

One-Pot Synthesis of β-Alanine from Maleic Acid via Three-Enzyme Cascade Biotransformation

School of Chemical and Environmental Engineering, Shanghai Institute of Technology, Shanghai 201418, China
*
Authors to whom correspondence should be addressed.
Catalysts 2023, 13(2), 267; https://0-doi-org.brum.beds.ac.uk/10.3390/catal13020267
Submission received: 13 December 2022
/
Revised: 18 January 2023
/
Accepted: 22 January 2023
/
Published: 24 January 2023
(This article belongs to the Special Issue Current State-of-the-Art of Biocatalysts)
Abstract
:A novel and efficient one-pot three-enzyme cascade method for the synthesis of β-alanine from maleic acid was developed. Two recombinant E. coli strains were constructed. The E. coli (MaiA-AspA) co-expressing maleic cis-trans isomerase (MaiA) and L-aspartase (AspA) catalyzed the biotransformation of maleic acid to L-aspartate via fumaric acid, and E. coli (ADC) expressing L-aspartate-α-decarboxylase (ADC) catalyzed the bioconversion of L-aspartate to β-alanine. After systematic optimization of reaction conditions for each strain, the whole cells of two strains were combined for one-pot synthesis of β-alanine. It was found that the ratio of the two kinds of cells as well as the cell amount play critical roles in the reaction rate and yield of β-alanine. Adding two kinds of cells in one-pot at the beginning of the reaction was better than adding step by step. Under optimal conditions, the concentration of β-alanine reached 751 mM after a 9 h reaction, corresponding to a 93.9% yield and 178 g/L/d space-time yield. The developed new route showed application potential for green and efficient biosynthesis of β-alanine from a cheap substrate by tandem biocatalysts.
1. Introduction
β-alanine is a kind of non-protein amino acid, which is widely used in chemical, pharmaceutical, material, feed, and food fields and has a large market demand [1,2]. It can be synthesized by a chemical method from non-renewable raw materials such as acrylonitrile [3] and β-aminopropionitrile [4]. The chemical methods have the defects of harsh reaction conditions, high energy consumption, and pollution with high raw material cost. Although there are natural pathways for the synthesis of β-alanine, their synthetic efficiency is often low, and the reaction process is complex and difficult to regulate. At present, it has been reported that β-alanine can be obtained by the catalysis of acrylic acid aminase [5], β-aminopropanitrile hydrolase [6], and L-aspartic acid-α-decarboxylase (ADC) [7,8] from different substrates. Enzymatic synthesis of β-alanine has the advantages of mild reaction conditions, high selectivity, fewer pollutants, etc. Among them, decarboxylation of L-aspartic acid to β-amino propionic acid using L-aspartic acid-α-decarboxylase is one of the most popular methods in current research [9,10]. However, the yield of β-alanine synthesized by the enzymatic method is not high, and there exist substrate inhibitory inactivation for the ADC from prokaryotic microbes [11]. On the other hand, the production cost of β-alanine with L-aspartic acid as a substrate is high. It is of great significance to develop a new and more economical process for the production of β-alanine.
The preparation process of fine chemicals and drugs usually involves multi-step reactions, and the use of the step reaction method is usually tedious and time-consuming. Multi-enzyme catalytic reactions show good application potential [12,13,14,15,16,17]. The establishment of a multi-enzyme reaction can transform the initial substrate into the target product through several steps of enzymatic reaction in the same reaction system [18], with high catalytic efficiency and strong specificity [19], saving the time and cost of the separation and purification of the reaction intermediates [20], and improving the problem of substrate inhibition to the reaction [21]. It can also adjust the thermodynamic equilibrium of some reversible reactions [22] to make them move to the direction of the target chemical reaction. Therefore, as more and more enzymes are discovered, multi-enzyme catalyzed tandem reactions have become one of the research hotspots in the field of biocatalysis [23,24].
In a relevant study, Long et al. [25] built recombinant E. coli/pRSFDuet -maiA-aspA co-expressing maleate cis-trans isomerase (MaiA) and L-aspartic acid amino lyase (AspA). The substrate maleic acid can be completely converted to L-aspartic acid via fumaric acid with more than 98% conversion. Gao et al. [26] used AspA and ADC in series to transform 0.1 M of fumaric acid into β-alanine with a 90.9% yield when the ratio of AspA to ADC was 1:80. Wang et al. [27] used mutated ADC cells to catalyze the conversion of L-aspartic acid to β-alanine. When 400 mM of substrate was added in five batches under the condition of 80 g/L cells, 94.5% conversion was obtained after 23 h of reaction. However, there was no report on the synthesis of β-alanine from maleic acid by multiple enzymes in one-pot up to now, to the best of our knowledge.
In this paper, we constructed a reaction pathway for the synthesis of β-alanine by using cheap maleic acid instead of L-aspartic acid as the substrate, and selected three enzymes from different sources to convert maleic acid into β-alanine. Firstly, maleic acid was converted to fumaric acid by MaiA from Serratia marcescens. Then, fumaric acid was converted to L-aspartic acid by AspA from E. coli. MaiA and AspA were co-expressed in E. coli since they have similar expression activity [25]. ADC from Tribolium castaneum was selected for the conversion of L-aspartic acid to β-alanine. ADC from eukaryotic sources is a class of pyridoxal phosphate dependent decarboxylase and no substrate inhibitory inactivation of enzyme was reported [28]. The mutant ADCtb (K221R) [27], with higher thermal stability compared to the wild-type, was selected, which will be more beneficial to practical application. Since the activity of ADCtb is much lower than that of MaiA or AspA, it was expressed separately for better flexibility of three-enzyme combination.
