3.2.1. CBDA and CBD Content
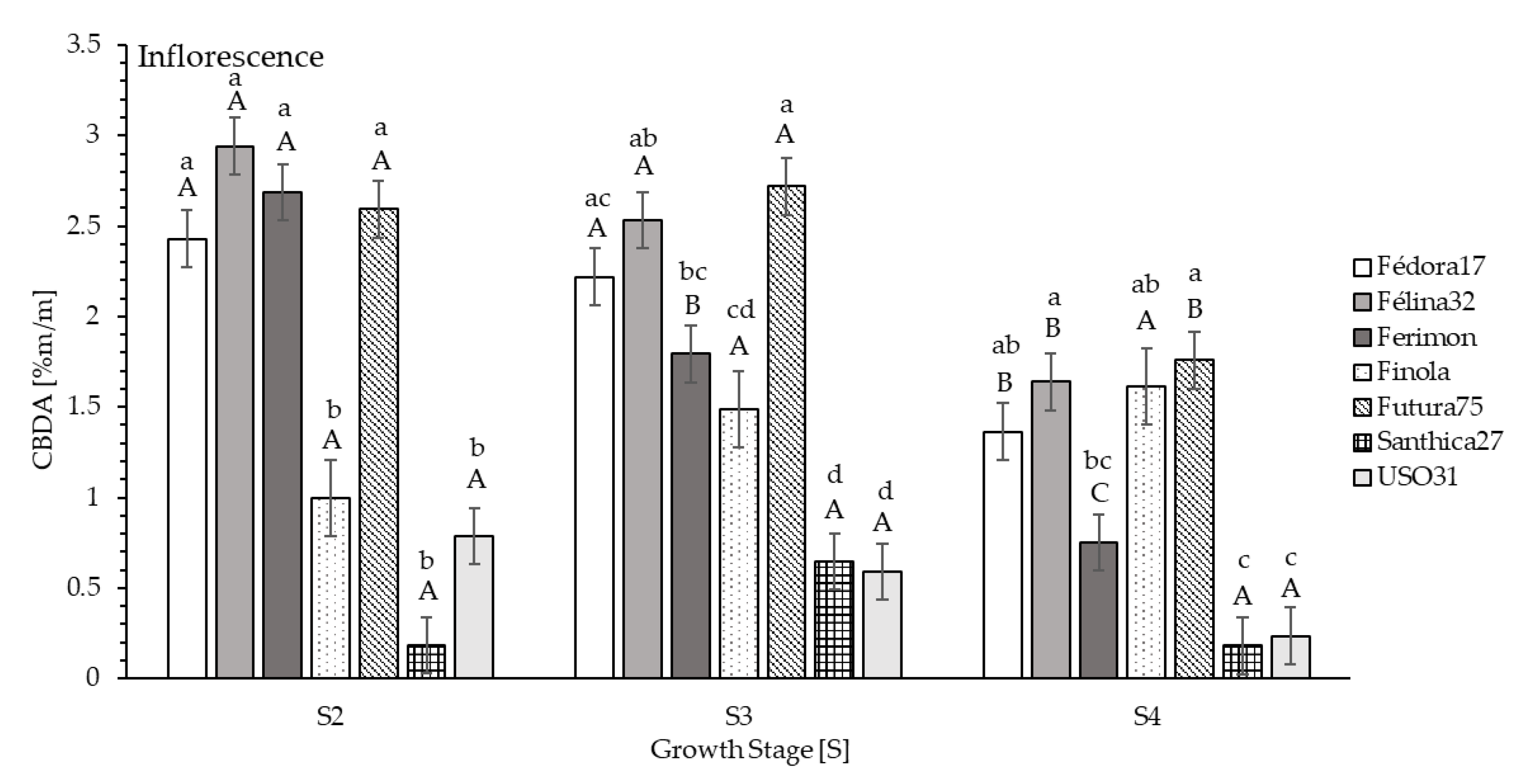
The highest content of CBDA in inflorescence at S2 was measured in genotype Félina32 (2.941%), followed by Ferimon (2.687%), Futura75 (2.593%) and Fédora17 (2.430%). The lowest content was found at S2 in dried inflorescence of Finola (0.995%), USO31 (0.785%) and Santhica27 (0.185%;
Figure 3). At full-flowering stage (S3), average CBDA contents differed significantly between genotypes. The highest contents were measured in Futura75 (2.719%), Félina32 (2.533%), followed by Fédora17 (2.220%) and Ferimon (1.793%;
Figure 3). The present results are in line with findings of Sikora et al. [
31], indicating a range of 1.444% to 2.039% cannabidiol in the upper third of the plant, including leaves and inflorescence of genotype Fédora19, Ferimon12 and Futura77. Santhica27 and USO31 exhibited the lowest values with 0.647% and 0.589%, respectively (
Figure 3). At seed maturity (S4), genotype Futura75 (1.759%), Félina32 (1.639%) and Fédora17 (1.363%), as well as Finola (1.613%) indicated the highest CBDA values in their inflorescence (
Figure 3).
