Equal K Amounts to N Achieved Optimal Biomass and Better Fiber Quality of Late Sown Cotton in Yangtze River Valley

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Cultivar
2.2. Climate
2.3. Experiment Design
2.4. Field Management
2.5. Data Collection
2.5.1. Cotton Phenology
2.5.2. Cotton Biomass Accumulation
2.5.3. Cotton Fiber Quality
2.6. Statistical Analysis
3. Results
3.1. Cotton Plant Phenology
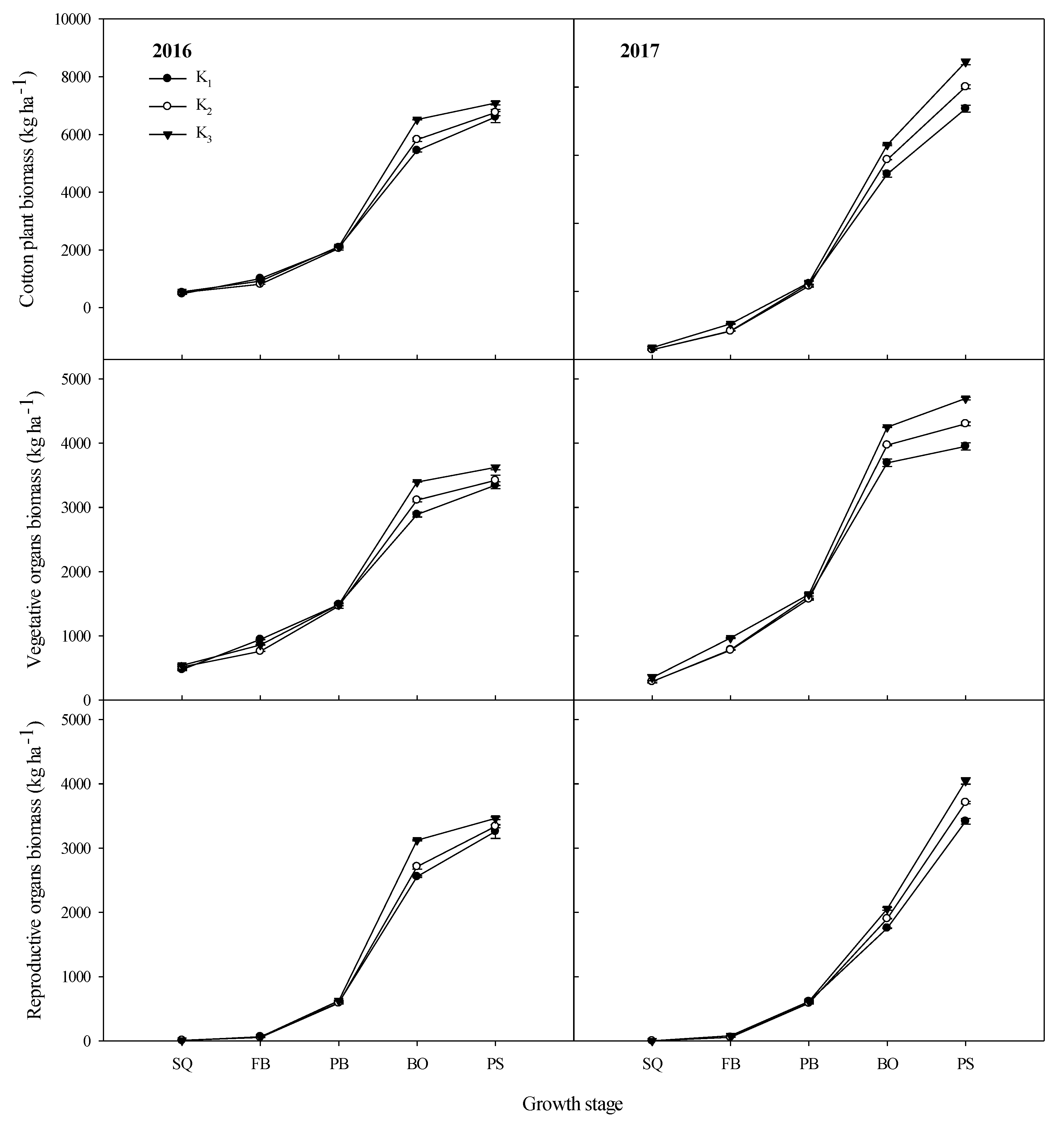
3.2. Cotton Plant Biomass Accumulation
3.3. Simulation of Biomass Accumulation
3.4. Fiber Quality
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Reddy, N.; Yang, Y. Properties and potential applications of natural cellulose fibers from the bark of cotton stalks. Bioresour. Technol. 2009, 100, 3563–3569. [Google Scholar] [CrossRef]
- Esteve-Turrillas, F.A.; Guardia, M.D.L. Environmental impact of recover cotton in textile industry. Resour. Conserv. Recycl. 2017, 116, 107–115. [Google Scholar] [CrossRef]
- Yang, G.; Tang, H.; Nie, Y.; Zhang, X. Responses of cotton growth, yield, and biomass to nitrogen split application ratio. Eur. J. Agron. 2011, 35, 164–170. [Google Scholar] [CrossRef]
- Yang, G.; Luo, X.; Nie, Y.; Zhang, X. Effects of plant density on yield and canopy micro environment in hybrid cotton. J. Integr. Agric. 2014, 13, 2154–2163. [Google Scholar] [CrossRef]
- Yang, G.; Zhou, M.Y. Multi-location investigation of optimum planting density and boll distribution of high-yielding cotton (Gossypium hirsutum L.) in Hubei province, China. Agric. Sci. China 2010, 9, 1749–1757. [Google Scholar] [CrossRef]
- Khan, A.; Najeeb, U.; Wang, L.; Tan, D.K.Y.; Yang, G.; Munsif, F.; Ali, S.; Hafeez, A. Planting density and sowing date strongly influence growth and lint yield of cotton crops. Field Crops Res. 2017, 209, 129–135. [Google Scholar] [CrossRef]
- Shah, A.N.; Yang, G.; Tanveer, M.; Iqbal, J. Leaf gas exchange, source-sink relationship, and growth response of cotton to the interactive effects of nitrogen rate and planting density. Acta Physiol. Plant. 2017, 39, 119. [Google Scholar] [CrossRef]
- Yang, G.; Tang, H.; Tong, J.; Nie, Y.; Zhang, X. Effect of fertilization frequency on cotton yield and biomass accumulation. Field Crops Res. 2012, 125, 161–166. [Google Scholar] [CrossRef]
- Hafeez, A.; Ali, S.; Ma, X.; Tung, S.A.; Shah, A.N.; Liu, A.; Zhang, Z.; Liu, J.; Yang, G. Sucrose metabolism in cotton subtending leaves influencedby potassium-to-nitrogen ratios. Nutr. Cycl. Agroecosyst. 2019, 113, 201–216. [Google Scholar] [CrossRef]
