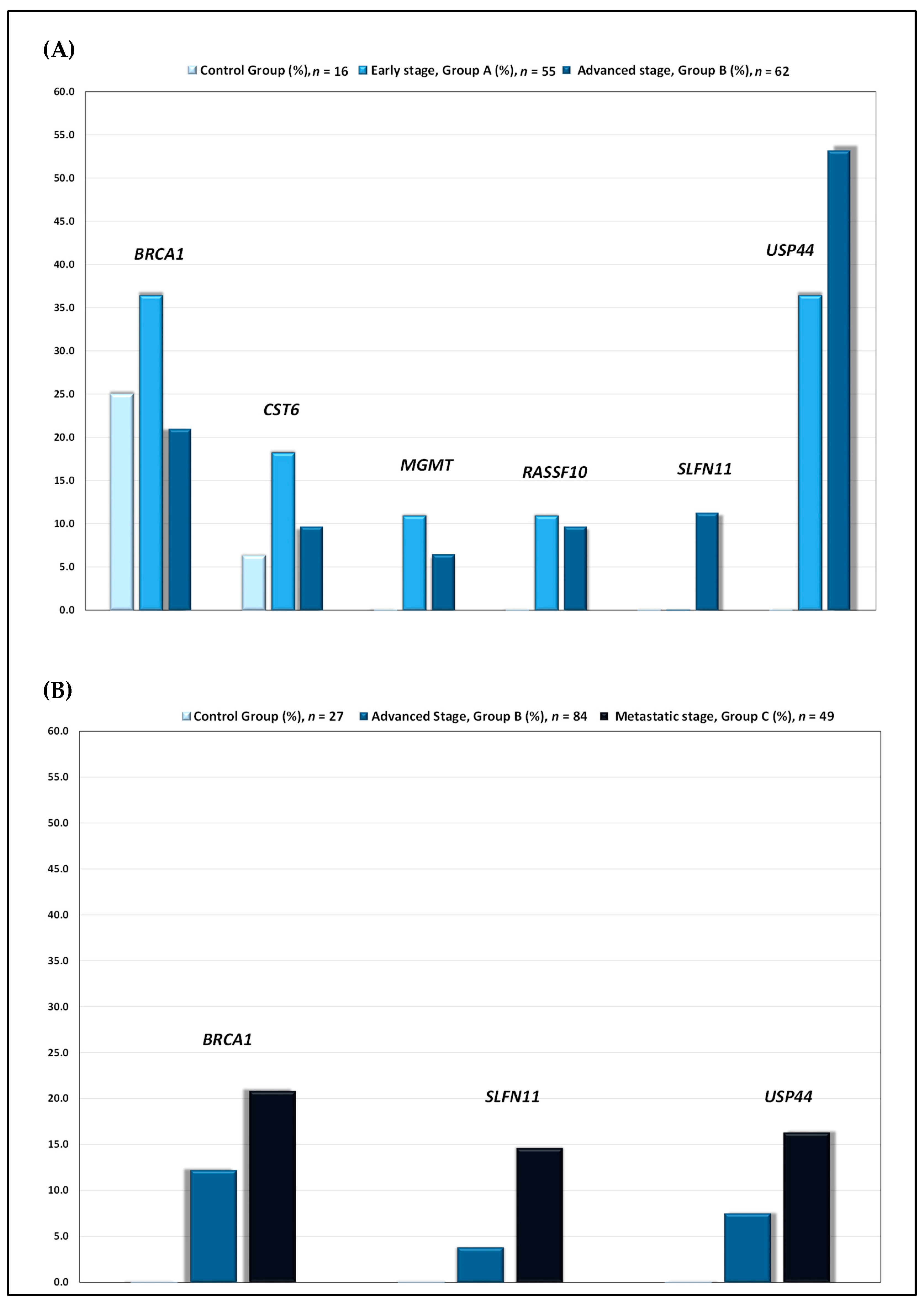
3.1. DNA Methylation Markers in HGSOC Primary Tumors and Plasma cfDNA
3.2. Evaluation of the Prognostic Significance of the DNA Methylation of BRCA1, SLFN11 and USP44 in HGSOC
3.3. Concordance of DNA Methylation between Primary Tumors and Corresponding Plasma cfDNA in Advanced Stage HGSOC
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Wood, G.E.; Ledermann, J.A. Adjuvant and post-surgical treatment in high-grade epithelial ovarian cancer. Best Pract. Res. Clin. Obstet. Gynaecol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Burger, R.A.; Brady, M.F.; Bookman, M.A.; Fleming, G.F.; Monk, B.J.; Huang, H.; Mannel, R.S.; Homesley, H.D.; Fowler, J.; Greer, B.E.; et al. Incorporation of Bevacizumab in the Primary Treatment of Ovarian Cancer. N. Engl. J. Med. 2011, 365, 2473–2483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oza, A.M.; Cook, A.D.; Pfisterer, J.; Embleton, A.; Ledermann, J.A.; Pujade-Lauraine, E.; Kristensen, G.; Carey, M.S.; Beale, P.; Cervantes, A.; et al. Standard chemotherapy with or without bevacizumab for women with newly diagnosed ovarian cancer (ICON7): Overall survival results of a phase 3 randomised trial. Lancet Oncol. 2015, 16, 928–936. [Google Scholar] [CrossRef]
- Odunsi, K. Immunotherapy in ovarian cancer. Ann. Oncol. 2017, 28, viii1. [Google Scholar] [CrossRef]
- Lorusso, D.; Ceni, V.; Daniele, G.; Salutari, V.; Pietragalla, A.; Muratore, M.; Nero, C.; Ciccarone, F.; Scambia, G. Newly diagnosed ovarian cancer: Which first-line treatment? Cancer Treat. Rev. 2020, 91, 102111. [Google Scholar] [CrossRef]
- Colombo, N.; Sessa, C.; du Bois, A.; Ledermann, J.; McCluggage, W.G.; McNeish, I.; Morice, P.; Pignata, S.; Ray-Coquard, I.; Vergote, I.; et al. ESMO–ESGO consensus conference recommendations on ovarian cancer: Pathology and molecular biology, early and advanced stages, borderline tumours and recurrent disease. Ann. Oncol. 2019, 30, 672–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patch, A.M.; Christie, E.L.; Etemadmoghadam, D.; Garsed, D.W.; George, J.; Fereday, S.; Nones, K.; Cowin, P.; Alsop, K.; Bailey, P.J.; et al. Whole-genome characterization of chemoresistant ovarian cancer. Nature 2015, 521, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Balasubramaniam, S.; Beaver, J.A.; Horton, S.; Fernandes, L.L.; Tang, S.; Horne, H.N.; Liu, J.; Liu, C.; Schrieber, S.J.; Yu, J.; et al. FDA Approval Summary: Rucaparib for the Treatment of Patients with Deleterious BRCA Mutation–Associated Advanced Ovarian Cancer. Clin. Cancer Res. 2017, 23, 7165–7170. [Google Scholar] [CrossRef] [Green Version]
- Moore, K.; Colombo, N.; Scambia, G.; Kim, B.-G.; Oaknin, A.; Friedlander, M.; Lisyanskaya, A.; Floquet, A.; Leary, A.; Sonke, G.S.; et al. Maintenance Olaparib in Patients with Newly Diagnosed Advanced Ovarian Cancer. N. Engl. J. Med. 2018, 379, 2495–2505. [Google Scholar] [CrossRef]
