Assessment of Tomato Recombinant Lines in Conventional and Organic Farming Systems for Productivity and Fruit Quality Traits

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
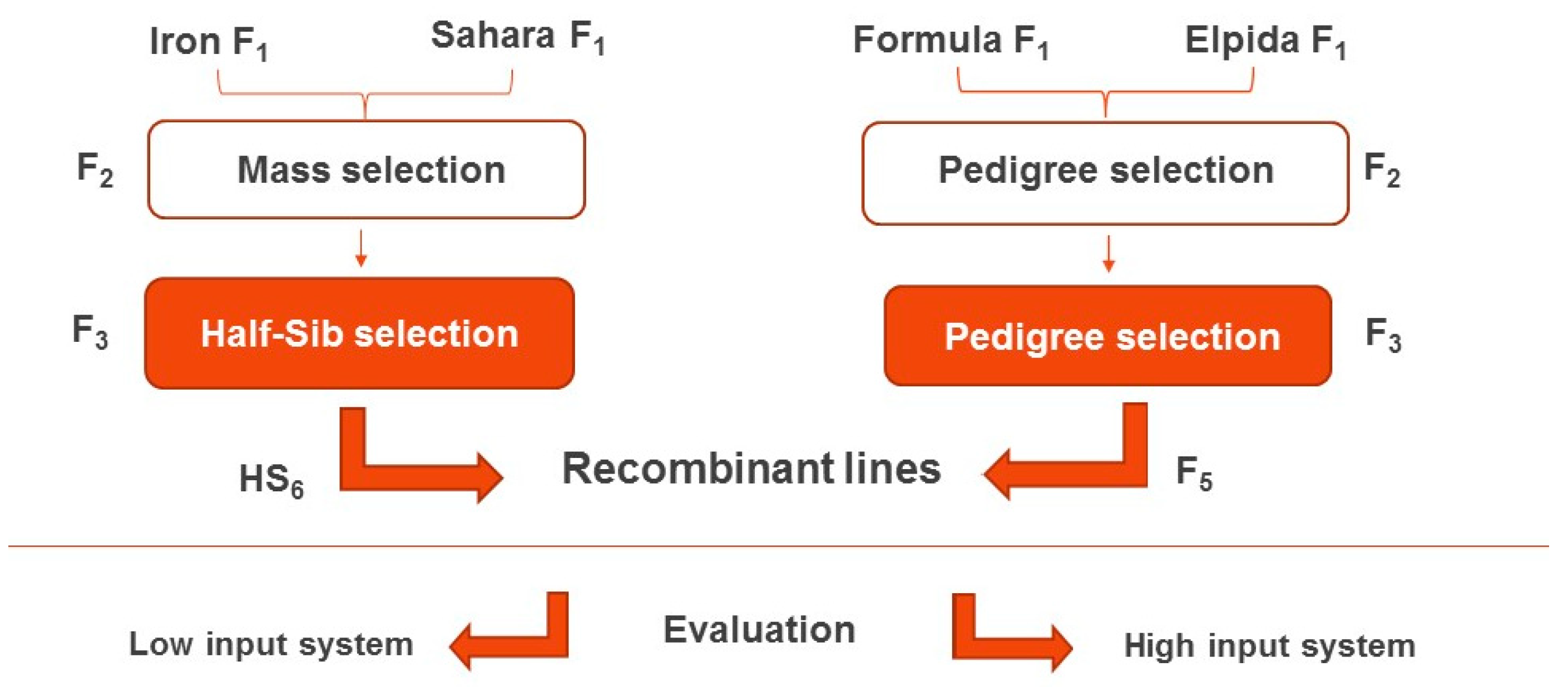
2.1. Plant Material
2.2. Methodology
2.3. Selection and Assessment Procedure
2.3.1. Farming System 1: Conventional with High-Input Conditions
2.3.2. Farming System 2: Organic (Low-Input Cultural Practices)
2.4. Traits Evaluated
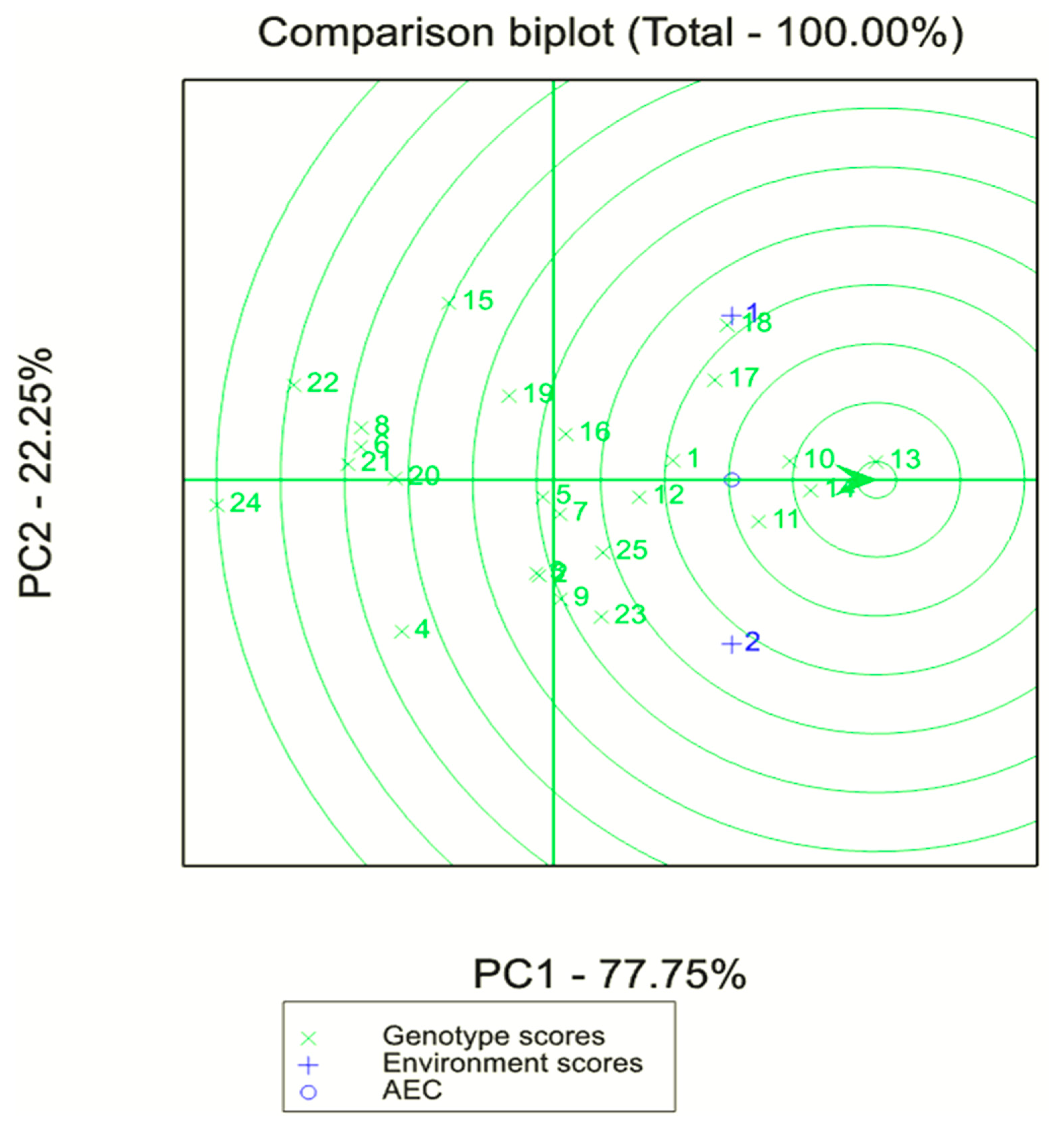
2.5. Statistical Analyses
3. Results
3.1. Farming System 1: Conventional High-Input System
3.2. Farming System 2: Low-Input System
3.3. Fruit Quality Traits
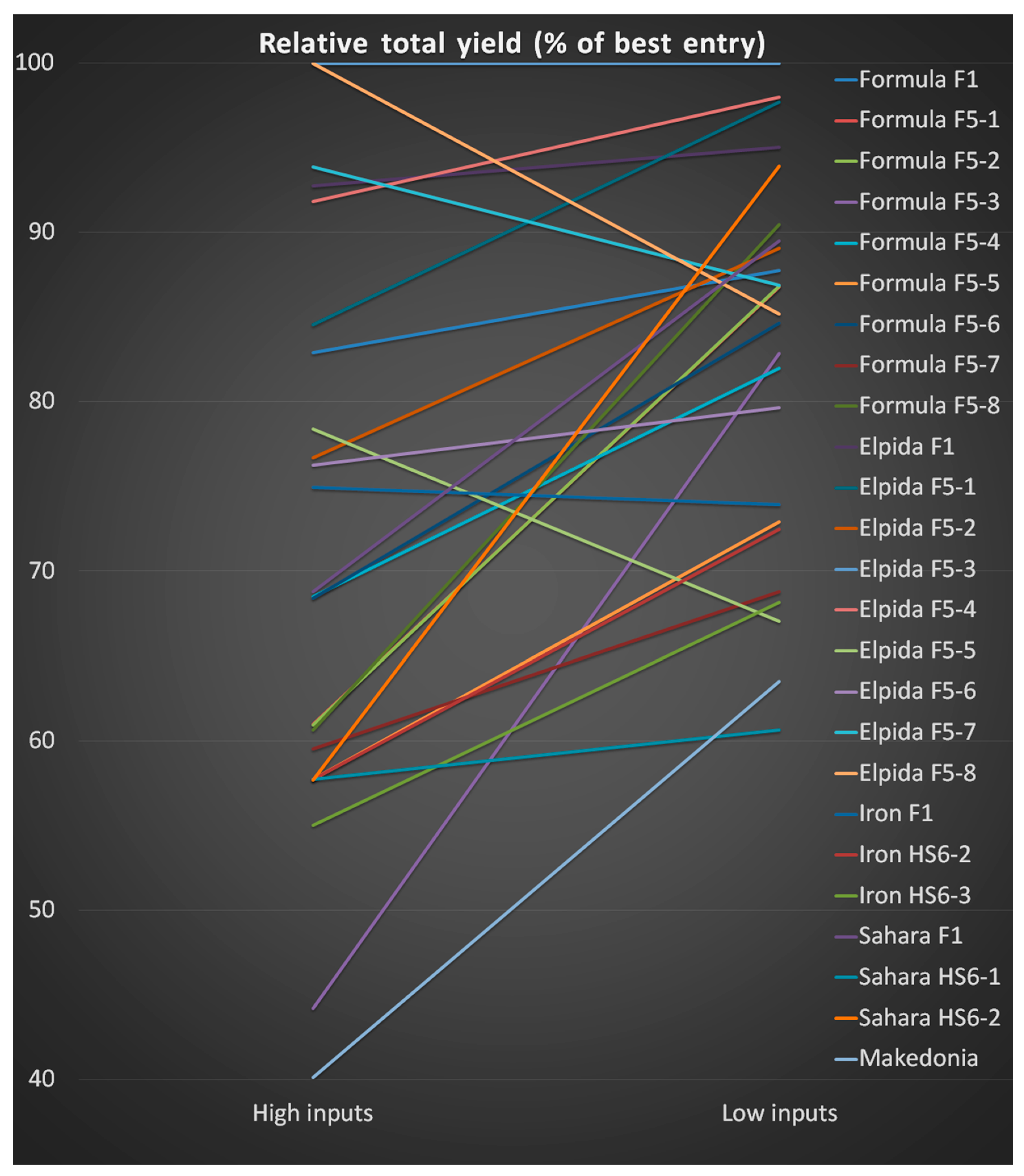
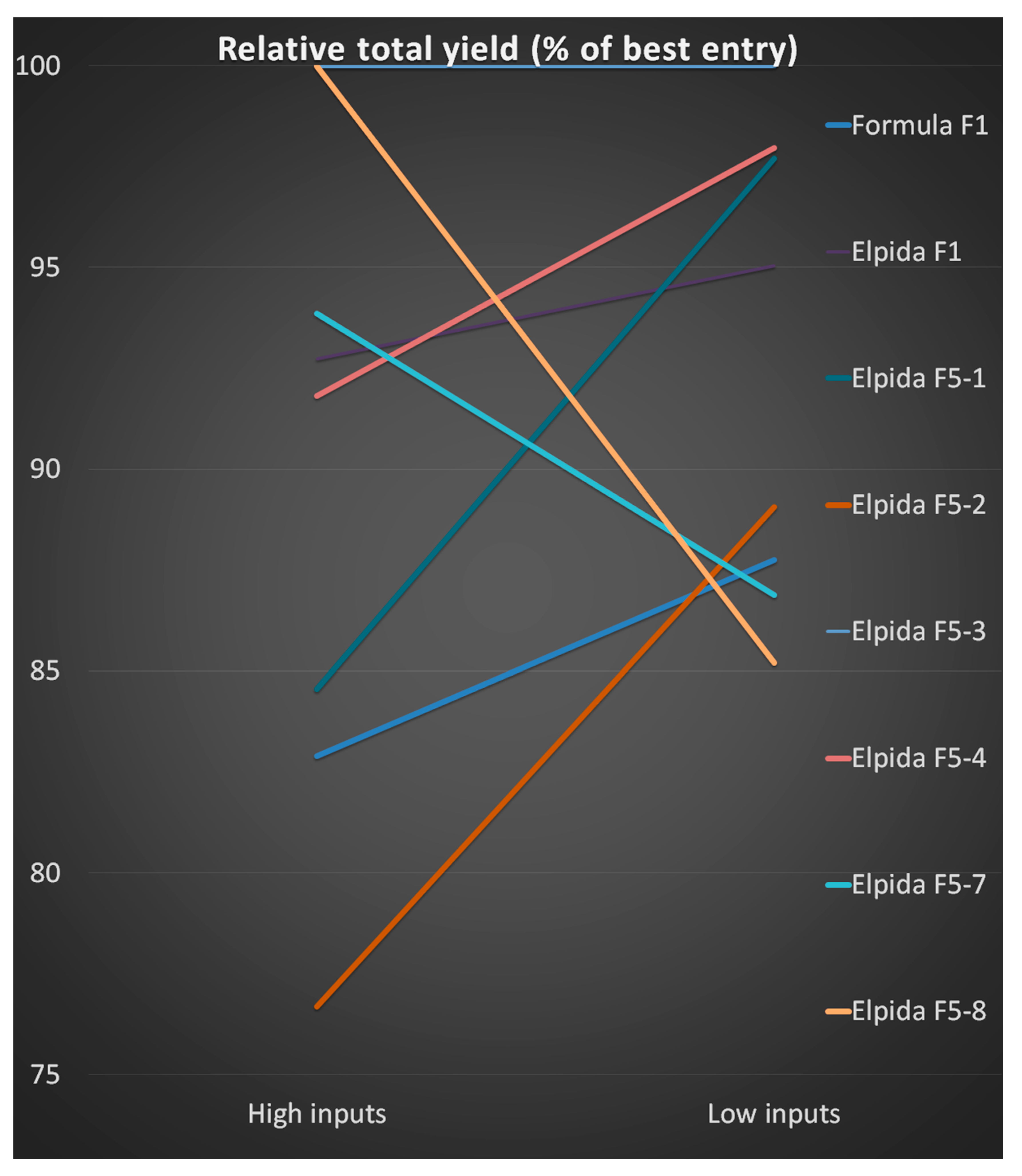
3.4. Comparing the Two Farming Systems
4. Discussion
4.1. Inbred Vigor
4.2. Methodology
4.3. Farming System
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xu, X.; Hou, L.; Lin, H.; Liu, W. Zoning of sustainable agricultural development in China. Agric. Syst. 2006, 87, 38–62. [Google Scholar] [CrossRef]
- Gafsi, M.; Legagneux, B.; Nguyen, G.; Robin, P. Towards sustainable farming systems: Effectiveness and deficiency of the French procedure of sustainable agriculture. Agric. Syst. 2006, 90, 226–242. [Google Scholar] [CrossRef]
- Van Bueren, E.T.L.; Østergård, H.; Goldringer, I.; Scholten, O. Plant breeding for organic and sustainable, low-input agriculture: Dealing with genotype–environment interactions. Euphytica 2008, 163, 321–322. [Google Scholar] [CrossRef]
- Van Bueren, E.T.L.; Jones, S.; Tamm, L.; Murphy, K.; Myers, J.; Leifert, C.; Messmer, M. The need to breed crop varieties suitable for organic farming, using wheat, tomato and broccoli as examples: A review. NJAS Wagening. J. Life Sci. 2011, 58, 193–205. [Google Scholar] [CrossRef]
- Wolfe, M.S.; Baresel, J.P.; Desclaux, D.; Goldringer, I.; Hoad, S.; Kovacs, G.; Löschenberger, F.; Miedaner, T.; Østergård, H.; Van Bueren, E.L. Developments in breeding cereals for organic agriculture. Euphytica 2008, 163, 323–346. [Google Scholar] [CrossRef] [Green Version]
