Bacterial Colonization on the Surface of Copper Sulfide Minerals Probed by Fourier Transform Infrared Micro-Spectroscopy

Abstract
:1. Introduction
2. Materials and Methods
2.1. Extraction of Extracellular Polymeric Substances (EPS) from Bacterial Cells
2.2. FTIR and Raman Microspectroscopy
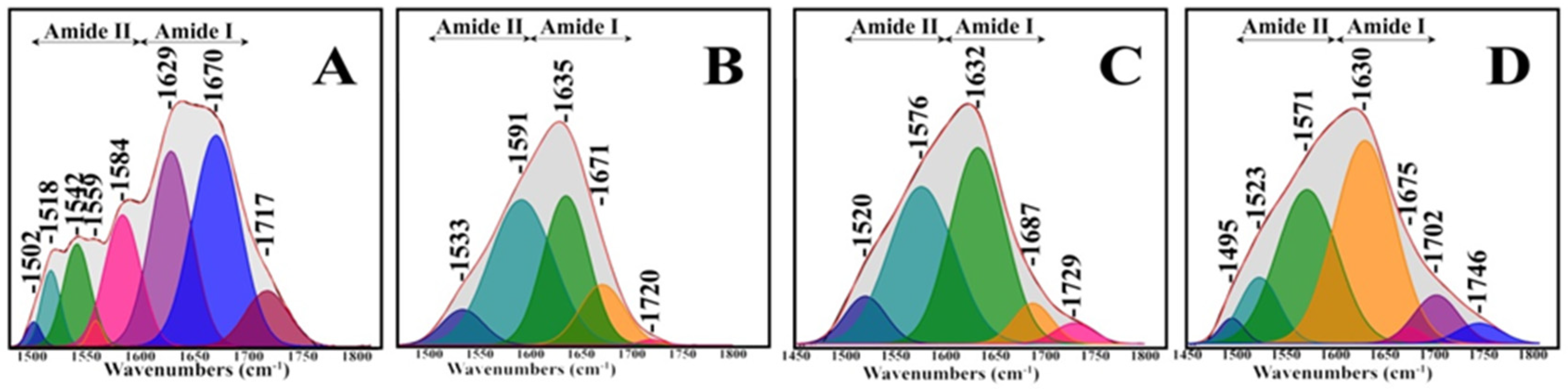
2.3. Deconvolution of Amide I
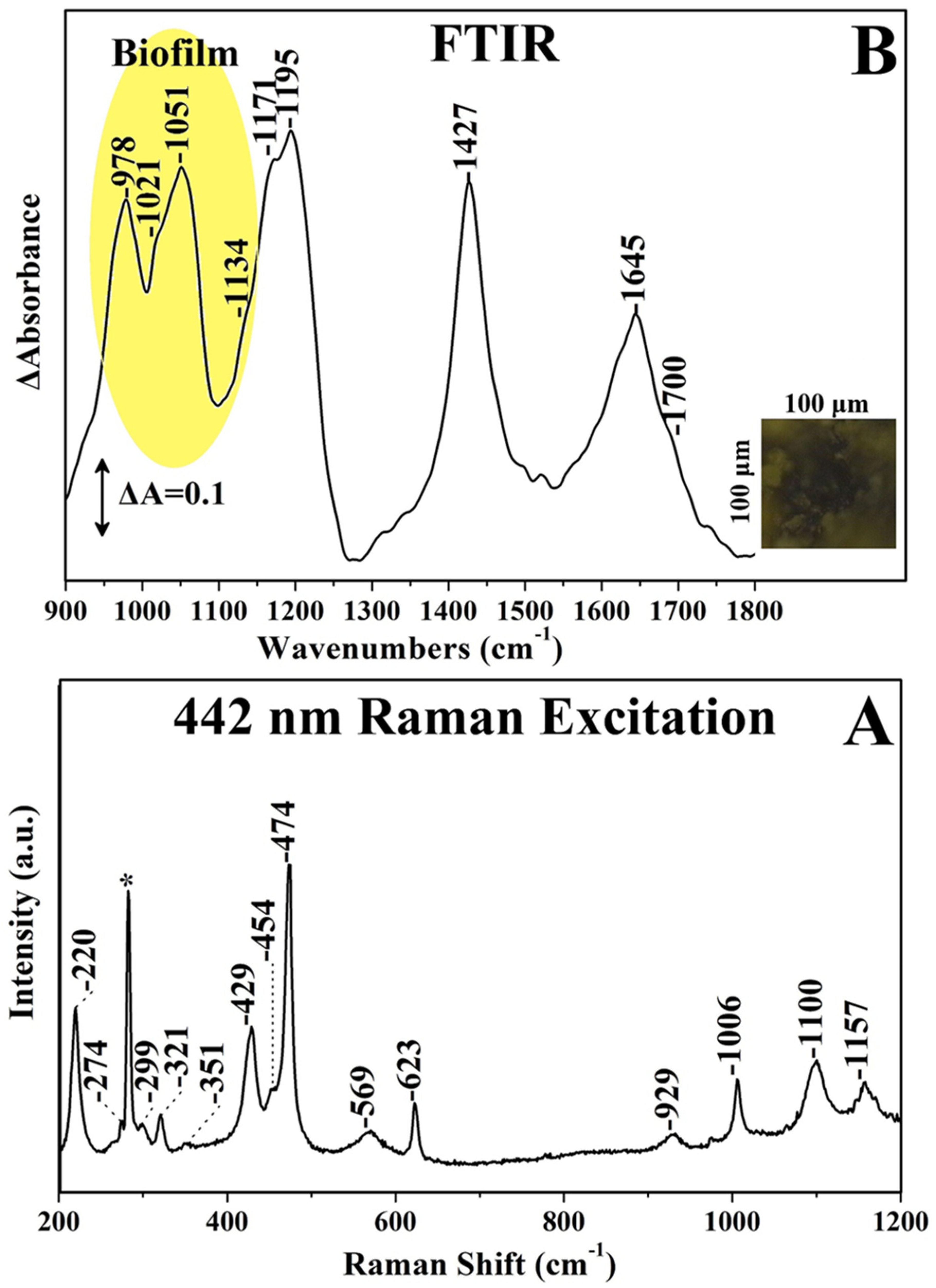
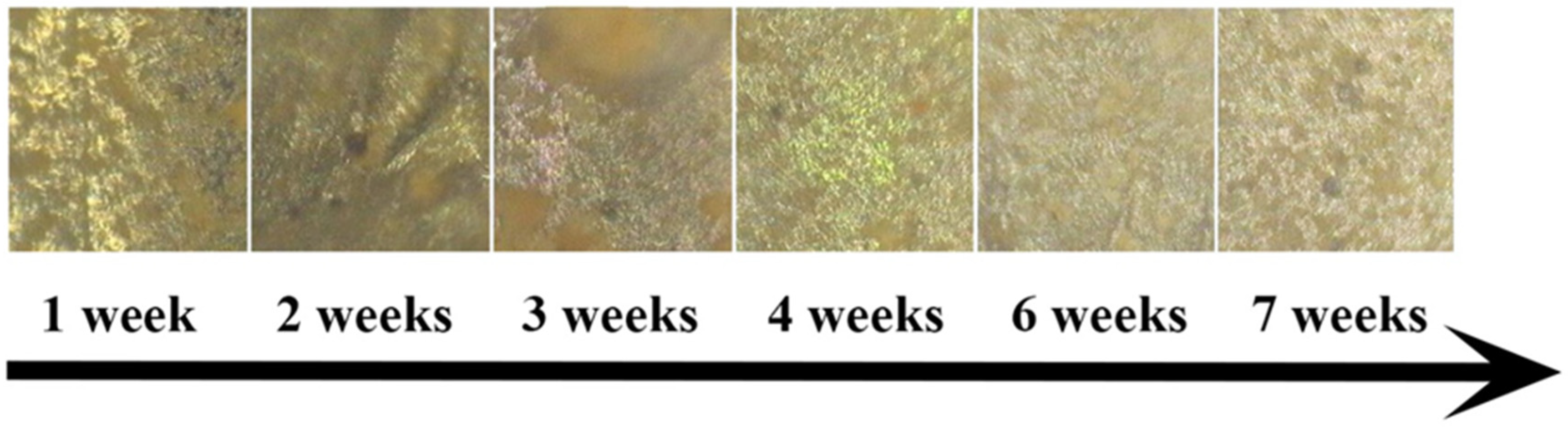
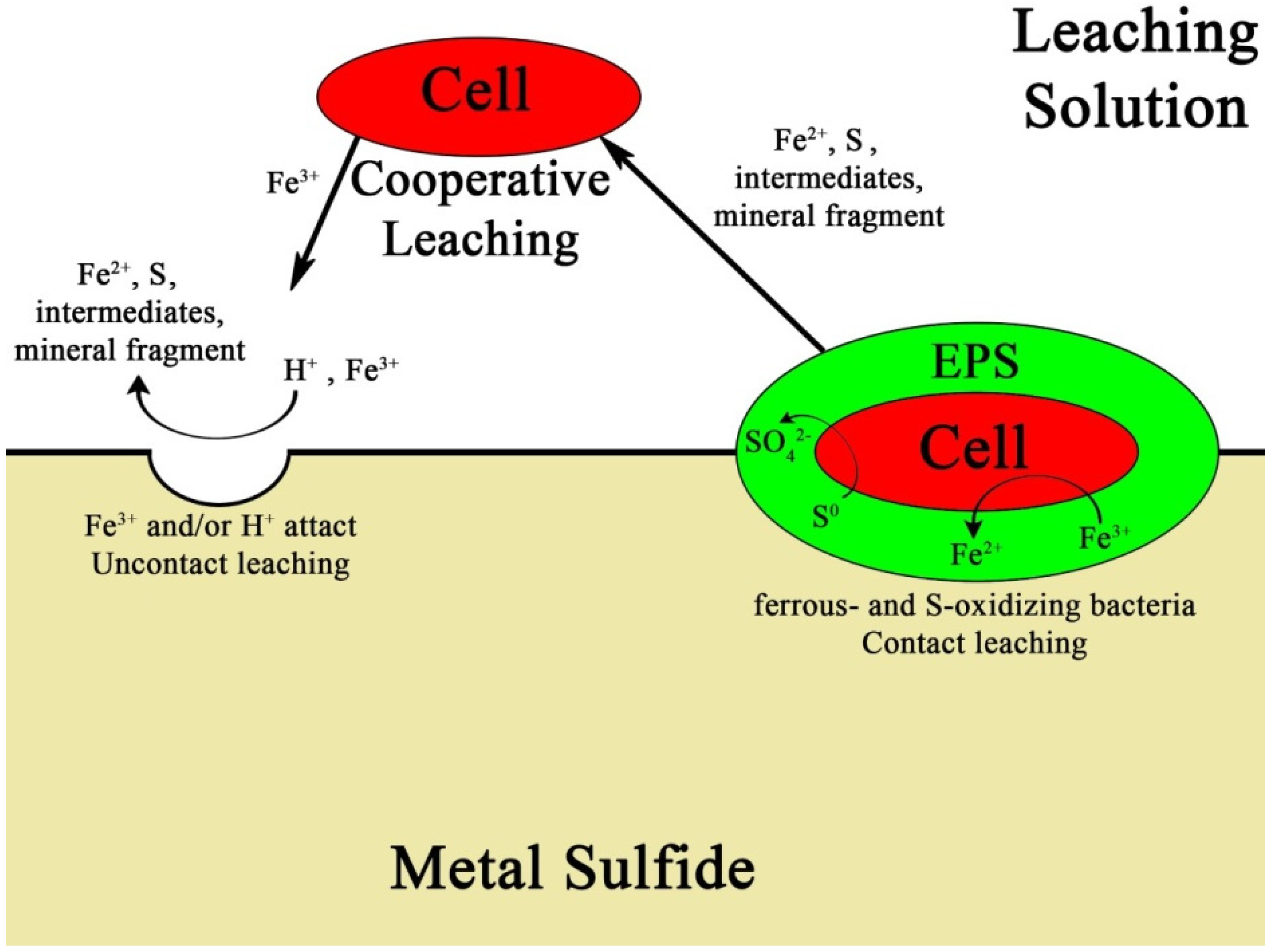
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jiang, L.; Zhou, H.; Peng, X. Bio-Oxidation of Pyrite, Chalcopyrite and Pyrrhotite by Acidithiobacillus Ferrooxidans. Chin. Sci. Bull. 2007, 52, 2702–2714. [Google Scholar] [CrossRef]
- Zhao, H.; Wang, J.; Gan, X.; Zheng, X.; Tao, L.; Hu, M.; Li, Y.; Qin, W.; Qiu, G. Effects of Pyrite and Bornite on Bioleaching of Two Different Types of Chalcopyrite in the Presence of Leptospirillum Ferriphilum. Bioresour. Technol. 2015, 194, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Adamou, A.; Manos, G.; Messios, N.; Georgiou, L.; Xydas, C.; Varotsis, C. Probing the Whole Ore Chalcopyrite–bacteria Interactions and Jarosite Biosynthesis by Raman and FTIR Microspectroscopies. Bioresour. Technol. 2016, 214, 852–855. [Google Scholar] [CrossRef] [PubMed]
- Adamou, A.; Nicolaides, A.; Varotsis, C. Bio-hydrometallurgy dynamics of copper sulfide-minerals probed by micro-FTIR mapping and Raman microspectroscopy. Miner. Eng. 2019, 132, 39–47. [Google Scholar] [CrossRef]
- Di Giambattista, L.; Grimaldi, P.; Udroiu, I.; Pozzi, D.; Cinque, G.; Giansanti, A.; Congiu Castellano, A. FTIR Spectral Imaging as a Probe of Ultrasound Effect on Cells in Vitro. arXiv 2009, 2, arXiv:1007.0864. [Google Scholar]
