Coronatine Enhances Stalk Bending Resistance of Maize, Thickens the Cell Wall and decreases the Area of the Vascular Bundles

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Experimental Design
2.2. Sampling and Measurements
2.2.1. Morphological Trait and Bending Resistant Strength
Plant Height and Ear Height
Characteristics of the Third Basal Internode
2.2.2. Histological Analysis
2.2.3. Phytohormone Quantification
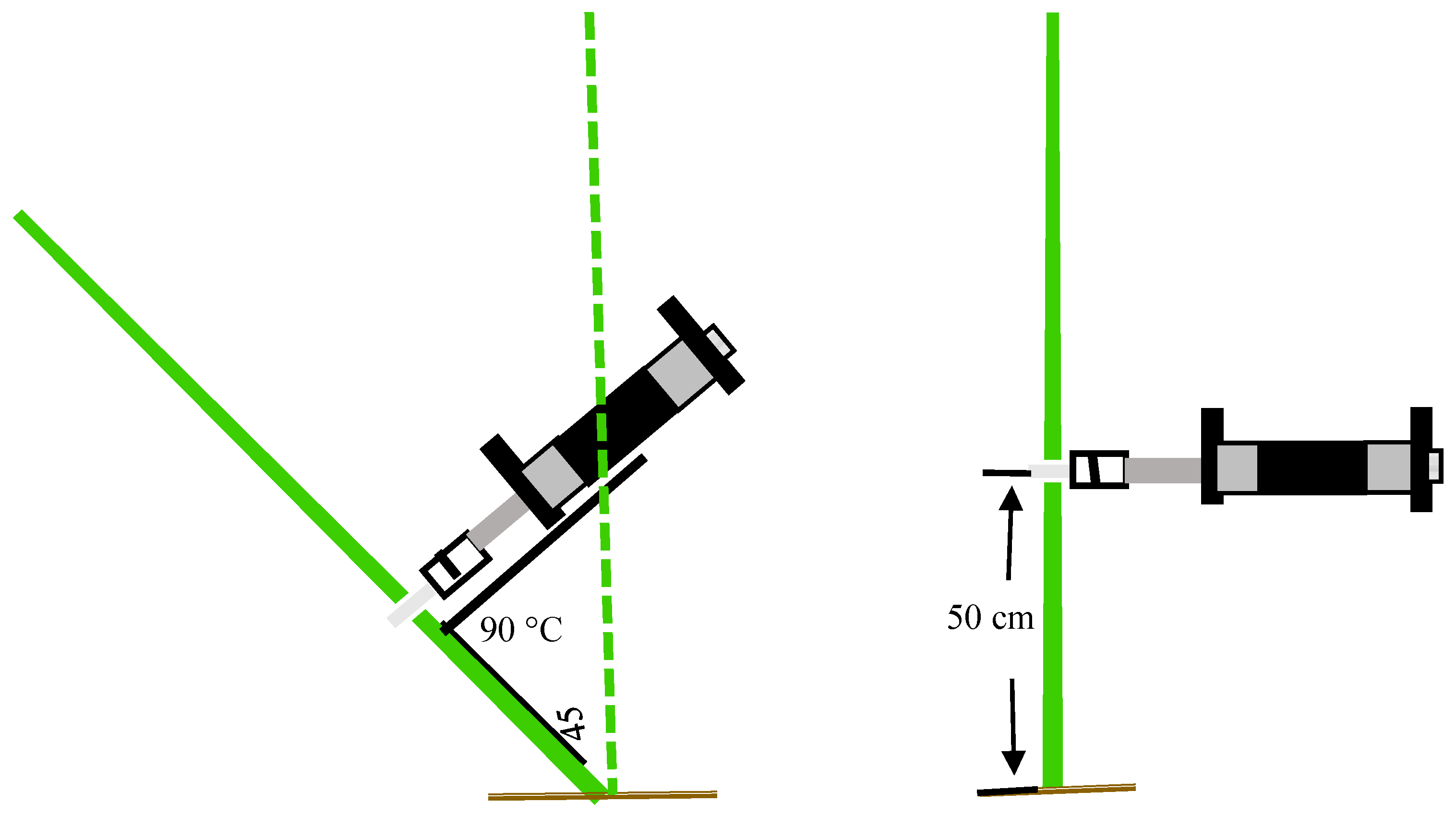
2.2.4. Measurement of Pushing Resistance
2.3. Statistical Analyses
3. Results
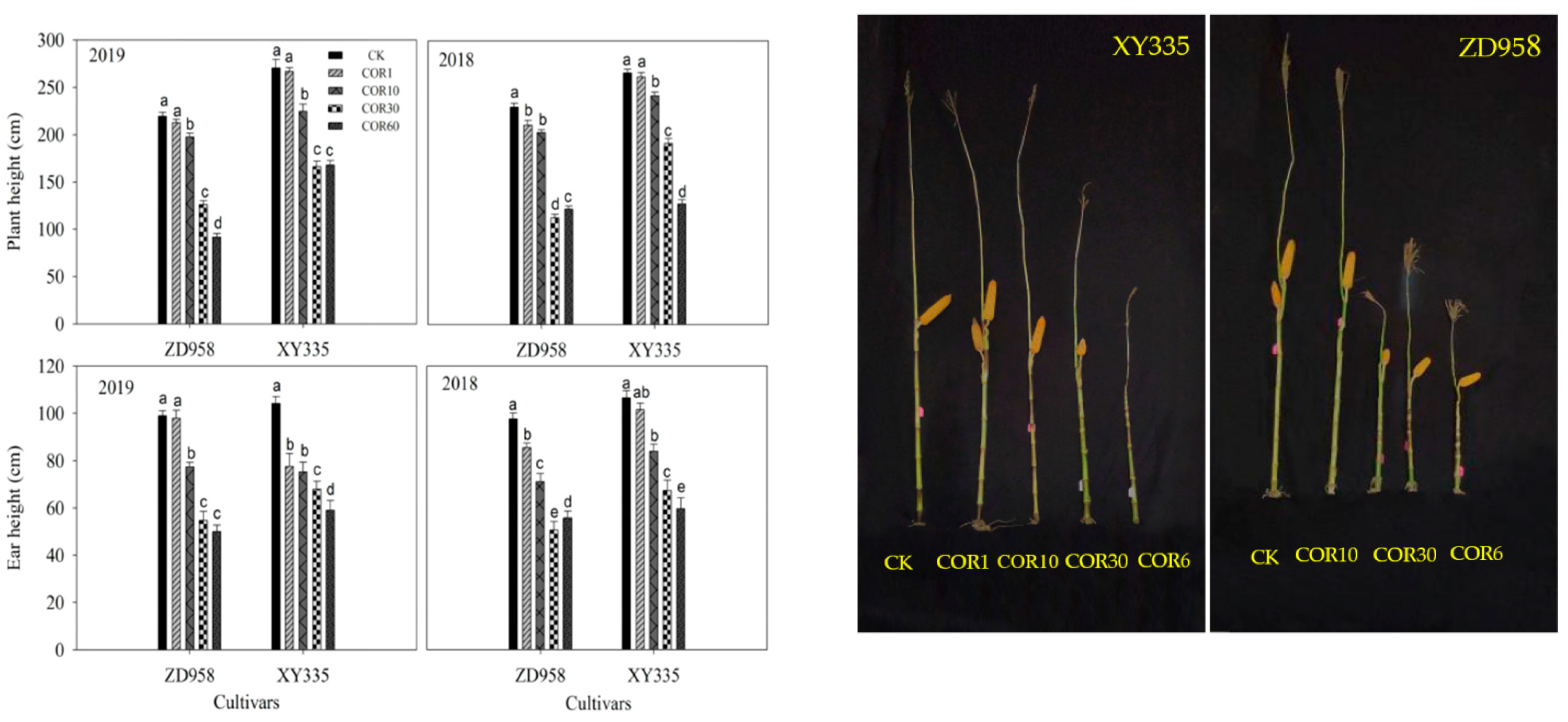
3.1. Plant Height and Ear Height
3.2. Agronomic Traits of the 3rd Basal Internodes
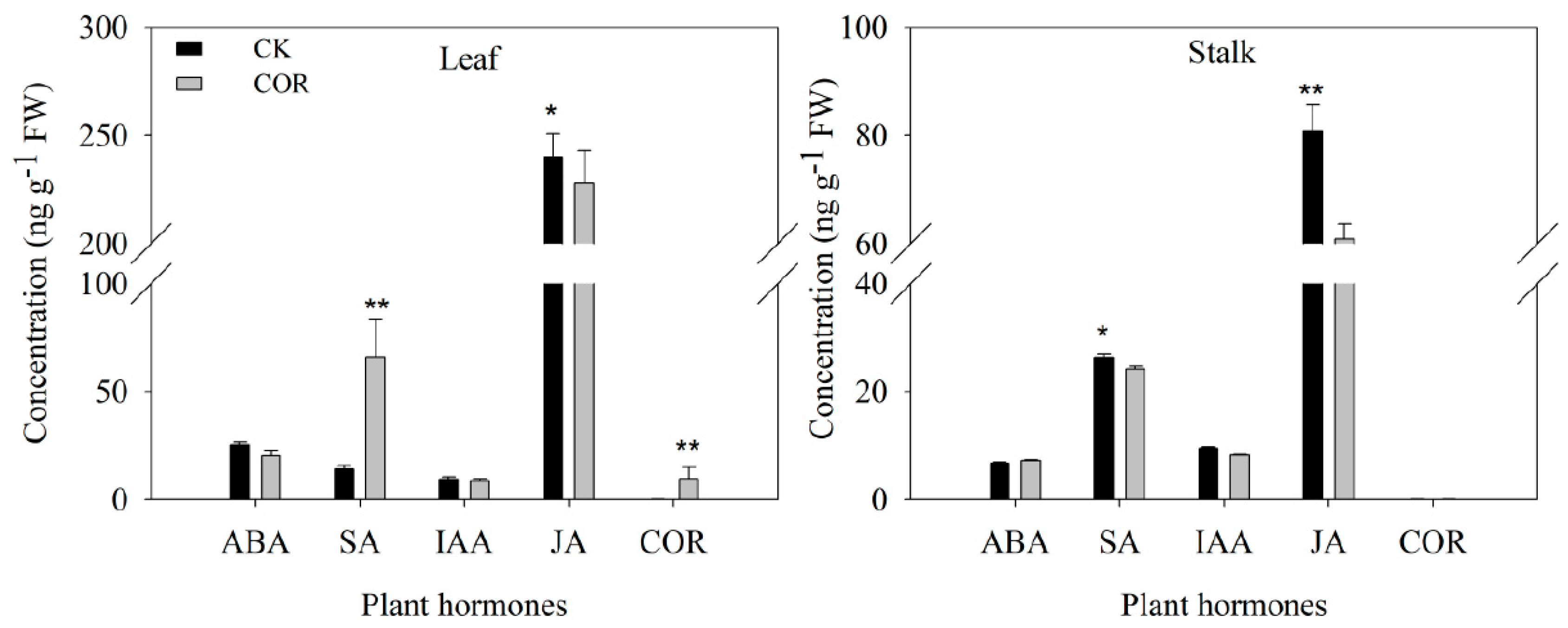
3.3. Detection of Endogenous Hormone Contents
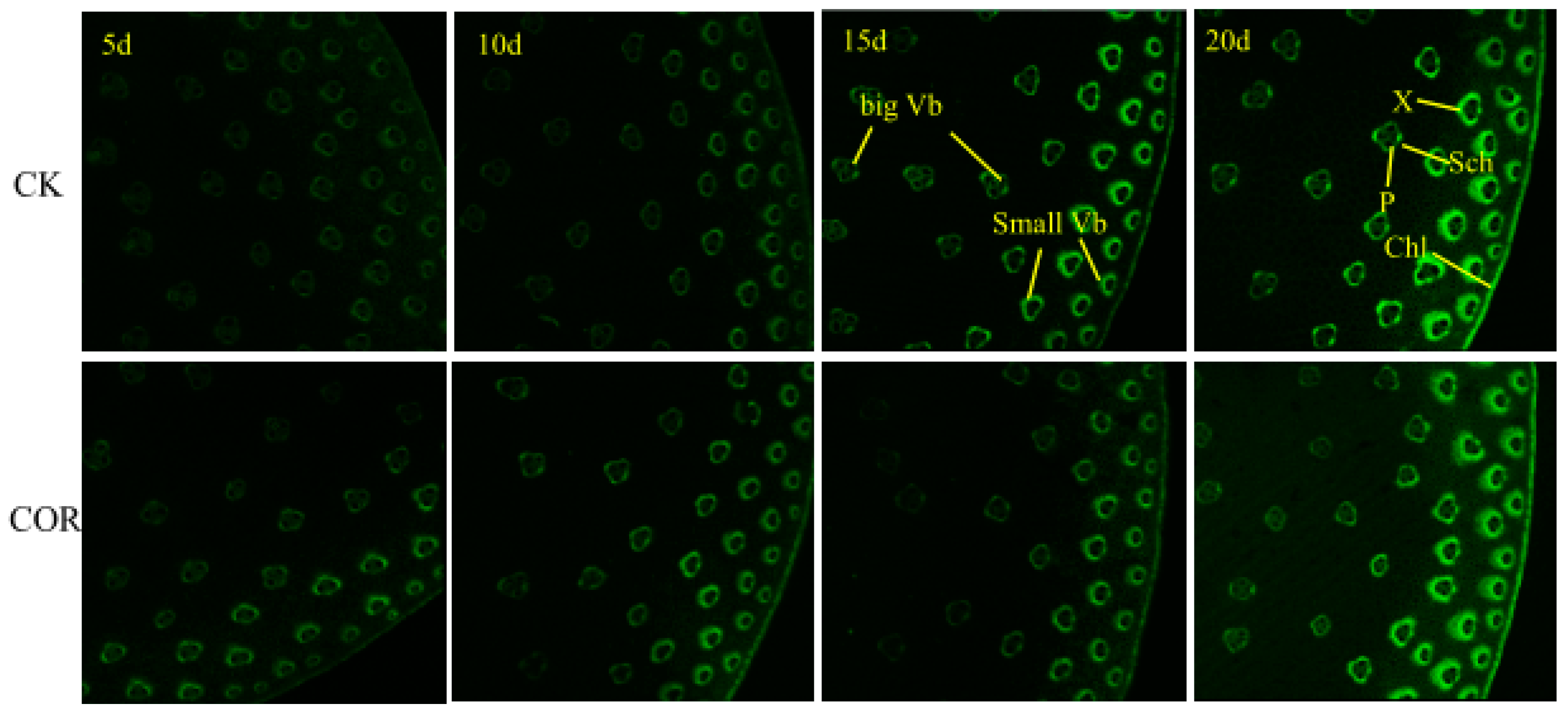
3.4. Microstructure Analysis of Maize Stalk