- Le Campion, A.; Oury, F.-X.; Heumez, E.; Rolland, B. Conventional versus organic farming systems: Dissecting comparisons to improve cereal organic breeding strategies. Org. Agric. 2019, 10, 63–74. [Google Scholar] [CrossRef] [Green Version]
- Fasoula, V.; Tokatlidis, I. Development of crop cultivars by honeycomb breeding. Agron. Sustain. Dev. 2012, 32, 161–180. [Google Scholar] [CrossRef] [Green Version]
- Priyadarshan, P.M. Breeding Self-Pollinated Crops. In Plant Breeding: Classical to Modern; Springer: Singapore, 2019. [Google Scholar] [CrossRef]
- Kaiwart, S.; Toppo, S.; Sahu, S. Evaluation of tomato (Solanum lycopersicum L.) Hybrids for quality parameter in Allahabad agroclimatic condition. J. Pharmacogn. Phytochem. 2020, 9, 2089–2091. [Google Scholar]
- Pattnaik, P.; Singh, A.K.; Kumar, B.; Mishra, D.; Singh, B.K.; Barman, K.; Pal, A.K. Analysis of heterotic pattern of F1′s in tomato (Solanum lycopersicum L.) for the improvement of yield and quality traits. Int. J. Chem. Stud. 2020, 8, 3160–3165. [Google Scholar] [CrossRef]
- Solieman, T.; El-Gabry, M.; Abido, A. Heterosis, potence ratio and correlation of some important characters in tomato (Solanum lycopersicum L.). Sci. Hortic. 2013, 150, 25–30. [Google Scholar] [CrossRef]
- Liu, J.; Li, M.; Zhang, Q.; Wei, X.; Huang, X. Exploring the molecular basis of heterosis for plant breeding. J. Integr. Plant Biol. 2020, 62, 287–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendrick, W.P.; Booth, N.O. Mendelian characters in tomatoes. Proc. Am. Soc. Hort. Sci. 1968, 5, 19–24. [Google Scholar]
- Bhatt, R.P.; Biswas, V.R.; Kumar, N. Heterosis, combining ability, genetics for vitamin C, total soluble solids and yield in tomato (Lycopersicon esculentum Mill.) at 1700 m altitude. J. Agric. Sci. 2001, 137, 71–75. [Google Scholar] [CrossRef]
- Bai, Y.; Lindhout, P. Domestication and breeding of tomatoes: What have we gained and what can we gain in the future? Ann. Bot. 2007, 100, 1085–1094. [Google Scholar] [CrossRef]