- Ison, G.; Howie, L.J.; Amiri-Kordestani, L.; Zhang, L.; Tang, S.; Sridhara, R.; Pierre, V.; Charlab, R.; Ramamoorthy, A.; Song, P.; et al. FDA Approval Summary: Niraparib for the Maintenance Treatment of Patients with Recurrent Ovarian Cancer in Response to Platinum-Based Chemotherapy. Clin. Cancer Res. 2018, 24, 4066–4071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombo, I.; Lheureux, S.; Oza, A.M. Rucaparib: A novel PARP inhibitor for BRCA advanced ovarian cancer. Drug Des. Devel. Ther. 2018, 12, 605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerliu, L.; Myruski, S.; Bhatti, A.; Soni, P.; Petrosius, P.; Pervanas, H.C.; Horton, E.R. Niraparib for the Treatment of Recurrent Epithelial Ovarian, Fallopian Tube, or Primary Peritoneal Cancer. Ann. Pharmacother. 2020, 54, 1010–1015. [Google Scholar] [CrossRef] [PubMed]
- Esteller, M. Molecular origins of cancer: Epigenetics in cancer. N. Engl. J. Med. 2008, 358, 1148–1159. [Google Scholar] [CrossRef]
- Dor, Y.; Cedar, H. Principles of DNA methylation and their implications for biology and medicine. Lancet 2018, 392, 777–786. [Google Scholar] [CrossRef]
- Feinberg, A.P.; Koldobskiy, M.A.; Göndör, A. Epigenetic modulators, modifiers and mediators in cancer aetiology and progression. Nat. Rev. Genet. 2016, 17, 284–299. [Google Scholar] [CrossRef] [PubMed]
- Earp, M.A.; Cunningham, J.M. DNA methylation changes in epithelial ovarian cancer histotypes. Genomics 2015, 106, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, A.; Peixoto, A.; Pinto, P.; Pinheiro, M.; Teixeira, M.R. Potential clinical applications of circulating cell-free DNA in ovarian cancer patients. Expert Rev. Mol. Med. 2018, 20, e6. [Google Scholar] [CrossRef]
- Wittenberger, T.; Sleigh, S.; Reisel, D.; Zikan, M.; Wahl, B.; Alunni-Fabbroni, M.; Jones, A.; Evans, I.; Koch, J.; Paprotka, T.; et al. DNA methylation markers for early detection of women’s cancer: Promise and challenges. Epigenomics 2014, 6, 311–327. [Google Scholar] [CrossRef] [Green Version]
- Hentze, J.L.; Høgdall, C.K.; Høgdall, E.V. Methylation and ovarian cancer: Can DNA methylation be of diagnostic use? Mol. Clin. Oncol. 2019, 10, 323. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Gupta, S.; Sachan, M. Epigenetic Biomarkers in the Management of Ovarian Cancer: Current Prospectives. Front. Cell Dev. Biol. 2019, 7, 182. [Google Scholar] [CrossRef] [Green Version]
- Lianidou, E.; Hoon, D. Circulating Tumor Cells and circulating Tumor DNA. In Tietz Textbook of Clinical Chemistry and Molecular Diagnostics; Nader, R., Horrath, A.R., Wittwer, C., Eds.; Elsevier Ltd.: Amsterdam, The Netherlands, 2017; pp. 1111–1144. [Google Scholar]
- Lianidou, E.; Pantel, K. Liquid biopsies. Genes Chromosom. Cancer 2019, 58, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Alix-Panabières, C.; Pantel, K. Liquid Biopsy: From Discovery to Clinical Application. Cancer Discov. 2021, 11, 858–873. [Google Scholar] [CrossRef]
