Elucidating Tricin-Lignin Structures: Assigning Correlations in HSQC Spectra of Monocot Lignins

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Syntheses of Model Compounds
2.3. Acetylation of Model Compounds and Lignin
2.4. Nuclear Magnetic Resonance (NMR) Characterization
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Boerjan, W.; Ralph, J.; Baucher, M. Lignin biosynthesis. Annu. Rev. Plant Biol. 2003, 54, 519–546. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Tobimatsu, Y.; Havkin-Frenkel, D.; Dixon, R.A.; Ralph, J. A polymer of caffeyl alcohol in plant seeds. Proc. Natl. Acad. Sci. USA 2012, 109, 1772–1777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kukkola, E.M.; Koutaniemi, S.; Pollanen, E.; Gustafsson, M.; Karhunen, P.; Lundell, T.K.; Saranpaa, P.; Kilpelainen, I.; Teeri, T.H.; Fagerstedt, K.V. The dibenzodioxocin lignin substructure is abundant in the inner part of the secondary wall in Norway spruce and silver birch xylem. Planta 2004, 218, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Bottcher, A.; Cesarino, I.; dos Santos, A.B.; Vicentini, R.; Mayer, J.L.S.; Vanholme, R.; Morreel, K.; Goeminne, G.; Moura, J.C.M.S.; Nobile, P.M.; et al. Lignification in sugarcane: Biochemical characterization, gene discovery, and expression analysis in two genotypes contrasting for lignin content. Plant Physiol. 2013, 163, 1539–1557. [Google Scholar] [CrossRef] [PubMed]
- Del Río, J.C.; Rencoret, J.; Prinsen, P.; Martinez, A.T.; Ralph, J.; Gutierrez, A. Structural characterization of wheat straw lignin as revealed by analytical pyrolysis, 2D-NMR, and reductive cleavage methods. J. Agric. Food Chem. 2012, 60, 5922–5935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, W.; Lu, F.C.; Regner, M.; Zhu, Y.M.; Rencoret, J.; Ralph, S.A.; Zakai, U.I.; Morreel, K.; Boerjan, W.; Ralph, J. Tricin, a flavonoid monomer in monocot lignification. Plant Physiol. 