Elevated Root-Zone Dissolved Inorganic Carbon Alters Plant Nutrition of Lettuce and Pepper Grown Hydroponically and Aeroponically


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Measurements
2.2. Nutrient Analysis
2.3. Total Nitrogen and NO3− Analysis
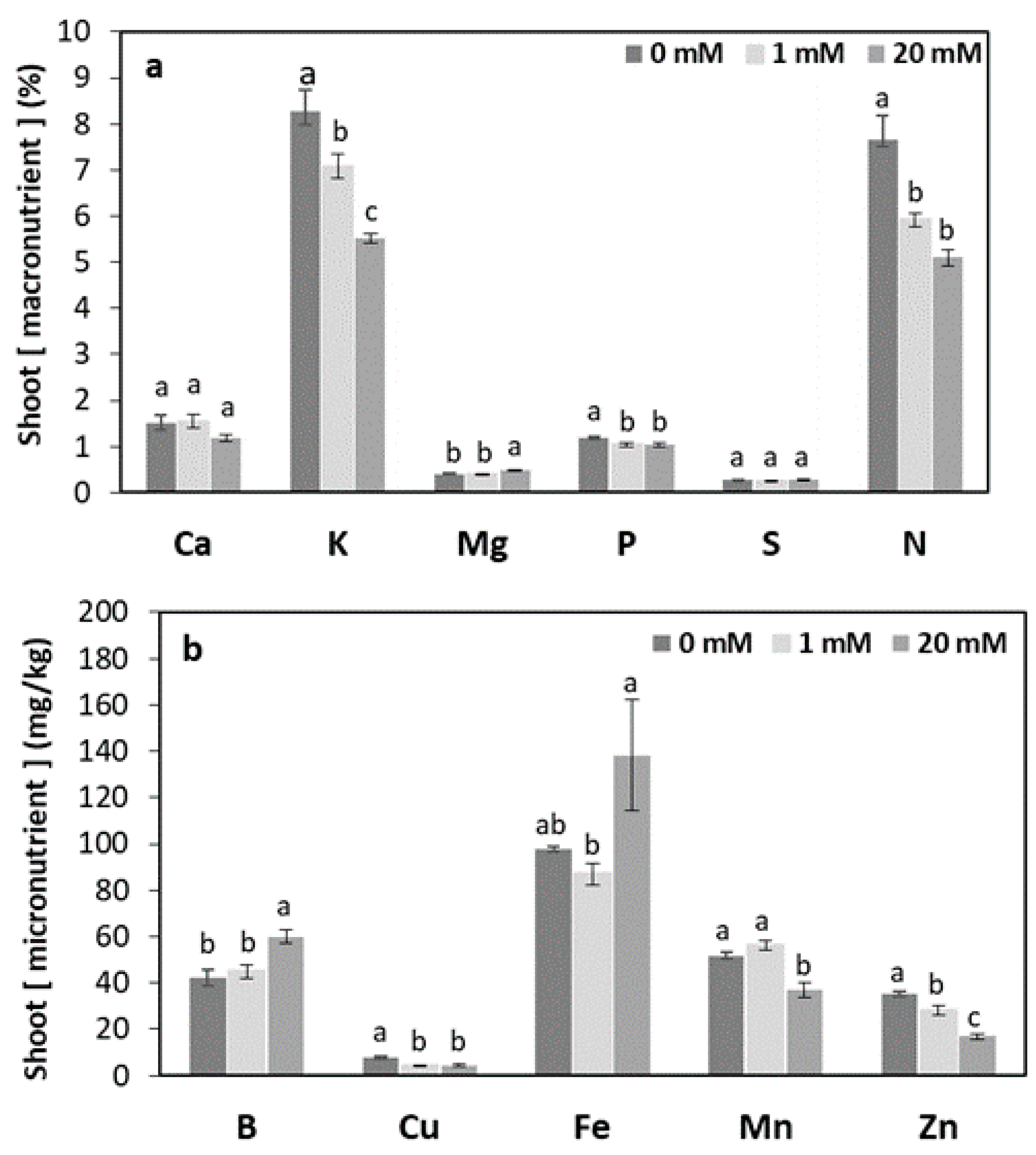
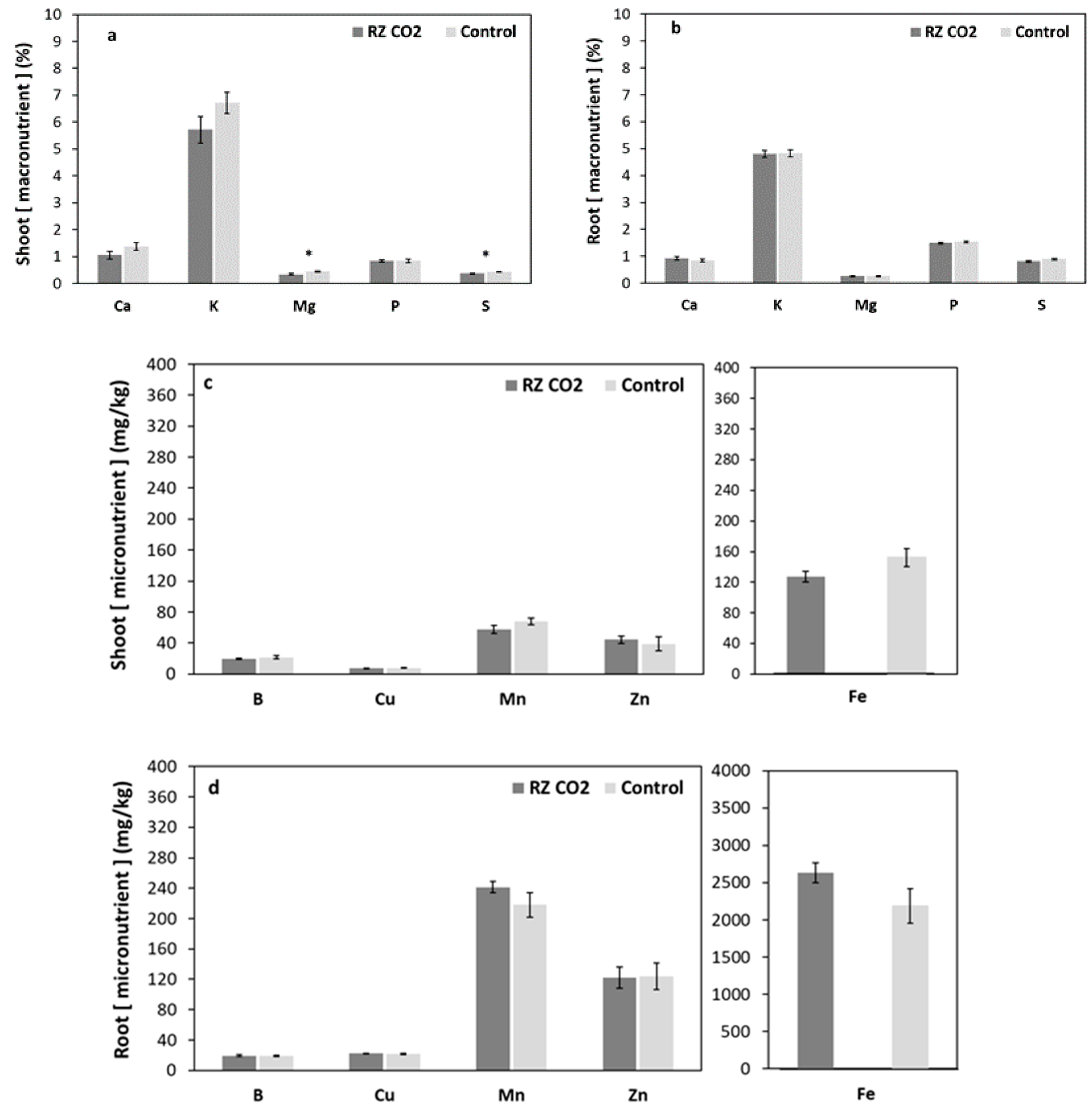
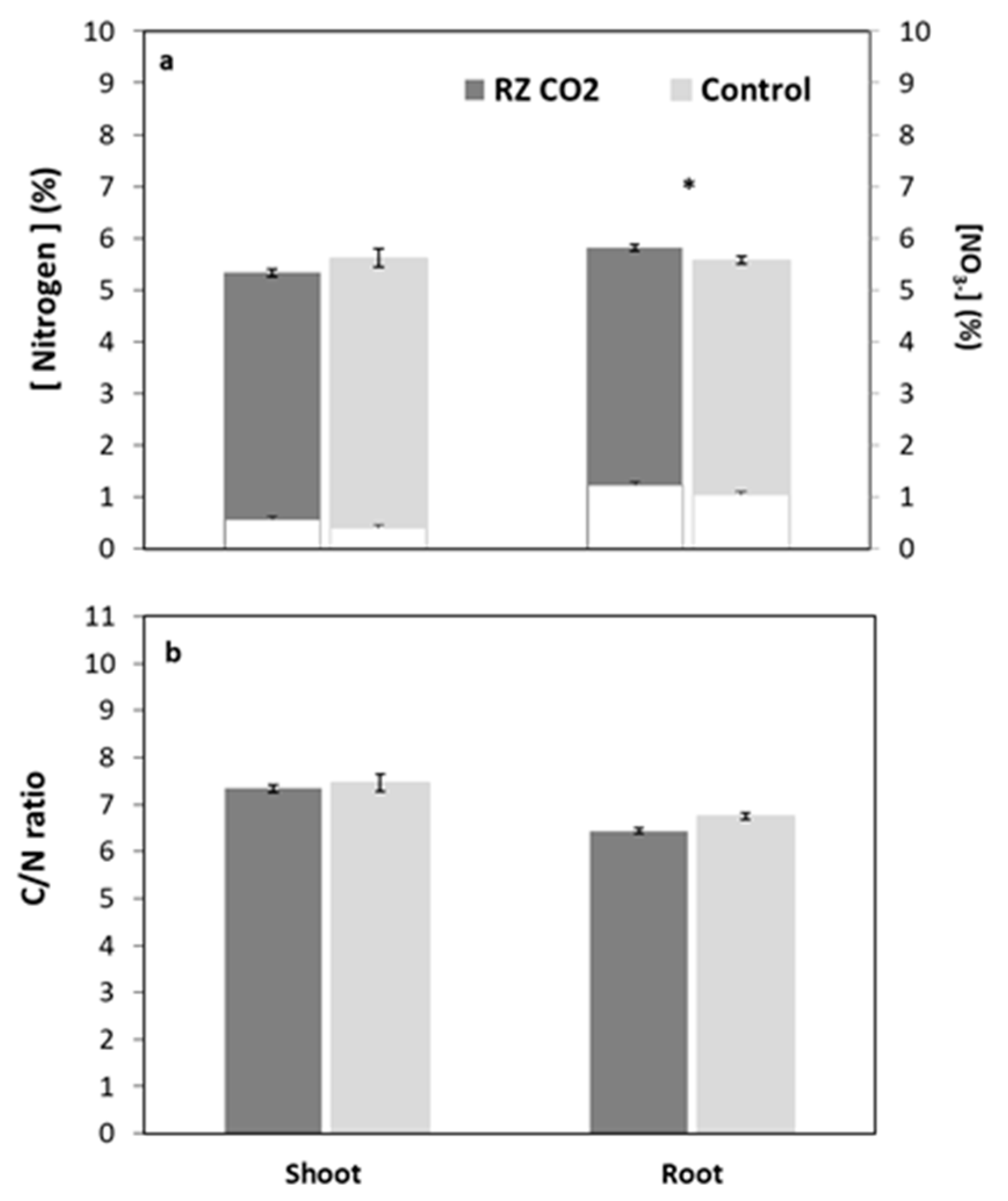
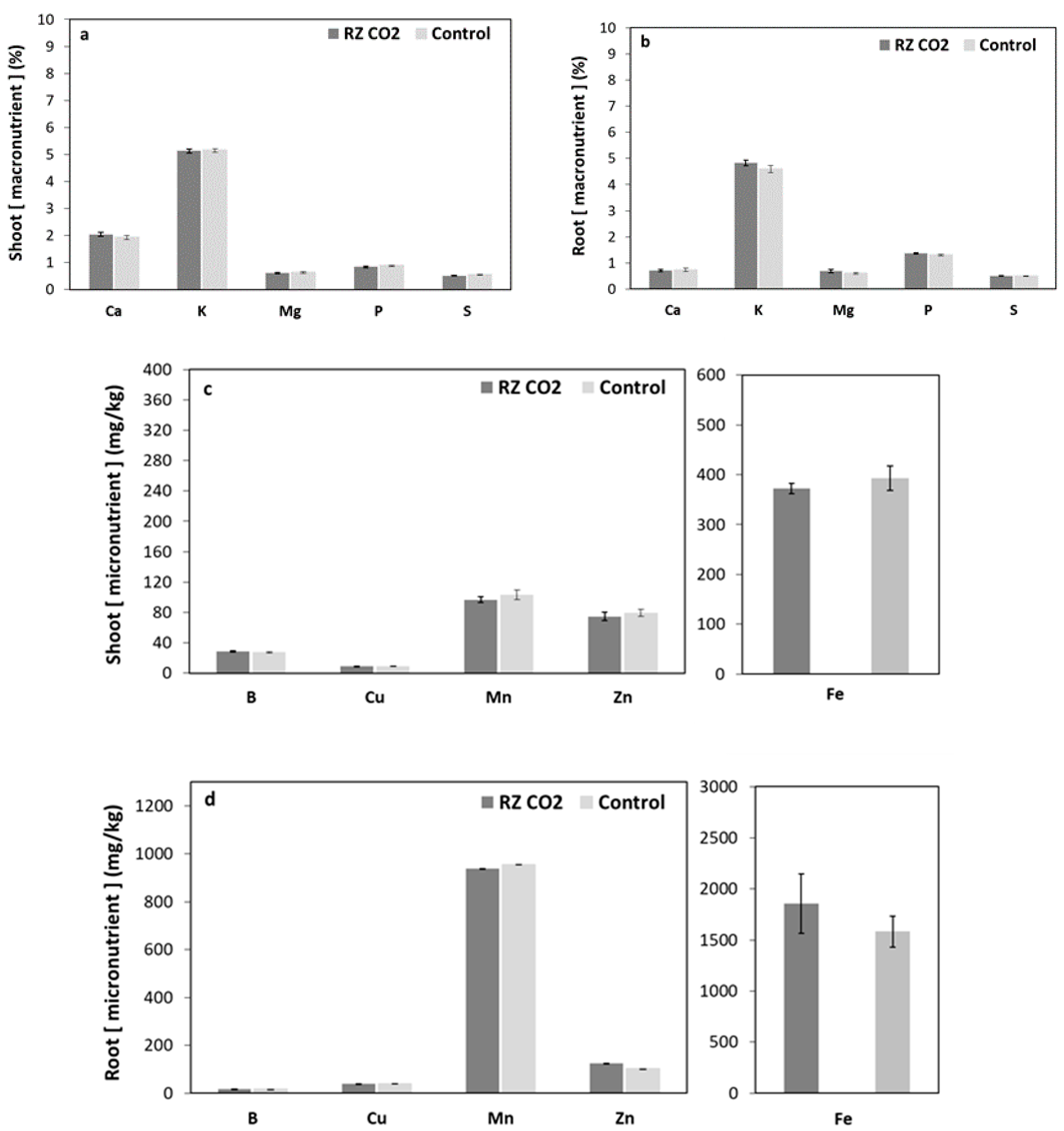
3. Results
3.1. Root-zone Bicarbonate Enrichment of Plants Grown Hydroponically
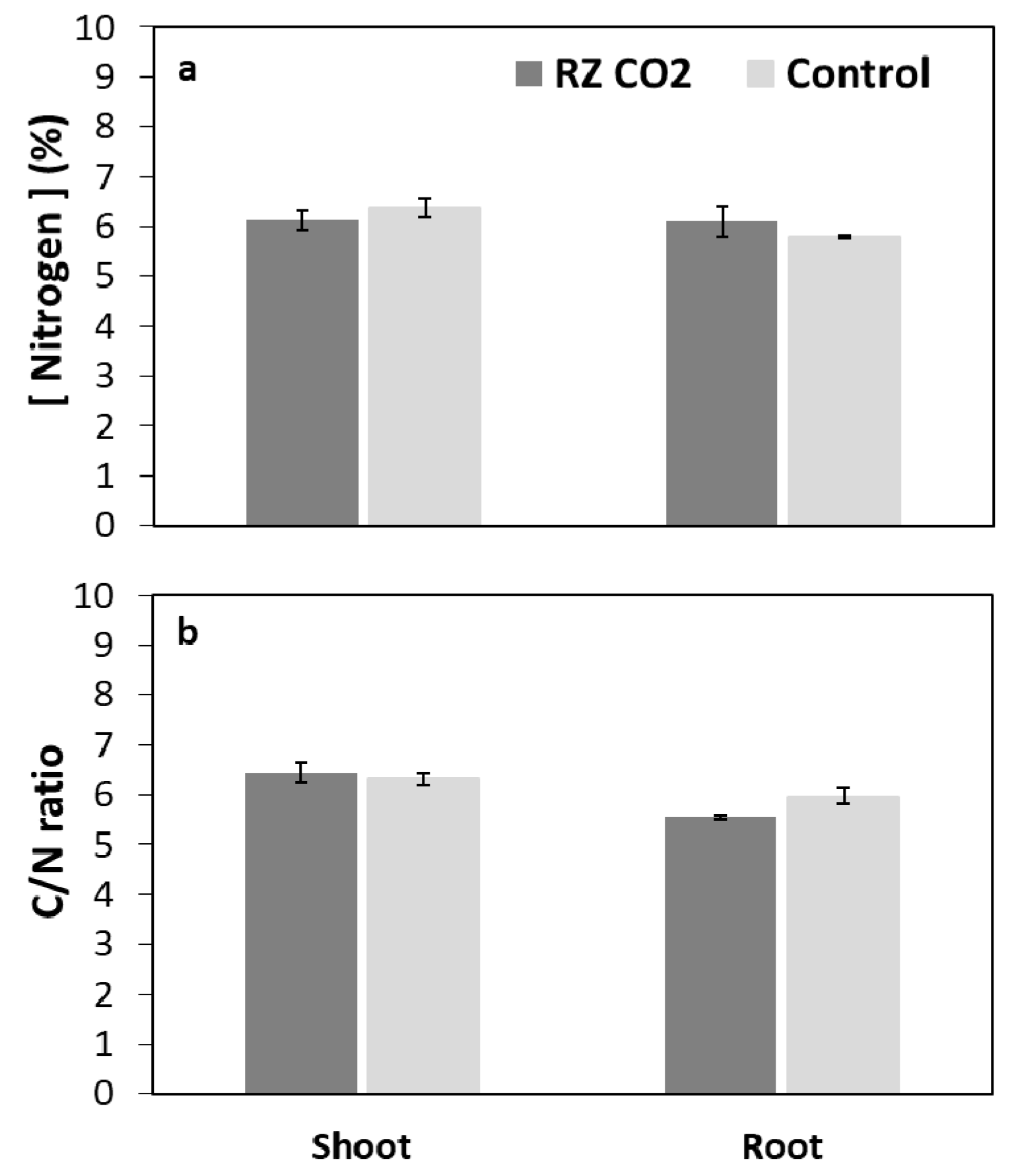
3.2. Root-Zone Carbon Dioxide Enrichment of Plants Grown Aeroponically
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Status | Macronutrients (%) | |||||
|---|---|---|---|---|---|---|
| N | P | K | Ca | Mg | S | |
| Deficient | <4 | 0.4 | 5 | 1 | 0.3 | |
| In range | 4–6 | 0.4–0.6 | 5–7 | 1–2 | 0.3–0.6 | 0.2–0.3 |
| High | >6 | 0.6 | 7 | 2 | 0.6 | |
| Micronutrients (mg/kg) | ||||||
| B | Mn | Cu | Zn | Fe | ||
| Deficient | 15 | 20 | 5 | 40 | <50 | |
| In range | 15–30 | 20–40 | 5–10 | 40–60 | 50–150 | |
| High | 30 | 40 | 10 | 60 | >150 | |
| Toxic | >100 | >250 | ||||
References
- Clark, I.; Fritz, P. Environmental Isotopes in Hydrogeology; Starkweather, A.W., Ed.; CRC Press LLC: Boca Raton, FL, USA, 1997. [Google Scholar]
- Bai, S.G.; Jiao, Y.; Yang, W.Z.; Gu, P.; Yang, J.; Liu, L.J. IOP Conference Series: Earth and Environmental. Science 2017, 100, 012129. [Google Scholar]
- Enoch, H.Z.; Olesen, J.M. Plant response to irrigation with water enriched carbon dioxide. New Phytol. 1993, 125, 249–258. [Google Scholar] [CrossRef]
- Bialczyk, J.; Lechowski, Z.; Libik, A. Growth of tomato seedlings under different HCO− concentration in the medium. J. Plant Nutr. 1994, 17, 801–816. [Google Scholar] [CrossRef]
- Bialczyk, J.; Lechowski, Z.; Libik, A. Early vegetative growth of tomato plants in media containing nitrogen source as nitrate, ammonium, or various nitrate-ammonium mixtures with bicarbonate addition. J. Plant Nutr. 2005, 27, 1687–1700. [Google Scholar] [CrossRef]
