Effects of Moderately-Reduced Water Supply and Picking Time on the Chemical Composition of Pickling Cucumber (Cucumis sativus L.) in Open Field Cultivation

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Open Field Trial
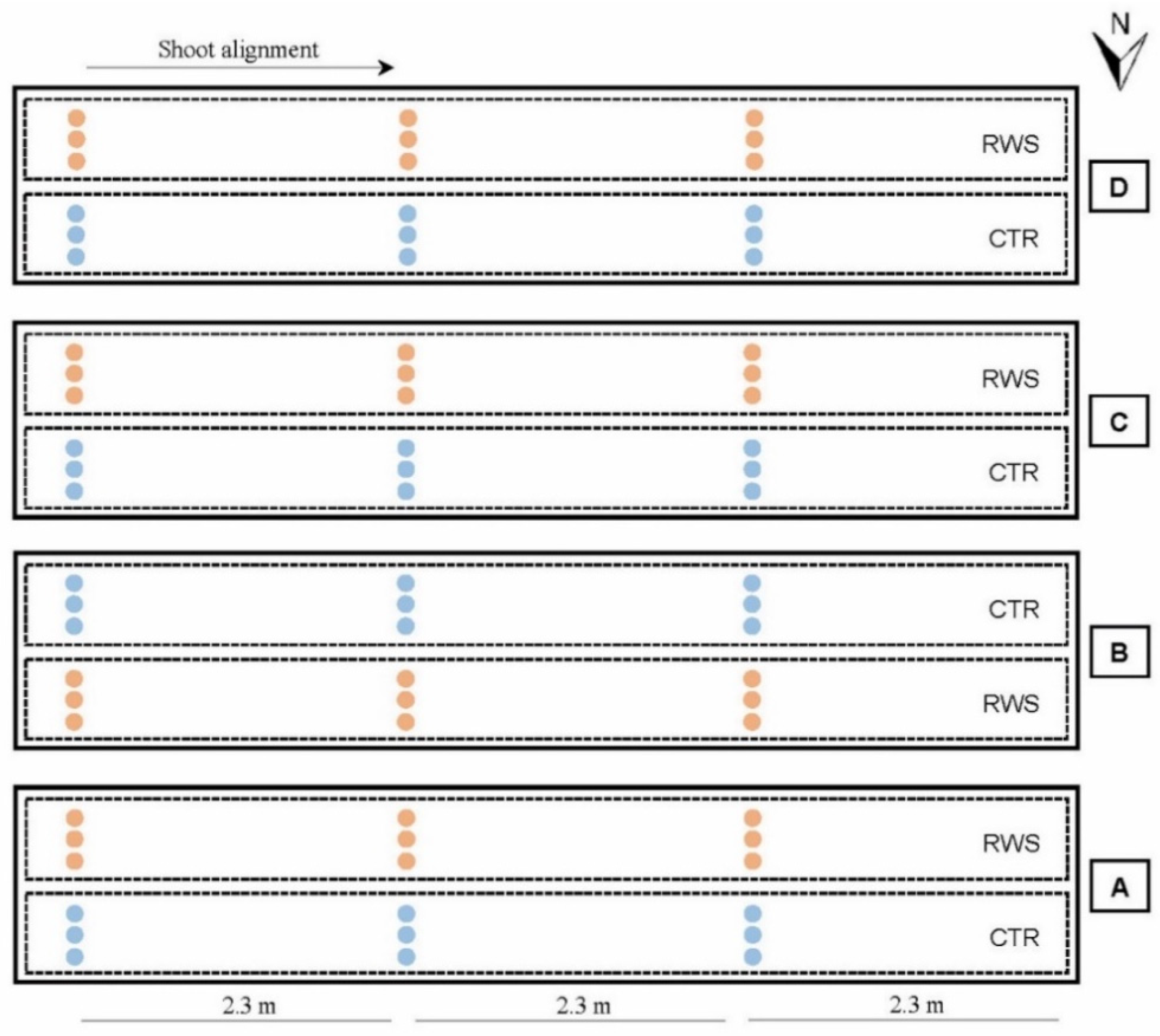
2.2.1. Plant Material and Field Experimental Design
2.2.2. Cultivation and Water Supply
2.2.3. Harvest
2.2.4. Climatic Data
2.2.5. Sample Preparation
2.2.6. Chemical Analysis
2.3. Greenhouse Trial
2.3.1. Plant Material and Experimental Cultivation Setup
2.3.2. Water Supply Treatments
2.3.3. Harvest
2.3.4. Sample Preparation and Chemical Analysis
2.4. Statistical analyses
3. Results and Discussion
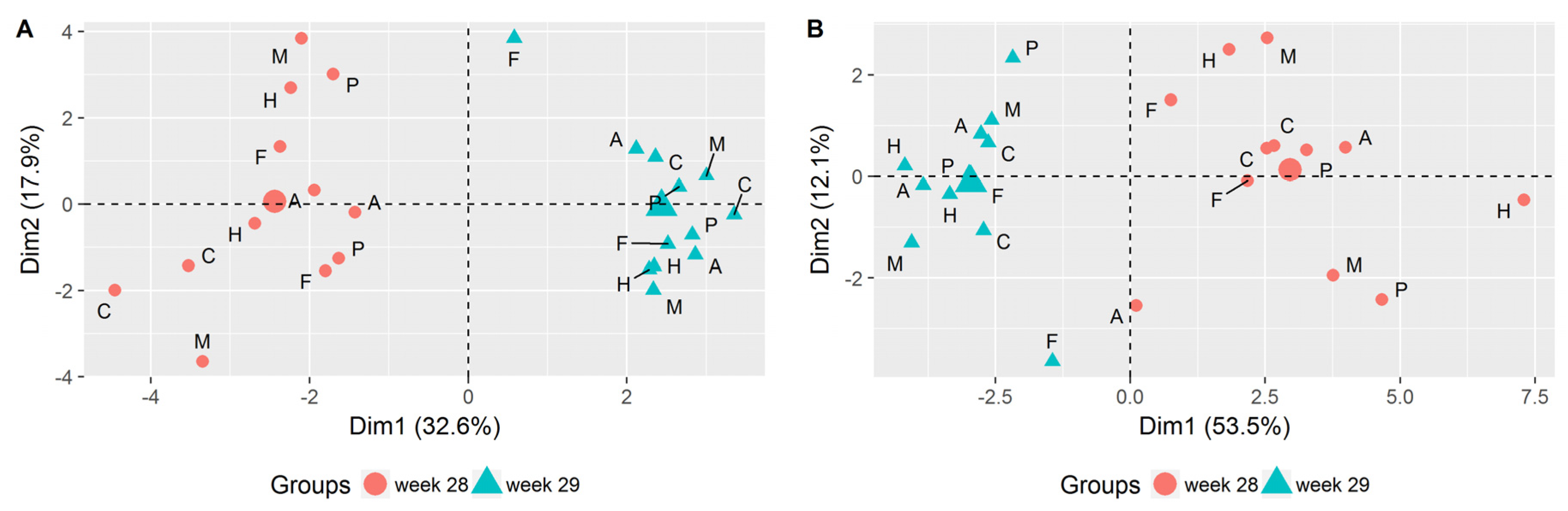
3.1. Evaluation of Picking Time
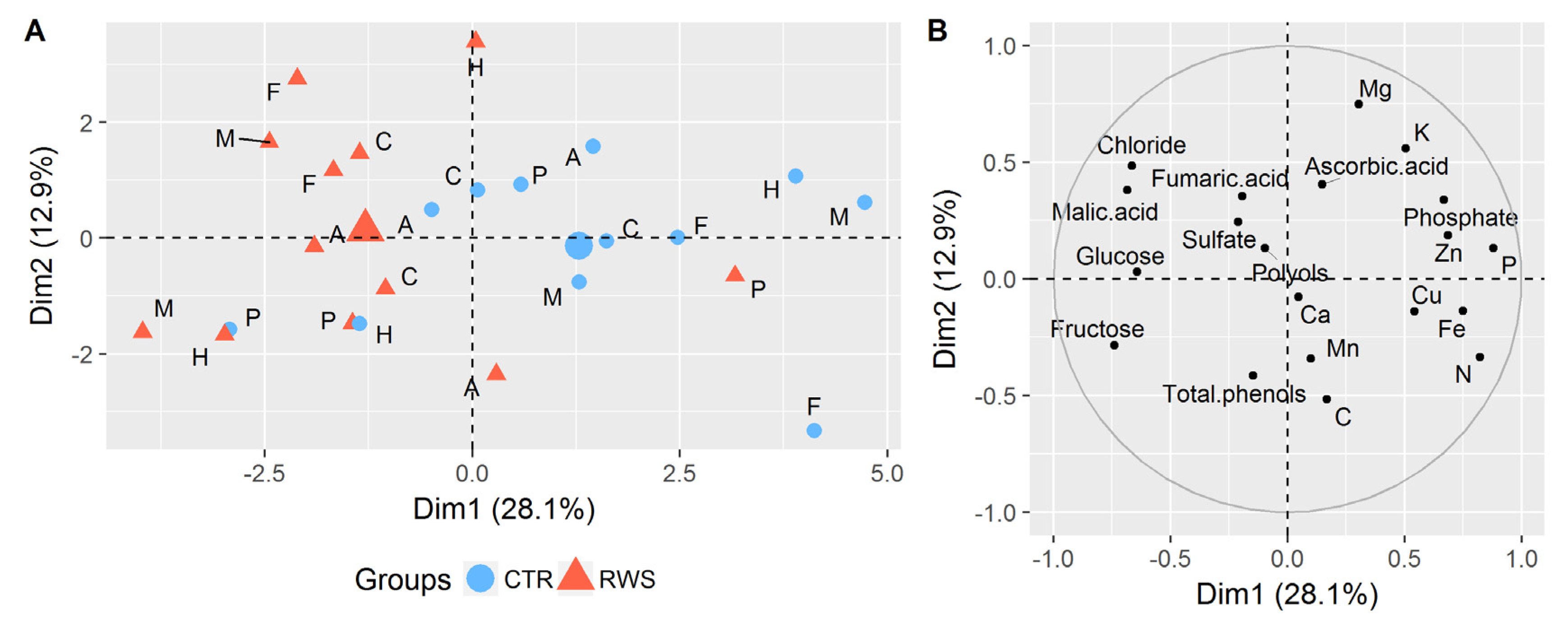
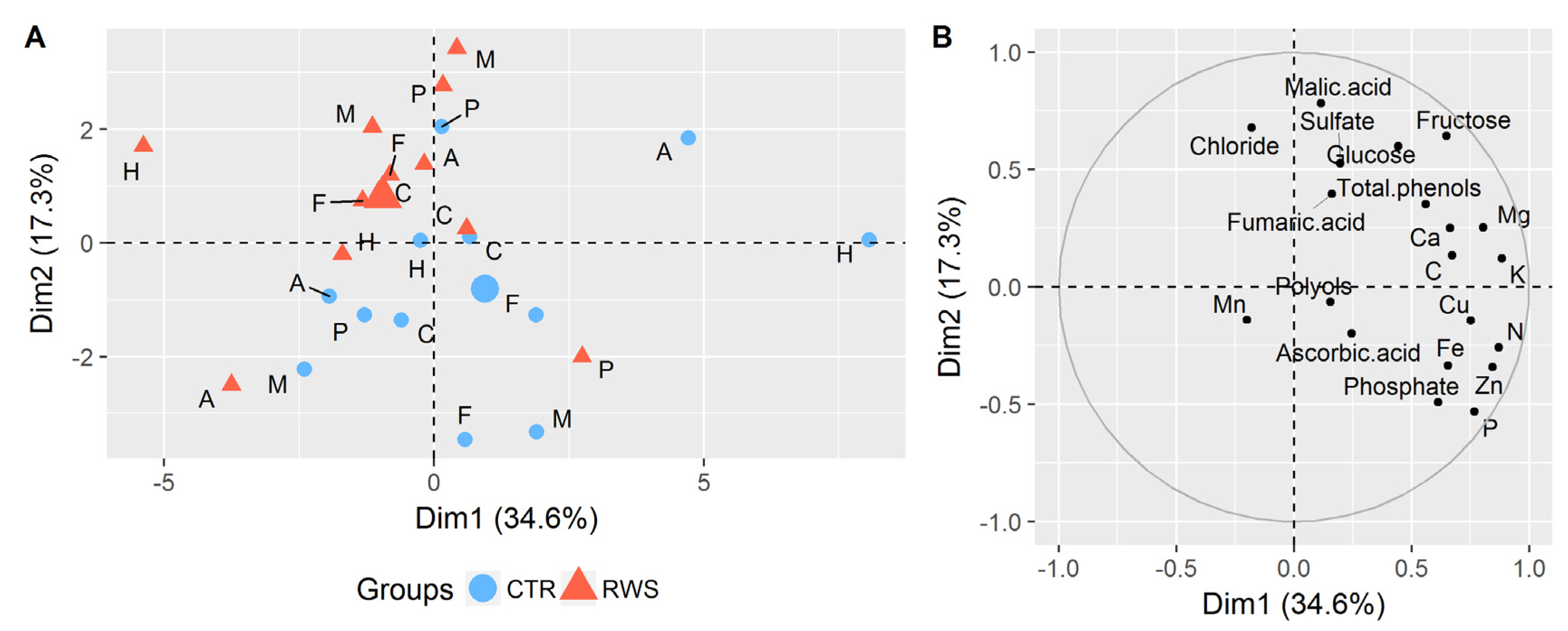
3.2. Effects of Reduced Water Supply on the Chemical Composition of Cucumber Biomass
3.2.1. Sugars and Polyols
3.2.2. Organic Acids
3.2.3. Inorganic Anions
3.2.4. Carbon
3.2.5. Nitrogen
3.2.6. Potassium
3.2.7. Phosphorous
3.2.8. Calcium
3.2.9. Magnesium
3.2.10. Micronutrients (Fe, Zn, Mn, Cu)
3.2.11. Phenolic Compounds
3.2.12. Carotenoids
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Olesen, J.E.; Carter, T.R.; Díaz-Ambrona, C.H.; Fronzek, S.; Heidmann, T.; Hickler, T.; Holt, T.; Minguez, M.I.; Morales, P.; Palutikof, J.P.; et al. Uncertainties in projected impacts of climate change on European agriculture and terrestrial ecosystems based on scenarios from regional climate models. Clim. Chang. 2007, 81, 123–143. [Google Scholar] [CrossRef]
- Gómez-López, M.D.; Fernández-Trujillo, J.P.; Baille, A. Cucumber fruit quality at harvest affected by soilless system, crop age and preharvest climatic conditions during two consecutive seasons. Sci. Hortic. 2006, 110, 68–78. [Google Scholar] [CrossRef]
- Soares, J.C.; Santos, C.S.; Carvalho, S.M.P.; Pintado, M.M.; Vasconcelos, M.W. Preserving the nutritional quality of crop plants under a changing climate: Importance and strategies. Plant Soil 2019, 443, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Schlering, C.; Dietrich, H.; Frisch, M.; Schreiner, M.; Schweiggert, R.; Will, F.; Zinkernagel, J. Chemical composition of field grown radish (Raphanus sativus L. var. sativus) as influenced by season and moderately reduced water supply. J. Appl. Bot. Food Qual. 2019, 343–354. [Google Scholar] [CrossRef]