- Otandault, A.; Anker, P.; Al Amir Dache, Z.; Guillaumon, V.; Meddeb, R.; Pastor, B.; Pisareva, E.; Sanchez, C.; Tanos, R.; Tousch, G.; et al. Recent advances in circulating nucleic acids in oncology. Ann. Oncol. 2019, 30, 374–384. [Google Scholar] [CrossRef]
- Warton, K.; Mahon, K.L.; Samimi, G. Methylated circulating tumor DNA in blood: Power in cancer prognosis and response. Endocr. Relat. Cancer 2016, 23, R157–R171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Widschwendter, M.; Zikan, M.; Wahl, B.; Lempiäinen, H.; Paprotka, T.; Evans, I.; Jones, A.; Ghazali, S.; Reisel, D.; Eichner, J.; et al. The potential of circulating tumor DNA methylation analysis for the early detection and management of ovarian cancer. Genome Med. 2017, 9, 116. [Google Scholar] [CrossRef] [PubMed]
- Corcoran, R.B.; Chabner, B.A. Application of Cell-free DNA Analysis to Cancer Treatment. N. Engl. J. Med. 2018, 379, 1754–1765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lianidou, E. Detection and relevance of epigenetic markers on ctDNA: Recent advances and future outlook. Mol. Oncol. 2021, 15, 1683–1700. [Google Scholar] [CrossRef] [PubMed]
- Chimonidou, M.; Strati, A.; Tzitzira, A.; Sotiropoulou, G.; Malamos, N.; Georgoulias, V.; Lianidou, E.S. DNA methylation of tumor suppressor and metastasis suppressor genes in circulating tumor cells. Clin. Chem. 2011, 57, 1169–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chimonidou, M.; Strati, A.; Malamos, N.; Georgoulias, V.; Lianidou, E.S. SOX17 promoter methylation in circulating tumor cells and matched cell-free DNA isolated from plasma of patients with breast cancer. Clin. Chem. 2013, 59, 270–279. [Google Scholar] [CrossRef] [Green Version]
- Chimonidou, M.; Tzitzira, A.; Strati, A.; Sotiropoulou, G.; Sfikas, C.; Malamos, N.; Georgoulias, V.; Lianidou, E. CST6 promoter methylation in circulating cell-free DNA of breast cancer patients. Clin. Biochem. 2013, 46, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Balgkouranidou, I.; Chimonidou, M.; Milaki, G.; Tsarouxa, E.G.; Kakolyris, S.; Welch, D.R.; Georgoulias, V.; Lianidou, E.S. Breast cancer metastasis suppressor-1 promoter methylation in cell-free DNA provides prognostic information in non-small cell lung cancer. Br. J. Cancer 2014, 110, 2054–2062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastoraki, S.; Strati, A.; Tzanikou, E.; Chimonidou, M.; Politaki, E.; Voutsina, A.; Psyrri, A.; Georgoulias, V.; Lianidou, E. ESR1 methylation: A Liquid biopsy-based epigenetic assay for the follow up of patients with metastatic breast cancer receiving endocrine treatment. Clin. Cancer Res. 2018, 24, 1500–1510. [Google Scholar] [CrossRef] [Green Version]
- Giannopoulou, L.; Chebouti, I.; Pavlakis, K.; Kasimir-Bauer, S.; Lianidou, E.S. RASSF1A promoter methylation in high-grade serous ovarian cancer: A direct comparison study in primary tumors, adjacent morphologically tumor cell-free tissues and paired circulating tumor DNA. Oncotarget 2017, 8, 21429–21443. [Google Scholar] [CrossRef] [Green Version]