3.4.1. Lignin Distribution in Stalks Identified by Autofluorescence Microscopy
3.4.2. Scanning Electron Micrographs of Vascular Bundles and Cell Walls
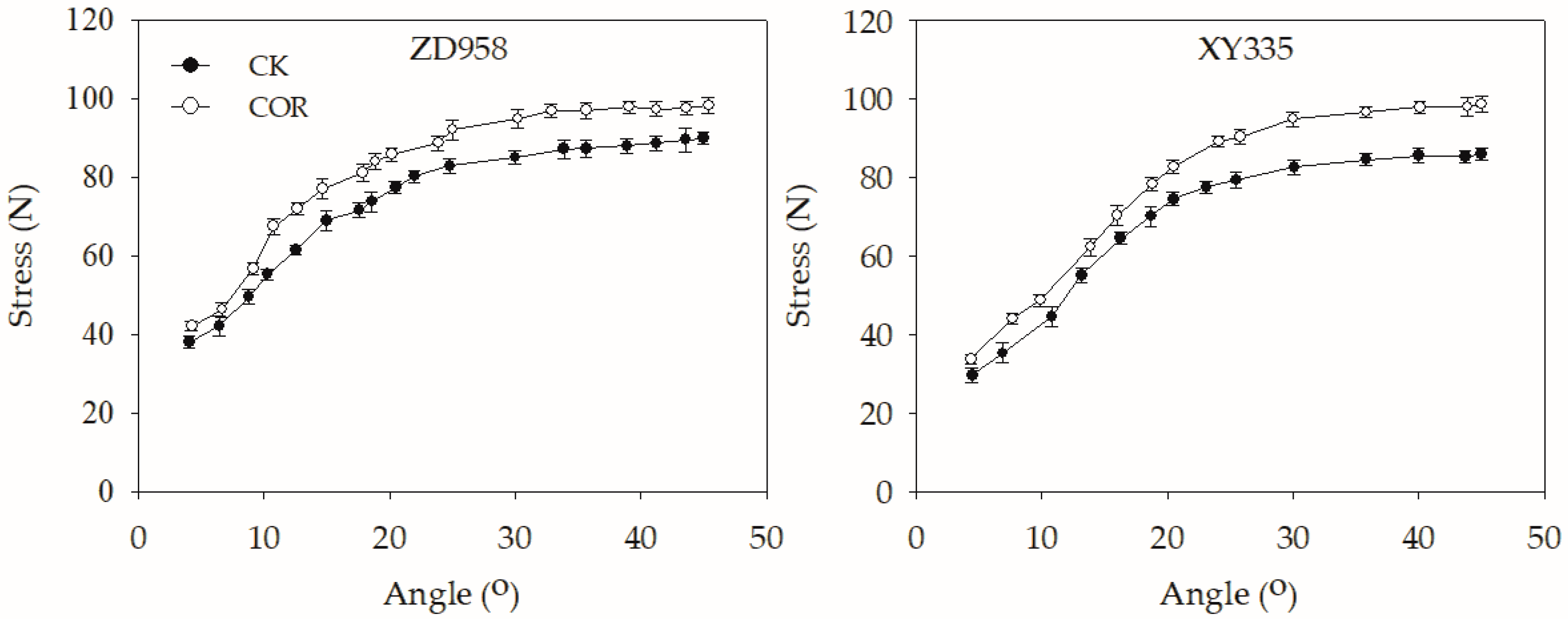
3.5. Pushing Resistance of Maize Stalks
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- FAO. FAOSTAT-Agriculture Database. 2012. Available online: http://faostat.fao.org/ (accessed on 5 May 2020).