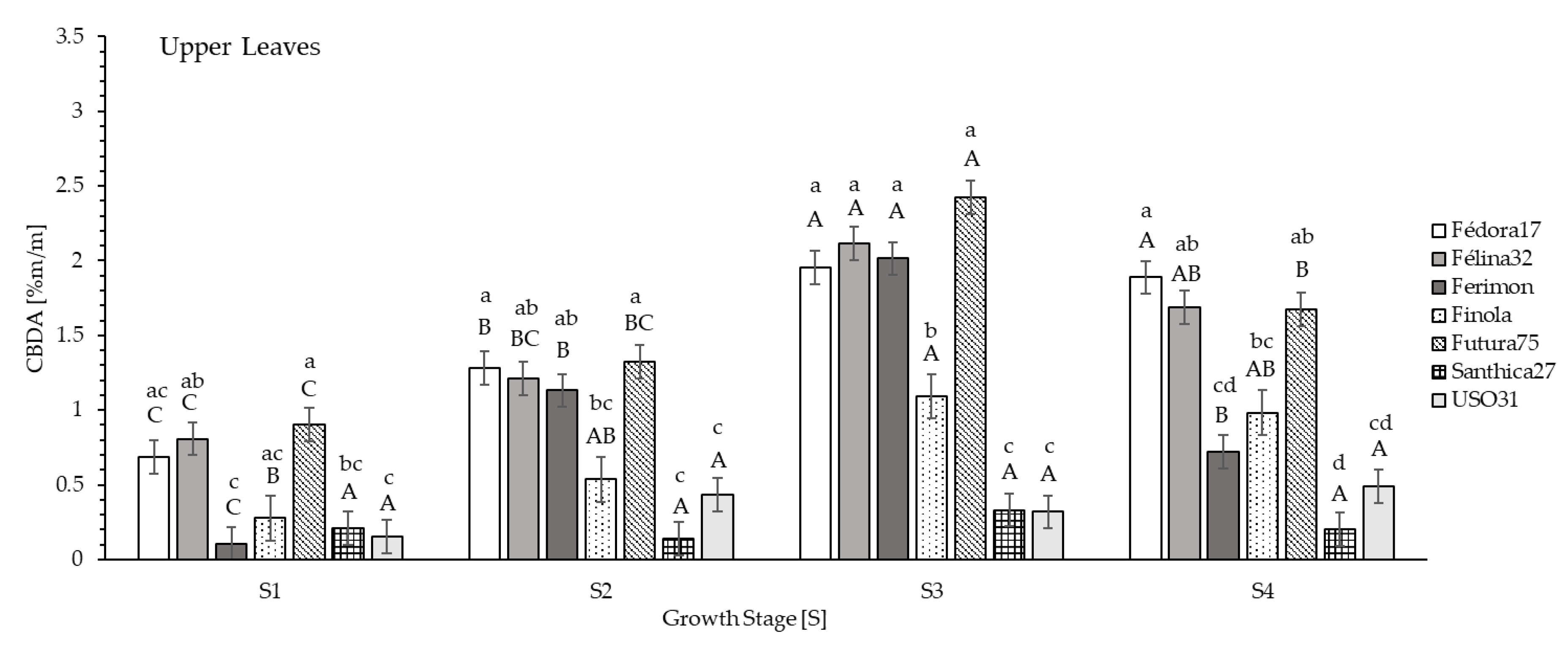
Average CBDA contents of dried upper leaves, in S1 ranged from 0.104% (Ferimon) to 0.902% (Futura75;
Figure 4). At S2 values ranged from 0.139% (Santhica27) to 1.322% (Futura75;
Figure 4). The highest CBDA content of the dried upper leaf fraction was obtained at S3. Genotype Futura75 (2.422%) indicated the highest contents, followed by Félina32 (2.116%), Ferimon (2.014%) and Fédora17 (1.954%;
Figure 4). At seed maturity (S4), CBDA contents of dried upper leaves decreased and ranged from 0.203% (Santhica27) to 1.888% (Fédora17;
Figure 4).