- Giannopoulou, L.; Mastoraki, S.; Buderath, P.; Strati, A.; Pavlakis, K.; Kasimir-Bauer, S.; Lianidou, E.S. ESR1 methylation in primary tumors and paired circulating tumor DNA of patients with high-grade serous ovarian cancer. Gynecol. Oncol. 2018, 150, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Al-Moghrabi, N.; Al-Showimi, M.; Al-Yousef, N.; Al-Shahrani, B.; Karakas, B.; Alghofaili, L.; Almubarak, H.; Madkhali, S.; Al Humaidan, H. Methylation of BRCA1 and MGMT genes in white blood cells are transmitted from mothers to daughters. Clin. Epigenetics 2018, 10, 99. [Google Scholar] [CrossRef]
- Esteller, M.; Garcia-Foncillas, J.; Andion, E.; Goodman, S.N.; Hidalgo, O.F.; Vanaclocha, V.; Baylin, S.B.; Herman, J.G. Inactivation of the DNA-Repair Gene MGMT and the Clinical Response of Gliomas to Alkylating Agents. N. Engl. J. Med. 2000, 343, 1350–1354. [Google Scholar] [CrossRef]
- Volodko, N.; Gordon, M.; Salla, M.; Ghazaleh, H.A.; Baksh, S. RASSF tumor suppressor gene family: Biological functions and regulation. FEBS Lett. 2014, 588, 2671–2684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.H.; Forster, M.; Liang, Y.; Yu, M.; Wang, H.C.; Robert, F.; Langlais, D.; Pelletier, J.; Clare, S.; Nijnik, A. USP44 is dispensable for normal hematopoietic stem cell function, lymphocyte development, and B-cell-mediated immune response in a mouse model. Exp. Hematol. 2019, 72, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Murai, J.; Tang, S.W.; Leo, E.; Baechler, S.A.; Redon, C.E.; Zhang, H.; Al Abo, M.; Rajapakse, V.N.; Nakamura, E.; Jenkins, L.M.M.; et al. SLFN11 Blocks Stressed Replication Forks Independently of ATR. Mol. Cell 2018, 69, 371–384.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lalmanach, G.; Kasabova-Arjomand, M.; Lecaille, F.; Saidi, A. Cystatin M/E (Cystatin 6): A Janus-Faced Cysteine Protease Inhibitor with Both Tumor-Suppressing and Tumor-Promoting Functions. Cancers 2021, 13, 1877. [Google Scholar] [CrossRef] [PubMed]
- Kalachand, R.D.; Stordal, B.; Madden, S.; Chandler, B.; Cunningham, J.; Goode, E.L.; Ruscito, I.; Braicu, E.I.; Sehouli, J.; Ignatov, A.; et al. BRCA1 Promoter Methylation and Clinical Outcomes in Ovarian Cancer: An Individual Patient Data Meta-Analysis. JNCI J. Natl. Cancer Inst. 2020, 112, 1190. [Google Scholar] [CrossRef]
- Qiao, B.; Zhang, Z.; Li, Y. Association of MGMT promoter methylation with tumorigenesis features in patients with ovarian cancer: A systematic meta-analysis. Mol. Genet. Genomic Med. 2018, 6, 69–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nogales, V.; Reinhold, W.C.; Varma, S.; Martinez-Cardus, A.; Moutinho, C.; Moran, S.; Heyn, H.; Sebio, A.; Barnadas, A.; Pommier, Y.; et al. Epigenetic inactivation of the putative DNA/RNA helicase SLFN11 in human cancer confers resistance to platinum drugs. Oncotarget 2016, 7, 3084–3097. [Google Scholar] [CrossRef] [Green Version]