- Schmitt, Y.; Hähl, H.; Gilow, C.; Mantz, H.; Jacobs, K.; Leidinger, O.; Bellion, M.; Santen, L. Structural Evolution of Protein-Biofilms: Simulations and Experiments. Biomicrofluidics 2010, 4, 32201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutch, L.A.; Watling, H.R.; Watkin, E.L.J. Microbial Population Dynamics of Inoculated Low-Grade Chalcopyrite Bioleaching Columns. Hydrometallurgy 2010, 104, 391–398. [Google Scholar] [CrossRef]
- Yang, Y.; Tan, S.N.; Glenn, A.M.; Harmer, S.; Bhargava, S.; Chen, M. A Direct Observation of Bacterial Coverage and Biofilm Formation by Acidithiobacillus Ferrooxidans on Chalcopyrite and Pyrite Surfaces. Biofouling 2015, 31, 575–586. [Google Scholar] [CrossRef]
- Vera, M.; Krok, B.; Bellenberg, S.; Sand, W.; Poetsch, A. Shotgun Proteomics Study of Early Biofilm Formation Process of Acidithiobacillus Ferrooxidans ATCC 23270 on Pyrite. Proteomics 2013, 13, 1133–1144. [Google Scholar] [CrossRef]
- Diao, M.; Taran, E.; Mahler, S.; Nguyen, A.V. A Concise Review of Nanoscopic Aspects of Bioleaching Bacteria-Mineral Interactions. Adv. Colloid Interface Sci. 2014, 212, 45–63. [Google Scholar] [CrossRef] [Green Version]
- Gallagher, W. FTIR Analysis of Protein Structure. Biochemistry 1997, 1958, 662–666. [Google Scholar]
- Litvinov, R.I.; Faizullin, D.A.; Zuev, Y.F.; Weisel, J.W. The α-Helix to β-Sheet Transition in Stretched and Compressed Hydrated Fibrin Clots. Biophys. J. 2012, 103, 1020–1027. [Google Scholar] [CrossRef] [Green Version]
