

Profiling the Adrenergic System in Breast Cancer and the Development of Metastasis
 , ,
, ,  , , ,
, , ,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Human Samples
2.2. Quantitative ELISA Analysis
2.3. Immunofluorescence Analysis
2.4. Transcriptomic Analysis
2.4.1. Human BC Tissue Samples Dataset
2.4.2. Human BC Cell Lines Dataset
2.5. Statistical Analysis
3. Results
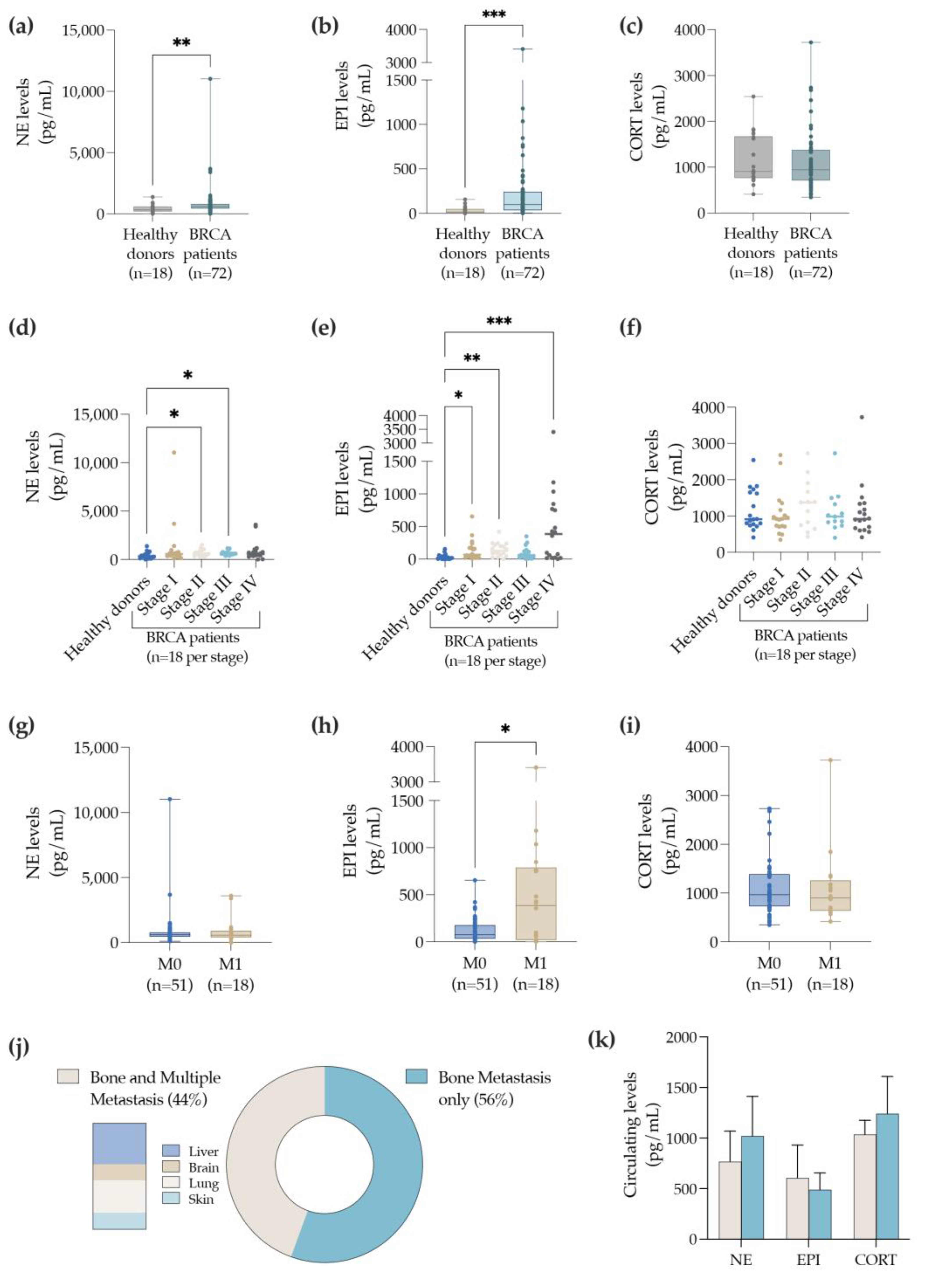
3.1. BC Patients with Bone Metastasis Exhibit Elevated Levels of Circulating EPI
3.2. Bone Metastasis Biopsies Retrieved from BC Patients Express Sympathetic Nerve Fibers and the Adrenergic Receptors—α2a and β2
3.3. ADR Genes Are Downregulated in BC Tissues and Correlate with a Worse Prognosis
3.4. The ADRA2A, ADRA2C and ADRB2 Genes Are Differentially Expressed by the Distinct BC Subtypes
3.5. Markers of Catecholamine Synthesis, Reuptake and Degradation Are also Downregulated in BC Tissues
3.6. Human BC Cell Lines Express Higher Levels of ADRA2C and COMT Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferlay, J.E.M.; Lam, F.; Colombet, M.; Mery, L.; Piñeros, M.; Znaor, A.; Soerjomataram, I.; Bray, F. Global Cancer Observatory: Cancer Today. Lyon, France: International Agency for Research on Cancer. 2020. Available online: https://gco.iarc.fr/today (accessed on 12 January 2021).