- Chimonidou, M.; Strati, A.; Malamos, N.; Kouneli, S.; Georgoulias, V.; Lianidou, E. Direct comparison study of DNA methylation markers in EpCAM-positive circulating tumour cells, corresponding circulating tumour DNA, and paired primary tumours in breast cancer. Oncotarget 2017, 8, 72054–72068. [Google Scholar] [CrossRef] [Green Version]
- Richter, A.M.; Walesch, S.K.; Dammann, R.H. Aberrant Promoter Methylation of the Tumour Suppressor RASSF10 and Its Growth Inhibitory Function in Breast Cancer. Cancers 2016, 8, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Londra, D.; Mastoraki, S.; Bournakis, E.; Zavridou, M.; Thanos, A.; Rampias, T.; Lianidou, E.S. USP44 Promoter Methylation in Plasma Cell-Free DNA in Prostate Cancer. Cancers 2021, 13, 4607. [Google Scholar] [CrossRef]
- Esteller, M.; Silva, J.M.; Dominguez, G.; Bonilla, F.; Matias-Guiu, X.; Lerma, E.; Bussaglia, E.; Prat, J.; Harkes, I.C.; Repasky, E.A.; et al. Promoter hypermethylation and BRCA1 inactivation in sporadic breast and ovarian tumors. J. Natl. Cancer Inst. 2000, 92, 564–569. [Google Scholar] [CrossRef]
- Esteller, M.; Hamilton, S.; Burger, P.; Baylin, S.B.; Herman, J.C. Inactivation of the DNA repair gene O6-methylguanine-DNA methyltransferase by promoter hypermethylation is a common event in primary human neoplasia. Cancer Res. 1999, 59, 793–797. [Google Scholar] [PubMed]
- Prieske, K.; Prieske, S.; Joosse, S.A.; Trillsch, F.; Grimm, D.; Burandt, E.; Mahner, S.; Schmalfeldt, B.; Milde-Langosch, K.; Oliveira-Ferrer, L.; et al. Loss of BRCA1 promotor hypermethylation in recurrent high-grade ovarian cancer. Oncotarget 2017, 8, 83063–83074. [Google Scholar] [CrossRef] [Green Version]
- Elazezy, M.; Prieske, K.; Kluwe, L.; Oliveira-Ferrer, L.; Peine, S.; Müller, V.; Woelber, L.; Schmalfeldt, B.; Pantel, K.; Joosse, S.A. BRCA1 promoter hypermethylation on circulating tumor DNA correlates with improved survival of patients with ovarian cancer. Mol. Oncol. 2021, 15, 3615–3625. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Zhao, L.; Lang, J. The BRCA1 Methylation and PD-L1 Expression in Sporadic Ovarian Cancer. Int. J. Gynecol. Cancer 2018, 28, 1514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.Q.; Yan, Q.; Zhang, J.R.; Li, S.-D.; Yang, Y.X.; Wan, X.P. Epigenetic inactivation of BRCA1 through promoter hypermethylation in ovarian cancer progression. J. Obstet. Gynaecol. Res. 2013, 39, 549–554. [Google Scholar] [CrossRef]
- Roh, H.J.; Suh, D.S.; Choi, K.U.; Yoo, H.J.; Joo, W.D.; Yoon, M.S. Inactivation of O6-methyguanine-DNA methyltransferase by promoter hypermethylation: Association of epithelial ovarian carcinogenesis in specific histological types. J. Obstet. Gynaecol. Res. 2011, 37, 851–860. [Google Scholar] [CrossRef] [PubMed]