- Fasoulas, A.C. Principles of Crop Breeding; Aristotle University Press: Thessaloniki, Greece, 1993. [Google Scholar]
- Fasoula, D.A.; Fasoula, V.A. Bridging the productivity gap between maize inbreds and hybrids by replacing gene and genome dichotomization with gene and genome integration. Maydica 2005, 50, 49–61. [Google Scholar]
- Souza, L.M.; Paterniani, M.; Melo, P.C.T.; Melo, A.M.T. Diallel cross among fresh market tomato inbreeding lines. Hortic. Bras. 2012, 30, 246–251. [Google Scholar] [CrossRef] [Green Version]
- Cappetta, E.; Andolfo, G.; Di Matteo, A.; Ercolano, M.R. Empowering crop resilience to environmental multiple stress through the modulation of key response components. J. Plant Physiol. 2020, 246, 153134. [Google Scholar] [CrossRef]
- Cappetta, E.; Andolfo, G.; Di Matteo, A.; Barone, A.; Frusciante, L.; Ercolano, M.R. Accelerating Tomato Breeding by Exploiting Genomic Selection Approaches. Plants 2020, 9, 1236. [Google Scholar] [CrossRef]
- Koutsika-Sotiriou, M.S.; Traka-Mavrona, E.A.; Evgenidis, G.L. Assessment of tomato source breeding material through mating designs. J. Agric. Sci. 2008, 146, 301–310. [Google Scholar] [CrossRef]
- Avdikos, I.D.; Tsivelika, N.; Gallidou, A.; Koutsika-Sotiriou, M.; Traka-Mavrona, E. Exploitation of heterosis through recurrent selection scheme applied in segregating generations of a tomato breeding program. Sci. Hortic. 2011, 130, 701–707. [Google Scholar] [CrossRef]
- Fasoula, V.A.; Fasoula, D.A. Principles underlying genetic improvement for high and stable crop yield potential. Field Crop. Res. 2002, 75, 191–209. [Google Scholar] [CrossRef]
- Tollenaar, M.; Wu, J. Yield improvement in temperate maize is attributable to greater stress tolerance. Crop. Sci. 1999, 39, 1597–1604. [Google Scholar] [CrossRef]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods, 8th ed.; Iowa State University Press: Ames, IA, USA, 1989; p. 491. [Google Scholar]
- Yan, W. GGEbiplot—A Windows application for graphical analysis of multi-environment trial data and other types of two-way data. Agron. J. 2001, 93, 1111–1118. [Google Scholar] [CrossRef] [Green Version]