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, F.; Kyriakides, S.; Ohno, S.; Penault-Llorca, F.; Poortmans, P.; Rubio, I.T.; Zackrisson, S.; Senkus, E. Early breast cancer: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2019, 30, 1194–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zielonke, N.; Gini, A.; Jansen, E.E.L.; Anttila, A.; Segnan, N.; Ponti, A.; Veerus, P.; de Koning, H.J.; van Ravesteyn, N.T.; Heijnsdijk, E.A.M. Evidence for reducing cancer-specific mortality due to screening for breast cancer in Europe: A systematic review. Eur. J. Cancer Oxf. Engl. 1990 2020, 127, 191–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carioli, G.; Malvezzi, M.; Rodriguez, T.; Bertuccio, P.; Negri, E.; La Vecchia, C. Trends and predictions to 2020 in breast cancer mortality in Europe. Breast 2017, 36, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Society, A.C. Breast Cancer Facts & Figures 2019–2020. American Cancer Society 2019. Available online: https://www.cancer.org (accessed on 23 February 2021).
- Cardoso, F.; Senkus, E.; Costa, A.; Papadopoulos, E.; Aapro, M.; André, F.; Harbeck, N.; Aguilar Lopez, B.; Barrios, C.H.; Bergh, J.; et al. 4th ESO-ESMO International Consensus Guidelines for Advanced Breast Cancer (ABC 4). Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2018, 29, 1634–1657. [Google Scholar] [CrossRef]
- Kohler, B.A.; Sherman, R.L.; Howlader, N.; Jemal, A.; Ryerson, A.B.; Henry, K.A.; Boscoe, F.P.; Cronin, K.A.; Lake, A.; Noone, A.-M.; et al. Annual Report to the Nation on the Status of Cancer, 1975–2011, Featuring Incidence of Breast Cancer Subtypes by Race/Ethnicity, Poverty, and State. JNCI J. Natl. Cancer Inst. 2015, 107, djv048. [Google Scholar] [CrossRef]
- SEER*Explorer: An Interactive Website for SEER Cancer Statistics. Surveillance Research Program, National Cancer Institute. Available online: https://seer.cancer.gov (accessed on 23 February 2021).