- Cramer, M.D.; Richards, M.D. The effect of rhizosphere dissolved inorganic carbon on gas exchange characteristics and growth rates of tomato seedlings. J. Exp. Bot. 1999, 50, 79–87. [Google Scholar] [CrossRef]
- He, J.; Austin, P.T.; Lee, S.K. Effects of elevated root zone CO2 and air temperature on photosynthetic gas exchange, nitrate uptake, and total reduced nitrogen content in aeroponically grown lettuce plants. J. Exp. Bot. 2010, 61, 3959–3969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Austin, P.T.; Nichols, M.A.; Lee, S.K. Elevated root-zone CO2 protects lettuce plants from midday depression of photosynthesis. Environ. Exp. Bot. 2007, 61, 94–110. [Google Scholar] [CrossRef]
- Zhao, K.; Wu, Y. Effects of zinc deficiency and bicarbonate on the growth and photosynthetic characteristic of four plant species. PLoS ONE 2017, 12, e0169812. [Google Scholar]
- Poschenrieder, C.; Fernández, J.A.; Rubio, L.; Perez, L.; Terés, J.; Barceló, J. Transport and use of bicarbonate in plants: Current knowledge and challenges ahead. Int. J. Mol. Sci. 2018, 19, 1352. [Google Scholar] [CrossRef] [Green Version]
- Fageria, N.K.; Nascente, A.S. Chapter six: Management of soil acidity of South American soils for sustainable crop production. Adv. Agron. 2014, 128, 221–275. [Google Scholar]
- Alhendawi, R.A.; Romheld, V.; Kirby, E.A.; Marschner, H. Influence of increasing bicarbonate concentrations on plant growth, organic acid accumulation in roots and iron uptake by barley, sorghum and maize. J. Plant Nutr. 1997, 20, 1721–1735. [Google Scholar] [CrossRef]
- Al mansouri, H.M.; Alhendawi, R.A. Effect of Increasing Concentration of Bicarbonate on Plant Growth and Nutrient Uptake by Maize Plants. Am. Eur. J. Agric. Environ. Sci. 2014, 14, 1–6. [Google Scholar]
- Zhao, X.; Li, T.L.; Sun, Z.P. Effects of prolonged root-zone CO2 treatment on morphological parameter and nutrient uptake of tomato grown in aeroponic system. J. Appl. Bot. Food Qual. 2010, 83, 212–216. [Google Scholar]