- Yan, W. Singular-value partitioning in biplot analysis of multienvironment trial data. Agron. J. 2002, 94, 990–996. [Google Scholar] [CrossRef] [Green Version]
- Patel, U.J.; Kathiria, K.B.; Patel, J.S.; Saiyad, I.M. Heterobeltiosis and inbreeding depression in tomato (Lycopersicon esculentum Mill.). Int. J. Plant Sci. 2010, 5, 636–638. [Google Scholar]
- Aragão, F.; Giordano, L.b.; Melo, P.C.T.; Boiteux, L.S. Desempenho de híbridos experimentais de tomateiro para processamento industrial nas condições edafoclimáticas do cerrado brasileiro. Hortic. Bras. 2004, 22, 529–533. [Google Scholar] [CrossRef] [Green Version]
- Santos, F.F.B.; Ribeiro, A.; Siqueira, W.J.; Melo, A.M.T. Desempenho agronômico de híbridos F1 de tomate de mesa. Hortic. Bras. 2011, 29, 304–310. [Google Scholar] [CrossRef]
- Schwarz, K.; Resende, J.T.V.; Preczenhak, A.P.; Paula, J.T.; Faria, M.V.; Dias, D.M. Desempenho agronômico e qualidade físico-química de híbridos de tomateiro em cultivo rasteiro. Hortic. Bras. 2013, 31, 410–418. [Google Scholar] [CrossRef] [Green Version]
- Zhuchenko, A.A. Genetics of the Tomato; Academic Press: Kishinev, Moldova, 1973; p. 644. [Google Scholar]
- Daskaloff, C.; Konstantinova, M. The inheritance of some quantitative characters determining tomato fruit quality in view of developing high quality lines and cultivars. In Proceedings of the Meeting of the Eucarpia Tomato Working Group, Avignon, France, 18–21 May 1981; pp. 121–128. [Google Scholar]
- Georgiev, H. Heterosis in tomato breeding. In Genetic Improvement of Tomato; Kalloo, G., Ed.; Springer: Berlin, Germany, 1991; pp. 83–98. [Google Scholar]
- Smith, H.H. Fixing transgressive vigor in Nicotiana Rustica. In Heterosis; Gowen, J.M., Ed.; Iowa State College Press: Ames, IA, USA, 1952; pp. 161–174. [Google Scholar]
- Powers, L. Gene recombination and heterosis. In Heterosis; Gowen, J.M., Ed.; Iowa State College Press: Ames, IA, USA, 1952; pp. 298–316. [Google Scholar]