- Zahalka, A.H.; Frenette, P.S. Nerves in cancer. Nat. Rev. Cancer 2020, 20, 143–157. [Google Scholar] [CrossRef]
- Conceicao, F.; Sousa, D.M.; Paredes, J.; Lamghari, M. Sympathetic activity in breast cancer and metastasis: Partners in crime. Bone Res. 2021, 9, 9. [Google Scholar] [CrossRef]
- Elefteriou, F. Chronic stress, sympathetic activation and skeletal metastasis of breast cancer cells. BoneKEy Rep. 2015, 4, 693. [Google Scholar] [CrossRef] [Green Version]
- Kamiya, A.; Hayama, Y.; Kato, S.; Shimomura, A.; Shimomura, T.; Irie, K.; Kaneko, R.; Yanagawa, Y.; Kobayashi, K.; Ochiya, T. Genetic manipulation of autonomic nerve fiber innervation and activity and its effect on breast cancer progression. Nat. Neurosci. 2019, 22, 1289–1305. [Google Scholar] [CrossRef]
- Cole, S.W.; Nagaraja, A.S.; Lutgendorf, S.K.; Green, P.A.; Sood, A.K. Sympathetic nervous system regulation of the tumour microenvironment. Nat. Rev. Cancer 2015, 15, 563–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulcrone, P.L.; Campbell, J.P.; Clément-Demange, L.; Anbinder, A.L.; Merkel, A.R.; Brekken, R.A.; Sterling, J.A.; Elefteriou, F. Skeletal Colonization by Breast Cancer Cells Is Stimulated by an Osteoblast and β2AR-Dependent Neo-Angiogenic Switch. J. Bone Miner. Res. 2017, 32, 1442–1454. [Google Scholar] [CrossRef] [PubMed]
- McCorry, L.K. Physiology of the autonomic nervous system. Am. J. Pharm. Educ. 2007, 71, 78. [Google Scholar] [CrossRef] [Green Version]
- McEwen, B.S. Neurobiological and Systemic Effects of Chronic Stress. Chronic Stress 2017, 1, 2470547017692328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thaker, P.H.; Sood, A.K.; Ramondetta, L.M. Importance of adrenergic pathways in women’s cancers. Cancer Biomark 2013, 13, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Insel, P.A. Adrenergic receptors, G proteins, and cell regulation: Implications for aging research. Exp. Gerontol. 1993, 28, 341–348. [Google Scholar] [CrossRef]
- Draoui, A.; Vandewalle, B.; Hornez, L.; Revillion, F.; Lefebvre, J. β-Adrenergic receptors in human breast cancer: Identification, characterization and correlation with progesterone and estradiol receptors. Anticancer. Res. 1991, 11, 677–680. [Google Scholar]
- Vandewalle, B.; Revillion, F.; Lefebvre, J. Functional β-adrenergic receptors in breast cancer cells. J. Cancer Res. Clin. Oncol. 1990, 116, 303–306. [Google Scholar] [CrossRef]
- Re, G.; Badino, P.; Girardi, C.; Di Carlo, F. Effects of a β2-agonist (clenbuterol) on cultured human (CG-5) breast cancer cells. Pharmacol. Res. 1992, 26, 377–384. [Google Scholar] [CrossRef]
- Re, G.; Badino, P.; Novelli, A.; Girardi, C.; Di Carlo, F. Evidence for functional β-adrenoceptor subtypes in CG-5 breast cancer cells. Pharmacol. Res. 1996, 33, 255–260. [Google Scholar] [CrossRef]
- Cakir, Y.; Plummer, H.K.; Tithof, P.K.; Schuller, H.M. Beta-adrenergic and arachidonic acid-mediated growth regulation of human breast cancer cell lines. Int. J. Oncol. 2002, 21, 153–157. [Google Scholar] [CrossRef]