- National Bureau of Statistics of China. Chinese Statistical Yearbook; China Statistics: Beijing, China, 2014.
- Norberg, O.S.; Mason, S.C.; Lowry, S.R. Ethephon influence on harvestable yield, grain quality, and lodging of corn. Agron. J. 1988, 80, 768–772. [Google Scholar] [CrossRef]
- Minami, M.; Ujihara, A. Effects of lodging on dry matter production, grain yield and nutritional composition at different growth stages in maize (Zea mays L.). Jpn. J. Crop Sci. 1991, 60, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Gu, W.; Dai, J. The effect of density on lodging of crop. J. Shenyang Agric. Univ. 1989, 4, 413–416. [Google Scholar]
- Ma, D.L.; Xie, R.Z.; Liu, X.; Niu, X.K.; Hou, P.; Wang, K.R.; Lu, Y.L.; Li, S.K. Lodging- related stalk characteristics of maize varieties in china since the 1950. Crop Sci. 2014, 54, 2805–2814. [Google Scholar] [CrossRef]
- Schluttenhofer, C.M.; Massa, G.D.; Mitchell, C.A. Use of uniconazole to control plant height for an industrial/pharmaceutical maize platform. Ind. Crop. Prod. 2011, 33, 720–726. [Google Scholar] [CrossRef]
- Xu, C.L.; Gao, Y.B.; Tian, B.J.; Ren, J.H.; Meng, Q.F.; Wang, P. Effects of EDAH, a novel plant growth regulator, on mechanical strength, stalk vascular bundles and grain yield of summer maize at high densities. Field Crops Res. 2017, 200, 71–79. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, L.; Evers, J.; van der Werf, W.; Zhang, W.; Duan, L. Maize yield and quality in response to plant density and application of anovel plant growth regulator. Field Crops Res. 2014, 164, 82–89. [Google Scholar] [CrossRef]
- Zeng, Q.; Jiang, L.; Wang, D.; Huang, S.; Yang, D. Camptothecin and 10- hydroxycamptothecin accumulation in tender leaves of Camptotheca acuminata saplings after treatment with plant growth regulators. Plant Growth Regul. 2012, 68, 467–473. [Google Scholar] [CrossRef]
- Naeem, M.; Khan, M.M.A.; Moinuddin. Triacontanol: A potent plant growth regulator in agriculture. J. Plant Interact. 2012, 7, 129–142. [Google Scholar] [CrossRef]
- Tripathi, S.C.; Sayre, K.D.; Kaul, J.N.; Narang, R.S. Growth and morphology of spring wheat (Triticum aestivum L.) culms and their association with lodging: Effects of genotypes, N levels and ethephon. Field Crop. Res. 2003, 84, 271–290. [Google Scholar] [CrossRef]
- Ramburan, S.; Greenfield, P.L. Use of ethephon and chlormequat chloride to manage plant height and lodging of irrigated barley (cv. Puma) when high rates of N-fertiliser are applied. S. Afr. J. Plant Soil. 2007, 24, 181–187. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, I.; Kamran, M.; Ali, S.; Bilegjargal, B.; Cai, T.; Ahmad, S.; Meng, X.P.; Su, W.N.; Liu, T.N.; Han, Q.F. Uniconazole application strategies to improve lignin biosynthesis, lodging resistance and production of maize in semiarid regions. Field Crop. Res. 2018, 222, 66–77. [Google Scholar] [CrossRef]
- Ichihara, A.; Shiraishi, K.; Sato, H.; Sakamura, S.; Nishiyama, K.; Sakai, R.; Furusaki, A.; Matsumoto, T. The structure of coronatine. J. Am. Chem. Sot. 1977, 99, 636–637. [Google Scholar] [CrossRef]
- Feys, B.J.F.; Benedetti, C.E.; Penfold, C.N.; Turner, J.G. Arabidopsis mutants selected for resistance to the phytotoxin coronatine are male sterile, insensitive to methyl jasmonate, and resistant to a bacterial pathogen. Plant Cell. 1994, 6, 751–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staswick, P.E.; Tiryaki, I. The oxylipin signal jasmonic acid is activated by an enzyme that conjugates it to isoleucine in Arabidopsis. Plant Cell. 2004, 16, 2117–2127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dittrich, H.; Kutchan, T.M.; Zenk, M.H. The jasmonate precursor, 12-oxo-phytodienoic acid, induces phytoalexin synthesis in Petroselinum crispum cell cultures. FEBS Lett. 1992, 309, 33–36. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.B.; Zhang, C.; Gu, M.; Bai, Z.Y.; Zhang, W.G.; Qi, T.C.; Cheng, Z.W.; Peng, W.; Luo, H.B.; Nan, F.J.; et al. The arabidopsis CORONATINE INSENSITIVE1 protein is a jasmonate receptor. Plant Cell. 2009, 21, 2220–2236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Z.X.; Duan, L.S.; Li, Z.H.; Wang, X.D.; Liu, X.J. Dose-dependent effects of coronatine on cotton seedling growth under salt stress. J. Plant Growth Regul. 2015, 34, 651–664. [Google Scholar] [CrossRef]