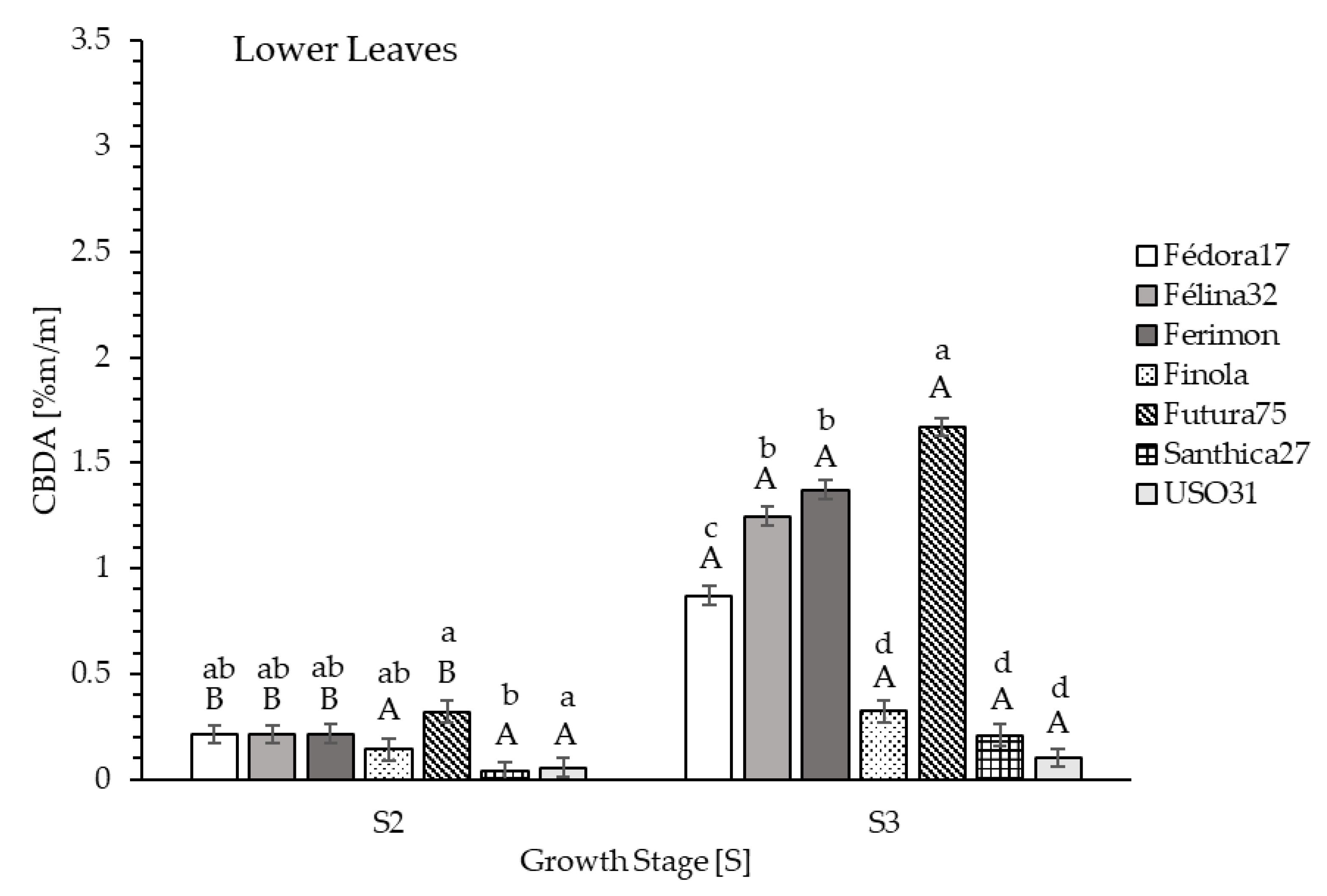
Generally, the CBDA content of the lower leaves was lowest in both growth stages. S2 ranged from 0.041% (Santhica27) to 0.319% (Futura75) and S3 ranged from 0.104% (USO31) to 1.670% (Futura75). The highest content of the lower leaves over all genotypes was measured at S3. Genotype Futura75 indicated the highest CBDA content (1.670%) compared to all other genotypes. The lowest content was obtained in Finola (0.324%), Santhica27 (0.208%), and USO31 (0.104%;
Figure 5).