- Troullier, A.; Reinstädler, D.; Dupont, Y.; Naumann, D.; Forge, V. Transient Non-Native Secondary Structures during the Refolding of α- Lactalbumin Detected by Infrared Spectroscopy. Nat. Struct. Biol. 2000, 7, 78–86. [Google Scholar] [PubMed]
- Williams, S.; Causgrove, T.P.; Gilmanshin, R.; Fang, K.S.; Callender, R.H.; Woodruff, W.H.; Dyer, R.B. Fast Events in Protein Folding: Helix Melting and Formation in a Small Peptide. Biochemistry 1996, 35, 691–697. [Google Scholar] [CrossRef]
- Amenabar, I.; Poly, S.; Nuansing, W.; Hubrich, E.H.; Govyadinov, A.A.; Huth, F.; Krutokhvostov, R.; Zhang, L.; Knez, M.; Heberle, J.; et al. ARTICLE Structural Analysis and Mapping of Individual Protein Complexes by Infrared Nanospectroscopy. Nat. Commun. 2013, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gong, A.S.; Bolster, C.H.; Benavides, M.; Walker, S.L. Extraction and Analysis of Extracellular Polymeric Substances: Comparison of Methods and Extracellular Polymeric Substance Levels in Salmonella Pullorum SA 1685. Environ. Eng. Sci. 2009, 26, 1523–1532. [Google Scholar] [CrossRef] [Green Version]
- Seshadri, S.; RituKhurana, R.; Fink, A.L. Fourier Transform Infrared spectroscopy of Protein Deposits. Methods Enzymol. 1999, 309, 559–576. [Google Scholar]
- Garidel, P.; Schott, H. Fourier-Transform Mid-infrared Spectroscopy for Analysis and Screening of Liquid Protein Formulations Part 2: Details Analysis and Applications. Bioprocess Int. 2006, 1, 48–55. [Google Scholar]
- Kong, J.; Yu, S. Fourier Transform Infrared Spectroscopic Analysis of Protein Secondary Structures. Acta Biochim. Biophys. Sin. 2007, 39, 549–559. [Google Scholar] [CrossRef] [Green Version]