- Losi, L.; Fonda, S.; Saponaro, S.; Chelbi, S.T.; Lancellotti, C.; Gozzi, G.; Alberti, L.; Fabbiani, L.; Botticelli, L.; Benhattar, J. Distinct DNA Methylation Profiles in Ovarian Tumors: Opportunities for Novel Biomarkers. Int. J. Mol. Sci. 2018, 19, 1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, H.-Y.; Lai, H.-C.; Lin, Y.-W.; Chou, Y.-C.; Liu, C.-Y.; Yu, M.-H. An epigenetic marker panel for screening and prognostic prediction of ovarian cancer. Int. J. Cancer 2009, 124, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.-W.; Fu, C.-F.; Chang, M.-C.; Lu, T.-P.; Lin, H.-P.; Chiang, Y.-C.; Chen, C.-A.; Cheng, W.-F. CDH1, DLEC1 and SFRP5 methylation panel as a prognostic marker for advanced epithelial ovarian cancer. Epigenomics 2018, 10, 1397–1413. [Google Scholar] [CrossRef]
- Montavon, C.; Gloss, B.S.; Warton, K.; Barton, C.A.; Statham, A.L.; Scurry, J.P.; Tabor, B.; Nguyen, T.V.; Qu, W.; Samimi, G.; et al. Prognostic and diagnostic significance of DNA methylation patterns in high grade serous ovarian cancer. Gynecol. Oncol. 2012, 124, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Shee, K.; Wells, J.D.; Jiang, A.; Miller, T.W. Integrated pan-cancer gene expression and drug sensitivity analysis reveals SLFN11 mRNA as a solid tumor biomarker predictive of sensitivity to DNA-damaging chemotherapy. PLoS ONE 2019, 14, e0224267. [Google Scholar] [CrossRef] [PubMed]
- Winkler, C.; King, M.; Berthe, J.; Ferraioli, D.; Garuti, A.; Grillo, F.; Rodriguez-Canales, J.; Ferrando, L.; Chopin, N.; Ray-Coquard, I.; et al. SLFN11 captures cancer-immunity interactions associated with platinum sensitivity in high-grade serous ovarian cancer. JCI Insight 2021, 6, e146098. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Clinicopathological Characteristics | Group A Early, (FIGO I–II) n = 55, (n%) | Group B Advanced (FIGO III) n = 104, (n%) | Group C Metastatic (FIGO IV) n = 49, (n%) |
|---|---|---|---|
| Histology | |||
| Serous | 55 (100.0) | 104 (100.0) | 49 (100.0) |
| Tumor grade (G) | |||
| G1 | - | - | - |
| G2 | 4 (7.3) | 22 (21.2) | 4 (8.2) |
| G3 | 51 (100.0) | 82 (78.9) | 45 (91.8) |
| FIGO stage | |||
| I | 14 (25.5) | - | |
| II | 31(56.5) | - | |
| III | - | 104 (100.0) | - |
| IV | - | 49 (100.0) | |
| Age | Median age = 57 | Medianage = 66 | Median age = 61 |
| ≥median age | 28 (50.9) | 52 (50.0) | 24 (49.0) |
| <median age | 27 (49.1) | 52 (50.0) | 25 (51.0) |
| Distant metastasis (M) | |||
| M0 | 55 (100.0) | 104 (100.0) | - |
| M1 | - | - | 49 (100.0) |
| Unknown | - | - | - |
| Gene | Primer Sequences 5′-3′ (For SB–Converted Sequences) | Detection | PCR Product, bp |
|---|---|---|---|
| BRCA1 | F: TCGTGGTAACGGAAAAGCGC R: AAATCTCAACGAACTCACGCCG | LC green | 75 |
| CST6 | F: TCGAGTTTCGTTTTATTTTAGGTC R: CATAACCGTCAATACCGTCG | FAM–TAGCGGGTAAAAGTGCGCGGTCTAAGTT–BBQ | 134 |
| MGMT | F: TTTCGACGTTCGTAGGTTTTCGC R: CGTTTCGTTTTCGGAAGAGTGC | LC green | 81 |
| RASSF10 | F: CGTCGTTTTAGTAGATTTCGGTC R: CGTCGAAACAAATAATACGACG | LC green | 143 |