- Cataldo, D.A.; Maroon, M.; Schrader, L.E.; Youngs, V.L. Rapid colorimetric determination of nitrate in plant-tissue by nitration of salicylic-acid. Commun. Soil Sci. Plant Anal. 1975, 6, 71–80. [Google Scholar] [CrossRef]
- Rehman, H.; Aziz, T.; Farooq, M.; Wakeel, A.; Rengel, Z. Zinc nutrition in rice production systems: A review. Plant Soil 2012, 361, 203–226. [Google Scholar] [CrossRef]
- Rathore, V.S. Effect of Zn2+ stress in rice (Oryza sativa cv. Manhar) on Growth and Photosynthetic Processes. Photosynthetica 1995, 31, 571–584. [Google Scholar]
- Randall, P.J.; Bouma, D. Zinc deficiency, carbonic anhydrase, and photosynthesis in leaves of spinach. Plant Physiol. 1973, 52, 229–232. [Google Scholar] [CrossRef] [Green Version]
- Van Eysinga, J.R.; Smilde, K.W. Nutritional Disorders in Glasshouse Tomatoes, Cucumbers and Lettuce; Centre for Agricultural Publications and Documentations: Wageningen, The Netherlands, 1981. [Google Scholar]
- Liu, D.; Jiang, W.; Zhang, L.; Li, L. Effects of boron ions on root growth and cell division of broad bean (Vicia faba L.). Isr. J. Plant Sci. 2000, 48, 47–51. [Google Scholar] [CrossRef]
- Lou, Y.; Yang, Y.; Xu, J. Effect of boron fertilization on B uptake and utilization by oilseed rape (Brassica napus L.) under different soil moisture regimes. J. Appl. Ecol. 2001, 12, 478–480. [Google Scholar]
- Ozturk, O.; Soylu, S.; Ada, R.; Gezgin, S.; Babaoglu, M. Studies on differential response of spring canola cultivars to B toxicity. J. Plant Nutr. 2010, 33, 1141–1154. [Google Scholar] [CrossRef]
- Bertoni, G.M.; Pissaloux, A.; Morard, P.; Sayag, D.R. Bicarbonate-pH relationship with iron chlorosis in white lupine. J. Plant Nutr. 1992, 15, 1509–1518. [Google Scholar] [CrossRef]
- De la Guardia, M.; Alcantara, E.V. Bicarbonate and low iron level increase root to total plant weight ratio in olive and peach rootstock. J. Plant Nutr. 2007, 25, 1021–1032. [Google Scholar] [CrossRef]