- Walther, F.J.; Waring, A.J.; Hernandez-Juviel, J.M.; Gordon, L.M.; Wang, Z.; Jung, C.-L.; Ruchala, P.; Clark, A.P.; Smith, W.M.; Sharma, S.; et al. Critical Structural and Functional Roles for the N-Terminal Insertion Sequence in Surfactant Protein B Analogs. PLoS ONE 2010, 5, e8672. [Google Scholar] [CrossRef]
- Naumann, D. Infrared Spectroscopy in Microbiology. 2000, pp. 102–131. Available online: http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.195.1805&rep=rep1&type=pdf (accessed on 5 November 2020). [CrossRef]
- Pinakoulaki, E.; Varotsis, C. Nitric oxide activation and reduction by heme–copper oxidoreductases and nitric oxide reductase. J. Inorg Biochem. 2008, 102, 1277–1287. [Google Scholar] [CrossRef]
- Pinakoulaki, E.; Varotsis, C. Time-Resolved Resonance Raman and Time-Resolved Step-Scan FTIR Studies of Nitric Oxide Reductase from Paracoccus denitrificans: Comparison of the Heme b3 -FeB Site to That of the Heme-CuB in Oxidases. Biochemistry 2003, 42, 14856–14861. [Google Scholar] [CrossRef]
- Varotsis, C.; Woodruff, W.H.; Babcock, G.T. Time-resolved Raman detection of. mu.(Fe-O) in an early intermediate in the reduction of oxygen by cytochrome oxidase [Erratum to document cited in CA111. J. Am. Chem. Soc. 1990, 112, 1297. [Google Scholar] [CrossRef]
- Koutsoupakis, C.; Pinakoulaki, E.; Stavrakis, S.; Daskalakis, V.; Varotsis, C. Time-resolved step-scan Fourier transform infrared investigation of heme-copper oxidases: Implications for O2 input and H2O/H+ output channels. Biochim. Biophys. Acta Bioenerg. 2004, 1655, 347–352. [Google Scholar] [CrossRef] [Green Version]