2. Results
2.1. Design the Cascade Biotransformation by Whole Cell Biocatalysts
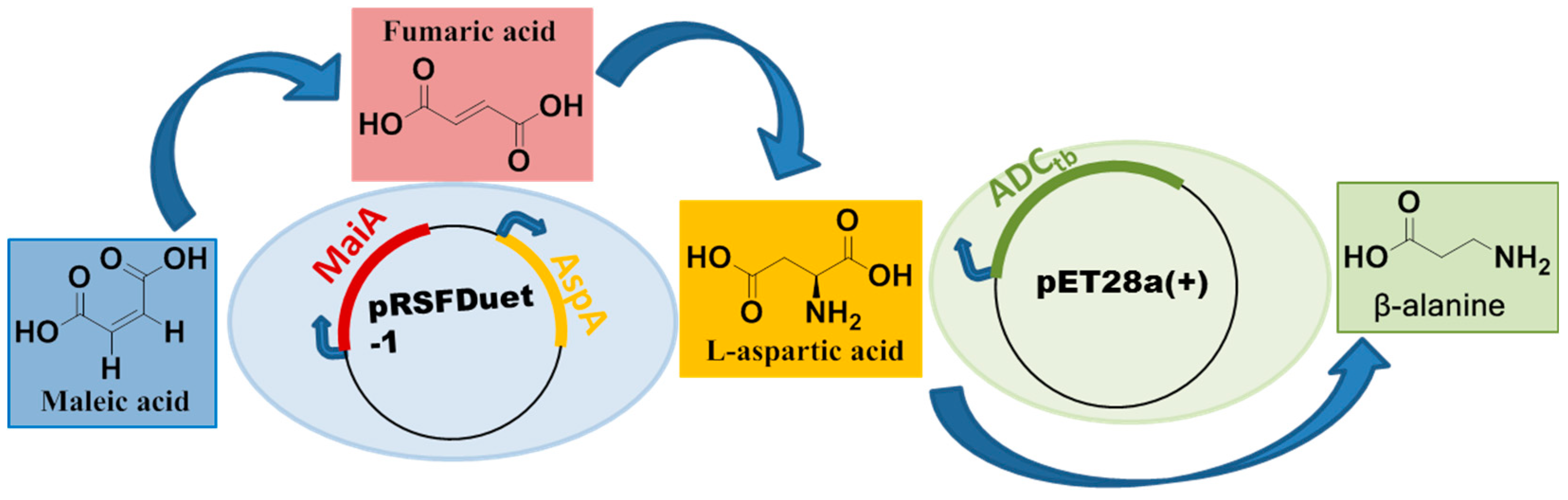
To synthesize β-alanine efficiently and economically, maleic acid was selected as the substrate to further reduce the cost of the substrate, since the price of maleic acid is about four fifths of fumaric acid and one tenth of L-aspartic acid. Maleic acid was first converted to L-aspartic acid via fumaric acid by whole cells of E. coli co-expressing MaiA and AspA, and then L-aspartic acid was transferred to β-alanine by the whole cell of another recombinant E. coli expressing ADCtb (Figure 1). The route uses resting whole cells for a one-pot biotransformation, which are available by cultivation at a low cost and provide a natural environment and cofactor regeneration for enzymes. One-pot biotransformation has multiple enzymes in the reactor to increase the local concentration of enzymes, which reduces the diffusion of intermediates in multistep reactions. In the preliminary experiments, it was found that the cellular enzyme activity of MaiA (48.01 U/mg) and AspA (103 U/mg) were high, while that of ADCtb (7.6 U/mg) was much lower than that of the first two enzymes. To achieve efficient conversion by the three-enzyme cascade reaction, the reaction speed for each step must be balanced. Therefore, MaiA and AspA with higher enzyme activity were co-expressed on a pRSFDuet-1 expression vector, which simplified the bacterial culture steps. ADCtb was independently expressed on pET28a(+) vector, so that the proportion of multiple enzymes could be flexibly adjusted in the reaction. At the same time, the permeable process of ADCtb was carried out to improve the enzyme activity of whole cells, and the decarboxylation condition by L-aspartic acid-α-decarboxylase was optimized. Finally, the three-enzyme catalyzed cascade reaction was studied to achieve the goal of efficient biosynthesis of β-alanine from cheap maleic acid.
2.2. Construction of Engineered Strains
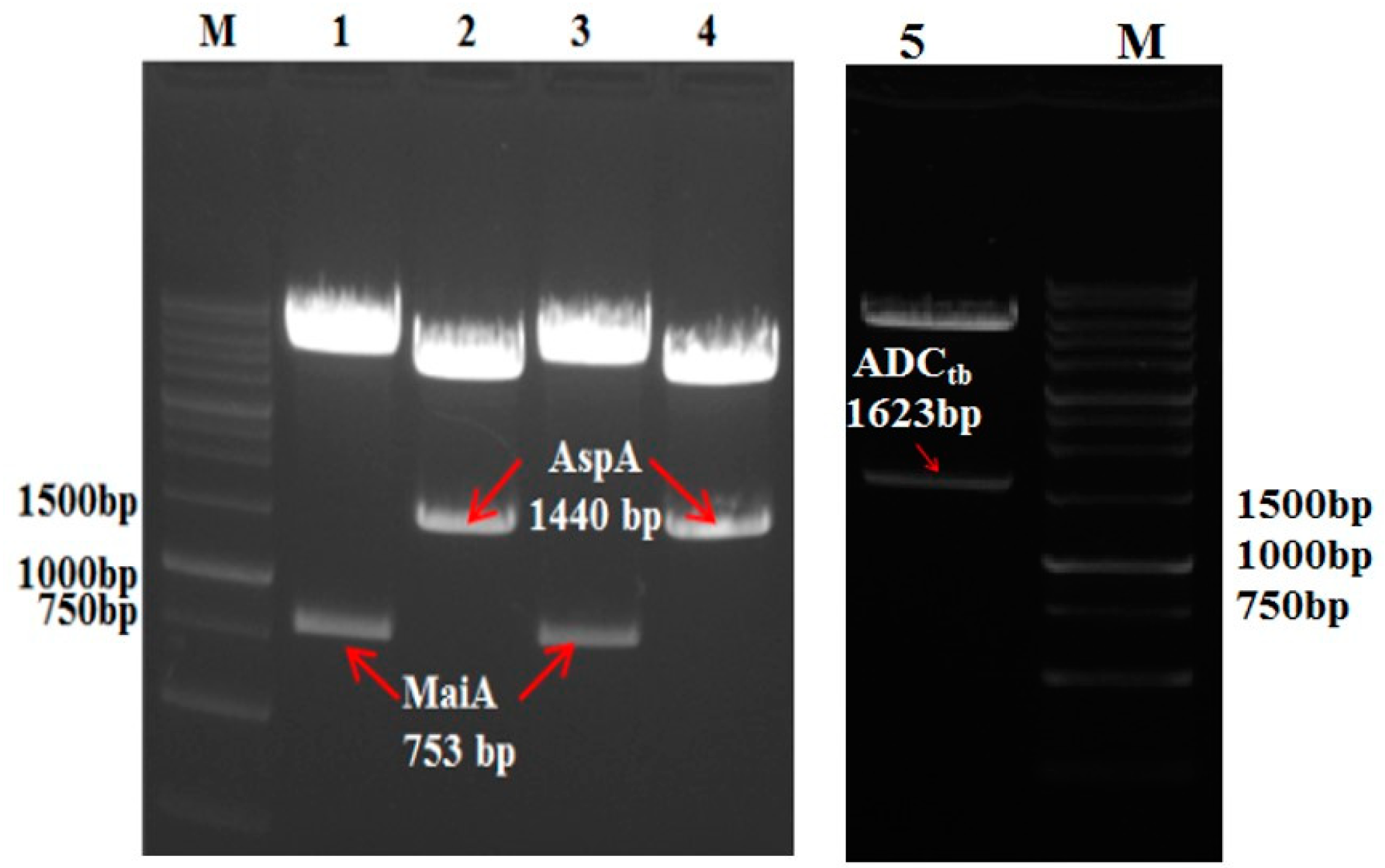
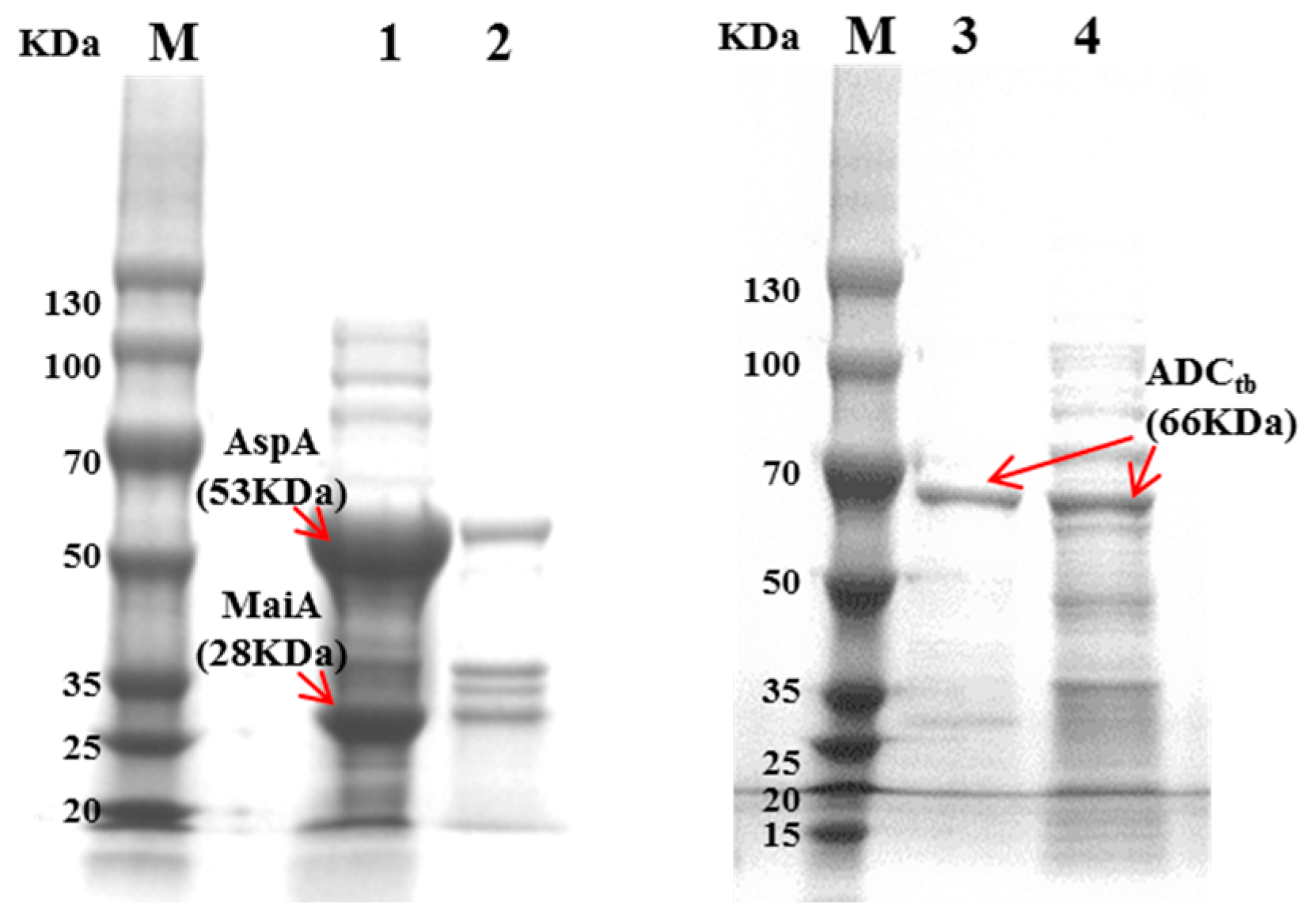
Strains of E. coli BL21 (DE3)-MaiA-AspA and E. coli BL21 (DE3)-ADCtb were constructed. The plasmid pRSFDuet-1-MaiA-AspA was digested with BamHI-HindIII and Ned I-Xho I double enzymes, respectively, and the plasmid pET28a-ADCtb was digested with Nco I/EcoR I, and then treated with 0.8% agarose gel electrophoresis verified. The size of the band was similar to the target band and the theoretical values were consistent. The electrophoresis results are shown in Figure 2 (AspA: 1440 bp, MaiA: 753 bp, ADCtb: 1623 bp). SDS-PAGE was conducted in order to verify the expression of recombinant strain E. coli BL21 (DE3)-MaiA-AspA and E. coli BL21 (DE3)-ADCtb. The size of the band was similar to the target band and the theoretical values were consistent. The protein electrophoresis results are shown in Figure 3 (AspA: 53 KDa, MaiA: 28 KDa, ADCtb: 66 KDa).
2.3. Optimizing the Bioconversion of Maleic Acid to L-Aspartic Acid by Whole Cells of E. coli (MaiA–AspA)