- Zhou, Y.Y.; Liu, Y.R.; Peng, C.X.; Li, X.W.; Zhang, M.C.; Tian, X.L.; Li, J.M.; Li, Z.H.; Duan, L.S. Coronatine enhances drought tolerance in winter wheat by maintaining high photosynthetic performance. J. Plant Physiol. 2018, 228, 59–65. [Google Scholar] [CrossRef]
- Bender, C.L.; Alarcon, C.F.; Gross, D.C. Pseudomonas syringae phytotoxins: Mode of action, regulation and biosynthesis by peptide and polypeptide synthetases. Microbiol. Mol. Biol. Rev. 1999, 63, 266–292. [Google Scholar] [CrossRef] [Green Version]
- Melotto, M.; Underwood, W.; He, S.Y. Role of stomata in plant innate immunity and foliar bacterial disease. Ann. Rev. Phytopathol. 2008, 46, 101–122. [Google Scholar] [CrossRef] [Green Version]
- Kenyon, J.S.; Turner, J.G. Physiological changes in nicotiana taba-cum leaves during development of chlorosis caused by coronatine. Physiol. Mol. Plant Pathol. 1990, 37, 463–477. [Google Scholar] [CrossRef]
- Kenyon, J.S.; Turner, J.G. The stimulation of ethylene synthesis in nicotiana tabacum leaves by the phytotoxin coronatine. Plant Physiol. 1992, 100, 219–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schüler, G.; Mithöfer, A.; Baldwin, I.T.; Berger, S.; Ebel, J.; Santos, J.G.; Herrmann, G.; Hölscher, D.; Kramell, R.; Kutchan, T.M.; et al. Coronalon: A powerful, tool in plant stress physiology. EFBS Lett. 2004, 563, 17–22. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.X.; Duan, L.S.; Tian, X.L.; Wang, B.M.; Eneji, A.E.; Li, Z.H. Corontine alleviates salinity stress in cotton by improving the antioxidative defense system and radical-scavenging activity. J. Plant Physiol. 2008, 165, 375–384. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Yang, F.Q.; Li, B.; Eneji, A.E.; Li, J.M.; Duan, L.S.; Wang, B.M.; Li, Z.H.; Tian, X.L. Coronatine-induced lateral-root formation in cotton (Gossypium hirsutum) seedlings under potassium-sufficient and deficient conditions in relation to auxin. J. Plant Nutr. Soil Sci. 2009, 172, 435–444. [Google Scholar] [CrossRef]
- Geng, X.Q.; Jin, L.; Shimada, M.; Kim, M.G.; Mackey, D. The phytotoxin coronatine is a multifunctional component of the virulence armament of Pseudomonas syringae. Planta 2014, 240, 1149–1165. [Google Scholar] [CrossRef] [Green Version]
- Uppalapati, S.R.; Ayoubi, P.; Weng, H.; Palmer, D.A.; Mitchell, R.E.; Jones, W.; Bender, C.L. The phytotoxin coronatine and methyl jasmonate impact multiple phytohormone pathways in tomato. Plant J. 2005, 42, 201–217. [Google Scholar] [CrossRef]
- Peiffer, J.A.; Flint-Garcia, S.A.; Leon, N.D.; McMullen, M.D.; Kaeppler, S.M.; Buckler, E.S. The genetic architecture of maize stalk strength. PLoS ONE 2013, 8, e67066. [Google Scholar] [CrossRef] [Green Version]
- Baker, C.J.; Sterling, M.; Berry, P. A generalised model of crop lodging. J. Theor. Biol. 2014, 363, 1–12. [Google Scholar] [CrossRef]
- Bosch, M.; Mayer, C.D.; Cookson, A.; Donnison, I.S. Identification of genes involved in cell wall biogenesis in grasses by differential gene expression profiling of elongating and non-elongating maize internodes. J. Exp. Bot. 2011, 62, 3545–3561. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, G.H.; Song, Y.P.; Liu, Z.H.; Yang, C.D.; Tang, S.; Zheng, C.Y.; Wang, S.H.; Ding, Y.F. Lodging resistance characteristics of high-yielding rice populations. Field Crops Res. 2014, 161, 64–74. [Google Scholar] [CrossRef]
