Successive Harvests Affect Yield, Quality and Metabolic Profile of Sweet Basil (Ocimum basilicum L.)

 , ,
, ,  , and
, and 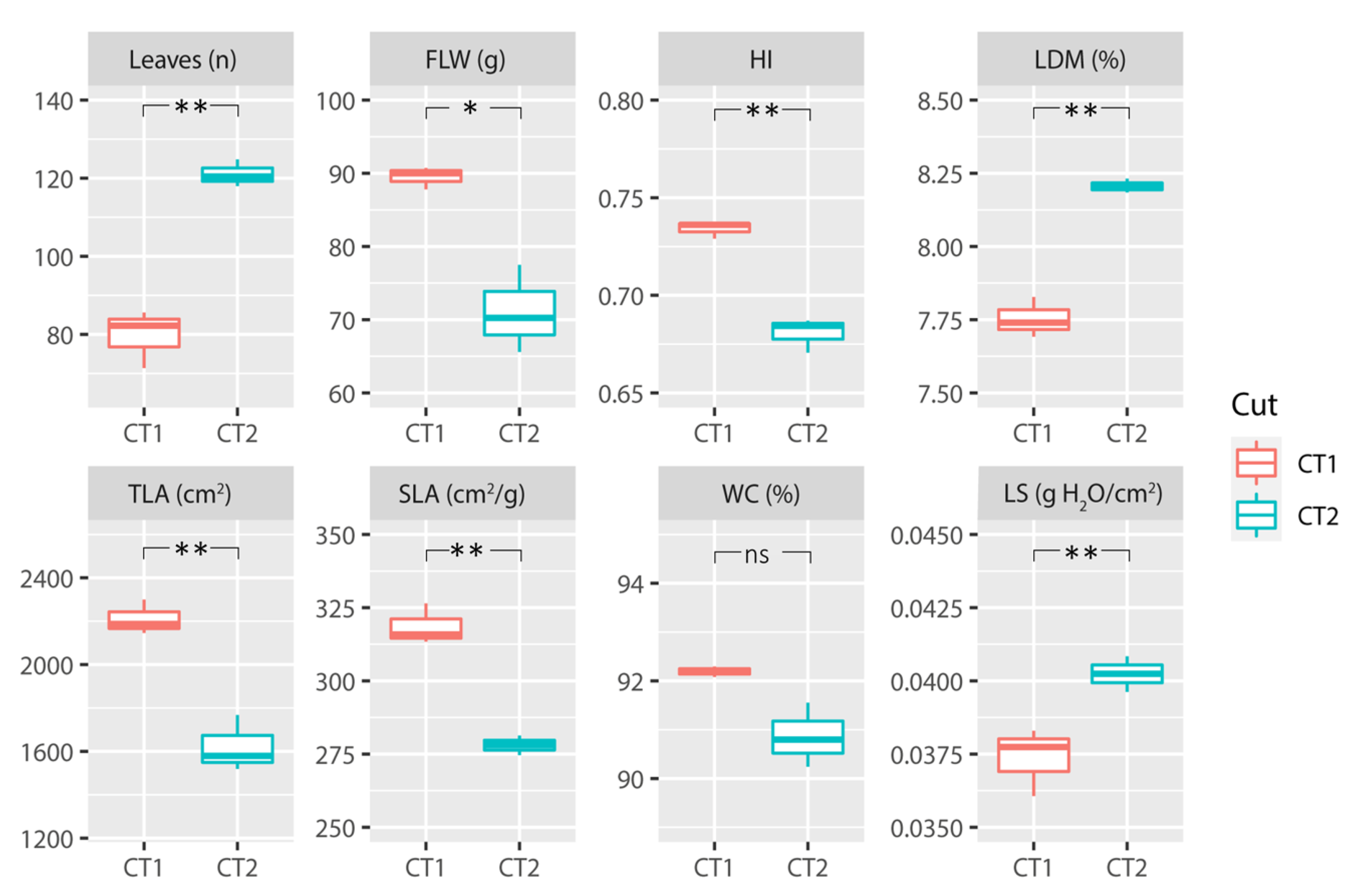{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Experimental Design and Conditions
2.2. Harvest and Growth Analysis
2.3. Leaf Gas Exchange
2.4. Nitrogen Determination and Nutritional Efficiency Indices
2.5. Antioxidant Capacity and Polyphenol Quantification
2.6. Metabolomics
2.7. Statistics
3. Results
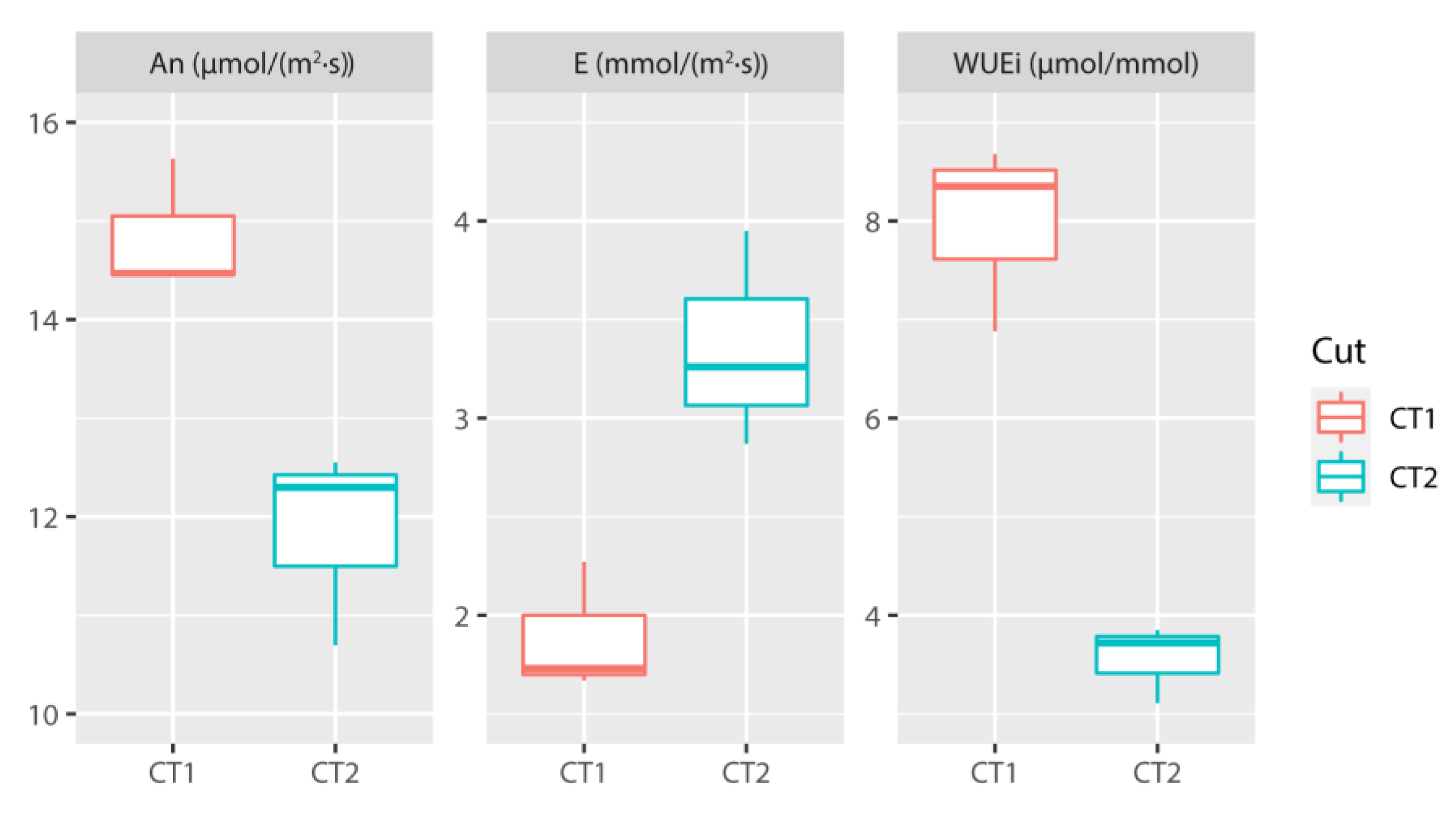
3.1. Analysis of Agronomic and Physiological Parameters
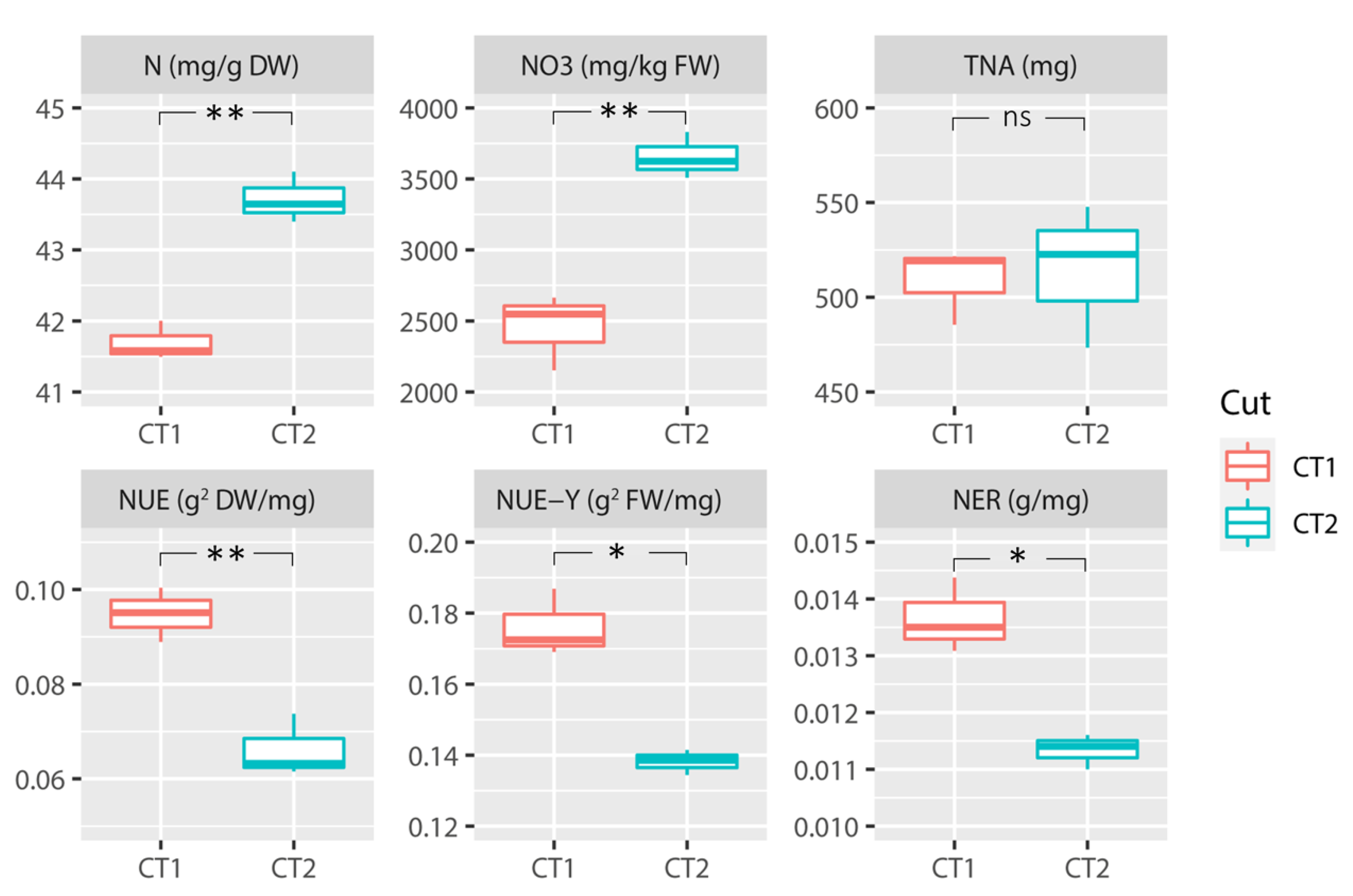
3.2. Nitrogen Accumulation
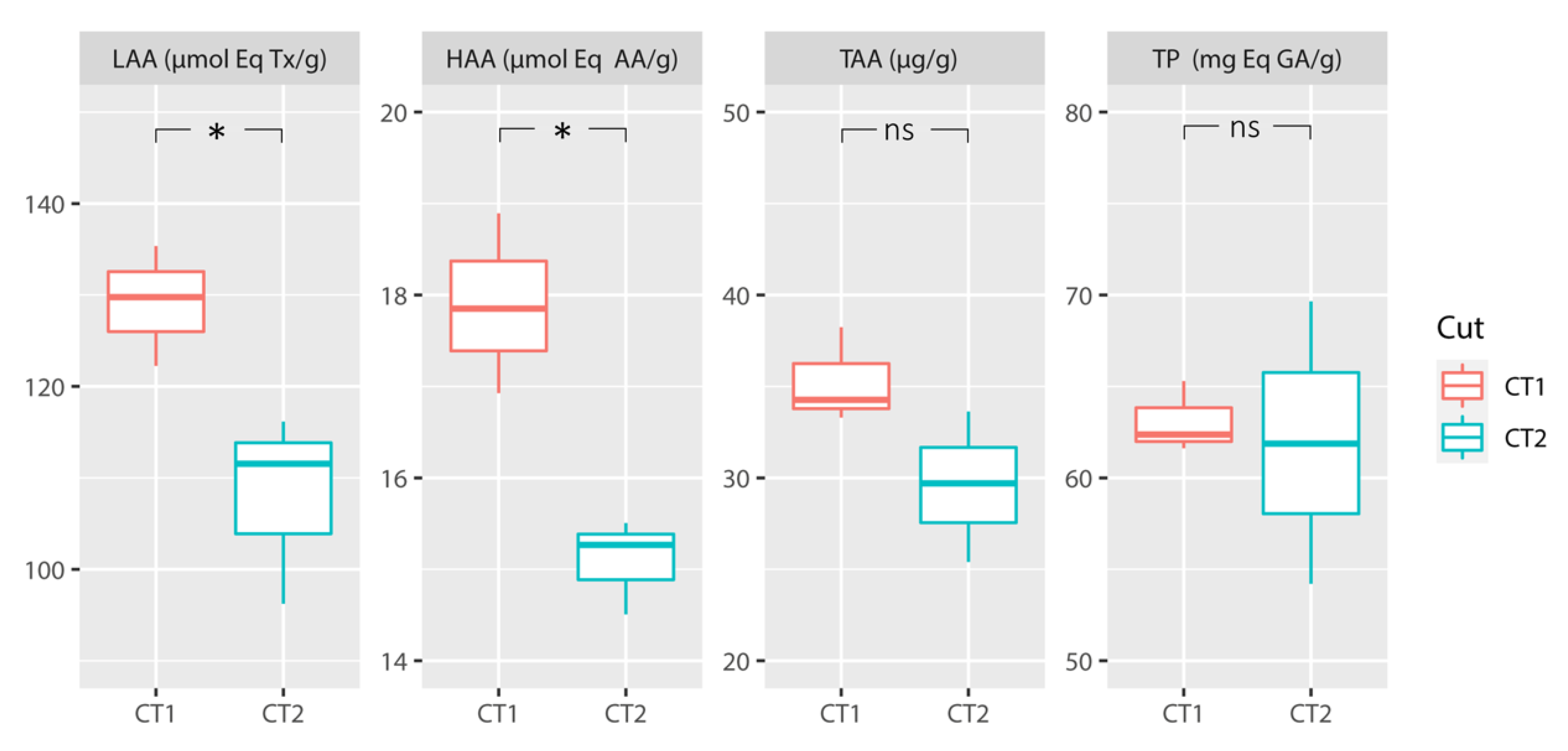
3.3. Antioxidants
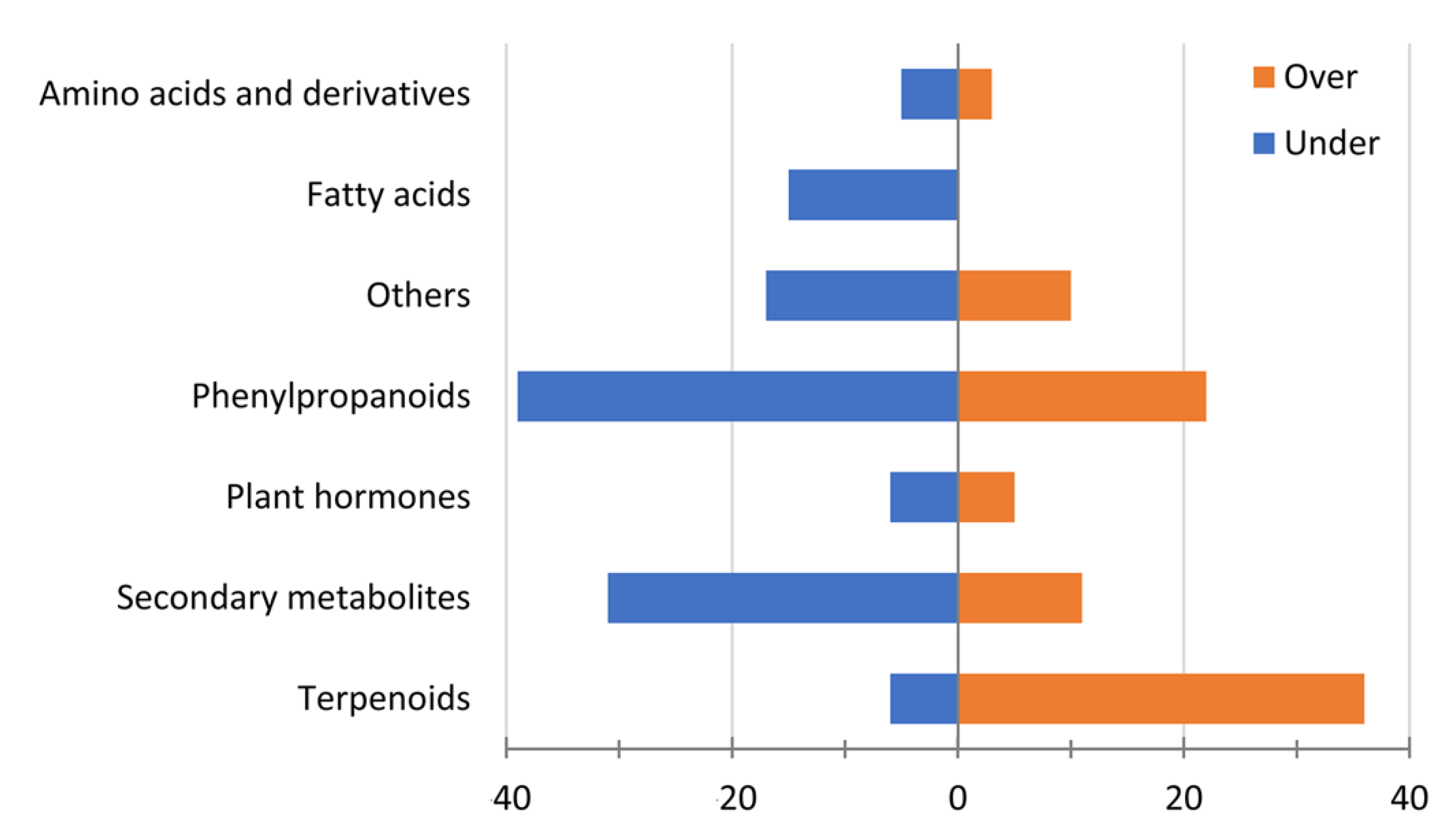
3.4. Metabolomics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hiltunen, R.; Holm, Y. Basil: The Genus Ocimum; CRC Press: Boca Raton, FL, USA, 1999. [Google Scholar]
- Acharya, T.P.; Reiter, M.S.; Welbaum, G.; Arancibia, R.A. Nitrogen Uptake and Use Efficiency in Sweet Basil Production under Low Tunnels. HortScience 2020, 55, 429–435. [Google Scholar] [CrossRef] [Green Version]
- Makri, O.; Kintzios, S. Ocimum sp. (basil): Botany, cultivation, pharmaceutical properties, and biotechnology. J. Herbs Spices Med. Plants 2008, 13, 123–150. [Google Scholar] [CrossRef]
- Simon, J.E.; Morales, M.R.; Phippen, W.B.; Vieira, R.F.; Hao, Z. Basil: A source of aroma compounds and a popular culinary and ornamental herb. Perspect. New Crops New Uses 1999, 16, 499–505. [Google Scholar]
- Martin, C.; Zhang, Y.; Tonelli, C.; Petroni, K. Plants, Diet, and Health. Annu. Rev. Plant Boil. 2013, 64, 19–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gang, D.R. Evolution of flavors and scents. Annu. Rev. Plant Boil. 2005, 56, 301–325. [Google Scholar] [CrossRef] [PubMed]
- Sgherri, C.; Cecconami, S.; Pinzino, C.; Navari-Izzo, F.; Izzo, R. Levels of antioxidants and nutraceuticals in basil grown in hydroponics and soil. Food Chem. 2010, 123, 416–422. [Google Scholar] [CrossRef]