Average CBD contents of inflorescence over the growth stages (S2-S4) ranged from 0.010% (Santhica27) to 0.497% (Ferimon). The highest CBD content was determined in inflorescence of Ferimon (0.497%) at growth stage S2, while e.g., Santhica27 only reached a CBD content of 0.045% at S2. No CBD was determined (n.d.) for the other genotypes at S2. At growth stage S3, CBD content ranged from 0.111% (USO31) to 0.331% (Fédora17) and at S4 from 0.010% (Santhica27) to 0.170% (Finola;
Table 3).
For the upper leaves at S1, the highest value was determined for Félina32 (0.155%), followed by USO31 (0.004%) and Fédora17 (0.003%). No CBD was determined for the other genotypes in upper leaves at the vegetative growth stage. The same applied for upper and lower leaves at growth stage S2, where the highest value was shown for Ferimon (0.060%), followed by Santhica27 (0.030%) and Futura75 (0.007%). At growth stage S3, CBD contents ranged from 0.016% (Santhica27) to 0.162% (Ferimon). The contents decreased at S4, in a range of 0.003% (Santhica27) to 0.090% (Félina32). In general, the mean CBD content of the lower leaf fraction obtained the lowest values. At S2, mostly no CBD was determined while at S3 the contents ranged from 0.080% (Futura75) to 0.006% (USO31;
Table 4).
Neutral cannabinoids do not exist at high concentrations in fresh plant material.
C. sativa L. biosynthesizes mainly the carboxylic acid forms of cannabidiol (CBD), cannabigerol (CBG), tetrahydrocannabinol (THC), namely cannabidiolic acid (CBDA), cannabigerolic acid (CBGA) and tetrahydrocannabinolic acid (THCA) [
32]. Cannabinoids are present in all aerial parts of the plant, correlated with the number of grandular trichomes, especially present on leaves and bracts [
33].
For CBDA, the acid precursor of CBD as decarboxylated form of the compound in growing plants, genotypes Futura75, Félina32 and Fédora17 indicated the highest values over all growth stages and all fractions of the plant (
Figure 3,
Figure 4 and
Figure 5). Genotype Ferimon likewise showed the highest CBDA content only at growth stage S2 and S3, whereas genotype Finola obtained the highest CBDA content in inflorescence at S4. In order to optimize CBD/A at field level, the growth stage at harvest and the plant fraction seems to be highly important. The concentration of cannabinoids depends on tissue type, genotype, age, harvest time and growth conditions as reported by Khan et al. [
34]. Stout et al. [
35] reported the highest CBD/A content in female flowers and a substantially lower content in the other tissues. In particular, dried inflorescence of genotype Futura75 contained the highest content of CBDA at growth stage S2 (2.593%) and S3 (2.719%). According to Hillig and Mahlberg [
36] phytocannabinoids are accumulated in inflorescence, which bear most of the trichomes produced by the hemp plant. In the upper leaf fraction, CBDA content increased over time, starting with the lowest value at growth stage S1 (0.902%), 1.322% at S2, while reaching the highest values at growth stage S3 (2.422%). At S4 the levels decreased to 1.674%. Both upper leaves and inflorescence showed the highest increase in CBDA at full-flowering stage (S3;
Figure 3 and
Figure 4). Results on the increase of CBDA in leaves to a maximum at S3 followed by a subsequent decrease with plant age and a maximum content in the upper leaves were in line with results of Pacifico et al. [
37]. The highest CBDA content was recorded in leaves of a fibre genotype with 2.40% 76 days from sowing. After 76 days, a decrease of the average CBDA content in leaves was observed [
37]. Mandolino and Ranalli [
38] reported that in proximity of full-flowering, the content of cannabinoids reached a maximum in trichome-rich organs like inflorescence. The same trend was observed for genotypes Fédora17 and Félina32. The content of CBDA in inflorescence showed a maximum at growth stage S2 (2.430% and 2.941%) and S3 (2.220% and 2.533%), respectively (
Figure 3).