The reaction conditions for the conversion of maleic acid by whole cells of recombinant E. coli co-expressing MaiA and AspA were explored. The effect of substrate concentration, cell amount, and pH value on the reaction were studied, respectively. The appropriate conditions for a double-enzyme reaction were explored (Table 1). Under the condition of 3 g/L cell amount, the conversion at 300 mM of substrate concentration can reach 93% after a 6 h reaction (Table 1, Entry 1). With an increase in the substrate concentration, the conversion decreased for the same reaction time. When the cell amount was lower than 6 g/L, the conversion at 400 mM could not reach more than 95%, while when the amount of cells was higher than 6 g/L, the conversion rate of ammonium maleate at 400 mM could reach 97.5-99.4% at 6 h (Table 1, Entry 8 and 9). To explore the influence of pH on the dual-enzyme catalyzed reaction, the reaction of maleic acid to L-aspartic acid at different pH value was explored (pH 5.5~9.5). After a 6 h reaction, the conversion was only 75.2% at pH 5.5 (Table 1, Entry 10), followed by 82.7% at pH 9.5 (Table 1, Entry 13). When the pH value is between 6.5 and 8.5, the conversion rate is over 85.4% (Table 1, Entry 11, 12, 2), which implied that MaiA–AspA could maintain a similar catalytic performance in the pH range of 6.5~8.5, and the catalytic reaction can be carried out well within this range.
2.4. Optimizing the Bioconversion of L-Aspartic Acid to β-Alanine by Whole Cells of E. coli (ADCtb)
In the three-enzyme tandem system, the enzyme activity of ADCtb was much lower than that of MaiA and AspA, and the decarboxylation of L-aspartic acid was considered as the rate-limiting step of the tandem reaction. Therefore, the conditions of the one-step decarboxylation of ADCtb were studied in detail. We found that the whole cell enzyme activity of ADCtb was quite low. After permeabilization of the ADCtb cell with 0.4% CTAB, the whole cell enzyme activity was dramatically increased from 5.8 U/mg to 141 U/mg. The very low apparent enzyme activity of ADCtb whole cells may be due to the diffusion limitation of cofactor pyridoxal phosphate (PLP). The effects of pH value, cell amount, PLP amount, and substrate concentration on the conversion of decarboxylation reaction were studied (Table 2). The results showed that, in the pH range of 6.5–8.5, the conversion decreased with an increase in pH (Table 2, Entry 1–5), the subsequent reaction conditions are selected when pH=6.5. When the addition level of coenzyme PLP was 1–4 mM (Table 2, Entry 10–13), the conversion showed a trend of rising with an increase in the coenzyme. However, when the addition level increased to 6 mM, the conversion rate decreased (Table 2, Entry 14), so the addition level of coenzyme was 4 mM. There is a correlation between the amount of catalyst and substrate concentration (Table 2, Entry 6–9). Comprehensive experimental results showed that under the condition of 30 g/L cell amount, the substrate concentration was 400 mM could be completely converted to product within 6 h and that more cells and longer time were needed for higher substrate concentrations (Table 2, Entry 1, 15–18).
2.5. Optimization of the One-Pot Three-Enzyme Cascade Reaction
Based on the single enzyme optimization experiment of the MaiA–AspA co-expressed strain and ADCtb strain, the tandem reaction catalyzed by three enzymes was investigated. A series of experiments were designed to explore the optimal conditions for this reaction (Table 3).