- Iwase, T.; Varotsis, C.; Shinzawa-Itoh, K.; Yoshikawa, S.; Kitagawa, T. Infrared evidence for CuB ligation of photodissociated CO of cytochrome c oxidase at ambient temperatures and accompanied deprotonation of a carboxyl side chain of protein T Iwase, C Varotsis, K Shinzawa-Itoh, S Yoshikawa, T Kitagawa. J. Am. Chem. Soc. 1999, 121, 1415–1416. [Google Scholar] [CrossRef]
- Pinakoulaki, E.; Ohta, T.; Soulimane, T.; Kitagawa, T.; Varotsis, C. Simultaneous Resonance Raman Detection of the Heme a3-Fe-CO and CuB-CO Species in CO-bound ba3-Cytochrome c Oxidase from Thermus thermophilus EVIDENCE FOR A CHARGE TRANSFER CuB. J. Biol. Chem. 2004, 279, 22791–22794. [Google Scholar] [CrossRef] [Green Version]
- Koutsoupakis, C.; Soulimane, T.; Varotsis, C. Docking site dynamics of ba3-cytochrome c oxidase from Thermus thermophiles. J. Biol. Chem. 2003, 278, 36806–36809. [Google Scholar] [CrossRef] [Green Version]
- Stavrakis, S.; Pinakoulaki, E.; Urbani, A.; Varotsis, C. Fourier transform infrared evidence for a ferric six-coordinate nitrosylheme b3 complex of cytochrome cbb3 oxidase from Pseudomonas stutzeri at ambient temperature. J. Phys. Chem. B 2002, 106, 12860–12862. [Google Scholar] [CrossRef]
- Varotsis, C.; Vamvouka, M. Resonance Raman and Fourier Transform Infrared Detection of Azide Binding to the Binuclear Center of Cytochrome bo3 Oxidase from Escherichia coli. J. Phys. Chem. B 1999, 103, 3942–3946. [Google Scholar] [CrossRef]
- Babcock, G.T.; Varotsis, C.; Zhang, Y. O2 activation in cytochrome oxidase and in other heme proteins. Biochim. Biophys. Acta Bioenerg. 1992, 1101, 192–194. [Google Scholar] [CrossRef]
- Pinakoulaki, E.; Yoshimura, H.; Yoshioka, S.; Aono, S.; Varotsis, C. Recognition and discrimination of gases by the oxygen-sensing signal transducer protein HemAT as revealed by FTIR spectroscopy. Biochemistry 2006, 45, 7763–7766. [Google Scholar] [CrossRef]