- Schlering, C.; Zinkernagel, J.; Dietrich, H.; Frisch, M.; Schweiggert, R. Alterations in the Chemical Composition of Spinach (Spinacia oleracea L.) as Provoked by Season and Moderately Limited Water Supply in Open Field Cultivation. Horticulturae 2020, 6, 25. [Google Scholar] [CrossRef] [Green Version]
- Chaves, M.M.; Maroco, J.P.; Pereira, J.S. Understanding plant responses to drought—From genes to the whole plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef]
- Cramer, G.R.; Urano, K.; Delrot, S.; Pezzotti, M.; Shinozaki, K. Effects of abiotic stress on plants: A systems biology perspective. BMC Plant Biol. 2011, 11, 163. [Google Scholar] [CrossRef] [Green Version]
- Högy, P.; Poll, C.; Marhan, S.; Kandeler, E.; Fangmeier, A. Impacts of temperature increase and change in precipitation pattern on crop yield and yield quality of barley. Food Chem. 2013, 136, 1470–1477. [Google Scholar] [CrossRef]
- Daryanto, S.; Wang, L.; Jacinthe, P.-A. Global synthesis of drought effects on cereal, legume, tuber and root crops production: A review. Agric. Water Manag. 2017, 179, 18–33. [Google Scholar] [CrossRef] [Green Version]
- Najarian, M.; Mohammadi-Ghehsareh, A.; Fallahzade, J.; Peykanpour, E. Responses of cucumber (Cucumis sativus L.) to ozonated water under varying drought stress intensities. J. Plant Nutr. 2018, 41, 1–9. [Google Scholar] [CrossRef]
- Pulupol, L.U.; Behboudian, M.H.; Fisher, K.J. Growth, Yield, and Postharvest Attributes of Glasshouse Tomatoes Produced under Deficit Irrigation. HortScience 1996, 31, 926–929. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.; Liu, M.; Wang, X.; Liu, C.; Hou, Z.; Shi, J. Effects of deficit irrigation on yield and water use of greenhouse grown cucumber in the North China Plain. Agric. Water Manag. 2003, 61, 219–228. [Google Scholar] [CrossRef]
- Li, J.; Zou, Z.; Wang, X. The present studying situation and existing problems of water-saving irrigation index for vegetable. Agric. Res. Arid Areas 2000, 18, 118–123. (In Chinese) [Google Scholar]
- Rahil, M.H.; Qanadillo, A. Effects of different irrigation regimes on yield and water use efficiency of cucumber crop. Agric. Water Manag. 2015, 148, 10–15. [Google Scholar] [CrossRef]
- Rouphael, Y.; Cardarelli, M.; Schwarz, D.; Franken, P.; Colla, G. Effects of Drought on Nutrient Uptake and Assimilation in Vegetable Crops. In Plant Responses to Drought Stress: From Morphological to Molecular Features; Aroca, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 171–195. ISBN 978-3-642-32653-0. [Google Scholar]
