Plant Yield Efficiency by Homeostasis as Selection Tool at Ultra-Low Density. A Comparative Study with Common Stability Measures in Maize

 , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Procedure
2.2. Data Analysis
2.2.1. Estimation of PYE, PYH, and PYEH
2.2.2. Attainable Crop Yield and Stability Statistics
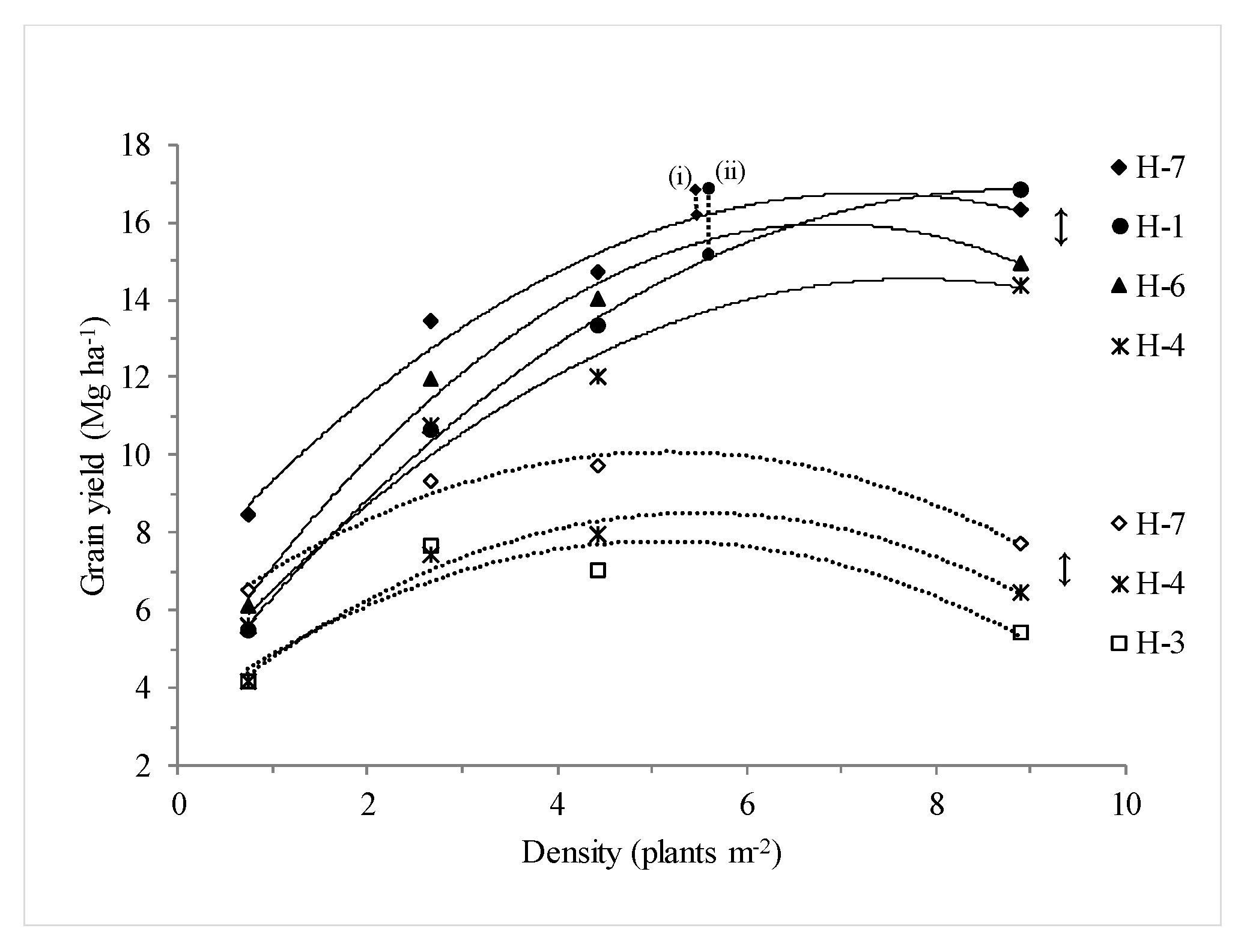
3. Results
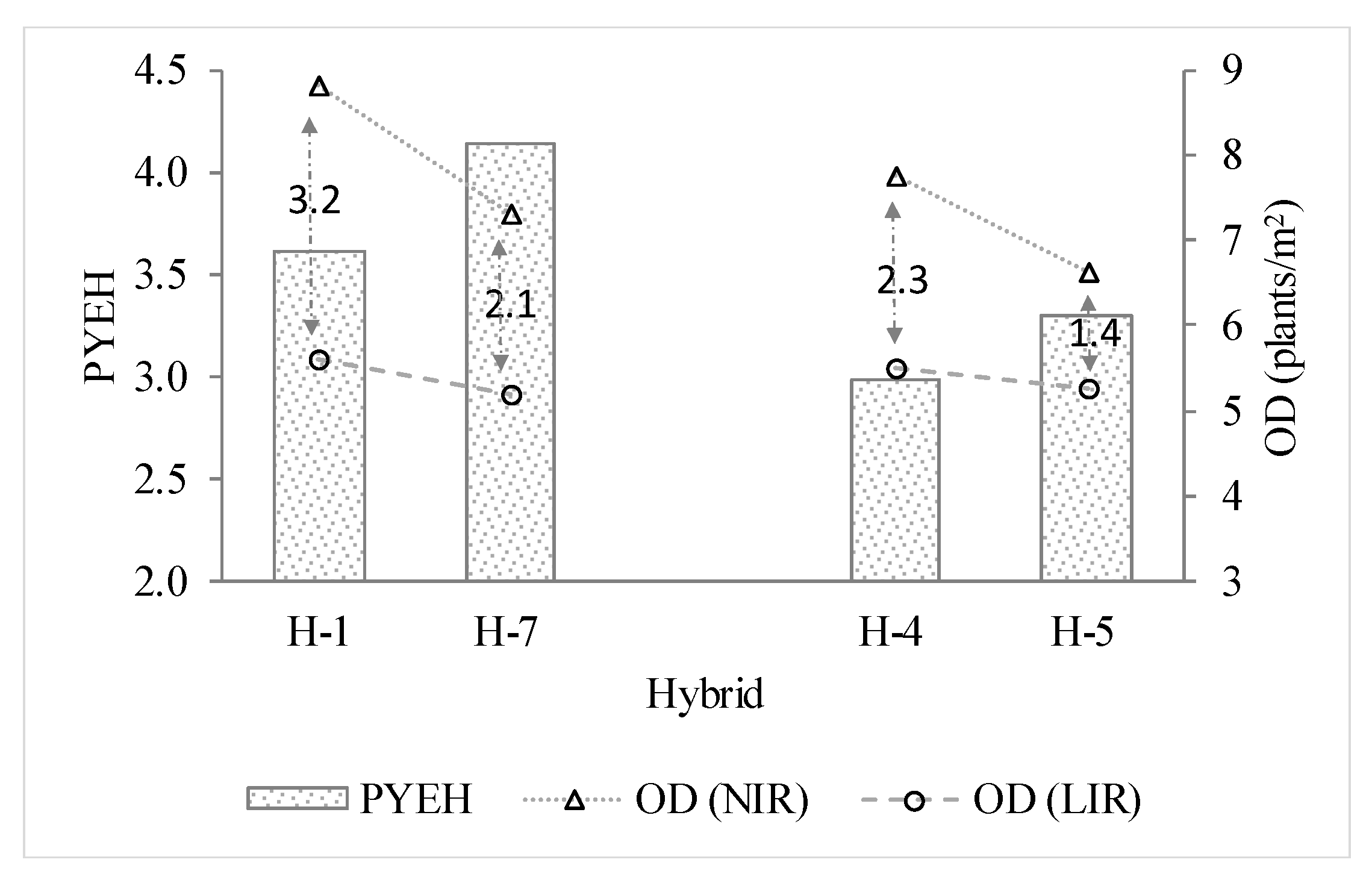
3.1. Single-Plant Performance for Yield and Homeostasis
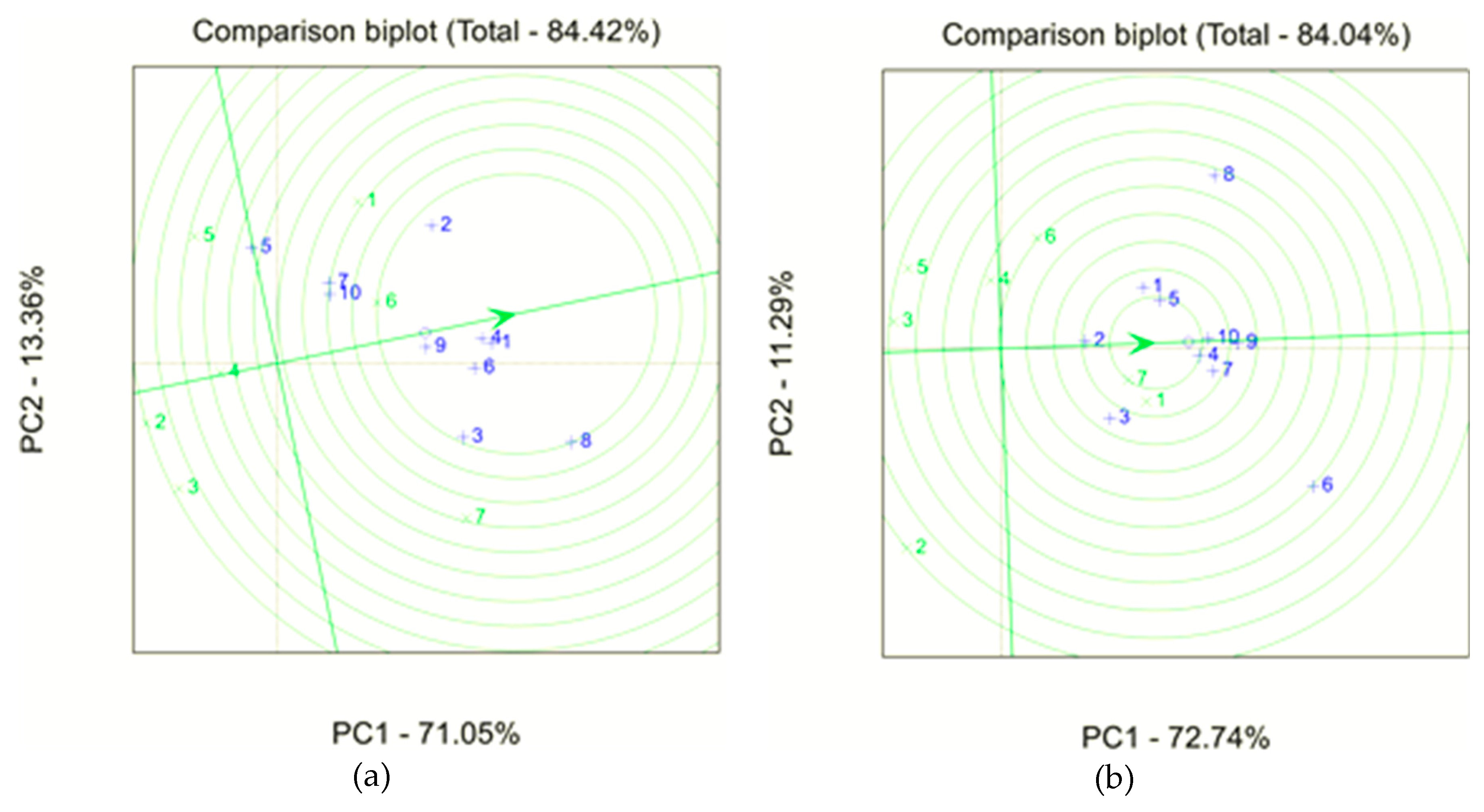
3.2. Crop Attainable Yield and Ranking Based on Stability
3.3. Breeding Density Versus Dense Stand
4. Discussion
4.1. Genotype Ranking on Stability Measures
4.2. The Prediction Value of the PYEH for Yield and Stability Across Densities
4.3. Implications of Breeding at the BD
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Baxevanos, D.; Goulas, C.; Tzortzios, S.; Mavromatis, A. Interrelationship among and repeatability of seven stability indices estimated from commercial cotton (Gossypium hirsutum L.) variety evaluation trials in three Mediterranean countries. Euphytica 2008, 161, 371–382. [Google Scholar] [CrossRef]
- Basu, S.K.; Acharya, S.N.; Bandara, M.S.; Friebel, D.; Thomas, J.E. Effects of genotype and environment on seed and forage yield in fenugreek (Trigonella foenum-graecum L.) grown in western Canada. Aust. J. Crop Sci. 2009, 3, 305–314. [Google Scholar]
- Akcura, M.; Kaya, Y.; Taner, S.; Ayranci, R. Parametric stability analyses for grain yield of durum wheat. Plant Soil Environ. 2006, 52, 254–261. [Google Scholar] [CrossRef] [Green Version]
- Sabaghnia, N.; Dehghani, H.; Sabaghpour, S.H. Nonparametric methods for interpreting genotype x environment interaction of lentil genotypes. Crop Sci. 2006, 46, 1100–1106. [Google Scholar] [CrossRef]
- Solomon, K.F.; Smit, H.A.; Malan, E.; Du Toit, W.J. Comparison study using rank based nonparametric stability statistics of durum wheat. World J. Agric. Sci. 2007, 3, 444–450. [Google Scholar]
- Fikere, M.; Tadesse, T.; Gebeyehu, S.; Hundie, B. Agronomic performances, disease reaction and yield stability of field pea (Pisum sativum L.) genotypes in Bale Highlands, Ethiopia. Aust. J. Crop Sci. 2010, 4, 238–246. [Google Scholar]
- Mohammadi, R.; Amri, A. Comparison of parametric and non-parametric methods for selecting stable and adapted durum wheat genotypes in variable environments. Euphytica 2008, 159, 419–432. [Google Scholar] [CrossRef]