- Kimbeng, C.A.; Bingham, E.T. Population improvement in lucerne (Medicago sativa L.): Components of inbreeding depression are different in original and improved populations. Aust. J. Exp. Agric. 1998, 38, 831–836. [Google Scholar] [CrossRef]
- Williams, W. Isolation of “pure lines” from F1 Hybrids of tomato and the problem of heterosis in breeding crop species. J. Agric. Sci. 1959, 53, 347–353. [Google Scholar] [CrossRef]
- Cuartero, J.; Baguena, M.; Costa, J.; Diezm, J.; Nuez, F. The use of tomato hybrids or varieties. Acta Hort. 1986, 191, 347–354. [Google Scholar] [CrossRef]
- Christakis, P.A.; Fasoulas, A.C. The recovery of recombinant inbreds outyielding the hybrid in tomato. J. Agric. Sci. 2001, 137, 179–183. [Google Scholar] [CrossRef]
- Christakis, P.A.; Fasoulas, A.C. The effects of the genotype by environmental interaction on the fixation of heterosis in tomato. J. Agric. Sci. 2002, 139, 55–60. [Google Scholar] [CrossRef]
- Ipsilandis, C.G.; Koutsika-Sotiriou, M. The combining ability of recombinant S-lines developed from an F2 maize population. J. Agric. Sci. 2000, 134, 191–198. [Google Scholar] [CrossRef]
- Hallauer, A.R.; Miranda, J.B. Quantitative Genetics in Maize Breeding; Iowa State University Press: Ames, IA, USA, 1981. [Google Scholar]
- Genter, C.F.; Alexander, M.W. Development and selection of productive S.I. inbred lines of com (Zea mays L.). Crop. Sci. 1966, 6, 429–431. [Google Scholar] [CrossRef]
- Ceccarelli, S. Adaptation to low/high input cultivation. Euphytica 1996, 92, 203–214. [Google Scholar] [CrossRef]
- Janick, J. Exploitation of heterosis: Uniformity and stability. In The Genetics and Exploitation of Heterosis in Crops; Coors, J.G., Pandey, S., Eds.; ASA-CSSA-SSSA: Madison, WI, USA, 1999; pp. 319–333. [Google Scholar]
- Murphy, K.M.; Campbell, K.G.; Lyon, S.R.; Jones, S.S. Evidence of varietal adaptation to organic farming systems. Field Crop. Res. 2007, 102, 172–177. [Google Scholar] [CrossRef] [Green Version]
- Reid, T.A.; Yang, R.; Salmon, D.F.; Navabi, A.; Spaner, D. Realized gains from selection for spring wheat grain yield are different in conventional and organically managed systems. Euphytica 2010, 177, 253–266. [Google Scholar] [CrossRef]