Under the same ratio of two kinds of cells, the yield of 3-AP was slightly increased when reaction volume was increased from 5 mL to 30 mL (Table 3, Entry 1 and 2). It indicated that the reaction volume could be enlarged without affecting the reaction efficiency. Adding two kinds of cells at the beginning in one step or in step-by-step mode were studied. Under the same substrate concentration and cell loading, the analytical yield of the final product reached over 93% regardless of one-step or two-step addition of enzymes (Table 3, Entry 2 and 3). However, the reaction time required to achieve the same conversion by stepwise addition of enzymes is relatively longer, which means that the corresponding space–time yield decreased. Therefore, the one-step addition of MaiA–AspA cells and ADCtb cells is better for the three-enzyme cascade reaction. In addition, the change of the initial pH value of the reaction from 6.5 to 8.5 had almost no effect on the final yield of 3-AP in the two-step addition of enzyme experiments (Table 3, Entry 3 and 4), indicating that the tandem reaction system could maintain similar catalytic activity within the pH range of 6.5~8.5. When the substrate concentration was increased from 400 mM to 600 mM and increasing the amount of cells in accordance with the ratio of MaiA–AspA cells to ADCtb cells was kept unchanged (1:5), the conversion remained nearly the same (Table 3, Entry 2 and 5). At 600 mM substrate concentration, the molar conversion of 94.7% can be obtained after a 6 h reaction (Table 3, Entry 5). Considering the consistency of total cells added to the system, the cell ratio was adjusted to 1:4 at 800 mM substrate concentration, and a molar conversion of 93.9% can be achieved after a 9 h reaction (Table 3, Entry 6).
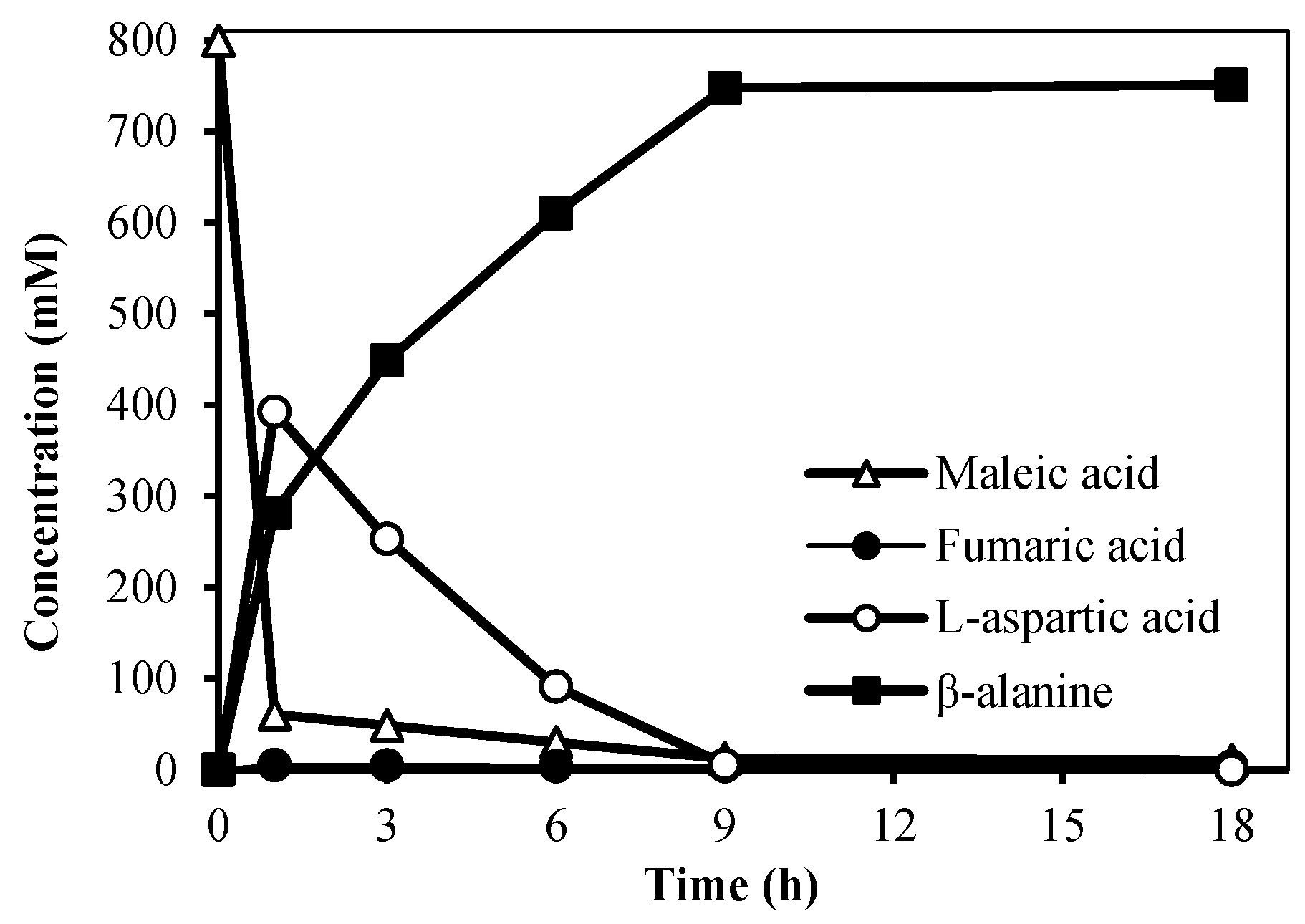
The three-enzyme cascade reaction was studied in detail at the substrate concentration of 800 mM. The reaction process was shown in Figure 4. After a 1 h reaction, ammonium maleate in the reaction system was transformed rapidly; there was almost no accumulation of fumaric acid and the accumulation of L-aspartic acid was 392.6 mM. The concentration of L-aspartic acid gradually decreased to 5.63 mM at 9 h, and the concentration of the final product β-alanine reached 751.2 mM at 9 h, corresponding to a 93.9% yield of β-alanine. At this time, the space–time yield was 178.4 g/L/d.
3. Discussion
In this study, β-alanine was converted from maleic acid through a one-pot three-enzyme cascade reaction by a combination of two kinds of resting cells. Maleic acid, which is much cheaper than fumaric acid and L-aspartic acid, was selected as the substrate to further reduce the cost of the reaction substrate. A multi-enzyme cascade reaction has many advantages, such as no separation of intermediates, saving reaction time, etc. The use of resting cells in the reaction also eliminates the need to add other coenzymes in the reaction. However, it is worth further studying because only the three-enzyme cascade reaction pathway was verified in this paper and was confirmed to be feasible at a high substrate concentration (800 mM). In the future, if the ADC activity is increased to the same order of magnitude as MaiA or AspA, we can try to express all three enzymes in one cell. In addition, only one kind of E. coli strain need be fermented for the reaction, saving the cost and time of the cell preparation process, which may be more convenient and meaningful.
In this paper, we constructed a reaction pathway for the synthesis of β-alanine with low cost and high efficiency by using maleic acid as a substrate and using MaiA, AspA, and ADCtb in one-pot. The reaction was conducted on high substrate concentration (800 mM) with high yield and productivity. It has potential application value for the biosynthesis of β-alanine from a cheap substrate.