- Hühn, M.; Leon, J. Non-parametric analysis of cultivar performance trials: Experimental results and comparison of different procedures based on ranks. Agron. J. 1995, 87, 627–632. [Google Scholar] [CrossRef]
- Finlay, K.W.; Wilkinson, G.N. The analysis of adaptation in a plant breeding programme. Aust. J. Agric. Res. 1963, 14, 742–754. [Google Scholar] [CrossRef] [Green Version]
- Eberhart, S.A.; Russell, W.A. Stability parameters for comparing varieties. Crop Sci. 1966, 6, 36–40. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.S.; Binns, M.R.; Lefkovitch, L.P. Stability analysis: Where do we stand? Crop Sci. 1986, 26, 894–900. [Google Scholar] [CrossRef] [Green Version]
- Becker, H.C.; Leon, J. Stability analysis in plant breeding. Plant Breed 1988, 101, 1–23. [Google Scholar] [CrossRef]
- Shukla, G.K. Some statistical aspects of partitioning genotype–environmental components of variability. Heredity 1972, 29, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Francis, T.R.; Kannenberg, L.W. Yield stability studied in short-season maize: I. A descriptive method for grouping genotypes. Can. J. Plant Sci. 1978, 58, 1029–1034. [Google Scholar] [CrossRef]
- Lin, C.S.; Binns, M.R. A method of analyzing cultivar x location x year experiments: A new stability parameter. Theor. Appl. Genet. 1988, 76, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Nassar, R.; Huehn, M. Studies on estimation of phenotypic stability: Tests of significance for nonparametric measures of phenotypic stability. Biometrics 1987, 43, 45–53. [Google Scholar] [CrossRef]
- Fox, P.N.; Skovmand, B.; Thompson, B.K.; Braun, H.J.; Cormier, R. Yield and adaptation of hexaploid spring triticale. Euphytica 1990, 47, 57–64. [Google Scholar] [CrossRef]
- Kang, M.S.; Pham, H.N. Simultaneous selection for high yielding and stable crop genotypes. Agron. J. 1991, 83, 161–165. [Google Scholar] [CrossRef]
- Gauch, H.G. Statistical analysis of yield trials by AMMI and GGE. Crop Sci. 2006, 46, 1488–1500. [Google Scholar] [CrossRef]
- Yan, W. Singular-value partition in biplot analysis of multienvironment trial data. Agron. J. 2002, 94, 990–996. [Google Scholar] [CrossRef] [Green Version]
- Purchase, J.L.; Hatting, H.; Van Deventer, C.S. Genotype × environment interaction of winter wheat in South Africa: II. Stability analysis of yield performance. S. Afr. J. Plant Soil 2000, 17, 101–107. [Google Scholar] [CrossRef]