- Zheljazkov, V.D.; Cantrell, C.L.; Evans, W.B.; Ebelhar, M.W.; Coker, C. Yield and Composition of Ocimum basilicum L. and Ocimum sanctum L. grown at four locations. HortScience 2008, 43, 737–741. [Google Scholar] [CrossRef] [Green Version]
- Khan, I.A.; Abourashed, E.A. Leung’s Encyclopedia of Common Natural Ingredients: Used in Food, Drugs and Cosmetics; Wiley: Hoboken, NJ, USA, 2009. [Google Scholar]
- Nicoletto, C.; Silvia, S.; Stefano, B.; Paolo, S. Influence of cut number on qualitative traits in different cultivars of sweet basil. Ind. Crop. Prod. 2013, 44, 465–472. [Google Scholar] [CrossRef]
- Murillo-Amador, B.; Nieto-Garibay, A.; Troyo-Diéguez, E.; Flores-Hernández, A.; Cordoba-Matson, M.V.; Villegas-Espinoza, A. Proximate analysis among 24 Ocimum cultivars under two cultivation environments: A comparative study. J. Food Agric. Environ. 2013, 11, 2842–2848. [Google Scholar]
- Resh, H.M. Hydroponic Food Production: A Definitive Guidebook for the Advanced Home Gardener and the Commercial Hydroponic Grower; CRC Press: Boca Raton, FL, USA, 2012. [Google Scholar]
- Tomasi, N.; Pinton, R.; Costa, L.D.; Cortella, G.; Terzano, R.; Mimmo, T.; Scampicchio, M.; Cesco, S. New ‘solutions’ for floating cultivation system of ready-to-eat salad: A review. Trends Food Sci. Technol. 2015, 46, 267–276. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.C. Enhancing quality of fresh vegetables through salinity eustress and biofortification applications facilitated by soilless cultivation. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Smith, J. Hydroponics: A Practical guide for the soilless grower. HortTechnology 2005, 15, 731. [Google Scholar] [CrossRef]
- Mason, J. Commercial Hydroponics: How to grow 86 different plants in hydroponics; Simon & Schuster Australia: New York, NY, USA, 2000. [Google Scholar]
- Putievsky, E.; Galambosi, B. Production systems of sweet basil. In Basil; CRC Press: Boca Raton, FL, USA, 1999; pp. 47–73. [Google Scholar]