- Hodges, D.M.; Lester, G.E. Curcurbits: Cucumber, Melon, Pumpkin and Squash. In Health-Promoting Properties of Fruit and Vegetables; [Enhanced Credo edition]; Terry, L.A., Ed.; Credo Reference: Wallingford, UK; Cambridge, MA, USA; CABI: Boston, MA, USA, 2013; pp. 118–134. ISBN 978-1-78064-422-6. [Google Scholar]
- Bates, D. Fitting Linear Mixed Models: The Newsletter of the R-project. Available online: http://CRAN.R-project.org/doc/Rnews/ (accessed on 3 December 2018).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- RStudio Team. RStudio: Integrated Development Environment for R; RStudio, Inc.: Boston, MA, USA, 2016. [Google Scholar]
- Lenth, R.V. Least-Squares Means: The R Package lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest: Tests in Linear Mixed Effects Models. R Package Version 2.0-33. Available online: https://CRAN.R-project.org/package=lmerTest (accessed on 26 March 2018).
- Le, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Kassambara, A.; Mundt, F. factoextra: Extract and Visualize the Results of Multivariate Data Analyses. R Package Version 1.0.5. Available online: https://CRAN.R-project.org/package=factoextra (accessed on 27 March 2018).
- Gómez, M.D.; Baille, A.; González-Real, M.M.; Mercader, J.M. Dry matter partitioning of greenhouse cucumber crops as affected by fruit load. Acta Hortic. 2003, 614, 573–578. [Google Scholar] [CrossRef]
- Gargallo-Garriga, A.; Sardans, J.; Pérez-Trujillo, M.; Rivas-Ubach, A.; Oravec, M.; Vecerova, K.; Urban, O.; Jentsch, A.; Kreyling, J.; Beierkuhnlein, C.; et al. Opposite metabolic responses of shoots and roots to drought. Sci. Rep. 2014, 4, 6829. [Google Scholar] [CrossRef] [Green Version]
- Akinci, S.; Lösel, D.M. The effects of water stress and recovery periods on soluble sugars and starch content in cucumber cultivars. Fresenius Environ. Bull. 2010, 19, 164–171. [Google Scholar]
- Bohnert, H.J.; Nelson, D.E.; Jensen, R.G. Adaptations to Environmental Stresses. Plant Cell 1995, 7, 1099–1111. [Google Scholar] [CrossRef]
- Nuccio, M.L.; Rhodest, D.; McNeil, S.D.; Hanson, A.D. Metabolic engineering of plants for osmotic stress resistance. Curr. Opin. Plant Biol. 1999, 2, 128–134. [Google Scholar] [CrossRef]
- McFeeters, R.F.; Fleming, H.P.; Thompson, R.L. Malic and Citric Acids in Pickling Cucumbers. J. Food Sci. 1982, 47, 1859–1861. [Google Scholar] [CrossRef]