- Siddiqi, M.Y.; Malhotra, B.; Min, X.; Glass, A.D.M. Effects of ammonium and inorganic carbon enrichment on growth and yield of a hydroponic to-mato crop. J. Plant Nutr. Soil Sci. 2002, 165, 191–197. [Google Scholar] [CrossRef]
- Verbruggen, N.; Hermans, C. Physiological and molecular responses to magnesium nutritional imbalance in plants. Plant Soil 2013, 368, 87–99. [Google Scholar] [CrossRef]
- Senbayram, M.; Gransee, A.; Wahle, V.; Thiel, H. Role of magnesium fertilisers in agriculture: Plant-soil continuum. Crop Pasture Sci. 2015, 66, 1219–1229. [Google Scholar] [CrossRef]
- Cakmak, I.; Yazici, A.M. Magnesium: A forgotten element in crop production. Better Crops 2010, 94, 23–25. [Google Scholar]
- Zhou, H.J.; Korcak, R.F.; Fan, F.; Faust, M. The effect of bicarbonate induced Fe chlorosis on mineral content and Ca45 uptake of apple seedlings. J. Plant Nutr. 1984, 7, 1355–1364. [Google Scholar] [CrossRef]
- Morales, F.; Grasa, R.; Abadia, A.; Abadia, J. Iron chlorosis paradox in fruit trees. J. Plant Nutr. 1998, 21, 815–825. [Google Scholar] [CrossRef]
- Roosta, H.R.; Pourebrahimi, M.; Hamidpour, M. Effects of bicarbonate and different Fe sources on vegetative growth and physiological characteristics of bell pepper (Capsicum annuum L.) plants in hydroponic system. J. Plant Nutr. 2015, 38, 397–416. [Google Scholar] [CrossRef]
- Mengel, K. Iron availability in plant tissues—Iron chlorosis on calcareous soils. Plant Soil 1994, 165, 275–283. [Google Scholar] [CrossRef]
- Romheld, V. The chlorosis paradox: Fe inactivation in leaves as a secondary event in Fe deficiency chlorosis. J. Plant Nutr. 2000, 23, 1629–1643. [Google Scholar] [CrossRef]
- Bialczyk, J.; Lechowski, Z.; Dziga, D. Composition of the xylem sap of tomato seedlings cultivated on media with HCO3− and nitrogen source as NO3− or NH4+. Plant Soil 2004, 263, 265–272. [Google Scholar] [CrossRef]