- Reddy, K.R.; Zhao, D. Interactive effects of elevated CO2 and potassium deficiency on photosynthesis, growth, and biomass partitioning of cotton. Field Crops Res. 2005, 94, 201–213. [Google Scholar] [CrossRef]
- Clementbailey, J.; Gwathmey, C.O. Potassium effects on partitioning, yield, and earliness of contrasting cotton cultivars. Agron. J. 2007, 99, 1130–1136. [Google Scholar] [CrossRef]
- Gerardeaux, E.; Jordan-Meille, L. Effect of carbon assimilation on dry weight production and partitioning during vegetative growth. Plant Soil. 2009, 324, 329–343. [Google Scholar] [CrossRef]
- Hu, W.; Yang, J.; Meng, Y.; Wang, Y.; Chen, B.; Zhao, W.; Oosterhuis, D.M.; Zhou, Z. Potassium application affects carbohydrate metabolism in the leaf subtending the cotton (Gossypium hirsutum L.) boll and its relationship with boll biomass. Field Crops Res. 2015, 179, 120–131. [Google Scholar] [CrossRef]
- Hu, W.; Zhao, W.; Yang, J.; Oosterhuis, D.M.; Loka, D.A.; Zhou, Z. Relationship between potassium fertilization and nitrogen metabolism in the leaf subtending the cotton (Gossypium hirsutum L.) boll during the boll development stage. Plant Physiol. Biochem. 2016, 101, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Gormus, O. Effects of rate and time of potassium application on cotton yield and quality in Turkey. J. Agron. Crop Sci. 2002, 188, 382–388. [Google Scholar] [CrossRef]
- Hu, W.; Coomer, T.D.; Loka, D.A.; Oosterhuis, D.M.; Zhou, Z. Potassium deficiency affects the carbon-nitrogen balance in cotton leaves. Plant Physiol. Biochem. 2017, 115, 408–417. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Dai, Z.; Yang, J.; Snider, J.L.; Wang, S.; Meng, Y.; Wang, Y.; Chen, B.; Zhao, W.; Zhou, Z. Cultivar sensitivity of cotton seed yield to potassium availability is associated with differences in carbohydrate metabolism in the developing embryo. Field Crops Res. 2017, 214, 301–309. [Google Scholar] [CrossRef]
- Gerardeaux, E.; Jordan-Meille, L.; Constantin, J.; Pellerin, S.; Dingkuhn, M. Changes in plant morphology and dry matter partitioning caused by potassium deficiency in Gossypium hirsutum (L.). Environ. Exp. Bot. 2010, 67, 451–459. [Google Scholar] [CrossRef]
- Onanuga, A.O.; Jiang, P.; Sina, A. Residual level of phosphorus and potassium nutrients in hydroponically grown cotton (Gossypium hirsutum L.). J. Agric. Sci. 2012, 4, 149–160. [Google Scholar] [CrossRef]