- Montoya, A.; Amaya, C.N.; Belmont, A.; Diab, N.; Trevino, R.; Villanueva, G.; Rains, S.; Sanchez, L.A.; Badri, N.; Otoukesh, S.; et al. Use of non-selective β-blockers is associated with decreased tumor proliferative indices in early stage breast cancer. Oncotarget 2017, 8, 6446–6460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillis, R.D.; Botteri, E.; Chang, A.; Ziegler, A.I.; Chung, N.-C.; Pon, C.K.; Shackleford, D.M.; Andreassen, B.K.; Halls, M.L.; Baker, J.G.; et al. Carvedilol blocks neural regulation of breast cancer progression in vivo and is associated with reduced breast cancer mortality in patients. Eur. J. Cancer Oxf. Engl. 1990 2021, 147, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Pérez Piñero, C.; Bruzzone, A.; Sarappa, M.G.; Castillo, L.F.; Lüthy, I.A. Involvement of α2- and β2-adrenoceptors on breast cancer cell proliferation and tumour growth regulation. Br. J. Pharmacol. 2012, 166, 721–736. [Google Scholar] [CrossRef] [Green Version]
- Slotkin, T.A.; Zhang, J.; Dancel, R.; Garcia, S.J.; Willis, C.; Seidler, F.J. Beta-adrenoceptor signaling and its control of cell replication in MDA-MB-231 human breast cancer cells. Breast Cancer Res. Treat. 2000, 60, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Drell IV, T.L.; Joseph, J.; Lang, K.; Niggemann, B.; Zaenker, K.S.; Entschladen, F. Effects of neurotransmitters on the chemokinesis and chemotaxis of MDA-MB-468 human breast carcinoma cells. Breast Cancer Res. Treat. 2003, 80, 63–70. [Google Scholar] [CrossRef]
- Lang, K.; Drell IV, T.L.; Lindecke, A.; Niggemann, B.; Kaltschmidt, C.; Zaenker, K.S.; Entschladen, F. Induction of a metastatogenic tumor cell type by neurotransmitters and its pharmacological inhibition by established drugs. Int. J. Cancer 2004, 112, 231–238. [Google Scholar] [CrossRef]
- Gruet, M.; Cotton, D.; Coveney, C.; Boocock, D.J.; Wagner, S.; Komorowski, L.; Rees, R.C.; Pockley, A.G.; Garner, A.C.; Wallis, J.D.; et al. β2-Adrenergic Signalling Promotes Cell Migration by Upregulating Expression of the Metastasis-Associated Molecule LYPD3. Biology 2020, 9, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rico, M.; Baglioni, M.; Bondarenko, M.; Cesatti Laluce, N.; Rozados, V.; André, N.; Carré, M.; Scharovsky, O.G.; Márquez, M.M. Metformin and propranolol combination prevents cancer progression and metastasis in different breast cancer models. Oncotarget 2016, 8, 2874–2889. [Google Scholar] [CrossRef] [Green Version]
- Dezong, G.; Zhongbing, M.; Qinye, F.; Zhigang, Y. Carvedilol suppresses migration and invasion of malignant breast cells by inactivating Src involving cAMP/PKA and PKCδ signaling pathway. J. Cancer Res. Ther. 2014, 10, 998–1003. [Google Scholar] [CrossRef]
- Sloan, E.K.; Priceman, S.J.; Cox, B.F.; Yu, S.; Pimentel, M.A.; Tangkanangnukul, V.; Arevalo, J.M.G.; Morizono, K.; Karanikolas, B.D.W.; Wu, L.; et al. The sympathetic nervous system induces a metastatic switch in primary breast cancer. Cancer Res. 2010, 70, 7042–7052. [Google Scholar] [CrossRef] [PubMed]
- Chang, A.; Le, C.P.; Walker, A.K.; Creed, S.J.; Pon, C.K.; Albold, S.; Carroll, D.; Halls, M.L.; Lane, J.R.; Riedel, B.; et al. β2-Adrenoceptors on tumor cells play a critical role in stress-enhanced metastasis in a mouse model of breast cancer. Brain Behav. Immun. 2016, 57, 106–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, A.; Fraser, S.P.; Djamgoz, M.B.A. Propranolol inhibits neonatal Nav1.5 activity and invasiveness of MDA-MB-231 breast cancer cells: Effects of combination with ranolazine. J. Cell Physiol. 2019, 234, 23066–23081. [Google Scholar] [CrossRef] [PubMed]