- Bhargava, A.; Shukla, S.; Dixit, B.S.; Bannerji, R.; Ohri, D. Variability and genotype × cutting interactions for different nutritional components in Chenopodium album L. Hortic. Sci. 2011, 33, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Shukla, S.; Bhargava, A.; Chatterjee, A.; Srivastava, A.; Singh, S.P. Genotypic variability in vegetable amaranth (Amaranthus tricolor L for foliage yield and its contributing traits over successive cuttings and years. Euphytica 2006, 151, 103–110. [Google Scholar] [CrossRef]
- Kmiecik, W.; Lisiewska, Z. Comparison of leaf yields and chemical composition of the Hamburg and leafy types of parsley. Leaf yields and their structure. Folia Hortic. 1999, 11. [Google Scholar]
- Kumar, S.; Gupta, S.; Singh, P.; Bajpai, P.; Gupta, M.; Singh, D.; Gupta, A.; Ram, G.; Shasany, A.; Sharma, S. High yields of artemisinin by multi-harvest of Artemisia annua crops. Ind. Crop. Prod. 2004, 19, 77–90. [Google Scholar] [CrossRef]
- Csizinszky, A.A. Yield response of herbs to nitrogen and potassium in sand in multiple harvests. J. Herbs, Spices Med. Plants 2000, 6, 11–22. [Google Scholar] [CrossRef]
- May, A.; Bovi, O.A.; Maia, N.B.; Barata, L.E.S.; Souza, R.D.C.Z.D.; de Souza, E.M.R.; de Moraes, A.R.A.; Pinheiro, M.Q. Basil plants growth and essential oil yield in a production system with successive cuts. Bragantia 2008, 67, 385–389. [Google Scholar] [CrossRef]
- Bączek, K.; Kosakowska, O.; Gniewosz, M.; Gientka, I.; Węglarz, Z. Sweet basil (Ocimum basilicum L.) productivity and raw material quality from organic cultivation. Agronomy 2019, 9, 279. [Google Scholar] [CrossRef] [Green Version]
- Gary, C.; Jones, J.; Longuenesse, J. Modelling daily changes in specific leaf area of tomato: The contribution of the leaf assimilate pool. In Proceedings of International Workshop on Greenhouse Crop Models 328; Acta Horticulturae: Saumane, France, 1991; pp. 205–210. [Google Scholar]
- Barrs, H.; Weatherley, P. A Re-Examination of the relative turgidity technique for estimating water deficits in leaves. Aust. J. Boil. Sci. 1962, 15, 413. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Colla, G.; Giordano, M.; El-Nakhel, C.; Kyriacou, M.C.; de Pascale, S. Foliar applications of a legume-derived protein hydrolysate elicit dose-dependent increases of growth, leaf mineral composition, yield and fruit quality in two greenhouse tomato cultivars. Sci. Hortic. 2017, 226, 353–360. [Google Scholar] [CrossRef]