- Oosterhuis, D.M.; Loka, D.A.; Raper, T.B. Potassium and stress alleviation: Physiological functions and management of cotton. J. Plant Nutr. Soil Sci. 2013, 176, 331–343. [Google Scholar] [CrossRef]
- Tsialtas, I.T.; Shabala, S.; Baxevanos, D.; Matsi, T. Effect of potassium fertilization on leaf physiology, fiber yield and quality in cotton (Gossypium hirsutum L.) under irrigated mediterranean conditions. Field Crops Res. 2016, 193, 94–103. [Google Scholar] [CrossRef]
- Chen, Y.; Li, Y.; Hu, D.; Zhang, X.; Wen, Y.; Chen, D. Spatial distribution of potassium uptake across the cotton plant affects fiber length. Field Crops Res. 2016, 192, 126–133. [Google Scholar] [CrossRef]
- Lokhande, S.; Reddy, K.R. Reproductive performance and fiber quality responses of cotton to potassium nutrition. Am. J. Plant Sci. 2015, 6, 911–924. [Google Scholar] [CrossRef] [Green Version]
- Gormus, O.; Yucel, C. Different planting date and potassium fertility effects on cotton yield and fiber properties in the Çukurova region, Turkey. Field Crops Res. 2002, 78, 141–149. [Google Scholar] [CrossRef]
- Tariq, M.; Afzal, M.N.; Muhammad, D.; Ahmad, S.; Shahzad, A.N.; Kiran, A.; Wakeel, A. Relationship of tissue potassium content with yield and fiber quality components of Bt cotton as influenced by potassium application methods. Field Crops Res. 2008, 229, 37–43. [Google Scholar] [CrossRef]
- Ali, S.; Hafeez, A.; Ma, X.; Tung, S.A.; Chattha, M.S.; Shah, A.N.; Luo, D.; Ahmad, S.; Liu, J.; Yang, G. Equal potassium-nitrogen ratio regulated the nitrogen metabolism and yield of high-density late-planted cotton (Gossypium hirsutum L.) in Yangtze River valley of China. Ind. Crops Prod. 2019, 129, 231–241. [Google Scholar] [CrossRef]
- Hafeez, A.; Ali, S.; Ma, X.; Tung, S.A.; Shah, A.N.; Liu, A.; Ahmed, S.; Chattha, M.S.; Yang, G. Potassium to nitrogen ratio favors photosynthesis in late-planted cotton at high planting density. Ind. Crops Prod. 2018, 124, 369–381. [Google Scholar] [CrossRef]
- Ali, S.; Hafeez, A.; Ma, X.; Tung, S.A.; Liu, A.; Shah, A.N.; Chattha, M.S.; Zhang, Z.; Yang, G. Potassium relative ratio to nitrogen considerably favors carbon metabolism in late-planted cotton at high planting density. Field Crops Res. 2018, 223, 48–56. [Google Scholar] [CrossRef]