- Mitchell, J.P.; Shennan, C.; Grattan, S.R. Developmental changes in tomato fruit composition in response to water deficit and salinity. Physiol. Plant. 1991, 83, 177–185. [Google Scholar] [CrossRef]
- Lobit, P.; Genard, M.; Soing, P.; Habib, R. Modelling malic acid accumulation in fruits: Relationships with organic acids, potassium, and temperature. J. Exp. Bot. 2006, 57, 1471–1483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cutler, J.M.; Rains, D.W. Effects of Water Stress and Hardening on the Internal Water Relations and Osmotic Constituents of Cotton Leaves. Physiol. Plant. 1978, 42, 261–268. [Google Scholar] [CrossRef]
- Davies, J.N. Effect of nitrogen, phosphorus and potassium fertilisers on the non-volatile organic acids of tomato fruit. J. Sci. Food Agric. 1964, 15, 665–673. [Google Scholar] [CrossRef]
- Młodzińska, E.; Zboińska, M. Phosphate Uptake and Allocation—A Closer Look at Arabidopsis thaliana L. and Oryza sativa L. Front. Plant Sci. 2016, 7, 1198. [Google Scholar] [CrossRef] [Green Version]
- Raghothama, K.G. Phosphate Acquisition. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 665–693. [Google Scholar] [CrossRef]
- Rausch, C.; Bucher, M. Molecular mechanisms of phosphate transport in plants. Planta 2002, 216. [Google Scholar] [CrossRef]
- Da Silva, E.C.; Nogueira, R.J.M.C.; Silva, M.; Albuquerque, M. Drought Stress and Plant Nutrition. Plant Stress 2010, 5, 32–41. [Google Scholar]
- Rouphael, Y.; Cardarelli, M.; Colla, G.; Rea, E. Yield, Mineral Composition, Water Relations, and Water Use Efficiency of Grafted Mini-watermelon Plants under Deficit Irrigation. HortScience 2008, 43, 730–736. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Rodríguez, E.; del Mar Rubio-Wilhelmi, M.; Cervilla, L.M.; Blasco, B.; Rios, J.J.; Leyva, R.; Romero, L.; Ruiz, J.M. Study of the ionome and uptake fluxes in cherry tomato plants under moderate water stress conditions. Plant Soil 2010, 335, 339–347. [Google Scholar] [CrossRef]
- Hu, Y.; Schmidhalter, U. Drought and salinity: A comparison of their effects on mineral nutrition of plants. J. Plant Nutr. Soil Sci. 2005, 168, 541–549. [Google Scholar] [CrossRef]
- Hedges, L.J.; Lister, C.E. Nutritional attributes of salad vegetables. Crop & Food Research Confidential Report No. 1473. Available online: http://www.freshvegetables.co.nz/assets/Members-pdfs/F001435797-2005-Nutritional-attributes-of-salad-vegetables-Copy.pdf (accessed on 30 April 2020).