- Flint-Garcia, S.A.; Jampatong, C.; Darrah, L.L.; McMullen, M.D. Quantitative trait locus analysis of stalk strength in four maize populations. Crop Sci. 2003, 43, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Gou, L.; Huang, J.J.; Sun, R.; Ding, Z.S.; Dong, Z.Q.; Zhao, M. Variation characteristic of stalk penetration strength of maize with different density tolerance varieties. Trans. CSAE 2010, 26, 156–162. [Google Scholar] [CrossRef]
- Wang, C.; Ruan, R.W.; Yuan, X.H.; Hu, D.; Yang, H.; Li, Y.; Yi, Z.L. Effects of nitrogen fertilizer and planting density on the lignin synthesis in the culm in relation to lodging resistance of buckwheat. Plant Prod. Sci. 2015, 18, 218–227. [Google Scholar] [CrossRef] [Green Version]
- Zheng, M.J.; Chen, J.; Shi, Y.H.; Li, Y.X.; Yin, Y.P.; Yang, D.Q.; Luo, Y.L.; Pang, D.W.; Xu, X.; Li, W.Q.; et al. Manipulation of lignin metabolism by plant den-sities and its relationship with lodging resistance in wheat. Sci. Rep. 2017, 7, 41805. [Google Scholar] [CrossRef]
- Peng, D.L.; Chen, X.G.; Yin, Y.P.; Lu, K.L.; Yang, W.B.; Tang, Y.H.; Wang, Z.L. Lodging resistance of winter wheat (Triticum aestivum L.): Lignin accumulation and its related enzymes activities due to the application of paclobutrazol or gibberellin acid. Field Crops Res. 2014, 157, 1–7. [Google Scholar] [CrossRef]
- Chen, X.G.; Shi, C.Y.; Yin, Y.P.; Wang, Z.L.; Shi, Y.H.; Peng, D.L.; Ni, Y.L.; Cai, T. Relationship between lignin metabolism and lodging resistance in wheat. Acta Agron. Sin. 2011, 37, 1616–1622. [Google Scholar] [CrossRef]
- Huang, J.L.; Liu, W.Y.; Zhou, F.; Peng, Y.J.; Wang, N.L. Mechanical properties of maize fibre bundles and their contribution to lodging resistance. Biosyst. Eng. 2016, 151, 298–307. [Google Scholar] [CrossRef]
- Wang, N.L.; Liu, W.Y.; Peng, Y. Gradual transition zone between cell wall layers and its influence on wood elastic modulus. J. Mater. Sci. 2013, 48, 5071–5084. [Google Scholar] [CrossRef]
- Donaldson, L.A.; Knox, J.P. Localization of cell wall polysaccharides in normal and compression wood of radiata pine: Relationships with lignification and microfibril orientation. Plant Physiol. 2012, 158, 642–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, K.; Fukushima, K. Distribution of lignin interunit bonds in the differentiating xylem of compression and normal woods of Pinus thunbergii. J. Wood Sci. 2005, 51, 246–251. [Google Scholar] [CrossRef]
- Kashiwagi, T.; Ishimaru, K. Identification and functional analysis of a locus for improvement of lodging resistance in rice. Plant Physiol. 2004, 134, 676–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, J.; Xie, R.Z.; Zhang, W.F.; Wang, K.R.; Hou, P.; Ming, B.; Gou, L.; Li, S.K. Research progress on reduced lodging of high-yield and -density maize. J. Integr. Agric. 2017, 16, 2717–2725. [Google Scholar] [CrossRef]
- Kokubo, A.; Kuraishi, S.; Sakurai, N. Culm strength of barley, correlation among maximum bending stress, cell wall dimensions, and cellulose content. Plant Physiol. 1989, 91, 876–882. [Google Scholar] [CrossRef] [Green Version]
- Zhong, R.Q.; Taylor, J.J.; Ye, Z.H. Disruption of interfascicular fiber differentiation in a Arabidopsis mutant. Plant Cell. 1997, 9, 2159–2170. [Google Scholar] [CrossRef] [Green Version]
- Kaack, K.; Schwarz, K.U.; Brander, P.E. Variation in morphology, anatomy and chemistry of stems of Miscanthus genotypes differing in mechanical properties. Ind. Crops Prod. 2003, 17, 131–142. [Google Scholar] [CrossRef]
- Weng, F.; Zhang, W.; Wu, X.; Xu, X.; Ding, Y.; Li, G.; Liu, Z.; Wang, S. Impact of low-temperature, overcast and rainy weather during the reproductive growth stage on lodging resistance of rice. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef]
- Khanna, V.K. Relationship of lodging resistance and yield to anatomical characters of stem in wheat, triticale and rye. Wheat Inf. Serv. 1991, 73, 19–24. [Google Scholar]
- Zuber, U.; Winzeler, H.; Messmer, M.M.; Keller, M.; Keller, B.; Schmid, J.E.; Stamp, P. Morphological traits associated with lodging resistance of spring wheat (Triticum aestivum L.). J. Agron. Crop Sci. 1999, 182, 17–24. [Google Scholar] [CrossRef]