- Tokatlidis, I.S. Adapting maize crop to climate change. Agron. Sustain. Dev. 2013, 33, 63–79. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Liu, C.; Wang, K.; Ruizhi, X.; Ming, B.; Wang, Y.; Zhang, G.; Liu, G.; Zhao, R.; Fan, P.; et al. Adjusting maize plant density to different climatic conditions across a large longitudinal distance in China. Field Crops Res. 2017, 212, 126–134. [Google Scholar] [CrossRef]
- Seyoum, S.; Rachaputi, R.; Fekybelu, S.; Chauhana, Y.; Prasanna, B. Exploiting genotype x environment x management interactions to enhance maize productivity in Ethiopia. Eur. J. Agron. 2019, 103, 165–174. [Google Scholar] [CrossRef]
- Duvick, D.N. The contribution of breeding to yield advances in maize (Zea mays L.). Adv. Agron. 2005, 86, 83–145. [Google Scholar]
- Brekke, B.; Edwards, J.; Knapp, A. Selection and adaptation to high plant density in the Iowa stiff stalk synthetic maize (Zea mays L.) population. Crop Sci. 2011, 51, 1965–1972. [Google Scholar] [CrossRef]
- Tokatlidis, I.S.; Dordas, C.; Papathanasiou, F.; Papadopoulos, I.; Pankou, C.; Gekas, F.; Ninou, E.; Mylonas, I.; Tzantarmas, C.; Petrevska, J.K.; et al. Improved plant yield efficiency is essential for maize rainfed production. Agron. J. 2015, 107, 1011–1018. [Google Scholar] [CrossRef] [Green Version]
- Mylonas, I.; Sinapidou, E.; Remountakis, E.; Sistanis, I.; Pankou, C.; Ninou, E.; Papadopoulos, I.; Papathanasiou, F.; Lithourgidis, A.; Gekas, F.; et al. Improved plant yield efficiency alleviates the erratic optimum density in maize. Agron. J. 2020, 112, 1690–1701. [Google Scholar] [CrossRef]
- Ciampitti, I.A.; Vyn, T.J. Understanding global and historical nutrient use efficiencies for closing maize yield gaps. Agron. J. 2014, 106, 2107–2117. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, V.H.; Tollenaar, M.; Bowman, A.; Good, B.; Lee, E.A. Maize yield potential and density tolerance. Crop Sci. 2018, 58, 472–485. [Google Scholar] [CrossRef]
- Assefa, Y.; Carter, P.; Hinds, M.; Bhalla, G.; Schon, R.; Jeschke, M.; Paszkiewicz, S.; Smith, S.; Ciampitti, I.A. Analysis of long term study indicates both agronomic optimal plant density and increase maize yield per plant contributed to yield gain. Sci. Rep. 2018, 8, 4937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berzsenyi, Z.; Tokatlidis, I.S. Density-dependence rather maturity determines hybrid selection in dryland maize production. Agron. J. 2012, 104, 331–336. [Google Scholar] [CrossRef]
- Fasoula, V.A.; Tokatlidis, I.S. Development of crop cultivars by honeycomb breeding. Agron. Sustain. Dev. 2012, 32, 161–180. [Google Scholar] [CrossRef] [Green Version]