- Abu-Reidah, I.M.; Arráez-Román, D.; Quirantes-Piné, R.; Fernández-Arroyo, S.; Segura-Carretero, A.; Fernández-Gutiérrez, A. HPLC–ESI-Q-TOF-MS for a comprehensive characterization of bioactive phenolic compounds in cucumber whole fruit extract. Food Res. Int. 2012, 46, 108–117. [Google Scholar] [CrossRef]
- Ripoll, J.; Urban, L.; Brunel, B.; Bertin, N. Water deficit effects on tomato quality depend on fruit developmental stage and genotype. J. Plant Physiol. 2016, 190, 26–35. [Google Scholar] [CrossRef]
- Johnson, E.J. A possible role for lutein and zeaxanthin in cognitive function in the elderly. Am. J. Clin. Nutr. 2012, 96, 1161S–1165S. [Google Scholar] [CrossRef] [Green Version]
- Johnson, E.J. Role of lutein and zeaxanthin in visual and cognitive function throughout the lifespan. Nutr. Rev. 2014, 72, 605–612. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Picking Time Period | Week 28 | Week 29 |
|---|---|---|
| Date of transplanting (Year-month-day) | 2016-06-02 | 2016-06-02 |
| Beginning of water supply differentiation (DAT) | 19 | 19 |
| Beginning of harvest period after planting (DAT) | 39 | 46 |
| Temperature sum (°C) | 775.7 | 922.9 |
| Daily mean air temperature (°C) | 18.9 | 19.2 |
| Mean relative air humidity (%) | 89.6 | 88.3 |
| Global radiation sum (MJ/m²) | 777.3 | 931.9 |
| Daily mean global radiation (MJ/m²) | 19.0 | 19.4 |
| Wind speed sum (m/s) at height of 2 m | 52.5 | 62.1 |
| Evapotranspiration sum (mm) z | 133.3 | 161.8 |
| Precipitation sum (mm) | 84 | 84 |
| Number of differentiated irrigation events | 10 | 15 |
| Total irrigation (mm) # | 29 | 47 |
| Total water amount (mm) of CTR incl. precipitation | 114 | 131 |
| Total water amount RWS (% of CTR) * | 90% | 85% |
| Cucumber Week 28 | Cucumber Week 29 | Cucumber Total | Effect of Harvest Period (Week) | Interaction of Water Supply (H2O) * Harvest Period (Week) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| CTR | RWS | p-Value | CTR | RWS | p-Value | CTR | RWS | p-Value | p-Value | p-Value | |
| Dry matter (% of FM) | 6.32 | 6.14 | 0.2180 | 5.29 | 5.37 | 0.2371 | 5.81 | 5.75 | 0.5039 | 0.000 *** | 0.000 *** |
| Sugars and polyols | |||||||||||
| Glucose (mg/g) | 179.0 | 181.0 | 0.7129 | 197.5 | 200.2 | 0.2706 | 188.2 | 190.6 | 0.3895 | 0.000 *** | 0.000 *** |
| Fructose (mg/g) | 209.6 | 211.8 | 0.3328 | 207.3 | 208.1 | 0.6342 | 208.5 | 210.0 | 0.2560 | 0.206 | 0.724 |
| Polyols total (mg/g) | 9.2 | 9.2 | 0.7729 | 8.9 | 8.6 | 0.1386 | 9.0 | 8.9 | 0.3768 | 0.007 ** | 0.857 |
| Inositol (mg/g) | 8.1 | 8.1 | 0.7879 | 7.9 | 7.5 | 0.1022 | 8.0 | 7.9 | 0.2820 | 0.015 * | 0.965 |
| Glycerol (mg/g) | 0.8 | 0.9 | 0.6344 | 0.8 | 0.8 | 0.2606 | 0.8 | 0.9 | 0.2250 | 0.667 | 0.816 |
| Erythritol (mg/g) | 0.3 | 0.3 | 1.0000 | 0.2 | 0.2 | 0.7294 | 0.2 | 0.2 | 0.8735 | 0.000 *** | 0.002 ** |
| Organic acids | |||||||||||
| Total acidity (mg/g) | 16.7 | 16.1 | 0.7602 | 11.3 | 11.6 | 0.6594 | 14.0 | 13.9 | 0.8784 | 0.000 *** | 0.002 ** |
| Malic acid (mg/g) | 36.3 | 37.8 | 0.2068 | 41.2 | 44.6 | 0.0095 ** | 38.7 | 41.2 | 0.0049 ** | 0.000 *** | 0.000 *** |
| Ascorbic acid (mg/g) | 1.2 | 1.2 | 0.5583 | 1.3 | 1.3 | 0.6535 | 1.2 | 1.2 | 0.9735 | 0.153 | 0.831 |