The higher CBDA content was found in genotypes Futura75, Fédora17, Félina32 and Ferimon. Note that, these industrial hemp genotypes, which comply with the 0.2% THC/A threshold set by the EU legislation restricts the choice of genotypes for European farmers, compared to producers in Switzerland, North America, Asia and Canada (limits from 0.3% up to 1%). However, the CBDA, differs in respect to DW yield between growth stages. Campbell et al. [
39] reported, that 83% of variance in CBD/A content resulted from genetic effects, making genotype selection important when seeking high CBD/A contents. Environmental factors such as the positive correlation between CBD/A content and water availability found by Calzolari et al. [
23] slightly increase CBD/A content, but the change was small enough that inflorescence yield is far more important to overall yield than a slight change in CBD/A percentage [
39].
While genotype Futura75 generated a 84% higher inflorescence yield, genotypes Fédora17, Félina32 and Ferimon reached a 86%, 83% and 78% higher inflorescence yield at S4, compared to S3. No significant differences were recorded for upper leaf DW between growth stage S3 and S4 (
Table 2). Taking these results into account, the calculated, total CBDA yield per square meter was higher at S4. In particular, genotype Futura75 reached a 76% higher CBDA yield at S4 (4568.26 mg m
−2) compared to S3 (1104.32 mg m
−2). The same trend was observed for the other three genotypes Fédora17, Félina32 and Ferimon, with a CBDA yield of 1350.20 mg m
−2, 1324.46 mg m
−2 and 1012.96 mg m
−2 at S3, in comparison to 6011.20 mg m
−2, 4975.60 mg m
−2 and 1929.60 mg m
−2 at S4, respectively.
The industrial hemp genotypes are mainly bred for fibre and seed production. Both, Futura75 and Fedora17 are candidate cultivars for a dual-purpose production in the EU, with Futura75 being more suitable for fibre production and Fédora17 for seed production [
28]. At full-flowering, stem yield for bast fibres is positively correlated with the duration of vegetative growth, with a tendency to be high in intermediate flowering genotypes, such as Futura75. Inflorescence yield and accordingly seed production of early flowering genotypes, such as Fédora17, Félina32, Fermion and USO31 are proven to be higher [
20].
This study found that industrial hemp genotypes are suitable for non-psychotropic cannabinoid production, namely CBD, CBG and their acid precursors. In addition to seed and fibre production, recently, hemp genotypes registered within the EU, have been cultivated for inflorescence to finally extract non-psychotropic cannabinoids [
9]. The utilization of harvested inflorescence for cannabinoid extraction, seems to exclude seed production at the same time as plants are harvested at full-flowering stage, thus limiting the full exploitation of the hemp crop [
7]. However, threshing residues of inflorescence obtained at seed maturity might offer a unexploited high-value product for the extraction of cannabinoids [
21].
Within the tested genotypes, CBD/A was determined in inflorescence at full-flowering stage, as well as in inflorescence resulting from seed threshing. In addition, higher dry matter yields of inflorescence per square meter at S4, supported the assumption, that these genotypes can be cultivated as multi-purpose crop: for seed, oil and fibre production as well as for the additional extraction of cannabinoids out of the remaining material. Cannabinoids can be extracted from the reproductive plant parts and foliage. The inflorescence material has a higher concentration of cannabinoids than foliage material, however foliage part have larger biomass of the hemp plant [
40]. The breeding of genotypes with superior characteristics is required to optimize both seed and fibre production, but also the quality and quantity of cannabinoids by residual inflorescence and upper leaves.