- Tokatlidis, I.S. Crop adaptation to density to optimise grain yield: Breeding implications. Euphytica 2017, 213, 92. [Google Scholar] [CrossRef]
- Fasoulas, A.C.; Fasoula, V.A. Honeycomb selection designs. Plant Breed Rev. 1995, 13, 87–139. [Google Scholar]
- Qian, C.; Yu, Y.; Gong, X.; Jiang, Y.; Zhao, Y.; Yang, Z.; Hao, Y.; Li, L.; Song, Z.; Zhang, W. Response of grain yield to plant density and nitrogen rate in spring maize hybrids released from 1970 to 2010 in Northeast China. Crop J. 2013, 4, 459–467. [Google Scholar] [CrossRef] [Green Version]
- Amelong, A.; Hernández, F.; Novoa, A.D.; Borrás, L. Maize stand density yield response of parental inbred lines and derived hybrids. Crop Sci. 2017, 57, 32–39. [Google Scholar] [CrossRef]
- Thennarasu, K. On Certain Non-Parametric Procedures for Studying Genotype-Environment Interactions and Yield Stability. Ph.D. Thesis, P.J. School, IARI, New Delhi, India, 1995. [Google Scholar]
- Payne, R.W. Genstat. Wiley Interdis. Rev. Comp. Stat. 2009, 1, 255–258. [Google Scholar] [CrossRef]
- Solomon, K.F.; Chauhan, Y.; Zeppa, A. Risks of yield loss due to variation in optimum density for different maize genotypes under variable environmental conditions. J. Agron. Crop Sci. 2017, 203, 519–527. [Google Scholar] [CrossRef]
- Meza, F.J.; Silva, D.; Vigil, H. Climate change impacts on irrigated maize in Mediterranean climates: Evaluation of double cropping as an emerging adaptation alternative. Agr. Syst. 2008, 98, 21–30. [Google Scholar] [CrossRef]
- Nájera, F.; Tapia, Y.; Baginsky, C.; Figueroa, V.; Cabeza, R.; Salazar, O. Evaluation of soil fertility and fertilisation practices for irrigated maize (Zea mays L.) under Mediterranean conditions in central Chile. J. Soil Sci. Plant Nut. 2015, 15, 84–97. [Google Scholar] [CrossRef] [Green Version]
- Maresma, A.; Ballesta, A.; Santiveri, F.; Lloveras, J. Sowing date affects maize development and yield in irrigated Mediterranean environments. Agriculture 2019, 9, 67. [Google Scholar] [CrossRef] [Green Version]
- Papadopoulos, I.I.; Tokatlidis, I.S. A novel statistic estimated in the absence of competition to foresee genotype performance at the farming conditions parallels the agronomic concept of stability. Austr. J. Crop Sci. 2011, 5, 822–830. [Google Scholar]
- Kebede, A.; Getahun, A. Adaptability and Stability Analysis of Groundnut Genotypes Using AMMI Model and GGE-biplot. J. Crop Sci. Biotech. 2017, 20, 343–349. [Google Scholar] [CrossRef]