- Shelton, A.C.; Tracy, W.F. Participatory plant breeding and organic agriculture: A synergistic model for organic variety development in the United States. Elem. Sci. Anthr. 2016, 4, 000143. [Google Scholar] [CrossRef] [Green Version]
- Hubbard, K.; Zystro, J. State of Organic Seed; Organic Seed Alliance: Port Townsend, WA, USA, 2016. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Entry | Early Fruit Yield in High Inputs | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| g/Plant | Fruit Number/Plant | g/Fruit | |||||||
| V/D | /s | V/D | /s | V/D | /s | ||||
| Formula F1 | 2540.13 a * | 100 | 3.01 | 11.40 a | 100 | 3.35 | 220.67 a | 100 | 6.37 |
| Formula F5-1 | 1111.60 c | 44 | 2.78 | 7.20 b | 63 | 3.53 | 156.67 b | 71 | 3.24 |
| Formula F5-2 | 1025.60 c | 40 | 2.29 | 6.80 b | 60 | 2.16 | 163.20 b | 74 | 2.67 |
| Formula F5-3 | 512.70 c | 20 | 2.14 | 3.57 b | 31 | 2.59 | 145.03 b | 66 | 2.64 |
| Formula F5-4 | 1087.87 c | 43 | 1.70 | 5.33 b | 47 | 1.98 | 196.20 a | 89 | 6.44 |
| Formula F5-5 | 861.07 c | 34 | 1.72 | 4.80 b | 42 | 1.96 | 177.20 b | 80 | 3.60 |
| Formula F5-6 | 1207.43 c | 48 | 2.56 | 7.93 b | 70 | 3.58 | 150.60 b | 68 | 4.89 |
| Formula F5-7 | 858.53 c | 34 | 1.74 | 5.20 b | 46 | 2.40 | 167.37 b | 76 | 2.94 |
| Formula F5-8 | 1956.6 b | 77 | 3.36 | 10.50 a | 92 | 2.47 | 198.90 a | 90 | 4.52 |
| Elpida F1 | 2018.37 b | 100 | 2.69 | 9.33 a | 100 | 3.28 | 212.60 a | 100 | 6.86 |
| Elpida F5-1 | 1600.13 b | 79 | 3.51 | 10.13 a | 109 | 3.56 | 163.80 b | 77 | 3.58 |
| Elpida F5-2 | 1644.73 b | 81 | 2.47 | 10.93 a | 117 | 2.92 | 152.03 b | 72 | 3.78 |
| Elpida F5-3 | 2194.60 a | 109 | 2.39 | 11.33 a | 121 | 2.34 | 194.47 a | 91 | 5.27 |
| Elpida F5-4 | 1479.27 b | 73 | 2.81 | 8.93 a | 96 | 2.96 | 172.70 b | 81 | 3.40 |
| Elpida F5-5 | 886.10 c | 44 | 2.45 | 7.77 b | 83 | 2.21 | 127.33 b | 60 | 2.69 |
| Elpida F5-6 | 1725.27 b | 85 | 2.65 | 12.00 a | 129 | 2.58 | 149.37 b | 70 | 4.67 |