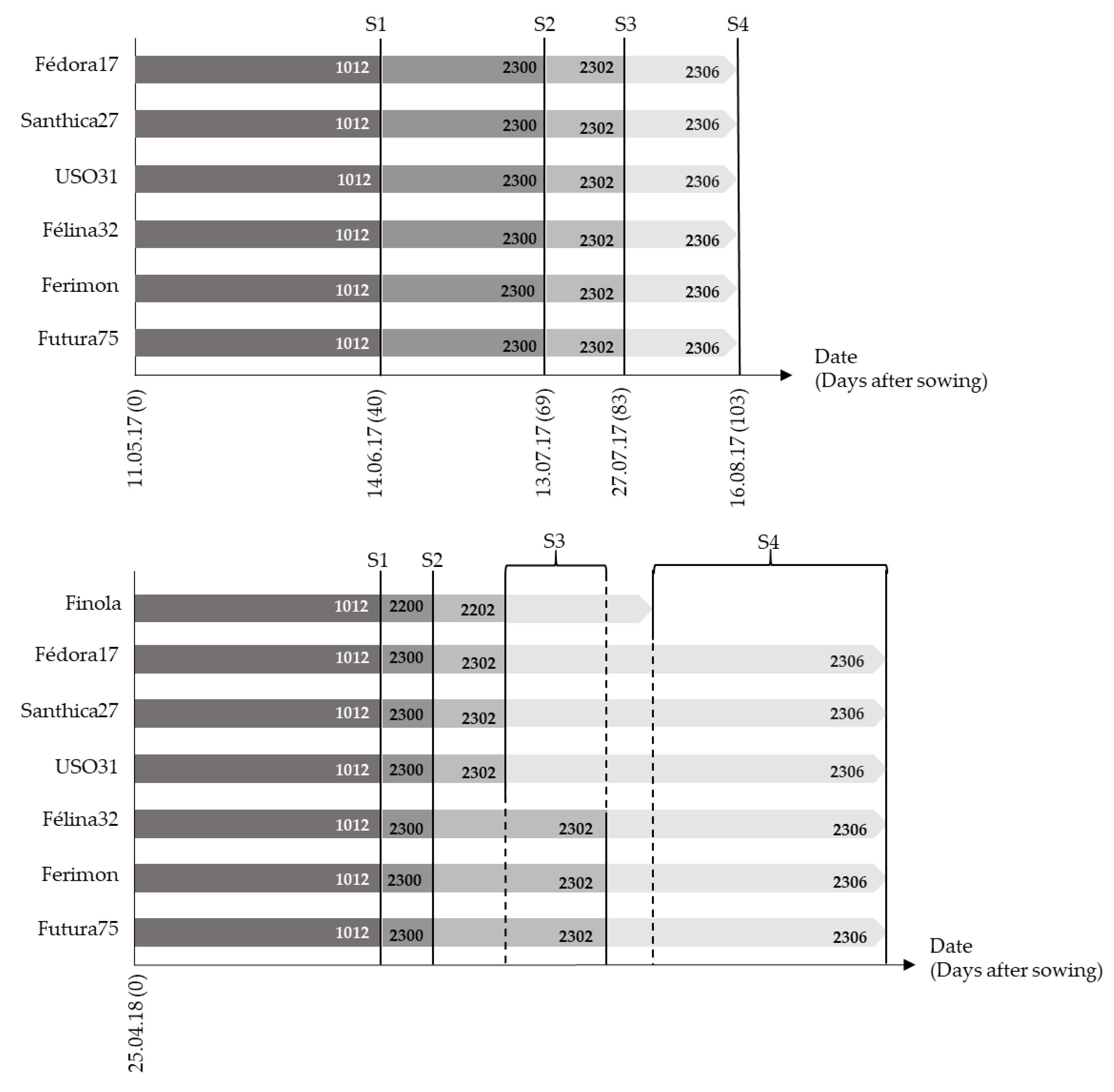
Furthermore, the only diocious industrial hemp genotype, Finola, which is specifically bred for oil production [
41], recorded a 85% higher DW yield of inflorescence at full-flowering stage (S3) in comparison to the average yield of the other genotypes (
Table 2). Subsequently, CBDA yield per square meter amounted to 5422.41 mg m
−2, and showed in average a 78% higher yield of CBDA per square meter compared with the other genotypes. No significant differences were shown for Finola in CBDA content between S3 and S4, as well as in DW of inflorescence at S4. If Finola is cultivated for single purpose only, cannabinoid extraction can be carried out from full-flowering to seed maturity. Aiming at dual or multi-purpose, Finola can be harvested after seed maturity. Notable is also the length of the cultivation period: while genotype Finola reached stage S3 after 62 days, together with Fédora17, Santhica27 and USO31, the other genotypes needed 80 days to reach full-flowering (S3) in 2018. Furthermore, after 80 days, Finola reached the seed maturity stage, whereas the other six genotypes reached seed maturity after 130 days. A short vegetation period, combined with a high DW yield and CBDA content, resulted in an optimized land utilization. Moreover, a 10:1 ratio of CBD/THC, is above the recent EU requirements of 2:1 [
30], what outlines Finola with a high CBDA/low THCA profile as an interesting genotype for cannabidiol [
35].