- Mekbib, F. Yield stability in common bean (Phaseoulus vulgaris L.) genotypes. Euphytica 2003, 130, 147–153. [Google Scholar] [CrossRef]
- Jalaluddin, M.D.; Harrison, S.A. Repeatability of stability statistics for grain yield in wheat. Crop Sci. 1993, 33, 720–725. [Google Scholar] [CrossRef]
- Mendes, M.H.S.; Ramalho, M.A.P. Repeatability of some phenotypic stability parameters - a resampling approach. Crop Breed Appl. Biot. 2018, 18, 139–147. [Google Scholar] [CrossRef] [Green Version]
- Dhaliwal, D.S.; Williams, M.M. Understanding variability in optimum plant density and recommendation domains for crowding stress tolerant processing sweet corn. PLoS ONE 2020, 15. [Google Scholar] [CrossRef] [Green Version]
- Sabaghnia, N.; Dehghani, H.; Alizadeh, B.; Mohghaddam, M. Diallele analysis of oil content and some agronomic traits in rapeseed (Brassica napus L.) based on the additive-dominance genetic model. Aust. J. Crop Sci. 2010, 4, 609–616. [Google Scholar]
- Romagosa, I.; Fox, P.N. Genotype × environment interaction and adaptation. In Plant breeding: Principles and Prospects; Hayward, M.D., Bosemark, N.O., Romagosa, I., Eds.; Chapman and Hall: London, UK, 1993; pp. 373–390. [Google Scholar]
- Baker, R.J. Tests for crossover genotype–environment interactions. Can J. Plant Sci. 1988, 68, 405–410. [Google Scholar] [CrossRef]
- Rosielle, A.A.; Hamblin, J. Theoretical aspects of selection for yield in stress and non-stress environments. Crop Sci. 1981, 21, 943–946. [Google Scholar] [CrossRef]
- Mansfield, B.D.; Mumm, R.H. Survey of plant density tolerance in U.S. maize germplasm. Crop Sci. 2014, 54, 157–173. [Google Scholar] [CrossRef] [Green Version]
- Ntanos, D.A.; Roupakias, D.G. Comparative efficiency of two breeding methods for yield and quality in rice. Crop Sci. 2001, 41, 345–350. [Google Scholar] [CrossRef]
- Tokatlidis, I.S.; Xynias, I.N.; Tsialtas, J.T.; Papadopoulos, I.I. Single-plant selection at ultra-low density to improve stability of a bread wheat cultivar. Crop Sci. 2006, 46, 90–97. [Google Scholar] [CrossRef]
- Fasoula, V.A.; Boerma, H.R. Intra-cultivar variation for seed weight and agronomic traits. Crop Sci. 2007, 47, 367–373. [Google Scholar] [CrossRef]
- Vlachostergios, D.N.; Lithourgidis, A.S.; Roupakias, D.G. Effectiveness of single plant selection at low density under organic environment. A field study with lentil. Crop Sci. 2011, 51, 41–51. [Google Scholar] [CrossRef]