- Loladze, I. Rising atmospheric CO2 and human nutrition: Toward globally imbalanced plant stoichiometry? Trends Ecol. Evol. 2002, 17, 457–461. [Google Scholar] [CrossRef]
- Leustek, T.; Martin, M.N.; Bick, J.A.; Davies, J.P. Pathways and regulation of sulfur metabolism revealed through molecular and genetic studies. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 141–165. [Google Scholar] [CrossRef] [PubMed]
- Saito, K. Biosynthesis of cysteine. In Plant Amino Acids: Biochemistry and Biotechnology; Singh, B., Ed.; Dekker: New York, NY, USA, 1999; pp. 267–291. [Google Scholar]
- Martin, T.; Oswald, O.; Graham, I.A. Arabidopsis seedling growth, storage lipid mobilization, and photosynthetic gene expression are regulated by carbon:nitrogen availability. Plant Physiol. 2002, 128, 472–481. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.L. Carbon and nitrogen nutrient balance signaling in plants. Plant Signal Behav. 2009, 4, 4584–4591. [Google Scholar] [CrossRef] [Green Version]
- Andrews, M. Nitrate and reduced-N concentrations in the xylem sap of Stellaria media, Xanthium strumarium and 6 legume species. Plant Cell Environ. 1986, 9, 605–608. [Google Scholar]
- Andrews, M.; Morton, J.D.; Lieffering, M.; Bisset, L. The partitioning of nitrate assimilation between root and shoot of a range of temperate cereals and pasture grasses. Ann. Bot. 1992, 70, 271–276. [Google Scholar] [CrossRef]
- Viktor, A.; Cramer, M.D. The influence of root assimilated inorganic carbon on nitrogen acquisition/assimilation and carbon partitioning. New Phytol. 2005, 165, 157–169. [Google Scholar] [CrossRef]
- Fageria, V.d. Nutrient interactions in crop plants. J. Plant Nutr. 2001, 24, 1269–1290. [Google Scholar] [CrossRef]
- Wang, J.; Liu, X.; Zhang, X.; Li, L. Changes in plant C, N and P ratios under elevated [CO2] and canopy warming in a rice winter wheat rotation system. Sci. Rep. 2019, 9, 5424. [Google Scholar] [CrossRef] [Green Version]