- Bremner, J. Semimicro-Kjeldahl Method. Chem. Microbiol. Prop. 1965, 2, 1171–1177. [Google Scholar]
- Sorgonà, A.; Abenavoli, M.R.; Gringeri, P.G.; Cacco, G. A comparison of nitrogen use efficiency definitions in Citrus rootstocks. Sci. Hortic. 2006, 109, 389–393. [Google Scholar] [CrossRef]
- Good, A.G.; Shrawat, A.K.; Muench, D.G. Can less yield more? Is reducing nutrient input into the environment compatible with maintaining crop production? Trends Plant Sci. 2004, 9, 597–605. [Google Scholar] [CrossRef]
- Siddiqi, M.Y.; Glass, A. Utilization index: A modified approach to the estimation and comparison of nutrient utilization efficiency in plants. J. Plant Nutr. 1981, 4, 289–302. [Google Scholar] [CrossRef]
- Gabelman, W.H.; Gerloff, G.C. The search for and interpretation of genetic controls that enhance plant growth under deficiency levels of a macronutrient. In Genetic Aspects of Plant Nutrition; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 1983; pp. 379–394. [Google Scholar]
- Fogliano, V.; Verde, V.; Randazzo, G.; Ritieni, A. Method for measuring antioxidant activity and its application to monitoring the antioxidant capacity of wines. J. Agric. Food Chem. 1999, 47, 1035–1040. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. In Methods in Enzymology; Elsevier BV: Amsterdam, The Netherlands, 1999; pp. 152–178. [Google Scholar]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Minutolo, M.; Chiaiese, P.; di Matteo, A.; Errico, A.; Corrado, G. Accumulation of ascorbic acid in tomato cell culture: Influence of the genotype, source explant and time of in vitro cultivation. Antioxidants 2020, 9, 222. [Google Scholar] [CrossRef] [Green Version]