| Citric acid (mg/g) | nd | nd | - | 0.3 | 0.4 | 0.1099 | nd | nd | - | - | - |
| Fumaric acid (mg/g) | 7.2 | 6.4 | 0.7665 | 5.7 | 7.2 | 0.3429 | 6.5 | 6.8 | 0.8185 | 1.000 | 1.000 |
| Anions | |||||||||||
| Nitrate (mg/g) | nd | nd | - | 0.5 | 0.5 | 0.6728 | nd | nd | - | - | - |
| Phosphate (mg/g) | 14.6 | 13.2 | 0.3732 | 16.7 | 15.0 | 0.0911 | 15.6 | 14.1 | 0.0751 | 0.112 | 0.612 |
| Sulfate (mg/g) | 5.3 | 5.3 | 0.8516 | 4.0 | 4.3 | 0.2664 | 4.6 | 4.8 | 0.2406 | 0.000 *** | 0.001 ** |
| Chloride (mg/g) | 2.1 | 2.0 | 0.6668 | 2.5 | 2.7 | 0.2750 | 2.3 | 2.4 | 0.5686 | 0.002 ** | 0.052 |
| Elements | |||||||||||
| Carbon (mg/g) | 392.4 | 392.1 | 0.7616 | 395.8 | 395.0 | 0.6843 | 394.1 | 393.3 | 0.4051 | 0.049 * | 0.316 |
| Macronutrients | |||||||||||
| Potassium (mg/g) | 40.39 | 39.61 | 0.1966 | 38.11 | 38.26 | 0.8241 | 39.25 | 38.93 | 0.4490 | 0.002 ** | 0.497 |
| Nitrogen (mg/g) | 27.44 | 26.59 | 0.2702 | 25.01 | 23.95 | 0.1639 | 26.22 | 25.27 | 0.0648 | 0.000 *** | 0.001 ** |
| Phosphorous (mg/g) | 6.71 | 6.49 | 0.3852 | 6.84 | 6.47 | 0.0217 * | 6.77 | 6.48 | 0.0365 * | 1.000 | 1.000 |
| Calcium (mg/g) | 4.33 | 4.31 | 0.9028 | 4.35 | 4.49 | 0.0282 * | 4.34 | 4.40 | 0.3417 | 0.609 | 1.000 |
| Magnesium (mg/g) | 2.48 | 2.45 | 0.3434 | 2.66 | 2.71 | 0.0694 | 2.57 | 2.58 | 0.6571 | 0.000 *** | 0.196 |
| Micronutrients | |||||||||||
| Iron (µg/g) | 72 | 64 | 0.2778 | 63 | 54 | 0.0479 * | 68 | 58 | 0.0350 * | 0.111 | 0.063 |
| Zinc (µg/g) | 34 | 33 | 0.4062 | 35 | 34 | 0.1611 | 34 | 33 | 0.1328 | 0.423 | 0.589 |
| Manganese (µg/g) | 17 | 17 | 0.5155 | 16 | 16 | 0.5891 | 16 | 16 | 0.3697 | 0.792 | 1.000 |
| Copper (µg/g) | 10 | 10 | 0.6336 | 11 | 11 | 0.5645 | 11 | 10 | 0.4407 | 0.327 | 0.793 |
| Phenolic compounds | |||||||||||
| Total phenols (mg/g) | 2.8 | 2.9 | 0.4941 | 2.7 | 2.7 | 0.8606 | 2.7 | 2.8 | 0.5028 | 0.021 * | 0.133 |
| Cucumber Week 28 | Cucumber Week 29 | Cucumber Total | Effect of Harvest Period (Week) | Interaction of Water Supply (H2O) * Harvest Period (Week) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| CTR | RWS | p-Value | CTR | RWS | p-Value | CTR | RWS | p-Value | p-Value | p-Value | |
| Dry matter (% of FM) | 6.32 | 6.14 | 0.2180 | 5.29 | 5.37 | 0.2371 | 5.81 | 5.75 | 0.5039 | 0.000 *** | 0.000 *** |
| Sugars and polyols | |||||||||||
| Glucose (g/100 g) | 1.13 | 1.11 | 0.6675 | 1.05 | 1.08 | 0.2438 | 1.09 | 1.09 | 0.8832 | 0.051 | 0.347 |
| Fructose (g/100 g) | 1.33 | 1.30 | 0.4618 | 1.10 | 1.12 | 0.3881 | 1.21 | 1.21 | 0.8238 | 0.000 *** | 0.000 *** |
| Polyols total (mg/100 g) | 57.9 | 56.6 | 0.2132 | 56.2 | 52.4 | 0.0607 | 57.0 | 54.7 | 0.0208 * | 0.019 * | 0.630 |
| Inositol (mg/100 g) | 51.0 | 49.8 | 0.2321 | 49.7 | 46.0 | 0.0395 * | 50.4 | 48.1 | 0.0156 * | 0.029 * | 0.675 |
| Glycerol (mg/100 g) | 5.3 | 5.2 | 0.6916 | 5.1 | 5.1 | 0.9237 | 5.2 | 5.2 | 0.8980 | 0.947 | 1.000 |
| Erythritol (mg/100 g) | 1.6 | 1.5 | 0.3627 | 1.4 | 1.3 | 0.2192 | 1.5 | 1.4 | 0.1761 | 0.000 *** | 0.000 *** |
| Organic acids | |||||||||||
| Total acidity (mg/100 g) | 105.3 | 99.0 | 0.5229 | 60.0 | 62.0 | 0.5784 | 82.7 | 80.5 | 0.6624 | 0.000 *** | 0.000 *** |