- Thoresby, P.; Thornton, I. Heavy metals and arsenic in soil, pasture herbage and barley in some mineralised areas in Britain. In Trace Substances in Environmental Health; Hemphill, D.D., Ed.; University of Missouri: Columbia, MO, USA, 1979. [Google Scholar]
- Kabata-Pendias, A.; Pendias, H. Trace Elements in Soils and Plants; CRC Press: Boca Raton, FL, USA, 2001. [Google Scholar]
- Broadley, M.R.; White, P.J.; Hammond, J.P.; Zelko, I.; Lux, A. Zinc in plants. New Phytol. 2007, 173, 677–702. [Google Scholar] [CrossRef] [PubMed]
- Brennan, R.F. Zinc Application and Its Availability to Plants. Ph.D. Thesis, School of Environmental Science, Division of Science and Engineering, Murdoch University, Murdoch, Australia, 2005. [Google Scholar]
- Pearson, J.N.; Rengel, Z.; Jenner, C.F.; Graham, R.D. Transport of zinc and manganese to developing wheat grains. Plant Physiol. 1995, 95, 449–455. [Google Scholar] [CrossRef]
- Haslett, B.S.; Reid, R.J.; Rengel, Z. Zinc mobility in wheat: Uptake and distribution of zinc applied to leaves or roots. Ann. Bot. 2001, 87, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Tao, Q.; Liang, C.; Yang, X. Elevated CO2 concentration increase the mobility of Cd and Zn in the rhizosphere of hyperaccumulator Sedum alfredii. Environ. Sci. Pollut. Res. 2014, 21, 5899–5908. [Google Scholar] [CrossRef]
- Rodriguez, J.H.; Klumpp, A.; Fangmeier, A.; Pignata, M.L. Effects of elevated CO2 concentrations and fly ash amended soils on trace element accumulation and translocation among roots, stems and seeds of Glycine max (L.) Merr. J. Hazard. Mater. 2011, 187, 58–66. [Google Scholar] [CrossRef] [Green Version]
- Hartz, T.; Johnstone, P.R.; Williams, E.; Smith, R.F. Establishing lettuce leaf and nutrient optimum ranges through DRIS analysis. HortScience 2007, 42, 143–146. [Google Scholar] [CrossRef]
- Hochmuth, G.; Maynard, D.; Vavrina, E.; Simmone, E. Plant Tissue Analysis and Interpretation for Vegetable Crops in Florida. UF/IFAS, Fla. Coop. Ext. Serv., HS964. Available online: http://edis.ifas.ufl.edu (accessed on 2 July 2019).