- Wright, P.R. Premature senescence of cotton (Gossypium hirsutum L.)-Predominantly a potassium disorder caused by an imbalance of source and sink. Plant Soil. 1999, 211, 231–239. [Google Scholar] [CrossRef]
- Bedrossian, S.; Singh, B. Potassium adsorption characteristics and potassium forms in some New South Wales soils in relation to early senescence in cotton. Aust. J. Soil Res. 2004, 42, 747–753. [Google Scholar] [CrossRef]
- Li, B.; Wang, Y.; Zhang, Z.; Wang, B.; Eneji, A.E.; Duan, L.; Li, Z.; Tian, X. Cotton shoot plays a major role in mediating senescence induced by potassium deficiency. J. Plant Physiol. 2012, 169, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Lv, X.; Yang, J.; Chen, B.; Zhao, W.; Meng, Y.; Wang, Y.; Zhou, Z. Effects of potassium deficiency on antioxidant metabolism related to leaf senescence in cotton (Gossypium hirsutum L.). Field Crops Res. 2016, 191, 139–149. [Google Scholar] [CrossRef]
- Read, J.J.; Reddy, K.R.; Jenkins, J.N. Yield and fiber quality of upland cotton as influenced by nitrogen and potassium nutrition. Eur. J. Agron. 2006, 24, 282–290. [Google Scholar] [CrossRef]
- Zahoor, R.; Dong, H.; Abid, M.; Zhao, W.; Wang, Y.; Zhou, Z. Potassium fertilizer improves drought stress alleviation potential in cotton by enhancing photosynthesis and carbohydrate metabolism. Environ. Exp. Bot. 2017, 137, 73–83. [Google Scholar] [CrossRef]
- Makhdum, M.I.; Pervez, H.; Ashraf, M. Dry matter accumulation and partitioning in cotton (Gossypium hirsutum L.) as influenced by potassium fertilization. Biol. Fertil. Soils. 2007, 43, 295–301. [Google Scholar] [CrossRef]
- Tang, F.; Wang, T.; Zhu, J. Carbohydrate profiles during cotton (Gossypium hirsutum L.) boll development and their relationships to boll characters. Field Crops Res. 2014, 164, 98–106. [Google Scholar] [CrossRef]
- Pettigrew, W.T.; Meredithjr, W.R. Dry matter production, nutrient uptake, and growth of cotton as affected by potassium fertilization. J. Plant Nutr. 1997, 20, 531–548. [Google Scholar] [CrossRef]
- Tung, S.A.; Huang, Y.; Hafeez, A.; Ali, S.; Khan, A.; Souliyanonh, B.; Song, X.; Liu, A.; Yang, G. Mepiquat chloride effects on cotton yield and biomass accumulation under late sowing and high density. Field Crops Res. 2018, 215, 59–65. [Google Scholar] [CrossRef]


{kind=link}
| Year | Treatment | Growing Stage (m/d) # | Growth Period (d) # | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Emergence | Squaring | First Bloom * | Opening | Seedling | Squaring | Flowering and Boll Setting | Total | ||