- Hansen, K.A.; Martin, J.M.; Lanning, S.P.; Talbert, L.E. Correlation of genotype performance for agronomic and physiological traits in space-planted versus densely seeded conditions. Crop Sci. 2005, 45, 1023–1028. [Google Scholar] [CrossRef]
- Liu, W.; Tollenaar, M. Response of yield heterosis to increasing plant density in maize. Crop Sci. 2009, 49, 1807–1816. [Google Scholar] [CrossRef]
- Martin, K.L.; Hodgen, P.J.; Freeman, K.W.; Melchiori, R.; Arnall, D.B.; Teal, R.K.; Mullen, R.W.; Desta, K.; Phillips, S.B.; Solie, J.B.; et al. Plant-to-plant variability in corn production. Agron. J. 2005, 97, 1603–1611. [Google Scholar] [CrossRef] [Green Version]
- Tokatlidis, I.S.; Remountakis, E. The Impacts of interplant variation on aboveground biomass, grain yield, and harvest index in maize. Inter. J. Plant Prod. 2020, 14, 57–65. [Google Scholar] [CrossRef]
- Zhai, L.; Xie, R.; Ming, B.; Li, S.; Ma, D. Evaluation and analysis of intraspecific competition in maize: A case study on plant density experiment. J. Integr. Agric. 2018, 17, 2235–2244. [Google Scholar] [CrossRef] [Green Version]
- Shao, H.; Shi, D.; Shi, W.; Ban, X.; Chen, Y.; Ren, W.; Chen, F.; Mi, G. Genotypic difference in the plasticity of root system architecture of field-grown maize in response to plant density. Plant and Soil 2019, 439, 201–217. [Google Scholar] [CrossRef]
- Ross, F.; Di Matteo, J.; Cerrudo, A. Maize prolificacy: A source of reproductive plasticity that contributes to yield stability when plant population varies in drought-prone environments. Field Crops Res. 2020, 247, 107699. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hybrid | Quadratic Equation | OD | ACY 1 | Quadratic Equation | OD | ACY |
|---|---|---|---|---|---|---|
| NIR | LIR | |||||
| H-1 | Y = −0.17x2 + 3.03x + 3.48 (0.99) | 8.81 | 16.82 a | Y = −0.21x2 + 2.34x + 3.21 (0.97) | 5.61 | 9.77 a |
| H-2 | Y = −0.21x2 + 2.95x + 3.23 (0.96) | 7.00 | 13.56 d | Y = −0.19x2 + 2.00x + 2.68 (0.83) | 5.27 | 7.94 c |
| H-3 | Y = −0.20x2 + 2.76x + 3.72 (0.97) | 7.05 | 13.46 d | Y = −0.17x2 + 1.75x + 3.31 (0.82) | 5.11 | 7.78 c |
| H-4 | Y = −0.17x2 + 2.72x + 3.99 (0.97) | 7.77 | 14.54 c | Y = −0.18x2 + 2.01x + 2.97 (0.96) | 5.51 | 8.51 b |
| H-5 | Y = −0.27x2 + 3.57x + 2.55 (0.99) | 6.64 | 14.38 c | Y = −0.21x2 + 2.26x + 2.33 (0.97) | 5.26 | 8.28 bc |
| H-6 | Y = −0.25x2 + 3.50x + 3.90 (0.98) | 6.89 | 15.95 b | Y = −0.22x2 + 2.32x + 3.69 (0.98) | 5.25 | 9.79 a |
| H-7 | Y = −0.19x2 + 2.71x + 6.82 (0.97) | 7.32 | 16.74 a | Y = −0.17x2 + 1.80x + 5.42 (0.97) | 5.18 | 10.08 a |
| LSD | 0.48 | 0.46 | ||||