2015, 167, 1284–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, W.; Morreel, K.; Lu, F.; Rencoret, J.; del Río, J.C.; Voorend, W.; Vermerris, W.; Boerjan, W.; Ralph, J. Maize tricin-oligolignol metabolites and their implications for monocot lignification. Plant Physiol. 2016, 171, 810–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, W.; Rencoret, J.; Lu, F.C.; Karlen, S.D.; Smith, B.G.; Harris, P.J.; del Río, J.C.; Ralph, J. Tricin-lignins: Occurrence and quantitation of tricin in relation to phylogeny. Plant J. 2016, 88, 1046–1057. [Google Scholar] [CrossRef] [PubMed]
- Eloy, N.B.; Voorend, W.; Lan, W.; Saleme, M.D.S.; Cesarino, I.; Vanholme, R.; Smith, R.A.; Goeminne, G.; Pallidis, A.; Morreel, K.; et al. Silencing CHALCONE SYNTHASE in maize impedes the incorporation of tricin into lignin and increases lignin content. Plant Physiol. 2017, 173, 998–1016. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.J.; Helms, G.L.; Gao, X.; Chen, S.L. Quantification of wheat straw lignin structure by comprehensive NMR analysis. J. Agric. Food Chem. 2013, 61, 10848–10857. [Google Scholar] [CrossRef] [PubMed]
- Zikeli, F.; Ters, T.; Fackler, K.; Srebotnik, E.; Li, J.B. Successive and quantitative fractionation and extensive structural characterization of lignin from wheat straw. Ind. Crop. Prod. 2014, 61, 249–257. [Google Scholar] [CrossRef]
- Rencoret, J.; Prinsen, P.; Gutierrez, A.; Martinez, A.T.; del Río, J.C. Isolation and structural characterization of the milled wood lignin, dioxane lignin, and cellulolytic lignin preparations from brewer’s spent grain. J. Agric. Food Chem. 2015, 63, 603–613. [Google Scholar] [CrossRef] [PubMed]
- Del Río, J.C.; Lino, A.G.; Colodette, J.L.; Lima, C.F.; Gutierrez, A.; Martinez, A.T.; Lu, F.C.; Ralph, J.; Rencoret, J. Differences in the chemical structure of the lignins from sugarcane bagasse and straw. Biomass Bioenergy 2015, 81, 322–338. [Google Scholar] [CrossRef] [Green Version]