- Corrado, G.; Lucini, L.; Miras-Moreno, M.; Chiaiese, P.; Colla, G.; de Pascale, S.; Rouphael, Y. Metabolic insights into the anion-anion antagonism in sweet basil: Effects of different nitrate/chloride ratios in the nutrient solution. Int. J. Mol. Sci. 2020, 21, 2482. [Google Scholar] [CrossRef] [Green Version]
- Chong, J.; Wishart, D.S.; Xia, J. Using MetaboAnalyst 4.0 for comprehensive and integrative metabolomics data analysis. Curr. Protoc. Bioinform. 2019, 68, e86. [Google Scholar] [CrossRef]
- Kim, S.; Thiessen, P.A.; Bolton, E.E.; Chen, J.; Fu, G.; Gindulyte, A.; Han, L.; He, J.; He, S.; Shoemaker, B.A.; et al. PubChem Substance and compound databases. Nucleic Acids Res. 2015, 44, 1202–1213. [Google Scholar] [CrossRef] [PubMed]
- Caspi, R.; Billington, R.; Keseler, I.M.; Kothari, A.; Krummenacker, M.; E Midford, P.; Ong, W.K.; Paley, S.; Subhraveti, P.; Karp, P.D. The MetaCyc database of metabolic pathways and enzymes—A 2019 update. Nucleic Acids Res. 2020, 48, 445–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoki, K.F.; Kanehisa, M. Using the KEGG Database Resource. Curr. Protoc. Bioinform. 2005, 11. [Google Scholar] [CrossRef] [PubMed]
- Team, R.; Core, R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Di Gioia, F.; Tzortzakis, N.; Rouphael, Y.; Kyriacou, M.C.; Sampaio, S.L.; Ferreira, I.C.F.R.; Petropoulos, S.A. Grown to be blue—Antioxidant properties and health effects of colored vegetables. Part II: Leafy, fruit, and other vegetables. Antioxidants 2020, 9, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, P.; Kalunke, R.M.; Giri, A.P. Towards comprehension of complex chemical evolution and diversification of terpene and phenylpropanoid pathways in Ocimum species. RSC Adv. 2015, 5, 106886–106904. [Google Scholar] [CrossRef]
- Gavrić, T.; Jurković, J.; Hamidović, S.; Haseljić, S.; Lalević, B.; Čorbo, A.; Bezdrob, M. Yield and contents of some bioactive components of basil (Ocimum basilicum L.) depending on time of cutting. Studia Universitatis" Vasile Goldis" Arad. Seria Stiintele Vietii 2018, 28, 192–197. [Google Scholar]