4. Materials and Methods
4.1. Strains, Primers, and Chemicals
The recombinant strains (E. coli BL21(DE3)-pRSFduet-maiA-aspA; E. coli BL21 (DE3)-TcADC; E. coli BL21(DE3)-ADCtb) were constructed and preserved in Biocatalysis and Biopharmaceutical laboratory (Shanghai Institute of Technology, Shanghai, China). Plasmids pRSFduet-1 and pET-28a(+) were preserved in Biocatalysis and Biopharmaceutical laboratory (Shanghai Institute of Technology, Shanghai, China). The primers used in this study are shown in Table 4; the primers in the table and genes of MaiA and TcADC were built from Shanghai Generay Biotech company (Shanghai, China). All strains were preserved at −80 °C prior to use. Maleic acid and fumaric acid were bought from Aladdin Company (Shanghai, China), at a purity >95%. L-aspartic acid and β-alanine were bought from Shanghai Titan Company (Shanghai, China), at a purity >98%.
4.2. Cell Culture, Expression, and Enzyme Activity Assay of MaiA, AspB, and ADCtb
4.2.1. Cell Culture and Expression of MaiA and AspB
A pure colony (E. coli BL21(DE3)-pRSFduet-maiA-aspA) was picked and cultivated overnight in 50 mL LB medium with 50 μg/mL kanamycin at 37 °C and 200 rpm. Then, 1 mL of culture was incubated in 100 mL of 2YT medium (Tryptone: 16 g/L, yeast extract powder: 10 g/L, NaCl: 5 g/L), with 50 μg/mL kanamycin, and the cells were cultured at 37 °C and 200 rpm for growth. When the OD600 value reached 0.4 to 0.6, IPTG (0.2 mM) was added to the culture to induce enzyme expression at 20 °C for 20 h. Harvested cells were suspended in 50 mM PBS buffer (pH 7.0) and then ultrasonicated on ice. Centrifugation was used to remove cell debris for 10 min at 12,000× g and 4 °C. The protein molecular mass was evaluated by SDS–PAGE.
4.2.2. Cell Culture and Expression of ADCtb
A pure colony (E. coli BL21(DE3)-ADCtb) was picked and cultivated overnight in 50 mL LB medium with 50 μg/mL kanamycin at 37 °C and 200 rpm. Then, 1 mL of culture was incubated in 100 mL of 2YT medium (Tryptone: 16 g/L, yeast extract powder: 10 g/L, NaCl: 5 g/L) with 50 μg/mL kanamycin and 10 g/L glucose, and the cells were cultured at 37 °C and 200 rpm for growth. When the OD600 value reached 0.4 to 0.6, IPTG (0.2 mM) was added to the culture to induce enzyme expression at 20 °C for 20 h. Harvested cells were suspended in 50 mM PBS buffer (pH 6.5) and then ultrasonicated on ice. Centrifugation was used to remove cell debris for 10 min at 12,000× g and 4 °C. The protein molecular mass was evaluated by SDS–PAGE.
4.2.3. Enzyme Activity Assay of MaiA, AspB, and ADCtb
MaiA activity assay: The resting cells were obtained by centrifugation from 1 mL fermentation solution. We added 50 μL maleic acid solution (2 mol/L, adjusted pH to 8.4 by KOH), filled the reaction system to 1 mL with Tris-HCL solution (pH = 8.4), created a full suspension, and the reaction was performed at 37 °C for 10 min followed by boiling for 10 min. After centrifugation at 12,000× g for 5 min, the reaction was filtered with a 0.45 μm organic filter membrane and assessed by HPLC. One unit of MaiA activity was defined as the amount of enzyme that catalyzes the conversion of 1 μmol of maleic acid per minute under the described conditions.
AspA activity assay: The resting cells were obtained by centrifugation from 1 ml fermentation solution. We added 100 mM ammonium fumarate and 1 mM Mg2+, filled the reaction system to 1 mL with PBS solution (pH = 7.0), created a full suspension, and the reaction was performed at 37 °C for 10 min followed by boiling for 10 min. After centrifugation at 12,000× g for 5 min, the reaction was filtered with an 0.45 μm organic filter membrane and assessed by HPLC. One unit of AspA activity was defined as the amount of enzyme that catalyzes the production of 1 μmol aspartic acid per minute under the described conditions.
ADC activity assay: The resting cells were obtained by centrifugation from 1 ml of fermentation solution. We added 100 mM L-aspartic acid sodium solution (pH = 6.5) and 1 mM pyridoxal phosphate (PLP), filled the reaction system to 1 mL with PBS solution (pH = 6.5), created a full suspension, and the reaction was performed at 37 °C for 20 min followed by boiling for 10 min. After centrifugation at 12,000× g for 5 min, the reaction was filtered with a 0.45 μm organic filter membrane and assessed by HPLC. One unit of ADC activity is defined as the amount of enzyme that catalyzes the production of 1 μmol β-alanine per minute under the described conditions.
4.3. Determination of Maleic Acid, Fumaric Acid, L-Aspartic Acid, and β-Alanine
The concentrations of maleic acid, fumaric acid, L-aspartic acid, and β-alanine were determined by HPLC (LC-20AT, Shimadzu, Kyoto, Japan). The determinate conditionsare shown in Table 5. Maleic acid and fumaric acid can be detected directly, while L-aspartic acid and β -alanine could be detected only after derivatization. The derivatization method is as follows: add 200 μL of 0.1 M phenyl isothiocyanate solution in ACN and 200 μL of 1 M triethylamine solution in ACN into 400 μL of reaction solution. Mix thoroughly and place in the dark for 1 h, then extract by 500 μL n-hexane for 30–60 s, ad leave to stand for 30 min. The lower layer solution is filtered through a 0.45 μm filter membrane for HPLC analysis (retention time: L-aspartic acid: 4.46 min; β-alanine: 9.60 min; Maleic acid: 5.47 min; Fumaric acid: 6.73 min).
4.4. Optimization of the Dual-Enzyme Catalyzed Reaction
4.4.1. Optimization of Reaction pH
The reaction was conducted in the conditions as follows: Substrate (ammonium maleate): 400 mM; Catalyst (MaiA–AspA resting cells): 3 gdcw/L; MgSO4: 1 mM; Buffer: PBS solution (0.1 M), Reaction volume: 5 mL. Reactions were conducted on the orbital shaker (37 °C, 200 r/min). The pH values were set as 5.5, 6.5; 7.5, 8.5, and 9.5.
4.4.2. Optimization of Substrate Concentration
The reaction was conducted in the conditions as follows: Catalyst (MaiA–AspA resting cells): 3 g/L, MgSO4: 1 mM, Buffer: PBS solution (0.1 M), pH: 8.5, Reaction volume: 5 mL. Reactions were conducted on the orbital shaker (37 °C, 200 r/min). The substrate concentrations were set as 300, 400, 600, 800, and 1000 mM.
4.4.3. Optimization of Catalyst Concentration
The reaction system was conducted in the conditions as follows: Substrate (ammonium maleate): 400 mM, MgSO4: 1 mM, Buffer: PBS solution (0.1 M), pH: 8.5, Reaction volume: 5 mL. Reactions were conducted on the orbital shaker (37 °C, 200 r/min). The catalyst (MaiA–AspA resting cells) concentration was set as 1.5, 3.0, 4.5, 6.0, and 10 g/L.
4.5. Optimization of ADCtb Catalyzed Reaction
4.5.1. Permeabilization of ADCtb Cells
The ADCtb resting cells were collected by centrifugation in fermentation solution and washed twice with 0.9% NaCl solution. Then, 0.4% CTAB solution in the same volume of fermentation solution was added and incubated at 30 °C for 30 min. The cells were centrifuged again and washed twice with 0.9% NaCl solution.