| SLFN11 | F: ATGGAGGGAGCGAGGAGAC R: AACGAATCTACTAAACCCGCG | LC green | 155 |
| USP44 | F: TATTTGTAGTGTCGTCGGGATAC R: GCGTTTCTACCAAACAATTCG | LC green | 143 |
| Patient ID | BRCA1 | SLFN11 | USP44 | |||
|---|---|---|---|---|---|---|
| FFPEs | Plasma-cfDNA | FFPEs | Plasma-cfDNA | FFPEs | Plasma-cfDNA | |
| #1 |  | NA | | | | |
| #2 | |  | | | | |
| #4 | | NA | | | | |
| #6 | | | | | | |
| #8 | | | | | | |
| #9 | | | | | | |
| #10 | | | | | | |
| #15 | | | | | | |
| #17 | | | | | | |
| #18 | | | | | | |
| #19 | | | | | | |
| #20 | | | | | | |
| #22 | | | | | | |
| #23 | | | | | | |
| #24 | | | | | | |
| #26 | | | | | | |
| #27 | | | | | | |
| #29 | | | | | | |
| #32 | | | | | | |
| #33 | | | | | | |
| #34 | | | | | | |
| #38 | | | | | | |
| #39 | | | | | | |
| #40 | | | | | | |
| #41 | | | | | | |
| #43 | | | | NA | | NA |
| #44 | | | | | | |
| #45 | | | | | | |
| #47 | | | | NA | | NA |
| #49 | | | | | | |
| #50 | | | | | | |
| #54 | | | | | | |
| #55 | | | | | | |
| #56 | | | NA | NA | NA | NA |
| #57 | | | | | | |
| #58 | | | | | | |
| #59 | | | NA | | NA | |
| #60 | | | | | | |
| #61 | | | | | | |
| #62 | | | | | | |
| #63 | | | | | | |
| #64 | | | | NA | | NA |
| Primary Tumor | Paired Plasma cfDNA | ||
|---|---|---|---|
| BRCA1 | Unmethylated | Methylated | Total |
| Unmethylated | 31 | 4 | 35 |
| Methylated | 5 | 0 | 5 |
| Total | 36 | 4 | 40 |
| Agreement | 31/40 (77.5%), p = 0.573, k = −0.125 | ||
| SLFN11 | Unmethylated | Methylated | Total |
| Unmethylated | 32 | 1 | 33 |
| Methylated | 4 | 0 | 4 |
| Total | 36 | 1 | 37 |
| Agreement | 32/37 (86.5%), p = 0.892, k = −0.045 | ||
| USP44 | Unmethylated | Methylated | Total |
| Unmethylated | 19 | 0 | 19 |
| Methylated | 16 | 2 | 18 |
| Total | 35 | 2 | 37 |
| Agreement | 21/37 (56.8%), p = 0.230, k = 0.114 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tserpeli, V.; Stergiopoulou, D.; Londra, D.; Giannopoulou, L.; Buderath, P.; Balgkouranidou, I.; Xenidis, N.; Grech, C.; Obermayr, E.; Zeillinger, R.; et al. Prognostic Significance of SLFN11 Methylation in Plasma Cell-Free DNA in Advanced High-Grade Serous Ovarian Cancer. Cancers 2022, 14, 4. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14010004
Tserpeli V, Stergiopoulou D, Londra D, Giannopoulou L, Buderath P, Balgkouranidou I, Xenidis N, Grech C, Obermayr E, Zeillinger R, et al. Prognostic Significance of SLFN11 Methylation in Plasma Cell-Free DNA in Advanced High-Grade Serous Ovarian Cancer. Cancers. 2022; 14(1):4. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14010004
Chicago/Turabian StyleTserpeli, Victoria, Dimitra Stergiopoulou, Dora Londra, Lydia Giannopoulou, Paul Buderath, Ioanna Balgkouranidou, Nikolaos Xenidis, Christina Grech, Eva Obermayr, Robert Zeillinger, and et al. 2022. "Prognostic Significance of SLFN11 Methylation in Plasma Cell-Free DNA in Advanced High-Grade Serous Ovarian Cancer" Cancers 14, no. 1: 4. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14010004