- Madden, K.S.; Szpunar, M.J.; Brown, E.B. β-Adrenergic receptors (β-AR) regulate VEGF and IL-6 production by divergent pathways in high β-AR-expressing breast cancer cell lines. Breast Cancer Res. Treat. 2011, 130, 747–758. [Google Scholar] [CrossRef] [Green Version]
- Nuevo-Tapioles, C.; Santacatterina, F.; Stamatakis, K.; Núñez de Arenas, C.; de Cedrón, M.G.; Formentini, L.; Cuezva, J.M. Coordinate β-adrenergic inhibition of mitochondrial activity and angiogenesis arrest tumor growth. Nat. Commun. 2020, 11, 3606. [Google Scholar] [CrossRef]
- Huang, J.-F.; Shen, J.; Li, X.; Rengan, R.; Silvestris, N.; Wang, M.; Derosa, L.; Zheng, X.; Belli, A.; Zhang, X.-L.; et al. Incidence of patients with bone metastases at diagnosis of solid tumors in adults: A large population-based study. Ann. Transl. Med. 2020, 8, 482. [Google Scholar] [CrossRef]
- Vasconcelos, D.M.; Ribeiro-da-Silva, M.; Mateus, A.; Alves, C.J.; Machado, G.C.; Machado-Santos, J.; Paramos-de-Carvalho, D.; Alencastre, I.S.; Henrique, R.; Costa, G.; et al. Immune response and innervation signatures in aseptic hip implant loosening. J. Transl. Med. 2016, 14, 205. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro-da-Silva, M.; Vasconcelos, D.M.; Alencastre, I.S.; Oliveira, M.J.; Linhares, D.; Neves, N.; Costa, G.; Henrique, R.; Lamghari, M.; Alves, C.J. Interplay between sympathetic nervous system and inflammation in aseptic loosening of hip joint replacement. Sci. Rep. 2018, 8, 16044. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Cheng, H.; Bai, Z.; Li, J. Breast cancer cell line classification and Its relevance with breast tumor subtyping. J. Cancer 2017, 8, 3131–3141. [Google Scholar] [CrossRef] [Green Version]
- Soni, A.; Ren, Z.; Hameed, O.; Chanda, D.; Morgan, C.J.; Siegal, G.P.; Wei, S. Breast cancer subtypes predispose the site of distant metastases. Am. J. Clin. Pathol. 2015, 143, 471–478. [Google Scholar] [CrossRef]
- Sui, L.; Sanders, A.; Jiang, W.G.; Ye, L. Deregulated molecules and pathways in the predisposition and dissemination of breast cancer cells to bone. Comput. Struct. Biotechnol. J. 2022, 20, 2745–2758. [Google Scholar] [CrossRef] [PubMed]
- Lourenco, C.; Conceicao, F.; Jeronimo, C.; Lamghari, M.; Sousa, D.M. Stress in Metastatic Breast Cancer: To the Bone and Beyond. Cancers 2022, 14, 1881. [Google Scholar] [CrossRef] [PubMed]
- Tank, A.W.; Lee Wong, D. Peripheral and central effects of circulating catecholamines. Compr. Physiol. 2015, 5, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Honke, N.; Lowin, T.; Opgenoorth, B.; Shaabani, N.; Lautwein, A.; Teijaro, J.R.; Schneider, M.; Pongratz, G. Endogenously produced catecholamines improve the regulatory function of TLR9-activated B cells. PLoS Biol. 2022, 20, e3001513. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, S.; Mortensen, O.V. Overview of Monoamine Transporters. Curr. Protoc. Pharm. 2017, 79, 12.16.11–12.16.17. [Google Scholar] [CrossRef]
- Lueboonthavatchai, P. Prevalence and psychosocial factors of anxiety and depression in breast cancer patients. J. Med. Assoc. Thail. 2007, 90, 2164–2174. [Google Scholar]