4.5.2. Optimization of Reaction pH
The reaction was conducted in the conditions as follows: Substrate (L-aspartic acid): 400 mM, Catalyst (ADCtb resting cells): 30 g/L, PLP: 1 mM, Buffer: PBS solution (0.1 M) or Tris-HCl solution (0.1M), Reaction volume: 5 mL. Reaction was conducted on the orbital shaker (37 °C, 200 r/min). The pH values were set as 6.5, 7.0, 7.5, 8.0, and 8.5 (Buffer pH 8.0 and 8.5 were Tris-HCl solution, others use PBS buffer).
4.5.3. Optimization of Catalyst Concentration
The reaction was conducted in the conditions as follows: Substrate (L-aspartic acid): 200 mM, PLP: 1 mM, Buffer: PBS solution (0.1 M), pH: 6.5, Reaction volume: 5 mL. Reaction was conducted on the orbital shaker (37 °C, 200 r/min). The catalyst concentration (ADCtb resting cells) was set as 20, 30, 40, and 50 g/L.
4.5.4. Optimization of PLP Concentration
The reaction system was conducted in the conditions as follows: Substrate (L-aspartic acid): 200 mM, Catalyst (ADCtb resting cells): 20 g/L, Buffer: PBS solution (0.1 M), pH: 6.5, Reaction volume: 5 mL. Reaction was conducted on the orbital shaker (37 °C, 200 r/min). The PLP concentration was set as 0.5, 1, 2, 4, and 6 mM.
4.5.5. Optimization of Substrate Concentration
The reaction was conducted in the conditions as follows: Catalyst (ADCtb resting cells): 30 g/L, PLP: 4 mM, Buffer: PBS solution (0.1 M), pH: 6.5, Reaction volume: 5 mL. Reaction was conducted on the orbital shaker (37 °C, 200 r/min). The substrate (L-aspartic acid) concentration was set as 400, 500, 600, 700, and 800 mM.
4.6. Optimization of the One-Pot Three-Enzyme Cascade Reaction
4.6.1. Selection of Two Resting Cells Addition Mode
Adding two kinds of cells simultaneously or step-by-step were studied to evaluate the effect of the adding mode of two cells. The reaction was conducted in the conditions as follows: Substrate (ammonium maleate): 400 mM, MgSO4: 1 mM, PLP: 4 mM, Catalyst (MaiA–AspA resting cells): 6 g/L, Catalyst (ADCtb resting cells): 30 g/L. Buffer: PBS solution (0.1 M, pH 6.5). Entry 1: Reaction volume was 5 mL, two catalysts were added at the same time, and the initial pH value of the reaction was adjusted to 6.5. Entry 2: Reaction volume was 30 mL, two catalysts were added at the same time, and the initial pH value of the reaction was adjusted to 6.5. Entry 3: Reaction volume was 30 mL, MaiA–AspA resting cells were added first, and the initial pH of the reaction was adjusted to 6.5. ADCtb was added after 2 h and the pH of the reaction was adjusted to 6.5. Entry 4: Reaction volume was 30 mL, MaiA–AspA resting cells were added first, and the initial pH of the reaction was adjusted to 8.5. ADCtb was added after 2 h and the pH of the reaction was adjusted to 6.5. The reactions were conducted on the orbital shaker (37 °C, 200 r/min).
4.6.2. Optimization of the Three-Enzyme Cascade Reaction
The effect of the simultaneous increase in the substrate concentration and cell volume on the reaction results was confirmed. The reaction was conducted in the conditions as follows: MgSO4: 1 mM, PLP: 4 mM, Buffer: 30 mL PBS solution (0.1 M, pH 6.5), Reaction volume was 30 mL. Entry 5: Substrate (ammonium maleate): 600 mM, Catalyst (MaiA–AspA resting cells): 7 g/L, Catalyst (ADCtb resting cells): 35 g/L. Entry 6: Substrate (ammonium maleate): 800 mM, Catalyst (MaiA–AspA resting cells): 10 g/L, Catalyst (ADCtb resting cells): 40 g/L. The reactions were conducted on the orbital shaker (37 °C, 200 r/min).
5. Conclusions
A novel and efficient one-pot three-enzyme cascade method for the synthesis of β-alanine from maleic acid was developed. Two recombinant E. coli strains were constructed. After systematic optimization of reaction conditions for each strain, the whole cells of two strains were combined for one-pot synthesis of β-alanine. Under optimal conditions, the concentration of β-alanine reached 751 mM after a 9 h reaction, corresponding to a 93.9% yield and 178 g/L/d space-time yield. The developed new route showed application potential for green and efficient biosynthesis of β-alanine from a cheap substrate by tandem biocatalysts.
Author Contributions
Conceptualization, Y.X. and B.-D.M.; methodology, Y.X. and B.-D.M.; validation, J.W.; formal analysis, J.W.; data curation, J.W.; writing—original draft preparation, J.W.; writing—review and editing, J.W. and Y.X.; funding acquisition, Y.X. and B.-D.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Open Project Funding of the State Key Laboratory of Biocatalysis and Enzyme Engineering (Grant No. SKLBEE2018008), Project of Leading Talents in Shandong Taishan Industry (Grant No. LJNY202019).