| Hybrid | GGE | ASVi | Pi | bi/=1 | bi/>1 | aCVi | Si | NPi | TMB | Ii | SUM | 1st | 2nd | 3rd | 4th | 5th | 6th | 7th | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LCD | |||||||||||||||||||||
| H-1 | 2 | 5 | 3 | 7 | 7 | 5 | 1 | 5 | 5 | 5 | 5 | 3 | 3 | 56 | 1 | 1 | 3 | 0 | 6 | 0 | 2 |
| H-2 | 7 | 4 | 7 | 2 | 5 | 4 | 6 | 6 | 4 | 2 | 3 | 6 | 7 | 63 | 0 | 2 | 1 | 3 | 1 | 3 | 3 |
| H-3 | 6 | 1 | 6 | 1 | 4 | 1 | 5 | 2 | 1 | 1 | 1 | 7 | 6 | 42 | 6 | 1 | 0 | 1 | 1 | 3 | 1 |
| H-4 | 4 | 2 | 4 | 4 | 6 | 2 | 3 | 1 | 2 | 3 | 2 | 5 | 5 | 43 | 1 | 4 | 2 | 3 | 2 | 1 | 0 |
| H-5 | 5 | 6 | 5 | 5 | 2 | 6 | 7 | 4 | 6 | 6 | 4 | 4 | 4 | 64 | 0 | 1 | 0 | 4 | 3 | 4 | 1 |
| H-6 | 1 | 3 | 2 | 3 | 3 | 3 | 2 | 3 | 3 | 4 | 6 | 1 | 2 | 36 | 2 | 3 | 6 | 1 | 0 | 1 | 0 |
| H-7 | 3 | 7 | 1 | 6 | 1 | 7 | 4 | 7 | 7 | 7 | 7 | 2 | 1 | 60 | 3 | 1 | 1 | 1 | 0 | 1 | 6 |
| Range | 0.07–6.35 | 1.21–8.35 | 0.01–0.14 | 0.86–1.10 | 0.05–3.97 | 0.22–0.31 | 0.68–5.82 | 0.45–1.66 | 0.67–6.55 | 0.20–1.26 | 12–27 | 10–25 | |||||||||
| HCD | |||||||||||||||||||||
| H-1 | 2 | 7 | 2 | 7 | 1 | 6 | 4 | 7 | 2 | 7 | 6 | 2 | 3 | 56 | 1 | 4 | 1 | 1 | 0 | 2 | 4 |
| H-2 | 7 | 6 | 6 | 6 | 7 | 7 | 5 | 5 | 7 | 2 | 1 | 6 | 6 | 71 | 1 | 1 | 0 | 0 | 2 | 5 | 4 |
| H-3 | 6 | 2 | 7 | 4 | 6 | 5 | 6 | 4 | 4 | 3 | 3 | 5 | 7 | 62 | 0 | 1 | 2 | 3 | 2 | 3 | 2 |
| H-4 | 4 | 5 | 4 | 2 | 4 | 3 | 3 | 1 | 5 | 4 | 5 | 4 | 4 | 48 | 1 | 1 | 2 | 6 | 3 | 0 | 0 |
| H-5 | 5 | 3 | 5 | 3 | 5 | 2 | 7 | 3 | 3 | 1 | 2 | 7 | 5 | 51 | 1 | 2 | 4 | 0 | 4 | 0 | 2 |
| H-6( | 3 | 4 | 3 | 1 | 3 | 4 | 2 | 2 | 6 | 5 | 4 | 3 | 2 | 42 | 1 | 3 | 4 | 3 | 1 | 1 | 0 |
| H-7 | 1 | 1 | 1 | 5 | 2 | 1 | 1 | 6 | 1 | 6 | 7 | 1 | 1 | 34 | 8 | 1 | 0 | 0 | 1 | 2 | 1 |
| Range | 0.81–1.47 | 0.14–6.78 | 0.01–0.17 | 0.89–1.17 | 0.58–1.46 | 0.39–0.45 | 0.86–4.18 | 0.39–1.10 | 1.45–4.74 | 0.33–1.19 | 14–28 | 5–25 | |||||||||
| Site2(NI) | Site3(NI) | Site4(NI) | Site5(NI) | Site1(LI) | Site2(LI) | Site3(LI) | Site4(LI) | Site5(LI) | |
|---|---|---|---|---|---|---|---|---|---|
| BD (0.74 plants/m2) | |||||||||
| Site1(NI) | 0.90 ** | 0.93 ** | 0.94 ** | 0.86 * | 0.79 * | 0.91 ** | 0.94 ** | 0.94 ** | 0.81 * |
| Site2(NI) | 0.82 * | 0.92 ** | 0.91 ** | 0.84 * | 0.88 ** | 0.91 ** | 0.95 ** | 0.82 * | |
| Site3(NI) | 0.86 * | 0.96 *** | 0.78 * | 0.97 *** | 0.88 ** | 0.96 *** | 0.85 * | ||
| Site4(NI) | 0.89 ** | 0.87 * | 0.92 ** | 0.90 ** | 0.76 * | 0.92 ** | |||
| Site5(NI) | 0.91 ** | 0.96 *** | 0.94 ** | 0.90 ** | 0.88 ** | ||||
| Site1(LI) | 0.89 ** | 0.93 ** | 0.84 * | 0.79 * | |||||