- Compas, B.E.; Stoll, M.F.; Thomsen, A.H.; Oppedisano, G.; Epping-Jordan, J.A.E.; Krag, D.N. Adjustment to breast cancer: Age-related differences in coping and emotional distress. Breast Cancer Res. Treat. 1999, 54, 195–203. [Google Scholar] [CrossRef]
- Toleutay, U.; Reznik, V.; Kalmatayeva, Z.; Smigelskas, K. Risk factors of breast cancer in kyzylorda oblast of Kazakhstan: A case-control study. Asian Pac. J. Cancer Prev. APJCP 2013, 14, 5961–5964. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Liao, W.-C.; Tsai, C.-J.; Wang, L.-R.; Mao, I.-F.; Chen, C.-C.; Kao, P.-F.; Yao, C.-C. The effects of perceived stress and life style leading to breast cancer. Women Health 2013, 53, 20–40. [Google Scholar] [CrossRef]
- Eckerling, A.; Ricon-Becker, I.; Sorski, L.; Sandbank, E.; Ben-Eliyahu, S. Stress and cancer: Mechanisms, significance and future directions. Nat. Rev. Cancer 2021, 21, 767–785. [Google Scholar] [CrossRef]
- Barron, T.I.; Connolly, R.M.; Sharp, L.; Bennett, K.; Visvanathan, K. Beta blockers and breast cancer mortality: A population-based study. J. Clin. Oncol. 2011, 29, 2635–2644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melhem-Bertrandt, A.; Chavez-MacGregor, M.; Lei, X.; Brown, E.N.; Lee, R.T.; Meric-Bernstam, F.; Sood, A.K.; Conzen, S.D.; Hortobagyi, G.N.; Gonzalez-Angulo, A.M. Beta-blocker use is associated with improved relapse-free survival in patients with triple-negative breast cancer. J. Clin. Oncol. 2011, 29, 2645–2652. [Google Scholar] [CrossRef] [PubMed]
- Powe, D.G.; Voss, M.J.; Zänker, K.S.; Habashy, H.O.; Green, A.R.; Ellis, I.O.; Entschladen, F. Beta-blocker drug therapy reduces secondary cancer formation in breast cancer and improves cancer specific survival. Oncotarget 2010, 1, 628–638. [Google Scholar] [CrossRef] [Green Version]
- Hiller, J.G.; Cole, S.W.; Crone, E.M.; Byrne, D.J.; Shackleford, D.M.; Pang, J.B.; Henderson, M.A.; Nightingale, S.S.; Ho, K.M.; Myles, P.S.; et al. Preoperative beta-Blockade with Propranolol Reduces Biomarkers of Metastasis in Breast Cancer: A Phase II Randomized Trial. Clin. Cancer Res. 2020, 26, 1803–1811. [Google Scholar] [CrossRef] [PubMed]
- Ganz, P.A.; Habel, L.A.; Weltzien, E.K.; Caan, B.J.; Cole, S.W. Examining the influence of beta blockers and ACE inhibitors on the risk for breast cancer recurrence: Results from the LACE cohort. Breast Cancer Res. Treat. 2011, 129, 549–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, S.M.; Carey, I.M.; Owen, C.G.; Harris, T.; Dewilde, S.; Cook, D.G. Does β-adrenoceptor blocker therapy improve cancer survival? Findings from a population-based retrospective cohort study. Br. J. Clin. Pharmacol. 2011, 72, 157–161. [Google Scholar] [CrossRef] [Green Version]
- Cardwell, C.R.; Pottegård, A.; Vaes, E.; Garmo, H.; Murray, L.J.; Brown, C.; Vissers, P.A.J.; O’Rorke, M.; Visvanathan, K.; Cronin-Fenton, D.; et al. Propranolol and survival from breast cancer: A pooled analysis of European breast cancer cohorts. Breast Cancer Res. 2016, 18, 119. [Google Scholar] [CrossRef] [Green Version]
- Lofling, L.L.; Stoer, N.C.; Sloan, E.K.; Chang, A.; Gandini, S.; Ursin, G.; Botteri, E. beta-blockers and breast cancer survival by molecular subtypes: A population-based cohort study and meta-analysis. Br. J. Cancer 2022, 127, 1086–1096. [Google Scholar] [CrossRef]