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Luo, J.X.; Xue, J.P.; Shen, Y.C. Synthesis and application of β-Alanine. Amino Acids Biotic Resour. 2005, 27, 52–55. (In Chinese) [Google Scholar]
- Buc, S.R.; Ford, J.H.; Wise, E.C. An improved synthesis of β-Alanine. J. Am. Chem. Soc. 1945, 67, 92–94. [Google Scholar] [CrossRef]
- Ren, Y.; Wang, Y.Q.; Shu, H.; Guo, C. New synthetic process of β-Aminopropanoic Acid. Liaoning Chem. Ind. 2006, 35, 187–188. (In Chinese) [Google Scholar]
- Ford, J.H. The alkaline hydrolysis of β-aminopropionitrile. J. Am. Chem. Soc. 1945, 67, 876–877. [Google Scholar] [CrossRef]
- Lou, J. Study on the production of β-alanine by biotransformation. Ph.D. Thesis, Zhejiang University of Technology, Hang Zhou, China, 2006. (In Chinese). [Google Scholar]
- Liang, L.Y.; Zheng, Y.G.; Shen, Y.C. Optimization of β-alanine production from β-aminopropionitrile by resting cells of Rhodococcus sp. G20 in a bubble column reactor using response surface methodology. Process Biochem. 2008, 43, 758–764. [Google Scholar] [CrossRef]
- Nozaki, S.; Webb, M.E.; Niki, H. An activator for pyruvoyl-dependent L-aspartate -α-decarboxylase is conserved in a small group of the γ-proteobacteria including Escherichia coli. MicrobiologyOpen 2012, 1, 298–310. [Google Scholar] [CrossRef]
- Shen, Y.; Zhao, L.Z.; Li, Y.R.; Zhang, L.; Shi, G.Y. Synthesis of β-alanine from L-aspartate using L-aspartate-α-decarboxylase from Corynebacterium glutamicum. Biotechnol. Lett. 2014, 36, 1681–1686. [Google Scholar] [CrossRef]
- Pei, W.L.; Zhang, J.L.; Deng, S.Y.; Tigu, F.; Li, Y.X.; Li, Q.; Cai, Z.; Li, Y. Molecular engineering of L-aspartate-α-decarboxylase for improved activity and catalytic stability. Appl. Microbiol. Biot. 2017, 101, 1–7. [Google Scholar] [CrossRef]
- Mo, Q.; Li, Y.R.; Wang, J.H.; Shi, G.Y. Identifcation of mutations restricting autocatalytic activation of bacterial L-aspartate-α-decarboxylase. Amino Acids 2018, 50, 1433–1440. [Google Scholar] [CrossRef]
- Zhao, L.Z.; Zhang, L.; Shi, G.Y. Expression of Corynebacterium glutamate L-aspartate-α-decarboxylase in Escherichia coli and enzyme transformation to produce β-alanine. Microbiol. China 2013, 40, 2161–2170. (In Chinese) [Google Scholar]
- Yao, P.Y.; Cui, Y.F.; Yu, S.S.; Du, Y.C.; Feng, J.H.; Wu, Q.Q.; Zhu, D.M. Efficient biosynthesis of (R)- or (S)-2-hydroxybutyrate from L-threonine through a synthetic biology approach. Adv. Synth. Catal. 2016, 358, 2923–2928. [Google Scholar] [CrossRef]
- Zhang, J.D.; Wu, S.; Wu, J.C.; Li, Z. Enantioselective cascade biocatalysis via epoxide hydrolysis and alcohol oxidation: One-pot synthesis of (R)-α-hydroxy ketones from meso- or racemic epoxides. ACS Catal. 2015, 5, 51–58. [Google Scholar] [CrossRef]
- Wu, S.; Liu, J.; Li, Z. Biocatalytic formal anti-markovnikov hydroamination and hydration of aryl alkenes. ACS Catal. 2017, 7, 5225–5233. [Google Scholar] [CrossRef]
- Busto, E.; Gerstmann, M.; Tobola, F.; Dittmann, E.; Wiltschi, B.; Kroutil, W. Systems biocatalysis: Para-alkenylation of unprotected phenols. Catal. Sci. Technol. 2016, 6, 8098–8103. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, K.; Bujons, J.; Joglar, J.; Charnock, S.J.; Pablo, D.D.M.; Fessner, W.D.; Clapés, P. Combining aldolases and transaminases for the synthesis of 2-Amino-4-hydroxybutanoic Acid. ACS Catal. 2017, 7, 1707–1711. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Sekar, B.S.; Wu, S.; Li, Z. Benzoic acid production via cascade biotransformation and coupled fermentation-biotransformation. Biotechnol. Bioeng. 2020, 117, 2340–2350. [Google Scholar] [CrossRef]
- Busto, E.; Richter, N.; Grischek, B.; Wolfgang, K. Biocontrolled formal inversion or retention of L-α-amino acids to enantiopure (R)- or (S)-hydroxyacids. Chem. Eur. J. 2014, 20, 11225–11228. [Google Scholar] [CrossRef]
- Zhang, J.D.; Zhao, J.W.; Gao, L.L.; Zhao, J.; Chang, H.H.; Wei, W.L. One-pot three-step consecutive transformation of L-α-amino acids to (R)- and (S)-vicinal 1,2-diols via combined chemical and biocatalytic process. ChemCatChem 2019, 11, 5032–5037. [Google Scholar] [CrossRef]
- Gourinchas, G.; Busto, E.; Killinger, M. A synthetic biology approach for the transformation of L-α-amino acids to the corresponding enantiopure (R)- or (S)-α-hydroxy acids. Chem. Commun. 2015, 51, 2828–2831. [Google Scholar] [CrossRef]
- Busto, E.; Simon, R.C.; Richter, N.; Kroutil, W. One-pot, two-module three-step cascade to transform phenol derivatives to enantiomerically pure (R)- or (S)-p-hydroxyphenyl lactic acids. ACS Catal. 2016, 6, 2393–2397. [Google Scholar] [CrossRef]
- Zhou, H.; Meng, L.; Yin, X.; Liu, Y.; Yang, L. Biocatalytic asymmetric synthesis of L-phosphinothricin using a one-pot three enzyme system and a continuous substrate fed-batch strategy. Appl. Catal. A Gen. 2019, 589, 117239. [Google Scholar] [CrossRef]
- Lukito, B.R.; Wu, S.; Saw, H.; Li, Z. One-pot production of natural 2-phenylethanol from L-phenylalanine via cascade biotransformations. ChemCatChem 2019, 11, 831–840. [Google Scholar] [CrossRef]
- Sun, Z.B.; Zhang, Z.J.; Li, F.L.; Nie, Y.; Yu, H.L.; Xu, J.H. One pot asymmetric synthesis of (R)-phenylglycinol from racemic styrene oxide via cascade biocatalysis. ChemCatChem 2019, 11, 3802–3807. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Chen, Y.; Zhou, L.; Zhou, Z.M. Whole-cell biocatalysis of maleic acid into L-aspartic acid by dual-enzyme coupling. Food Ferment. Ind. 2018, 44, 20–26. (In Chinese) [Google Scholar]
- Gao, Y.; Liu, Z.M.; Liu, K.; Zhou, Z.M.; Cui, W.J. Biocatalytic access to β-alanine by a two-enzyme cascade synthesis. Chin. J. Biotech. 2017, 33, 875–879. (In Chinese) [Google Scholar]
- Wang, C.; Ye, W.Q.; Xue, L.; Liu, Z.M.; Zhou, Z.M. Molecular modification of aspartate α-decarboxylase from Tribolium castaneum and its application in the production of β-alanine. Food Ferment. Ind. 2019, 45, 7–13. (In Chinese) [Google Scholar]
- Liu, P.Y.; Ding, H.Z.; Bruce, M.C.; Li, J.Y. Cysteine sulfinic acid decarboxylase activity of Aedes aegypti aspartate 1-decarboxylase: The structural basis of its substrate selectivity. Insect Biochem. Mol. Biol. 2012, 42, 396–403. [Google Scholar] [CrossRef]
Figure 1.
Enzymatic synthesis of β-alanine from maleic acid by a three-enzyme catalyzed cascade reaction in one pot.
Figure 1.
Enzymatic synthesis of β-alanine from maleic acid by a three-enzyme catalyzed cascade reaction in one pot.

Figure 2.
Nucleic acid electrophoresis assay of recombinant MaiA-AspA and ADCtb. (M: DNA marker; 1/3: pRSF-MaiA-AspA BamH I/Hind III digestion; 2/4: pRSF-MaiA-AspA Ned I/Xho I digestion; 5: pET28a-ADCtb Nco I / EcoR I digestion).
Figure 2.
Nucleic acid electrophoresis assay of recombinant MaiA-AspA and ADCtb. (M: DNA marker; 1/3: pRSF-MaiA-AspA BamH I/Hind III digestion; 2/4: pRSF-MaiA-AspA Ned I/Xho I digestion; 5: pET28a-ADCtb Nco I / EcoR I digestion).

Figure 3.
SDS-PAGE assay of recombinant MaiA-AspA and ADCtb. (M: protein marker; 1: supernatant of cell extract from E. coli (MaiA-AspA); 2: precipitation of cell extract from E. coli (MaiA-AspA); 3: supernatant of cell extract from E. coli (ADCtb); 4: precipitation of cell extract from E. coli (ADCtb).
Figure 3.