- Petropoulos, S.A.; Fernandes, A.; Karkanis, A.; Ntatsi, G.; Barros, L.; Ferreira, I.C.F.R. Successive harvesting affects yield, chemical composition and antioxidant activity of Cichorium spinosum L. Food Chem. 2017, 237, 83–90. [Google Scholar] [CrossRef] [Green Version]
- Garnier, E.; Laurent, G.; Bellmann, A.; Debain, S.; Berthelier, P.; Ducout, B.; Roumet, C.; Navas, M. Consistency of species ranking based on functional leaf traits. New Phytol. 2001, 152, 69–83. [Google Scholar] [CrossRef]
- Poorter, H.; de Jong, R. A comparison of specific leaf area, chemical composition and leaf construction costs of field plants from 15 habitats differing in productivity. New Phytol. 1999, 143, 163–176. [Google Scholar] [CrossRef] [Green Version]
- Wilson, P.J.; Thompson, K.; Hodgson, J.G. Specific leaf area and leaf dry matter content as alternative predictors of plant strategies. New Phytol. 1999, 143, 155–162. [Google Scholar] [CrossRef]
- Vendramini, F.; Díaz, S.; Gurvich, D.E.; Wilson, P.J.; Thompson, K.; Hodgson, J.G. Leaf traits as indicators of resource-use strategy in floras with succulent species. New Phytol. 2002, 154, 147–157. [Google Scholar] [CrossRef]
- Terashima, I.; Hikosaka, K. Comparative ecophysiology of leaf and canopy photosynthesis. Plant Cell Environ. 1995, 18, 1111–1128. [Google Scholar] [CrossRef]
- Reich, P.B.; Ellsworth, D.S.; Walters, M.B. Leaf structure (Specific Leaf Area) modulates photosynthesis-nitrogen relations: Evidence from within and across species and functional groups. Funct. Ecol. 1998, 12, 948–958. [Google Scholar] [CrossRef]
- Locascio, S.J.; Wiltbank, W.J.; Gull, D.D.; Maynard, D.N.; Hauck, R.D. Fruit and vegetable quality as affected by nitrogen nutrition. Soil Surv. Land Use Plann. 2015, 617–626. [Google Scholar] [CrossRef]
- Ullrich, W.R.; Rigano, C.; Fuggi, A.; Aparicio, P.J. Inorganic Nitrogen in Plants and Microorganisms: Uptake and Metabolism; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Millard, P. The accumulation and storage of nitrogen by herbaceous plants. Plant Cell Environ. 1988, 11, 1–8. [Google Scholar] [CrossRef]