- Spini, A.; Roberto, G.; Gini, R.; Bartolini, C.; Bazzani, L.; Donnini, S.; Crispino, S.; Ziche, M. Evidence of beta-blockers drug repurposing for the treatment of triple negative breast cancer: A systematic review. Neoplasma 2019, 66, 963–970. [Google Scholar] [CrossRef]
- Rivero, E.M.; Martinez, L.M.; Bruque, C.D.; Gargiulo, L.; Bruzzone, A.; Lüthy, I.A. Prognostic significance of α- and β2-adrenoceptor gene expression in breast cancer patients. Br. J. Clin. Pharmacol. 2019, 85, 2143–2154. [Google Scholar] [CrossRef]
- Grosman-Rimon, L.; Wright, E.; Sabovich, S.; Rimon, J.; Gleitman, S.; Sudarsky, D.; Lubovich, A.; Gabizon, I.; Lalonde, S.D.; Tsuk, S.; et al. Relationships among norepinephrine levels, exercise capacity, and chronotropic responses in heart failure patients. Heart Fail. Rev. 2022, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lutgendorf, S.K.; DeGeest, K.; Dahmoush, L.; Farley, D.; Penedo, F.; Bender, D.; Goodheart, M.; Buekers, T.E.; Mendez, L.; Krueger, G.; et al. Social isolation is associated with elevated tumor norepinephrine in ovarian carcinoma patients. Brain Behav. Immun. 2011, 25, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Lutgendorf, S.K.; DeGeest, K.; Sung, C.Y.; Arevalo, J.M.; Penedo, F.; Lucci, J.; Goodheart, M.; Lubaroff, D.; Farley, D.M.; Sood, A.K.; et al. Depression, social support, and beta-adrenergic transcription control in human ovarian cancer. Brain Behav. Immun. 2009, 23, 176–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szpunar, M.J.; Belcher, E.K.; Dawes, R.P.; Madden, K.S. Sympathetic innervation, norepinephrine content, and norepinephrine turnover in orthotopic and spontaneous models of breast cancer. Brain Behav. Immun. 2016, 53, 223–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vázquez, S.M.; Mladovan, A.G.; Pérez, C.; Bruzzone, A.; Baldi, A.; Lüthy, I.A. Human breast cell lines exhibit functional α2-adrenoceptors. Cancer Chemother. Pharmacol. 2006, 58, 50–61. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Healthy Donors | BC Patients | |
|---|---|---|
| Patients (n) | 18 | 72 |
| Median age at diagnosis (min–max) | 57 (46–65) | 59 (30–93) |
| Median tumor size (mm) (min–max) | − | 26 (0.3–67) |
| Molecular subtype * (%) | ||
| Luminal A | − | 39 (54.2) |
| Luminal B | − | 29 (40.3) |
| HER2-enriched | − | 2 (2.8) |
| Basal-like/TNBC | − | 2 (2.8) |
| Histological type (%) | ||
| Invasive ductal carcinoma | − | 53 (73.6) |
| Invasive lobular carcinoma | − | 14 (19.4) |
| Mixed-type carcinoma | − | 2 (2.8) |
| Special subtype | − | 3 (4.2) |
| Grade (%) | ||
| G1 | − | 1 (1.4) |
| G2 | − | 42 (58.3) |
| G3 | − | 28 (38.9) |
| Gx | − | 1 (1.4) |
| T stage (%) | ||
| T1 | − | 31 (43.1) |
| T2 | − | 19 (26.4) |
| T3 | − | 14 (19.4) |
| T4 | − | 8 (11.1) |
| N stage (%) | ||
| N0 | − | 32 (44.4) |
| N1 | − | 15 (20.8) |
| N2 | − | 8 (11.1) |
| N3 | − | 8 (11.1) |
| Nx | − | 9 (12.5) |
| M stage (%) | ||
| M0 | − | 51 (70.8) |
| M1 | − | 18 (25.0) |
| Mx | − | 3 (4.2) |
| Menopause Status (%) | ||
| Pre | − | 16.7 |
| Post | − | 83.3 |
| Total | TH+ | α2a+ | β2+ | TH/α2a/β2+ | |
|---|---|---|---|---|---|
| Patients (n) | 44 | 21 | 18 | 43 | 13 |
| Mean age at diagnosis (min-max) | 52 (28–76) | 52 (28–76) | 51 (32–73) | 51 (28–76) | 51 (32–73) |
| Tumor size (mm) (min-max) | 30 (9–100) | 33 (10–100) | 32 (9–100) | 30 (9–100) | 37 (15–100) |