3.2.2. CBGA and CBG Content of Genotype Santhica27
The first cannabinoid synthetized is cannabigerol (CBG), produced by condensation of a phenol-derived olivetolic acid and a terpene-based geranyl diphosphate. The process is catalysed by geranyldiphosphate:olivetolate geranyltransferase (GOT) [
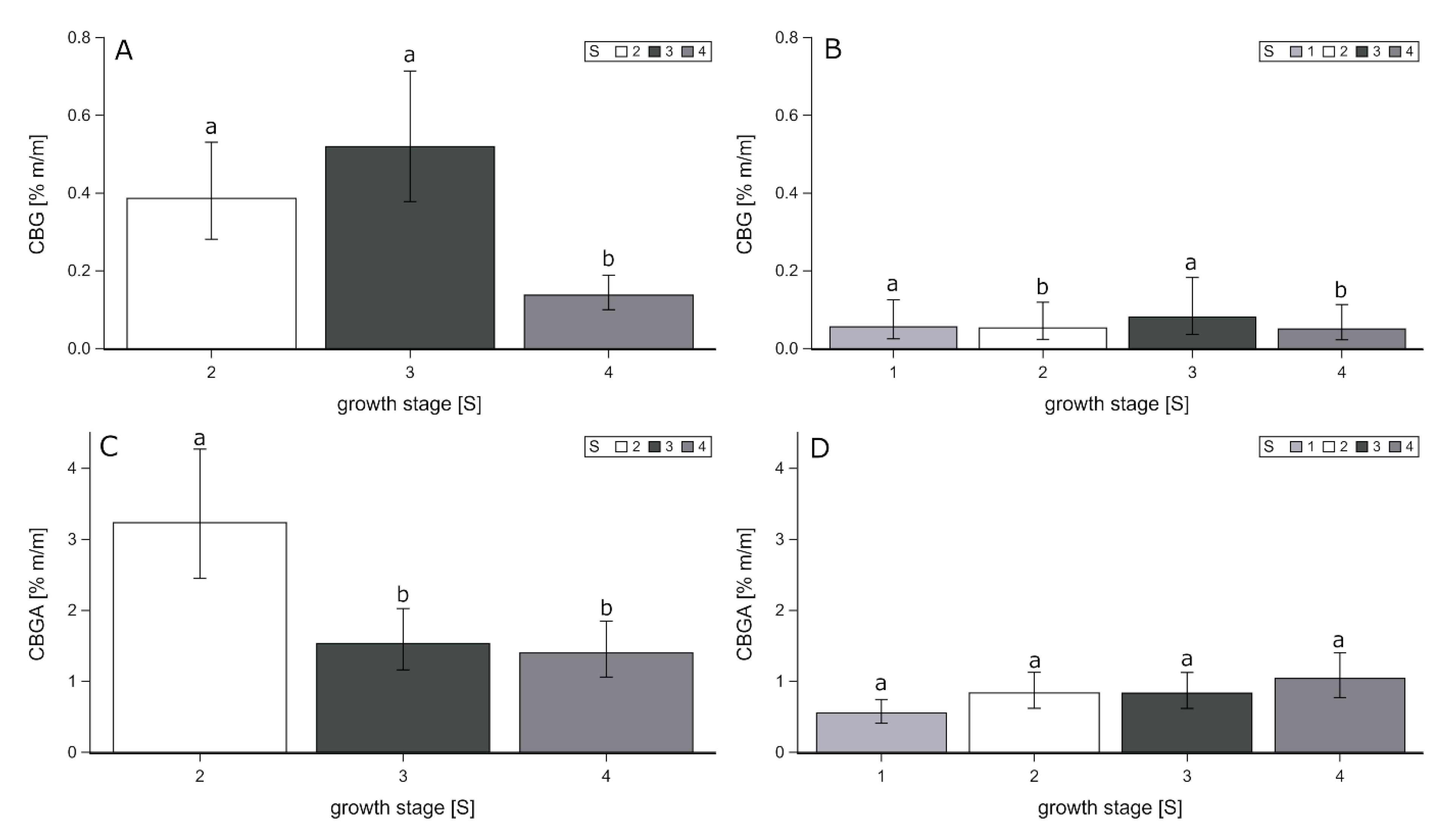
42]. CBG was only determined in considerable amounts in genotype Santhica27 (
Figure 6A–D). Fournier et al. [
43] reported a new chemotype, initially found in a French fibre hemp population, normally predominant in CBD, having CBG as the major constituent. In these genotypes the pathway CBG to tetrahydrocannabinol (THC) or cannabidiol (CBD) is largely obstructed [
44]. In the present study, average CBG contents in the inflorescence over the growth stages S2 to S4 ranged from 0.137% (S4) to 0.520% (S3;
Figure 6A). The highest contents were found at growth stage S2 (0.386%) and S3 (0.520%). In the upper leaf fraction the contents ranged from 0.050% (S2) to 0.081% (S1;
Figure 6B). The highest contents were found at S1 (0.056%) and S3 (0.081%). The lowest amounts were determined in lower leaves; maximum levels were recorded at growth stage S3 with 0.008%. The content of cannabigerolic acid (CBGA), the acid precursor of CBG, showed the highest content in the inflorescence at growth stage S2, with 3.235%, followed by S3, with 1.534%. The lowest content was determined at growth stage S4, with 1.399%, compared to S2 (
Figure 6C). In the upper leaves the content of CBGA did not show any statistical differences among the growth stage S2 to S4, while the values ranged from 0.552% (S1) to 1.040% (S4;