| 2016 | K1 | 5/28 | 7/15 | 8/1 | 9/23 | 48a * | 17a | 53a | 118a |
| K2 | 5/28 | 7/15 | 8/1 | 9/22 | 48a | 17a | 52a | 117a | |
| K3 | 5/28 | 7/15 | 8/1 | 9/22 | 48a | 17a | 52a | 117a | |
| 2017 | K1 | 5/18 | 6/20 | 7/15 | 8/24 | 33a | 25a | 40a | 98a |
| K2 | 5/18 | 6/20 | 7/15 | 8/25 | 33a | 25a | 41a | 99a | |
| K3 | 5/18 | 6/20 | 7/15 | 8/25 | 33a | 25a | 41a | 99a | |
| Year | Treatment | Biomass Accumulation (kg ha−1) | RPB/CPB (%) | ||||
|---|---|---|---|---|---|---|---|
| Root | Stem | Leaves | Reproductive Parts | Total | |||
| 2016 | K1 | 815.6c * | 1950.1b | 406.0b | 3258.8a | 6603.9b | 48.35a |
| K2 | 909.8b | 1991.3b | 494.6a | 3338.6a | 6759.6b | 49.39a | |
| K3 | 1017.1a | 2206.3a | 517.1a | 3464.0a | 7086.8a | 48.86a | |
| Average | 914.2 | 2049.2 | 499.6 | 3353.8 | 6816.8 | 48.87 | |
| 2017 | K1 | 1169.1a | 2374.0c | 406.0b | 3417.1b | 7366.2c | 44.18b |
| K2 | 1103.1a | 2703.3b | 494.6a | 3709.1ab | 8010.1b | 46.32a | |
| K3 | 1176.7a | 3000.8a | 517.1a | 4044.6a | 8739.2a | 46.29a | |
| Average | 1149.6 | 2692.7 | 472.6 | 3723.6 | 8038.5 | 45.60 | |
| Year | Treatment | Regression Eqs. W = kg/ha, t = DAE | p-Value | Fast Accumulation Period | ||||
|---|---|---|---|---|---|---|---|---|
| t1 (DAE) | t2 (DAE) | Δt | VT (kg/ha d−1) | VM (kg/ha d−1) | ||||
| Cotton plant | ||||||||
| 2016 | K1 | W=6771.764/(1+4.98e−0.050t) | 0.0006 | 72.7 | 125.0 | 52.3 | 74.7 | 85.2 |
| K2 | W=6850.748/(1+5.58e−0.057t) | 0.0007 | 74.1 | 120.0 | 45.8 | 86.3 | 98.4 | |
| K3 | W=7170.924/(1+6.19e−0.066t) | 0.0004 | 74.0 | 114.0 | 40.0 | 103.5 | 118.0 | |
| Average | 73.6 | 119.7 | 46.0 | 88.2 | 100.6 | |||
| 2017 | K1 | W=7387.145/(1+6.18e−0.073t) | 0.0003 | 66.7 | 102.9 | 36.1 | 118.0 | 134.6 |
| K2 | W=8036.283/(1+6.54e−0.076t) | 0.0003 | 68.7 | 103.3 | 34.6 | 134.0 | 152.8 | |
| K3 | W=8785.985/(1+6.33e−0.073t) | 0.0005 | 68.6 | 104.6 | 36.0 | 140.8 | 160.6 | |
| Average | 68.0 | 103.6 | 35.6 | 130.9 | 149.3 | |||
| Vegetative parts | ||||||||
| 2016 | K1 | W=3399.527/(1+4.00e−0.046t) | 0.0008 | 58.7 | 116.4 | 57.7 | 34.0 | 38.8 |
| K2 | W=3477.372/(1+4.61e−0.053t) | 0.0010 | 62.6 | 112.7 | 50.0 | 40.1 | 45.8 | |
| K3 | W=3714.961/(1+4.71e−0.054t) | 0.0010 | 63.2 | 112.2 | 49.0 | 43.8 | 49.9 | |
| Average | 61.5 | 113.7 | 52.3 | 39.3 | 44.8 | |||