| Molecular subtype * (%) | |||||
| Luminal A | 32 | 24 | 39 | 33 | 38 |
| Luminal B | 66 | 76 | 56 | 65 | 62 |
| HER2-enriched | 0 | 0 | 0 | 0 | 0 |
| Basal-like/TBNC | 2 | 0 | 5 | 2 | 0 |
| Grade (%) | |||||
| G1 | 6.8 | 4.8 | 5.6 | 7.0 | 0 |
| G2 | 52.3 | 52.4 | 50.0 | 53.5 | 53.8 |
| G3 | 40.9 | 42.8 | 44.4 | 39.5 | 46.2 |
| ADRA1A | ADRA1B | ADRA1D | ADRA2A | ADRA2B | ADRA2C | ADRB1 | ADRB2 | ADRB3 | |
|---|---|---|---|---|---|---|---|---|---|
| TH | |||||||||
| Pearson’s correlation | −0.012 | 0.072 * | 0.033 | −0.091 ** | 0.002 | 0.127 *** | 0.015 | −0.028 | 0.002 |
| p-value | 0.736 | 0.047 | 0.360 | 0.010 | 0.961 | 0.000 | 0.692 | 0.438 | 0.966 |
| DBH | |||||||||
| Pearson’s correlation | 0.126 *** | −0.008 | −0.008 | 0.087 * | 0.190 *** | 0.068 | 0.136 *** | 0.249 *** | 0.155 *** |
| p-value | 0.000 | 0.816 | 0.816 | 0.012 | 0.000 | 0.056 | 0.000 | 0.000 | 0.000 |
| PNMT | |||||||||
| Pearson’s correlation | 0.007 | 0.020 | 0.020 | 0.168 *** | 0.040 | 0.166 *** | 0.042 | 0.136 *** | 0.056 |
| p-value | 0.845 | 0.595 | 0.595 | 0.000 | 0.280 | 0.000 | 0.272 | 0.000 | 0.129 |
| COMT | |||||||||
| Pearson’s correlation | 3.4925 *** | 2.070 * | 1.321 | −2.463 * | 4.163 | 5.470 *** | −0.979 | −1.161 | 0.897 |
| p-value | 0.000 | 0.039 | 0.187 | 0.014 | 0.000 | 0.000 | 0.328 | 0.246 | 0.370 |
| MAO-A | |||||||||
| Pearson’s correlation | 7.426 *** | 0.595 | 2.605 ** | 7.647 *** | 0.978 | 2.227 * | 6.060 *** | 10.056 *** | 5.329 *** |
| p-value | 0.000 | 0.5519 | 0.009 | 0.000 | 0.328 | 0.026 | 0.000 | 0.000 | 0.000 |
| MAO-B | |||||||||
| Pearson’s correlation | 0.415 | 1.901 | 4.271 *** | 7.300 *** | 1.809 | 1.596 | 2.323 * | 7.185 *** | 0.705 |
| p-value | 0.677 | 0.058 | 0.000 | 0.000 | 0.071 | 0.111 | 0.020 | 0.000 | 0.481 |
| NET | |||||||||
| Pearson’s correlation | 4.595 *** | 1.085 | 5.750 *** | 1.348 | 7.091 *** | 1.138 | 5.685 *** | 8.675 *** | 5.856 *** |
| p-value | 0.000 | 0.278 | 0.000 | 0.178 | 0.000 | 0.255 | 0.000 | 0.000 | 0.000 |
| VMAT2 | |||||||||
| Pearson’s correlation | 5.338 *** | −0.196 | 3.541 *** | 10.442 *** | −2.013 * | 1.236 | 2.070 * | 10.345 *** | 1.058 |
| p-value | 0.000 | 0.884 | 0.000 | 0.000 | 0.044 | 0.217 | 0.039 | 0.000 | 0.290 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sousa, D.M.; Fernandes, V.; Lourenço, C.; Carvalho-Maia, C.; Estevão-Pereira, H.; Lobo, J.; Cantante, M.; Couto, M.; Conceição, F.; Jerónimo, C.; et al. Profiling the Adrenergic System in Breast Cancer and the Development of Metastasis. Cancers 2022, 14, 5518. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14225518
Sousa DM, Fernandes V, Lourenço C, Carvalho-Maia C, Estevão-Pereira H, Lobo J, Cantante M, Couto M, Conceição F, Jerónimo C, et al. Profiling the Adrenergic System in Breast Cancer and the Development of Metastasis. Cancers. 2022; 14(22):5518. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14225518
Chicago/Turabian StyleSousa, Daniela M., Veronica Fernandes, Catarina Lourenço, Carina Carvalho-Maia, Helena Estevão-Pereira, João Lobo, Mariana Cantante, Marina Couto, Francisco Conceição, Carmen Jerónimo, and et al. 2022. "Profiling the Adrenergic System in Breast Cancer and the Development of Metastasis" Cancers 14, no. 22: 5518. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14225518