| Elpida F5-7 | 2425.40 a | 120 | 2.73 | 12.13 a | 130 | 2.88 | 201.93 a | 95 | 5.42 |
| Elpida F5-8 | 1746.60 b | 87 | 1.85 | 7.47 b | 80 | 1.84 | 234.73 a | 110 | 4.70 |
| Iron F1 | 1406.41 c | 100 | 1.77 | 6.13 b | 100 | 1.92 | 216.00 a | 100 | 4.96 |
| Iron HS6-2 | 811.17 c | 62 | 1.72 | 5.13 b | 84 | 1.90 | 152.03 b | 70 | 3.36 |
| Iron HS6-3 | 915.37 c | 70 | 2.22 | 6.27 b | 102 | 2.88 | 143.23 b | 66 | 4.45 |
| Sahara F1 | 928.97 c | 100 | 1.71 | 4.77 b | 100 | 1.62 | 192.23 a | 100 | 3.41 |
| Sahara HS6-1 | 865.77 c | 93 | 1.67 | 7.67 b | 161 | 1.50 | 115.17 b | 60 | 3.30 |
| Sahara HS6-2 | 901.37 c | 97 | 1.69 | 5.77 b | 121 | 1.54 | 165.57 b | 86 | 3.32 |
| Makedonia | 968.80 c | — | 1.56 | 5.60 b | — | 1.97 | 168.27 b | — | 4.00 |
| Entry | Total Yield in High Inputs | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| g/Plant | Fruit Number/Plant | g/Fruit | |||||||
| V/D | /s | V/D | /s | V/D | /s | ||||
| Formula F1 | 5542.20 b * | 100 | 7.40 | 25.17 b | 100 | 6.88 | 221.53 a | 100 | 10.66 |
| Formula F5-1 | 4076.07 c | 74 | 4.37 | 26.47 b | 105 | 6.02 | 155.83 c | 70 | 4.41 |
| Formula F5-2 | 4075.20 c | 74 | 5.20 | 27.13 b | 108 | 4.99 | 152.47 c | 69 | 5.61 |
| Formula F5-3 | 2953.97 d | 53 | 2.77 | 18.43 c | 73 | 2.99 | 160.13 c | 72 | 4.63 |
| Formula F5-4 | 4580.67 c | 83 | 7.12 | 23.33 b | 93 | 5.77 | 198.50 a | 90 | 10.01 |
| Formula F5-5 | 3860.20 c | 70 | 4.70 | 24.47 b | 97 | 5.04 | 158.27 c | 71 | 11.48 |
| Formula F5-6 | 4572.73 c | 83 | 7.06 | 30.03 b | 119 | 5.19 | 155.10 c | 70 | 8.17 |
| Formula F5-7 | 3978.80 c | 72 | 5.33 | 23.03 b | 91 | 5.78 | 173.37 b | 78 | 9.67 |
| Formula F5-8 | 4052.77 c | 73 | 5.61 | 22.43 b | 89 | 3.86 | 185.87 b | 84 | 6.63 |
| Elpida F1 | 6201.20 a | 100 | 9.88 | 32.50 a | 100 | 7.64 | 193.17 a | 100 | 9.71 |
| Elpida F5-1 | 5653.87 b | 91 | 5.90 | 38.47 a | 118 | 6.61 | 147.97 c | 77 | 7.51 |
| Elpida F5-2 | 5127.70 b | 83 | 5.42 | 36.13 a | 111 | 4.76 | 145.27 c | 75 | 6.21 |
| Elpida F5-3 | 6686.87 a | 108 | 7.34 | 37.47 a | 115 | 5.66 | 181.87 b | 94 | 6.26 |