- Ohta, T.; Pinakoulaki, E.; Soulimane, T.; Kitagawa, T.; Varotsis, C. Detection of a Photostable Five-Coordinate Heme a3-Fe−CO Species and Functional Implications of His384/α10 in CO-Bound ba3-Cytochrome c Oxidase from Thermus thermophiles. J. Phys. Chem. B 2004, 108, 5489–5491. [Google Scholar] [CrossRef]
- Papageorgiou, M.; Tselios, C.; Varotsis, C. Photosensitivity responses of Sagittula stellata probed by FTIR, fluorescence and Raman microspectroscopy. RSC Adv. 2019, 9, 27391–27397. [Google Scholar] [CrossRef] [Green Version]
- Tselios, C.; Papageorgiou, M.; Varotsis, C. Extracellular electron uptake from carbon-based π electron surface-donors: Oxidation of graphite sheets by Sulfobacillus thermosulfidooxidans probed by Raman and FTIR spectroscopy. RSC Adv. 2019, 9, 19121–19125. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component Band | Composition % | Assignments |
|---|---|---|
| 1502 | 1 | -CH2 and -CH3 bending modes of lipids and proteins |
| 1518 | 5 | β-sheet structures |
| 1542 | 9 | α-helical |
| 1559 | 1 | Asymmetric stretching -COO- |
| 1584 | 16 | Carboxylate str. of aspartate and glutamate |
| 1629 | 27 | Β-sheet structures |
| 1670 | 33 | Β-turns |
| 1717 | 8 | C=O stretching |
| Week of Bio-Interaction | Component Band | Composition % | Assignments |
|---|---|---|---|
| 1 week | 1501 | 1 | -CH2 and -CH3 bending modes of lipids and proteins |
| 1521 | 2 | β-sheet | |
| 1537 | 1 | β-sheet | |
| 1554 | 10 | α-helical | |
| 1600 | 51 | DNA/RNA components | |
| 1638 | 28 | β-sheet | |
| 1678 | 4 | β-sheet | |