SDS-PAGE assay of recombinant MaiA-AspA and ADCtb. (M: protein marker; 1: supernatant of cell extract from E. coli (MaiA-AspA); 2: precipitation of cell extract from E. coli (MaiA-AspA); 3: supernatant of cell extract from E. coli (ADCtb); 4: precipitation of cell extract from E. coli (ADCtb).

Figure 4.
Three-enzyme cascade catalytic synthesis of β-alanine.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Enzymatic synthesis of L-aspartic acid from maleic acid by co-expressed MaiA and AspA under different conditions.
Table 1.
Enzymatic synthesis of L-aspartic acid from maleic acid by co-expressed MaiA and AspA under different conditions.
| Entry | Maleic Acid (mM) | Cell Amount (g/L) | pH | Conv. (%) |
|---|---|---|---|---|
| 1 | 300 | 3 | 8.5 | 93.3 |
| 2 | 400 | 3 | 8.5 | 85.4 |
| 3 | 600 | 3 | 8.5 | 75.2 |
| 4 | 800 | 3 | 8.5 | 73.5 |
| 5 | 1000 | 3 | 8.5 | 57.0 |
| 6 | 400 | 1.5 | 8.5 | 77.5 |
| 7 | 400 | 4.5 | 8.5 | 95.1 |
| 8 | 400 | 6 | 8.5 | 97.5 |
| 9 | 400 | 10 | 8.5 | 99.4 |
| 10 | 400 | 3 | 5.5 | 75.2 |
| 11 | 400 | 3 | 6.5 | 88.0 |
| 12 | 400 | 3 | 7.5 | 90.6 |
| 13 | 400 | 3 | 9.5 | 82.7 |
Table 2.
Enzymatic synthesis of β-alanine from L-aspartic acid by permeabilized ADCtb under different conditions.
Table 2.
Enzymatic synthesis of β-alanine from L-aspartic acid by permeabilized ADCtb under different conditions.
| Entry | Substrate (mM) | E. coli (g/L) | pH | PLP (mM) | Time (h) | Conv. (%) |
|---|---|---|---|---|---|---|
| 1 | 400 | 30 | 6.5 | 4 | 6 | 100.0 |
| 2 | 400 | 30 | 7.0 | 4 | 6 | 72.2 |
| 3 | 400 | 30 | 7.5 | 4 | 6 | 58.8 |
| 4 | 400 | 30 | 8.0 | 4 | 6 | 45.3 |
| 5 | 400 | 30 | 8.5 | 4 | 6 | 36.5 |
| 6 | 200 | 20 | 6.5 | 1 | 1 | 65.2 |
| 7 | 200 | 30 | 6.5 | 1 | 1 | 80.2 |
| 8 | 200 | 40 | 6.5 | 1 | 1 | 94.2 |
| 9 | 200 | 50 | 6.5 | 1 | 1 | 98.8 |
| 10 | 200 | 20 | 6.5 | 0.5 | 6 | 89.2 |
| 11 | 200 | 20 | 6.5 | 1 | 6 | 91.2 |
| 12 | 200 | 20 | 6.5 | 2 | 6 | 89.4 |
| 13 | 200 | 20 | 6.5 | 4 | 6 | 93.1 |
| 14 | 200 | 20 | 6.5 | 6 | 6 | 90.0 |
| 15 | 500 | 30 | 6.5 | 4 | 24 (6) | 100 (89.7) |
| 16 | 600 | 30 | 6.5 | 4 | 24 | 100 |
| 17 | 700 | 30 | 6.5 | 4 | 24 | 97.1 |
| 18 | 800 | 30 | 6.5 | 4 | 24 | 91.2 |
Table 3.
One-pot synthesis of β-alanine from maleic acid by three-enzyme cascade catalysis under different conditions.
Table 3.
One-pot synthesis of β-alanine from maleic acid by three-enzyme cascade catalysis under different conditions.
| Entry | Sub. Conc. /mM | MaiA–AspA (g/L) | ADCtb (g/L) | Vol. (mL) | Time (h) | Yield of 3-AP (%) | Space–Time Yield (g/L/d) | Ways to Add Enzymes | Initial pH |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 400 | 6 | 30 | 5 | 6 | 94.0 | 134.0 | One step | 6.5 |
| 2 | 400 | 6 | 30 | 30 | 6 | 95.6 | 136.3 | One step | 6.5 |
| 3 | 400 | 6 | 30 | 30 | 9 | 93.9 | 89.2 | Two step | 6.5 |
| 4 | 400 | 6 | 30 | 30 | 9 | 94.0 | 89.3 | Two step | 8.5 |
| 5 | 600 | 7 | 35 | 30 | 6 | 94.7 | 202.5 | One step | 6.5 |
| 6 | 800 | 10 | 40 | 30 | 9 | 93.9 | 178.4 | One step | 6.5 |
Table 4.
Primers used for the construction of strains.
| Gene | Primer | Restriction Site | Sequence(5′→3′) |
|---|---|---|---|
| MaiA | F1 | BamH I | CGGGATCCGATGAGTAATCATTATCGTATTGGTCAGA |
| R1 | Hind Ⅲ | CCCAAGCTTTTAATATGCGCCAGAGAGAAGGG | |
| AspA | F2 | Ned I | GGAATTCCATATGATGTCAAACAACATTCGTATCGAAG |
| R2 | Sac I | CGAGCTCTTACTGTTCGCTTTCATCAGTATAG | |
| R2’ | Xho I | CCGCTCGAGTTACTGTTCGCTTTCATCAGTATAGCG | |
| ADCtb | F3 | - | CCGGATATTAAAAAAAGAGGCTTACATAGTCTG |
| R3 | - | CAGACTATGTAAGCCTCTTTTTTTAATATCCGG |
Table 5.
HPLC analysis methods.
| Compound | L-Aspartic Acid & β-Alanine | Maleic Acid & Fumaric Acid |
|---|---|---|
| Column | C18 column (Diamonsil plus, 4.6 mm × 250 mm × 5 µm) | |
| Phase | Phase A: 80% ACN-H2O solution Phase B: 97:3 (0.1 M Sodium acetate: ACN solution; adjust pH to 6.5 by acetic acid) | 25 mM KH2PO4 solution (pH = 2.5) |
| Methods | 0–15 min: 95%–65% phase B 15–20 min: 65%–95% phase B 20–30 min: 95% phase B | 10 min |
| Flow rate | 1 mL/min | 1 mL/min |
| Wavelength | 254 nm | 210 nm |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wu, J.; Ma, B.-D.; Xu, Y. One-Pot Synthesis of β-Alanine from Maleic Acid via Three-Enzyme Cascade Biotransformation. Catalysts 2023, 13, 267. https://0-doi-org.brum.beds.ac.uk/10.3390/catal13020267
AMA Style
Wu J, Ma B-D, Xu Y. One-Pot Synthesis of β-Alanine from Maleic Acid via Three-Enzyme Cascade Biotransformation. Catalysts. 2023; 13(2):267. https://0-doi-org.brum.beds.ac.uk/10.3390/catal13020267
Chicago/Turabian StyleWu, Jia, Bao-Di Ma, and Yi Xu. 2023. "One-Pot Synthesis of β-Alanine from Maleic Acid via Three-Enzyme Cascade Biotransformation" Catalysts 13, no. 2: 267. https://0-doi-org.brum.beds.ac.uk/10.3390/catal13020267
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.