Figure 6D). The lowest content was determined again in lower leaves, ranging from 0.085% (S2) to 0.801% (S3). This is in agreement with results on CBG concentrations between 0.4 and 1.2% in leaves and inflorescence of threshing residues reported by Calzolari et al. [
23].
Overall, the highest amounts of CBG and the acid precursor CBGA were determined in inflorescence of Santhica27, at growth stage S2 and S3. The CBG content of upper leaves showed a maximum at growth stage S1 and S3, with no statistical differences and a maximum of CBGA at growth stages S2 to S4. Referring to CBG accumulation Pacificio et al. [
37], stated that in high-CBG plants, a maximum level of CBG accumulation proceeded before the maximum CBD accumulation in leaves was obtained. With regard to a high exploitation of CBG, over all fractions, a harvest at growth stage S2 or S3 can be recommended. This is in contrast to studies of Calzolari et al. [
23], where harvest time did not have an effect on CBG content of genotype Santhica27. Particularly, with regard to DW yield, genotype Santhica27 showed a 93% higher DW yield of inflorescence per square meter at S4, compared to S3. In this respect, a CBGA yield of 416.79 mg m
−2 was calculated for S3, compared to 5721.77 mg m
−2 at S4, with a DW yield of 408.99 g m
−2. For S3 a CBG yield of 141.28 mg m
−2 was calculated, while at S4 a yield of 560.32 mg m
−2 CBG was reached. Whilst CBG/A was found exclusively in genotype Santhica27, it shows the potential of this genotype to be used for CBG/A extraction as well as seed and fibre production.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}