| 1773 | 2 | C=O stretching mode of lipids | |
| 3 weeks | 1533 | 7 | β-sheet |
| 1591 | 45 | Stretching mode of C=C | |
| 1635 | 33 | β-sheet | |
| 1671 | 13 | β-turn | |
| 1720 | 1 | Free EPS | |
| 6 weeks | 1520 | 10 | β-sheet |
| 1576 | 34 | Carboxylate str. of aspartate/ glutamate | |
| 1632 | 42 | β-sheet | |
| 1687 | 9 | Antiparallel β-sheet | |
| 1729 | 4 | Free EPS | |
| 7 weeks | 1495 | 5 | Side chain vibrations |
| 1523 | 12 | β-sheet structures | |
| 1571 | 29 | Carboxylate str. of aspartate and glutamate | |
| 1630 | 38 | β-sheet structures | |
| 1675 | 3 | β-turns | |
| 1702 | 9 | Antiparallel β-sheet structures | |
| 1746 | 4 | Bound EPS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varotsis, C.; Papageorgiou, M.; Tselios, C.; Yiannakkos, K.A.; Adamou, A.; Nicolaides, A. Bacterial Colonization on the Surface of Copper Sulfide Minerals Probed by Fourier Transform Infrared Micro-Spectroscopy. Crystals 2020, 10, 1002. https://0-doi-org.brum.beds.ac.uk/10.3390/cryst10111002
Varotsis C, Papageorgiou M, Tselios C, Yiannakkos KA, Adamou A, Nicolaides A. Bacterial Colonization on the Surface of Copper Sulfide Minerals Probed by Fourier Transform Infrared Micro-Spectroscopy. Crystals. 2020; 10(11):1002. https://0-doi-org.brum.beds.ac.uk/10.3390/cryst10111002
Chicago/Turabian StyleVarotsis, Constantinos, Marios Papageorgiou, Charalampos Tselios, Konstantinos A. Yiannakkos, Anastasia Adamou, and Antonis Nicolaides. 2020. "Bacterial Colonization on the Surface of Copper Sulfide Minerals Probed by Fourier Transform Infrared Micro-Spectroscopy" Crystals 10, no. 11: 1002. https://0-doi-org.brum.beds.ac.uk/10.3390/cryst10111002