| Site2(LI) | 0.87 * | 0.88 ** | 0.75 | ||||||
| Site3(LI) | 0.86 * | 0.87 * | |||||||
| Site4(LI) | 0.95 *** | ||||||||
| LCD (4.44 plants/m2) | |||||||||
| Site1(NI) | 0.33 | 0.16 | −0.26 | 0.95 ** | 0.82 * | −0.01 | 0.63 | 0.59 | 0.95 ** |
| Site2(NI) | −0.55 | 0.66 | −0.35 | 0.90 ** | 0.32 | −0.01 | 0.24 | 0.58 | |
| Site3(NI) | 0.79 * | 0.66 | −0.57 | 0.23 | 0.48 | 0.75 | 0.47 | ||
| Site4(NI) | 0.79 * | 0.41 | 0.39 | 0.28 | 0.88 ** | 0.79 * | |||
| Site5(NI) | 0.82 * | 0.04 | 0.20 | 0.85 * | 0.93 ** | ||||
| Site1(LI) | 0.13 | 0.46 | −0.34 | 0.90 ** | |||||
| Site2(LI) | 0.38 | 0.57 | −0.28 | ||||||
| Site3(LI) | 0.95 ** | 0.53 | |||||||
| Site4(LI) | 0.89 ** | ||||||||
| HCD (8.89 plants/m2) | |||||||||
| Site1(NI) | 0.13 | 0.49 | 0.90 ** | 0.68 | 0.69 | 0.56 | 0.68 | 0.76 * | 0.88 ** |
| Site2(NI) | 0.74 | 0.65 | 0.68 | 0.81 * | 0.80 * | 0.74 | 0.73 | 0.92 ** | |
| Site3(NI) | 0.61 | 0.26 | 0.74 | 0.61 | 0.77 * | 0.45 | 0.86 * | ||
| Site4(NI) | 0.73 | 0.25 | 0.81 * | 0.87 * | 0.81 * | 0.78 * | |||
| Site5(NI) | 0.38 | 0.22 | 0.91 ** | 0.72 | 0.89 ** | ||||
| Site1(LI) | 0.75 | 0.46 | 0.72 | 0.87 * | |||||
| Site2(LI) | 0.78 * | 0.72 | 0.92 ** | ||||||
| Site3(LI) | 0.62 | 0.51 | |||||||
| Site4(LI) | 0.73 | ||||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sinapidou, E.; Pankou, C.; Gekas, F.; Sistanis, I.; Tzantarmas, C.; Tokamani, M.; Mylonas, I.; Papadopoulos, I.; Kargiotidou, A.; Ninou, E.; et al. Plant Yield Efficiency by Homeostasis as Selection Tool at Ultra-Low Density. A Comparative Study with Common Stability Measures in Maize. Agronomy 2020, 10, 1203. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10081203
Sinapidou E, Pankou C, Gekas F, Sistanis I, Tzantarmas C, Tokamani M, Mylonas I, Papadopoulos I, Kargiotidou A, Ninou E, et al. Plant Yield Efficiency by Homeostasis as Selection Tool at Ultra-Low Density. A Comparative Study with Common Stability Measures in Maize. Agronomy. 2020; 10(8):1203. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10081203
Chicago/Turabian StyleSinapidou, Evaggelia, Chrysanthi Pankou, Fotakis Gekas, Iosif Sistanis, Constantinos Tzantarmas, Maria Tokamani, Ioannis Mylonas, Ioannis Papadopoulos, Anastasia Kargiotidou, Elissavet Ninou, and et al. 2020. "Plant Yield Efficiency by Homeostasis as Selection Tool at Ultra-Low Density. A Comparative Study with Common Stability Measures in Maize" Agronomy 10, no. 8: 1203. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10081203