| HCO3− | 0 mM | 1 mM | 5 mM | 10 mM | 20 mM |
|---|---|---|---|---|---|
| Shoot fresh weight (g) | 33.2 ± 0.8 b | 43.7 ± 0.8 a | 44.3 ± 1.4 a | 36.6 ± 2.8 b | 21.7 ± 2 c |
| Shoot dry weight (g) | 2.1 ± 0.0 b | 2.5 ± 0.0 a | 2.4± 0.0 a | 2.0± 0.1 b | 1.4 ± 0.1 c |
| Leaf area (cm2) | 820 ± 18.2 b | 978 ± 27.6 a | 965 ± 31.6 a | 786 ± 51.1 b | 508 ± 42.4 c |
| Root dry weight (g) | 0.4 ± 0.0 c | 0.4 ± 0.0 bc | 0.5± 0.0 ab | 0.6± 0.0 a | 0.6 ± 0.0 a |
| HCO3− | 0 mM | 1 mM | 5 mM | 10 mM | 20 mM |
|---|---|---|---|---|---|
| Shoot fresh weight (g) | 59.5 ± 1.8 b | 70.6 ± 2.3 a | 60.5 ± 1.6 b | 55.2 ± 2.8 b | 45.3 ± 1.7 c |
| Shoot dry weight (g) | 5.6 ± 0.2 b | 6.5 ± 0.2 a | 5.6 ± 0.1 b | 5.2 ± 0.2 b | 4.3 ± 0.2 c |
| Leaf area (cm2) | 1190 ± 27 abc | 1286 ± 43 a | 1172 ± 27 bc | 1090 ± 47 c | 915 ± 29 d |
| Root dry weight (g) | 1.3 ± 0.0 b | 1.6 ± 0.0 a | 1.5 ± 0.0 a | 1.7± 0.1 a | 1.5 ± 0.0 a |
| HCO3− | 0 mM | 1 mM | 20 mM |
|---|---|---|---|
| Shoot fresh weight (g) | 28.2 ± 0.7 a | 32.3 ± 1.6 a | 13.3 ± 0.9 b |
| Shoot dry weight (g) | 1.70 ± 0.04 a | 1.94 ± 0.09 a | 0.80 ± 0.05 b |
| Leaf area (cm2) | 543 ± 32 b | 677 ± 34 a | 279 ± 16 c |
| Plant Growth Rate (mg g−1 /d) | 2.52 ± 0.03 b | 3 ± 0.1 a | 1.3 ± 0.1 c |
| mg | Ca | K | Mg | P | S | N |
|---|---|---|---|---|---|---|
| Control | 25.5 ± 3.1 a | 140.1 ± 10.1 a | 6.7 ± 0.5 a | 20.2 ± 0.8 a | 4.6 ± 0.2 a | 129.3 ± 7.5 a |
| 1 mM | 26.6 ± 4.7 a | 120.9 ± 16.1 a | 6.6 ± 0.8 a | 17.5 ± 2.0 a | 4.2 ± 0.5 a | 101.3 ± 13.3 a |
| 20 mM | 9.5 ± 1.1 b | 44.3 ± 3.7 b | 3.82 ± 0.4 b | 8.3 ± 0.7 b | 2.12 ± 0.2 a | 40.9 ± 3.6 b |
| µg | B | Cu | Fe | Mn | Zn | |
| Control | 87.7± 4.3 a | 1.3± 0.1 a | 59.4 ± 3.5 a | 165.5 ± 4.6 a | 71.7 ± 7.5 b | |
| 1 mM | 95.8 ±12.9 a | 0.7± 0.1 b | 47.2 ± 3.6 b | 149.3 ±22 a | 77.3 ± 12.2 ab | |
| 20 mM | 30.1 ± 4.5 b | 0.4 ± 0.0 c | 13.6 ± 1.7 c | 112.5 ± 22.9 a | 48.4 ± 5.1 bc |
| Treatment | Shoot Fresh Weight (g) ± SE | Shoot Dry Weight (g) ± SE | Root Dry Weight (g) ± SE | |
|---|---|---|---|---|
| Experiment 4 Lettuce | Control | 25.08 ± 2.04 | 1.24 ± 0.09 | 0.32 ± 0.03 |
| RZ CO2 | 28.38 ± 2.01 | 1.53 ± 0.09 * | 0.32 ± 0.02 | |
| Experiment 5 Lettuce | Control | 46.37 ± 1.45 | 2.48 ± 0.07 | 0.51 ± 0.13 |
| RZ CO2 | 51.94 ± 2.89 | 2.97 ± 0.19 * | 0.42 ± 0.13 | |
| Experiment 6 Lettuce | Control | 62.47 ± 5.19 | 3.01 ± 0.25 | 0.75 ± 0.15 |
| RZ CO2 | 76.34 ± 4.60 | 3.77 ± 0.19 * | 0.72 ± 0.21 | |
| Experiment 7 Lettuce | Control | 71.30 ± 4.28 | 3.40 ± 0.21 | 0.65 ± 0.17 |
| RZ CO2 | 86.86 ± 2.56 ** | 4.17 ± 0.12 ** | 0.56 ± 0.11 |
| Treatment | Shoot Fresh Weight (g) ± SE | Shoot Dry Weight (g) ± SE | Total Leaf Area (cm) ± SE | Root Dry Weight (g) ± SE | |