- Dunn, G.; Briggs, K.G. Variation in culm anatomy among barley cultivars differing in lodging resistance. Can. J. Bot. 1989, 67, 1838–1843. [Google Scholar] [CrossRef]
- Mwaikambo, L.Y. Tensile properties of alkalised jute fibres. Bioresource 2009, 4, 566–588. [Google Scholar]
- Esau, K. Anatomy of Seed Plants, 2nd ed.; Wiley: New York, NY, USA, 1977. [Google Scholar]
- Okuno, A.; Hirano, K.; Asano, K.; Takase, W.; Masuda, R.; Morinaka, Y.; Ueguchi-Tanaka, M.; Kitano, H.; Matsuoka, M. New approach to increasing rice lodging resistance and biomass yield through the use of high gibberellin producing varieties. PLoS ONE 2014, 9, e86870. [Google Scholar] [CrossRef] [PubMed]
- Didi, V.; Jackson, P.; Hejatko, J. Hormonal regulation of secondary cell wall for-mation. J. Exp. Bot. 2015, 66, 5015–5027. [Google Scholar] [CrossRef]
- Lin, D.S.; Cao, L.Y.; Zhou, Z.Z.; Zhu, L.; Ehrhardt, D.; Yang, Z.B.; Fu, Y. Rho GTPase signaling activates microtubule severing to promote microtubule ordering in Arabidopsis. Curr. Biol. 2013, 23, 290–297. [Google Scholar] [CrossRef] [Green Version]
- Cecchetti, V.; Altamura, M.M.; Brunetti, P.; Petrocelli, V.; Falasca, G.; Ljung, K.; Costantino, P.; Cardarelli, M. Auxin controls Arabidopsis anther dehiscence by regulating endothecium lignification and jasmonic acid biosynthesis. Plant J. 2013, 74, 411–422. [Google Scholar] [CrossRef]
- Vanacker, H.; Lu, H.; Rate, D.N.; Greenberg, J.T. A role for salicylic acid and NPR1 in regulating cell growth in Arabidopis. Plant J. 2001, 28, 209–216. [Google Scholar] [CrossRef]
- Swiatek, A.; Lenjou, M.; Van, B.D.; Dirk, I.; Harry, V.O. Differential Effect of Jasmonic Acid and Abscisic Acid on Cell Cycle Progression in Tobacco BY-2 Cells. Plant Physiol. 2002, 128, 201–211. [Google Scholar] [CrossRef]
- Liu, Y.R.; Zhou, Y.Y.; Huang, G.M.; Zhu, N.N.; Li, Z.H.; Zhang, M.C.; Duan, L.S. Coronatine inhibits mesocotyl elongation by promoting ethylene production in etiolated maize seedlings. Plant Growth Regul. 2019, 90, 51–61. [Google Scholar] [CrossRef]
- Shibaoka, H. Plant hormone-induced changes in the orientation of cortical microtubules- alterations in the cross-linking between microtubules and the plasma-membrane. Annu. Rev. Plant Physiol. Plant Mol. Bio. 1994, 45, 527–544. [Google Scholar] [CrossRef]
- Kalve, S.; Fotschki, J.; Beeckman, T.; Vissenberg, K.; Beemster, G.T.S. Three-di-mensional patterns of cell division and expansion throughout the development of Arabidopsis thaliana leaves. J. Exp. Bot. 2014, 65, 6385–6397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosgrove, D.J. Expansive growth of plant cell walls. Plant Physiol. Biochem. 2000, 38, 109–124. [Google Scholar] [CrossRef]
- You, T.T.; Mao, J.Z.; Yuan, T.Q.; Wen, J.L.; Xu, F. Structural elucidation of the lignins from stems and foliage of arundo donax Linn. J. Agric. Food Chem. 2013, 61, 5361–5370. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Years | Cultivers | Treatment | Length (cm) | Diameter (mm) | Volume (mL) | Cross Section Area (cm2) | Fresh Weight (g) | Fresh Weight Density (g cm−3) | Dry Weight (g) | Dry Weight per cm (g cm−1) | Ring Puncture Strength (Nmm−2) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2019 | ZD958 | Control | 11.8 ± 0.8a | 25.7 ± 0.3c | 55.6 ± 1.2b | 4.73 ± 0.5c | 51.5 ± 1.3c | 0.93 ± 0.03c | 8.36 ± 0.5b | 0.71 ± 0.04b | 52.2 ± 4.2c |
| COR1 | 11.8 ± 1.0a | 28.0 ± 0.5a | 61.0 ± 2.8a | 5.22 ± 0.8a | 54.8 ± 2.5b | 0.90 ± 0.04c | 8.00 ± 0.7b | 0.74 ± 0.02b | 60.1 ± 3.8a | ||
| COR10 | 9.52 ± 0.9c | 28.9 ± 0.2a | 51.0 ± 1.3c | 5.06 ± 0.4b | 48.4 ± 3.0d | 0.95 ± 0.06c | 7.20 ± 0.4c | 0.75 ± 0.05b | 55.2 ± 5.5b | ||
| COR30 | 9.46 ± 0.7c | 25.5 ± 0.6c | 42.4 ± 0.9c | 4.43 ± 0.3d | 44.8 ± 1.9e | 1.06 ± 0.04b | 6.82 ± 0.6d | 0.76 ± 0.07b | 51.4 ± 4.6c | ||