| Malic acid (mg/100 g) | 227.2 | 232.9 | 0.5577 | 218.6 | 239.1 | 0.0219 * | 222.9 | 236.0 | 0.0436 * | 1.000 | 0.429 |
| Ascorbic acid (mg/100 g) | 7.5 | 7.0 | 0.2268 | 6.9 | 7.2 | 0.5088 | 7.2 | 7.1 | 0.7140 | 1.000 | 1.000 |
| Citric acid (mg/100 g) | nd | nd | - | 1.6 | 1.9 | 0.1144 | nd | nd | - | - | - |
| Fumaric acid (mg/100 g) | 45.0 | 39.4 | 0.7437 | 30.3 | 38.5 | 0.3139 | 37.6 | 38.9 | 0.8840 | 1.000 | 1.000 |
| Anions | |||||||||||
| Nitrate (mg/100 g) | nd | nd | - | 2.4 | 2.8 | 0.6398 | nd | nd | - | - | - |
| Phosphate (mg/100 g) | 91.1 | 81.2 | 0.3087 | 88.6 | 80.4 | 0.0897 | 89.9 | 80.8 | 0.0732 | 1.000 | 1.000 |
| Sulfate (mg/100 g) | 32.9 | 32.4 | 0.4909 | 21.1 | 23.3 | 0.2022 | 27.0 | 27.8 | 0.3540 | 0.000 *** | 0.000 *** |
| Chloride (mg/100 g) | 13.1 | 12.4 | 0.5563 | 13.2 | 14.5 | 0.2116 | 13.2 | 13.4 | 0.6985 | 0.489 | 1.000 |
| Elements | |||||||||||
| Carbon (g/100 g) | 2.5 | 2.4 | 0.1987 | 2.1 | 2.1 | 0.3342 | 2.3 | 2.3 | 0.4776 | 0.000 *** | 0.000 *** |
| Macronutrients | |||||||||||
| Potassium (mg/100 g) | 255.24 | 243.11 | 0.1372 | 201.69 | 205.23 | 0.1457 | 228.47 | 224.17 | 0.3231 | 0.000 *** | 0.000 *** |
| Nitrogen (mg/100 g) | 173.27 | 163.20 | 0.1218 | 132.37 | 128.60 | 0.4133 | 152.82 | 145.90 | 0.0829 | 0.000 *** | 0.000 *** |
| Phosphorous (mg/100 g) | 42.33 | 39.81 | 0.1845 | 36.23 | 34.70 | 0.0059 ** | 39.28 | 37.25 | 0.0296 * | 0.000 *** | 0.048 * |
| Calcium (mg/100 g) | 27.35 | 26.39 | 0.2697 | 22.99 | 24.10 | 0.0646 | 25.17 | 25.25 | 0.8904 | 0.000 *** | 0.052 |
| Magnesium (mg/100 g) | 15.66 | 14.98 | 0.1385 | 14.04 | 14.55 | 0.0095 ** | 14.85 | 14.77 | 0.7585 | 0.003 ** | 0.263 |
| Micronutrients | |||||||||||
| Iron (µg/100 g) | 455 | 390 | 0.2138 | 334 | 290 | 0.0739 | 399 | 340 | 0.0404 * | 0.001 ** | 0.002 ** |
| Zinc (µg/100 g) | 213 | 202 | 0.1443 | 183 | 182 | 0.1399 | 198 | 192 | 0.0848 | 0.000 *** | 0.000 *** |
| Manganese (µg/100 g) | 104 | 105 | 0.8338 | 84 | 87 | 0.4063 | 94 | 96 | 0.4567 | 0.001 ** | 0.008 ** |
| Copper (µg/100 g) | 65 | 61 | 0.3896 | 58 | 57 | 0.7708 | 61 | 59 | 0.3917 | 0.093 | 0.282 |
| Phenolic compounds | |||||||||||
| Total phenols (mg/100 g) | 17.7 | 17.5 | 0.8832 | 14.1 | 14.3 | 0.6144 | 15.9 | 15.9 | 0.8957 | 0.000 *** | 0.000 *** |
| Exocarp | Mesocarp | Endocarp | |||||||
|---|---|---|---|---|---|---|---|---|---|
| CTR | RWS | p-Value | CTR | RWS | p-Value | CTR | RWS | p-Value | |
| Dry matter (% of FM) | 6.73 | 6.78 | 0.5601 | 4.86 | 4.89 | 0.7417 | 4.86 | 5.00 | 0.2588 |
| Sugars and polyols | |||||||||
| Glucose (mg/g) | 28.5 | 30.0 | 0.5997 | 271.4 | 275.6 | 0.2227 | 252.3 | 260.7 | 0.0092 ** |
| Fructose (mg/g) | 35.9 | 36.2 | 0.9084 | 288.1 | 288.3 | 0.9307 | 270.3 | 274.8 | 0.0201 * |
| Polyols total (mg/g) | 4.9 | 5.4 | 0.0383 * | 7.0 | 7.5 | 0.0288 * | 9.0 | 9.2 | 0.2718 |
| Inositol (mg/g) | 2.9 | 3.4 | 0.0245 * | 6.4 | 6.9 | 0.0354 * | 7.9 | 8.2 | 0.2135 |
| Glycerol (mg/g) | 1.3 | 1.3 | 0.8460 | 0.5 | 0.6 | 0.0805 | 1.0 | 1.0 | 0.4950 |
| Erythritol (µg/g) | 770 | 678 | 0.1298 | 90 | 73 | 0.0689 | 30 | 35 | 0.6042 |
| Organic acids | |||||||||
| Total acidity (mg/g) | 24.6 | 15.4 | 0.0005 *** | 13.4 | 14.5 | 0.7520 | 26.4 | 24.5 | 0.6365 |