- European Food safety Authority (EFSA). Nitrate in vegetables-Scientific opinion of the panel on contaminants in the food chain. EFSA J. 2008, 6, 689. [Google Scholar] [CrossRef]
- Colla, G.; Kim, H.-J.; Kyriacou, M.C.; Rouphael, Y. Nitrate in fruits and vegetables. Sci. Hortic. 2018, 237, 221–238. [Google Scholar] [CrossRef]
- Ferraro, D.O.; Oesterheld, M. Effect of defoliation on grass growth. A quantitative review. Oikos 2002, 98, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Chen, S.-P.; Lin, G. Compensatory growth responses to clipping defoliation in Leymus chinensis (Poaceae) under nutrient addition and water deficiency conditions. Plant Ecol. 2008, 196, 85–99. [Google Scholar] [CrossRef]
- Green, R.A.; Detling, J.K. Defoliation-induced enhancement of total aboveground nitrogen yield of grasses. Oikos 2000, 91, 280–284. [Google Scholar] [CrossRef]
- Schmitt, A.; Pausch, J.; Kuzyakov, Y. C and N allocation in soil under ryegrass and alfalfa estimated by 13C and 15N labelling. Plant Soil 2012, 368, 581–590. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.-L.; Guo, X.-L.; Hou, X.-G.; Zhao, W.; Xu, G.-W.; Li, Z.-Q. Effects of leaf zeatin and zeatin riboside induced by different clipping heights on the regrowth capacity of ryegrass. Ecol. Res. 2013, 29, 167–180. [Google Scholar] [CrossRef] [Green Version]
- Skalák, J.; Vercruyssen, L.; Claeys, H.; Hradilová, J.; Černý, M.; Novák, O.; Plačková, L.; Saiz-Fernández, I.; Skaláková, P.; Coppens, F.; et al. Multifaceted activity of cytokinin in leaf development shapes its size and structure in Arabidopsis. Plant J. 2019, 97, 805–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Bris, M. Hormones in Growth and Development. In Reference Module in Life Sciences; Elsevier BV: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Puértolas, J.; Albacete, A.; Dodd, I.C. Irrigation frequency transiently alters whole plant gas exchange, water and hormone status, but irrigation volume determines cumulative growth in two herbaceous crops. Environ. Exp. Bot. 2020, 176, 104101. [Google Scholar] [CrossRef]