|---|---|---|---|---|---|
| Experiment 8 Pepper | Control | 1.54 ± 0.09 | 0.27 ± 0.01 | 65 ± 2 | 0.27 ± 0.01 |
| RZ CO2 | 1.60 ± 0.12 | 0.27 ± 0.02 | 71 ± 7 | 0.29 ± 0.01 | |
| Experiment 9 Pepper | Control | nd | 3.12 ± 0.24 | 544 ± 30 | 0.96 ± 0.18 |
| RZ CO2 | nd | 3.30 ± 0.28 | 541 ± 33 | 1.21 ± 0.23 | |
| Experiment 10 Pepper | Control | 9.52 ± 0.38 | 1.10 ± 0.05 | 215 ± 7 | 0.64 ± 0.17 |
| RZ CO2 | 9.44 ± 0.39 | 1.10 ± 0.03 | 215 ± 9 | 0.54 ± 0.07 | |
| Experiment 11 Pepper | Control | 10.50 ± 0.75 | 1.26 ± 0.1 | 276 ± 21 | nd |
| RZ CO2 | 8.47 ± 0.76 | 1.01 ± 0.1 | 219 ± 16 | nd |
| Shoot | Root | Total | ||||
|---|---|---|---|---|---|---|
| RZ CO2 | Control | RZ CO2 | Control | RZ CO2 | Control | |
| mg | ||||||
| Ca | 43.4 ± 5.9 a | 44.8 ± 5.1 a | 4.1 ± 0.6 a | 5.1 ± 1.3 a | 47.5 ± 5.2 a | 49.9 ± 5.4 a |
| K | 237.9 ± 21.3 a | 216.4 ± 18.4 a | 21.7 ± 2.8 a | 24.5 ± 7.8 a | 259.6 ± 21.7 a | 240.9 ± 22.8 a |
| Mg | 14.1 ± 1.0 a | 15.3 ± 0.6 a | 1.2 ± 0.2 a | 1.6 ± 0.4 a | 15.3 ± 0.8 a | 16.9 ± 1 a |
| P | 34.6 ± 2.3 a | 25.6 ± 1.9 b | 6.7 ± 0.9 a | 7.8 ± 2.5 a | 41.3 ± 2.1 a | 33.4 ± 5.1 a |
| S | 15.3 ± 0.6 a | 13.6 ± 0.7 a | 3.5 ± 0.5 a | 4.6 ± 1.4 a | 18.8 ± 0.8 a | 18.2 ± 1.8 a |
| N | 225 ± 6.7 a | 179.9 ± 6.8 b | 26.9 ± 3.7 a | 28.9 ± 9.5 a | 251.9 ± 11.5 a | 208.8 ± 17.8 a |
| µg | ||||||
| B | 84.8 ± 4.2 a | 72.7 ± 12.4 a | 8.7 ± 1.7 a | 10 ± 2.4 a | 93.5 ± 4.8 a | 82.7 ± 13.7 a |
| Cu | 29.9 ± 0.95 a | 24.5 ± 1.5 b | 10.2 ± 1.5 a | 11.5 ± 3.2 a | 40.1 ± 2.4 a | 36 ± 4.6 a |
| Fe | 528.8 ± 25.2 a | 466.8 ± 44.3 a | 1234.1± 207.9 a | 1286.5 ± 371.3 a | 1763.2 ± 321.1 a | 1753.3 ± 241 a |
| Mn | 238.9 ± 22.8 a | 211.6 ± 18.5 a | 110.3± 14.6 a | 127.6 ± 34.3 a | 349.2 ± 28.4 a | 339.2 ± 37.4 a |
| Zn | 436.8 ± 26.4 a | 288.5 ± 43.9 b | 87.7 ± 16.7 a | 92.6 ± 20.1 a | 524.5 ± 42.7 a | 381.1 ± 63.2 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leibar-Porcel, E.; McAinsh, M.R.; Dodd, I.C. Elevated Root-Zone Dissolved Inorganic Carbon Alters Plant Nutrition of Lettuce and Pepper Grown Hydroponically and Aeroponically. Agronomy 2020, 10, 403. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10030403
Leibar-Porcel E, McAinsh MR, Dodd IC. Elevated Root-Zone Dissolved Inorganic Carbon Alters Plant Nutrition of Lettuce and Pepper Grown Hydroponically and Aeroponically. Agronomy. 2020; 10(3):403. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10030403
Chicago/Turabian StyleLeibar-Porcel, Estibaliz, Martin R. McAinsh, and Ian C. Dodd. 2020. "Elevated Root-Zone Dissolved Inorganic Carbon Alters Plant Nutrition of Lettuce and Pepper Grown Hydroponically and Aeroponically" Agronomy 10, no. 3: 403. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10030403