| COR60 | 10.8 ± 1.2b | 27.8 ± 1.2b | 53.8 ± 1.5bc | 5.03 ± 0.2b | 58.0 ± 4.5a | 1.08 ± 0.02a | 10.5 ± 0.3a | 0.97 ± 0.04a | 50.5 ± 3.7c | ||
| XY335 | Control | 14.7 ± 0.5b | 24.3 ± 0.4c | 59.8 ± 2.4d | 4.33 ± 0.7c | 51.4 ± 2.8c | 0.86 ± 0.06b | 9.96 ± 0.6d | 0.68 ± 0.02c | 58.0 ± 2.6b | |
| COR1 | 15.6 ± 0.7a | 26.4 ± 0.6a | 76.3 ± 1.5a | 6.24 ± 0.3a | 60.5 ± 3.2a | 0.80 ± 0.07c | 10.1 ± 0.7c | 0.71 ± 0.01c | 60.0 ± 6.2a | ||
| COR10 | 13.2 ± 0.6c | 26.8 ± 1.2a | 64.8 ± 2.0bc | 4.89 ± 1.1b | 57.1 ± 1.7b | 0.89 ± 0.05b | 10.8 ± 0.5c | 0.82 ± 0.06b | 61.7 ± 5.2a | ||
| COR30 | 13.2 ± 1.1c | 25.1 ± 0.8b | 65.8 ± 1.4b | 4.97 ± 0.5b | 57.8 ± 3.4b | 0.87 ± 0.07b | 11.7 ± 0.6b | 0.89 ± 0.04a | 58.0 ± 1.9b | ||
| COR60 | 13.5 ± 0.9c | 24.9 ± 1.1bc | 62.0 ± 2.8c | 4.90 ± 0.8b | 57.3 ± 4.1b | 0.92 ± 0.06a | 12.6 ± 0.9a | 0.93 ± 0.06a | 41.9 ± 6.4c | ||
| Source of variation | C | ** | * | ** | ** | ** | ** | ** | ** | ** | |
| T | ** | ** | ** | ** | ** | ** | ** | ** | ** | ||
| C×T | ** | ** | ** | ** | ** | ** | ** | ** | ** | ||
| 2018 | ZD958 | Control | 10.3 ± 0.6a | 26.0 ± 1.2b | 46.4 ± 4.2c | 4.48 ± 0.3b | 45.5 ± 2.7a | 0.95 ± 0.02b | 7.56 ± 0.5bc | 0.73 ± 0.03b | 45.4 ± 3.3b |
| COR1 | 10.6 ± 0.8a | 26.4 ± 0.5b | 48.8 ± 1.8a | 4.68 ± 0.6b | 45.0 ± 3.0d | 0.92 ± 0.04b | 8.63 ± 0.4a | 0.81 ± 0.02a | 46.9 ± 4.0a | ||
| COR10 | 9.20 ± 1.0b | 28.3 ± 0.8a | 47.2 ± 0.9b | 5.13 ± 0.4a | 47.9 ± 1.8ab | 1.01 ± 0.06a | 7.56 ± 0.6bc | 0.82 ± 0.05a | 45.9 ± 1.9b | ||
| COR30 | 6.58 ± 1.2c | 25.5 ± 1.4c | 34.8 ± 1.0d | 5.16 ± 0.5c | 29.5 ± 4.5e | 0.85 ± 0.08c | 6.46 ± 0.5c | 0.97 ± 0.06b | 40.3 ± 2.4c | ||
| COR60 | 10.2 ± 0.7a | 25.3 ± 0.6c | 46.0 ± 1.7c | 4.48 ± 0.8b | 46.9 ± 1.6c | 1.02 ± 0.06a | 7.80 ± 0.7b | 0.76 ± 0.08b | 26.5 ± 6.3d | ||
| XY335 | Control | 14.7 ± 0.8a | 24.5 ± 0.9b | 66.4 ± 2.9a | 4.51 ± 0.3b | 52.3 ± 2.6c | 0.80 ± 0.04c | 10.3 ± 0.5bc | 0.70 ± 0.09c | 55.4 ± 7.5b | |
| COR1 | 14.6 ± 1.0a | 23.5 ± 0.5c | 63.4 ± 3.5b | 4.32 ± 0.5d | 50.5 ± 3.4d | 0.81 ± 0.03b | 10.7 ± 0.4b | 0.75 ± 0.04b | 59.7 ± 6.4a | ||
| COR10 | 13.8 ± 0.4a | 25.4 ± 0.3a | 63.8 ± 5.0b | 4.61 ± 0.4a | 53.8 ± 1.6b | 0.84 ± 0.05b | 10.1 ± 0.6c | 0.73 ± 0.06c | 56.2 ± 2.9b | ||
| COR30 | 13.9 ± 0.9b | 24.2 ± 0.7bc | 61.8 ± 3.0c | 4.44 ± 0.6c | 52.3 ± 2.4c | 0.85 ± 0.04b | 9.76 ± 0.3d | 0.72 ± 0.04c | 48.7 ± 4.2c | ||
| COR60 | 13.7 ± 0.6b | 24.7 ± 0.6b | 60.0 ± 1.4d | 4.39 ± 0.5d | 54.5 ± 3.3a | 0.91 ± 0.06a | 11.9 ± 0.2a | 0.88 ± 0.05a | 14.5 ± 3.1d | ||
| Source of variation | C | ** | * | ** | ** | ** | ** | ** | ** | ** | |
| T | ** | ** | ** | * | ** | ** | ** | ** | ** | ||
| C×T | ** | ** | ** | * | ** | ** | ** | ** | ** | ||
| Areas (µm2) | Treatments | 5d | 10d | 15d | 20 |
|---|---|---|---|---|---|
| Vascular bundle | CK | 35,443.2 ± 102.7 | 50,076.9 ± 149.2 | 39,799.2 ± 450.6 | 50,822.6 ± 550.9 |
| C | 28,436.4 ± 203.5 | 30,795.6 ± 198.4 | 38,761.9 ± 342.6 | 38,754.2 ± 489.2 | |
| Protoxylem vessel | CK | 4012.6 ± 198.4 | 10,346.5 ± 368.6 | 71,41.6 ± 185.4 | 13,479.9 ± 358.2 |
| COR | 2407.1 ± 105.2 | 3867.4 ± 282.4 | 6487.5 ± 196.7 | 1885.4 ± 102.3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Huang, G.; Guo, Y.; Zhou, Y.; Duan, L. Coronatine Enhances Stalk Bending Resistance of Maize, Thickens the Cell Wall and decreases the Area of the Vascular Bundles. Agronomy 2020, 10, 807. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10060807
Li Y, Huang G, Guo Y, Zhou Y, Duan L. Coronatine Enhances Stalk Bending Resistance of Maize, Thickens the Cell Wall and decreases the Area of the Vascular Bundles. Agronomy. 2020; 10(6):807. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10060807
Chicago/Turabian StyleLi, Yanxia, Guanmin Huang, Yuling Guo, Yuyi Zhou, and Liusheng Duan. 2020. "Coronatine Enhances Stalk Bending Resistance of Maize, Thickens the Cell Wall and decreases the Area of the Vascular Bundles" Agronomy 10, no. 6: 807. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10060807