- Del Río, J.C.; Prinsen, P.; Rencoret, J.; Nieto, L.; Jimenez-Barbero, J.; Ralph, J.; Martinez, A.T.; Gutierrez, A. Structural characterization of the lignin in the cortex and pith of elephant grass (Pennisetum purpureum) stems. J. Agric. Food Chem. 2012, 60, 3619–3634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rencoret, J.; Ralph, J.; Marques, G.; Gutierrez, A.; Martinez, A.T.; del Río, J.C. Structural characterization of lignin isolated from coconut (Cocos nucifera) coir fibers. J. Agric. Food Chem. 2013, 61, 2434–2445. [Google Scholar] [CrossRef] [PubMed] [Green Version]



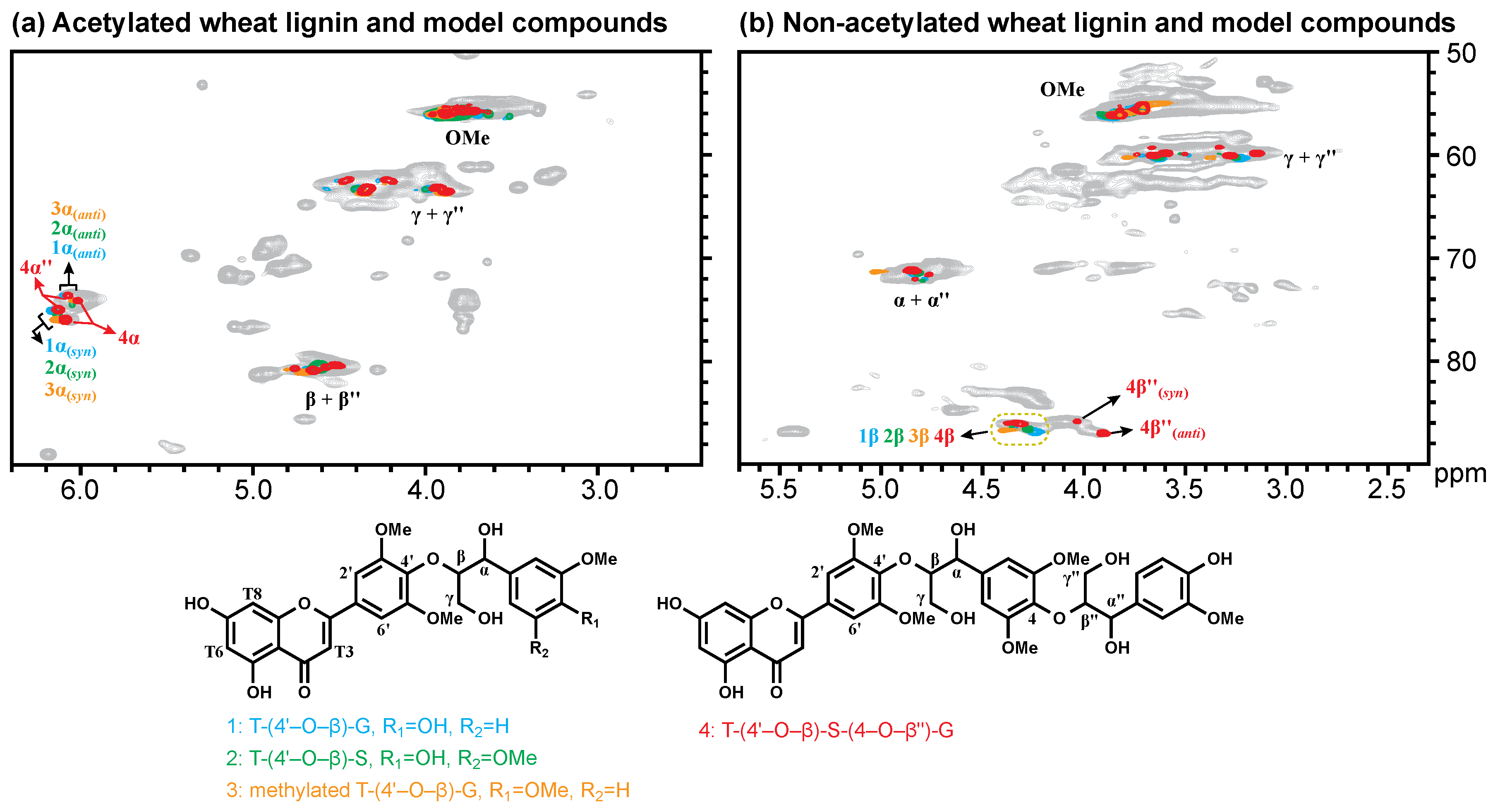{kind=link}
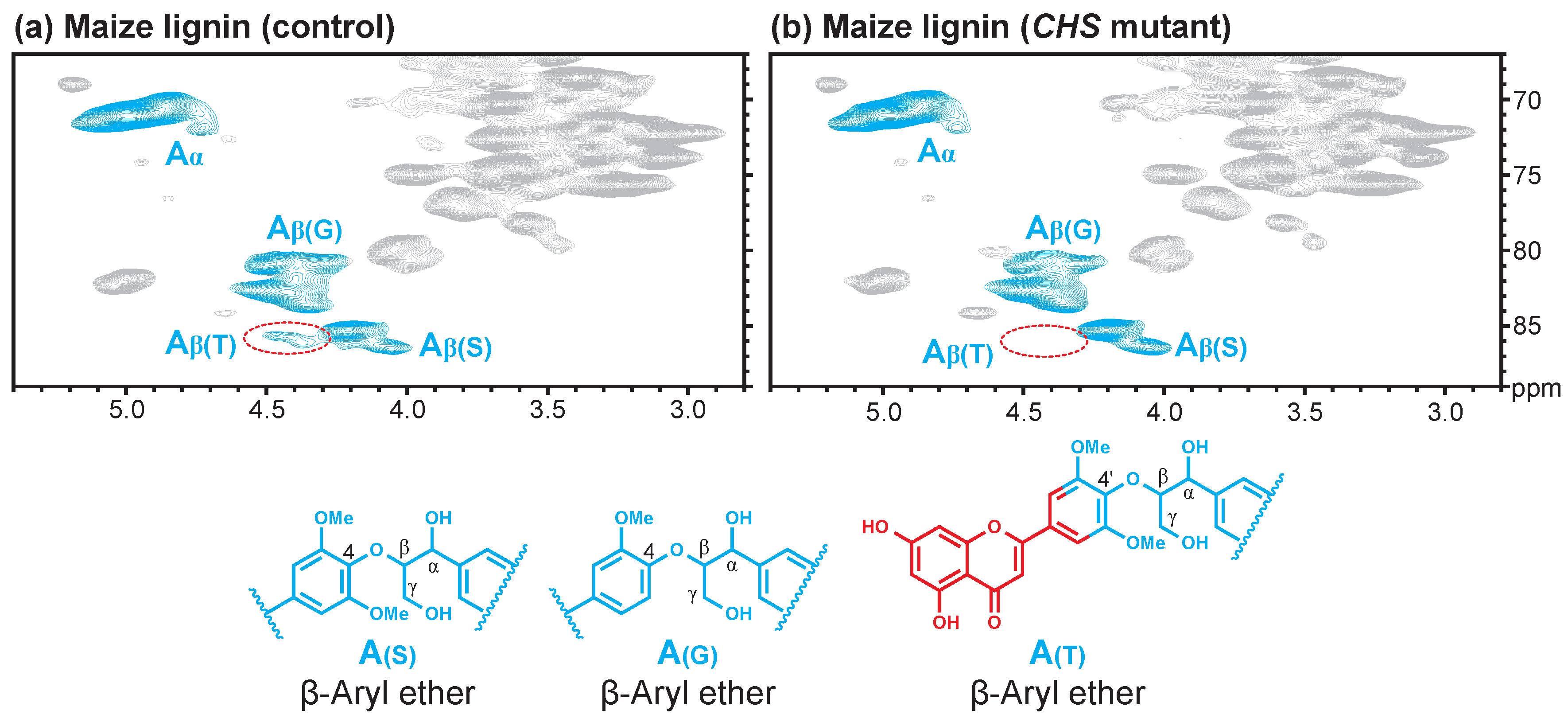{kind=link}
| Title 1 | Cβ/Hβ | Cβ″/Hβ″ |
|---|---|---|
| 1. T-(4′–O–β)-G | 86.94/4.24 | - |
| 86.38/4.34 | ||
| 2. T-(4′–O–β)-S | 86.63/4.27 | - |
| 86.36/4.36 | ||
| 3. 4–O-methylated T-(4′–O–β)-G | 86.74/4.39 | - |
| 86.21/4.49 | ||
| 4. T-(4′–O–β)-S-(4–O–β″)-G | 86.11/4.31 | 87.01/3.91 |
| 86.04/4.38 | 85.85/4.03 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lan, W.; Yue, F.; Rencoret, J.; Del Río, J.C.; Boerjan, W.; Lu, F.; Ralph, J. Elucidating Tricin-Lignin Structures: Assigning Correlations in HSQC Spectra of Monocot Lignins. Polymers 2018, 10, 916. https://0-doi-org.brum.beds.ac.uk/10.3390/polym10080916
Lan W, Yue F, Rencoret J, Del Río JC, Boerjan W, Lu F, Ralph J. Elucidating Tricin-Lignin Structures: Assigning Correlations in HSQC Spectra of Monocot Lignins. Polymers. 2018; 10(8):916. https://0-doi-org.brum.beds.ac.uk/10.3390/polym10080916
Chicago/Turabian StyleLan, Wu, Fengxia Yue, Jorge Rencoret, José Carlos Del Río, Wout Boerjan, Fachuang Lu, and John Ralph. 2018. "Elucidating Tricin-Lignin Structures: Assigning Correlations in HSQC Spectra of Monocot Lignins" Polymers 10, no. 8: 916. https://0-doi-org.brum.beds.ac.uk/10.3390/polym10080916