| 2017 | K1 | W=4036.745/(1+6.64e−0.087t) | 0.0041 | 61.0 | 91.2 | 30.2 | 77.2 | 88.1 |
| K2 | W=4390.263/(1+7.01e−0.090t) | 0.0044 | 63.2 | 92.4 | 29.2 | 86.7 | 98.9 | |
| K3 | W=4801.295/(1+6.43e−0.082t) | 0.0066 | 62.3 | 94.3 | 32.1 | 86.5 | 98.6 | |
| Average | 62.1 | 92.6 | 30.5 | 83.5 | 95.2 | |||
| Reproductive parts | ||||||||
| 2016 | K1 | W=3315.932/(1+6.97e−0.064t) | 0.0021 | 88.0 | 129.0 | 41.0 | 46.7 | 53.3 |
| K2 | W=3374.119/(1+7.34e−0.069t) | 0.0017 | 87.7 | 126.0 | 38.4 | 50.8 | 57.9 | |
| K3 | W=3450.948/(1+8.52e−0.085t) | 0.0009 | 84.8 | 115.8 | 31.0 | 64.3 | 73.3 | |
| Average | 86.8 | 123.6 | 36.8 | 53.9 | 61.5 | |||
| 2017 | K1 | W=3445.530/(1+6.85e−0.070t) | 0.0020 | 79.3 | 117.0 | 37.7 | 52.7 | 60.1 |
| K2 | W=3733.200/(1+7.25e−0.074t) | 0.0012 | 80.4 | 116.1 | 35.7 | 60.4 | 68.9 | |
| K3 | W=4072.099/(1+7.27e−0.074t) | 0.0009 | 80.7 | 116.4 | 35.7 | 65.9 | 75.1 | |
| Average | 80.1 | 116.5 | 36.4 | 59.7 | 68.0 | |||
| Year | Treatment | Length (mm) | Uniformity (%) | Micronaire | Strength (g/tex) | Elongation (%) |
|---|---|---|---|---|---|---|
| 2016 | K1 | 23.3b * | 83.8a | 4.6a | 25.8b | 6.57a |
| K2 | 25.0ab | 83.8a | 4.8a | 27.6ab | 6.60a | |
| K3 | 25.4a | 84.1a | 4.5a | 28.7a | 6.60a | |
| Average | 24.6 | 83.9 | 4.6 | 27.4 | 6.6 | |
| 2017 | K1 | 22.2b | 84.6a | 5.2a | 24.6b | 6.57a |
| K2 | 23.1a | 85.1a | 4.4b | 26.0a | 6.60a | |
| K3 | 23.6a | 84.9a | 4.3b | 26.4a | 6.60a | |
| Average | 23.0 | 84.9 | 4.6 | 25.7 | 6.6 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, X.; Ali, S.; Hafeez, A.; Liu, A.; Liu, J.; Zhang, Z.; Luo, D.; Noor Shah, A.; Yang, G. Equal K Amounts to N Achieved Optimal Biomass and Better Fiber Quality of Late Sown Cotton in Yangtze River Valley. Agronomy 2020, 10, 112. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10010112
Ma X, Ali S, Hafeez A, Liu A, Liu J, Zhang Z, Luo D, Noor Shah A, Yang G. Equal K Amounts to N Achieved Optimal Biomass and Better Fiber Quality of Late Sown Cotton in Yangtze River Valley. Agronomy. 2020; 10(1):112. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10010112
Chicago/Turabian StyleMa, Xiaolei, Saif Ali, Abdul Hafeez, Anda Liu, Jiahao Liu, Zhao Zhang, Dan Luo, Adnan Noor Shah, and Guozheng Yang. 2020. "Equal K Amounts to N Achieved Optimal Biomass and Better Fiber Quality of Late Sown Cotton in Yangtze River Valley" Agronomy 10, no. 1: 112. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10010112