- Miceli, A.; Moncada, A.; Sabatino, L.; Vetrano, F. Effect of gibberellic acid on growth, yield, and quality of leaf lettuce and rocket grown in a floating system. Agronomy 2019, 9, 382. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Kapteyn, J.; Gang, D.R. A systems biology investigation of the MEP/terpenoid and shikimate/phenylpropanoid pathways points to multiple levels of metabolic control in sweet basil glandular trichomes. Plant J. 2008, 54, 349–361. [Google Scholar] [CrossRef]
- Ceccanti, C.; Landi, M.; Rocchetti, G.; Miras-Moreno, M.; Lucini, L.; Incrocci, L.; Pardossi, A.; Guidi, L. Hydroponically Grown Sanguisorba minor Scop.: Effects of cut and storage on fresh-cut produce. Antioxidants 2019, 8, 631. [Google Scholar] [CrossRef] [Green Version]
- Ceccanti, C.; Landi, M.; Rocchetti, G.; Moreno, M.B.M.; Lucini, L.; Incrocci, L.; Pardossi, A.; Guidi, L. Effect of cut on secondary metabolite profile in hydroponically-grown Rumex acetosa L. seedlings: A metabolomic approach. Nat. Prod. Res. 2020, 1–5. [Google Scholar] [CrossRef]
- Basile, B.; Andreotti, C.; Rogers, H.; Rouphael, Y. Ushering horticulture into a new era of research-based novelty. Italus Hortus 2020, 27, 1. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corrado, G.; Chiaiese, P.; Lucini, L.; Miras-Moreno, B.; Colla, G.; Rouphael, Y. Successive Harvests Affect Yield, Quality and Metabolic Profile of Sweet Basil (Ocimum basilicum L.). Agronomy 2020, 10, 830. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10060830
Corrado G, Chiaiese P, Lucini L, Miras-Moreno B, Colla G, Rouphael Y. Successive Harvests Affect Yield, Quality and Metabolic Profile of Sweet Basil (Ocimum basilicum L.). Agronomy. 2020; 10(6):830. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10060830
Chicago/Turabian StyleCorrado, Giandomenico, Pasquale Chiaiese, Luigi Lucini, Begoña Miras-Moreno, Giuseppe Colla, and Youssef Rouphael. 2020. "Successive Harvests Affect Yield, Quality and Metabolic Profile of Sweet Basil (Ocimum basilicum L.)" Agronomy 10, no. 6: 830. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10060830