| Malic acid (mg/g) | 19.2 | 28.5 | 0.0007 *** | 26.4 | 39.9 | 0.0002 *** | 47.4 | 53.5 | 0.0027 ** |
| Citric acid (mg/g) | 3.9 | 3.3 | 0.1001 | 2.3 | 3.2 | 0.1350 | 0.2 | 1.1 | 0.0009 *** |
| Fumaric acid (mg/g) | 3.5 | 4.1 | 0.0495 * | 3.9 | 0.8 | 0.0018 ** | 4.5 | 2.9 | 0.4826 |
| Anions | |||||||||
| Nitrate (mg/g) | 2.9 | 2.1 | 0.1577 | 1.1 | 0.9 | 0.3363 | nd | nd | 0.3762 |
| Phosphate (mg/g) | 15.0 | 18.6 | 0.0058 ** | 7.4 | 10.1 | 0.0299 * | 19.7 | 18.6 | 0.6379 |
| Sulfate (mg/g) | 12.1 | 14.5 | 0.0061 ** | 4.7 | 4.7 | 0.9687 | 4.7 | 9.3 | 0.0446 * |
| Chloride (mg/g) | 9.7 | 9.2 | 0.0226 * | 3.3 | 3.8 | 0.0410 * | 6.1 | 5.7 | 0.4458 |
| Elements | |||||||||
| Carbon (mg/g) | 407.7 | 394.5 | 0.0547 | 387.2 | 381.5 | 0.0020 ** | 389.8 | 387.1 | 0.0035 ** |
| Macronutrients | |||||||||
| Potassium (mg/g) | 48.98 | 54.60 | 0.0016 ** | 19.40 | 23.89 | 0.0002 *** | 33.92 | 35.39 | 0.0764 |
| Nitrogen (mg/g) | 43.84 | 36.23 | 0.0052 ** | 20.23 | 15.43 | 0.0037 ** | 30.46 | 26.98 | 0.0011 ** |
| Phosphorous (mg/g) | 7.79 | 8.99 | 0.0146 * | 3.44 | 4.05 | 0.0027 ** | 6.71 | 6.78 | 0.1434 |
| Calcium (mg/g) | 7.66 | 8.00 | 0.1574 | 4.86 | 4.88 | 0.8896 | 1.58 | 1.76 | 0.0210 * |
| Magnesium (mg/g) | 2.86 | 3.01 | 0.2182 | 2.46 | 2.57 | 0.0390 * | 2.13 | 2.16 | 0.4860 |
| Micronutrients | |||||||||
| Iron (µg/g) | 86 | 87 | 0.9618 | 43 | 51 | 0.3444 | 47 | 40 | 0.3233 |
| Zinc (µg/g) | 43 | 36 | 0.0039 ** | 26 | 20 | 0.0056 ** | 40 | 35 | 0.0047 ** |
| Manganese (µg/g) | 46 | 51 | 0.2572 | 16 | 18 | 0.1112 | 18 | 21 | 0.0175 * |
| Copper (µg/g) | 16 | 12 | 0.0004 *** | 6 | 5 | 0.1965 | 10 | 7 | 0.0166 * |
| Phenolic compounds | |||||||||
| Total phenols (mg/g) | 4.0 | 3.7 | 0.0219 * | 3.2 | 3.8 | 0.0572 | 6.1 | 5.7 | 0.4458 |
| Carotenoids | |||||||||
| Violaxanthin (µg/g) | 16 | 13 | 0.0577 | nd | nd | - | nd | nd | - |
| Neoxanthin (µg/g) | 75 | 56 | 0.0215 * | nd | nd | - | nd | nd | - |
| Lutein (µg/g) | 380 | 274 | 0.0105 * | nd | nd | - | nd | nd | - |
| ß-Carotene (µg/g) | 166 | 135 | 0.0110 * | nd | nd | - | nd | nd | - |
| Total carotenoids (µg/g) | 637 | 477 | 0.0111 * | nd | nd | - | nd | nd | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schlering, C.; Schweiggert, R.; Dietrich, H.; Frisch, M.; Zinkernagel, J. Effects of Moderately-Reduced Water Supply and Picking Time on the Chemical Composition of Pickling Cucumber (Cucumis sativus L.) in Open Field Cultivation. Agronomy 2020, 10, 1097. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10081097
Schlering C, Schweiggert R, Dietrich H, Frisch M, Zinkernagel J. Effects of Moderately-Reduced Water Supply and Picking Time on the Chemical Composition of Pickling Cucumber (Cucumis sativus L.) in Open Field Cultivation. Agronomy. 2020; 10(8):1097. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10081097
Chicago/Turabian StyleSchlering, Christine, Ralf Schweiggert, Helmut Dietrich, Matthias Frisch, and Jana Zinkernagel. 2020. "Effects of Moderately-Reduced Water Supply and Picking Time on the Chemical Composition of Pickling Cucumber (Cucumis sativus L.) in Open Field Cultivation" Agronomy 10, no. 8: 1097. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10081097