| Elpida F5-4 | 6139.07 a | 99 | 5.63 | 37.60 a | 116 | 7.89 | 163.07 c | 84 | 8.39 |
| Elpida F5-5 | 5242.87 b | 85 | 4.43 | 38.73 a | 119 | 4.44 | 136.47 c | 71 | 8.08 |
| Elpida F5-6 | 5098.20 b | 82 | 4.98 | 34.20 a | 105 | 8.11 | 149.23 c | 77 | 5.96 |
| Elpida F5-7 | 6275.73 a | 101 | 6.65 | 34.20 a | 105 | 8.61 | 183.53 b | 95 | 10.64 |
| Elpida F5-8 | 6685.53 a | 108 | 6.69 | 34.60 a | 106 | 6.80 | 195.37 a | 101 | 6.74 |
| Iron F1 | 5013.07 b | 100 | 5.69 | 25.33 b | 100 | 4.70 | 197.77 a | 100 | 7.61 |
| Iron HS6-2 | 3859.57 c | 77 | 4.38 | 24.13 b | 95 | 4.68 | 160.13 c | 81 | 9.02 |
| Iron HS6-3 | 3678.63 c | 73 | 6.16 | 24.70 b | 98 | 6.52 | 150.17 c | 76 | 6.55 |
| Sahara F1 | 4579.63 c | 100 | 3.52 | 25.67 b | 100 | 4.54 | 178.00 b | 100 | 4.02 |
| Sahara HS6-1 | 3859.63 c | 84 | 3.46 | 28.80 b | 112 | 5.79 | 133.03 c | 75 | 4.53 |
| Sahara HS6-2 | 4174.47 c | 91 | 2.90 | 28.00 b | 109 | 2.61 | 156.33 c | 88 | 3.52 |
| Makedonia | 2683.27 d | — | 2.73 | 17.70 c | — | 4.01 | 147.83 c | — | 5.88 |
| Entry | Early Fruit Yield in Low Inputs | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| g/Plant | FRUIT Number/Plant | g/Fruit | |||||||
| V/D | /s | V/D | /s | V/D | /s | ||||
| Formula F1 | 1393.76 a * | 100 | 3.41 | 8.24 a | 100 | 3.41 | 169.70 a | 100 | 5.52 |
| Formula F5-1 | 806.63 b | 58 | 2.12 | 5.38 b | 65 | 2.84 | 146.63 b | 86 | 4.07 |
| Formula F5-2 | 703.07 b | 50 | 2.25 | 5.07 b | 62 | 2.13 | 143.54 b | 85 | 4.01 |
| Formula F5-3 | 875.00 b | 63 | 3.44 | 5.20 b | 63 | 4.11 | 169.46 a | 100 | 5.34 |
| Formula F5-4 | 1087.81 a | 78 | 1.95 | 6.81 b | 83 | 1.99 | 156.19 a | 92 | 4.81 |
| Formula F5-5 | 766.64 b | 55 | 1.17 | 5.07 b | 62 | 1.19 | 152.60 b | 90 | 5.11 |
| Formula F5-6 | 1254.31 a | 90 | 2.39 | 9.19 a | 112 | 2.98 | 135.44 b | 80 | 5.23 |
| Formula F5-7 | 686.50 b | 49 | 1.23 | 5.14 b | 62 | 1.47 | 122.59 b | 72 | 3.14 |
| Formula F5-8 | 1377.56 a | 99 | 2.88 | 9.69 a | 118 | 2.95 | 142.63 b | 84 | 5.71 |
| Elpida F1 | 1079.18 a | 100 | 3.64 | 7.88 a | 100 | 3.72 | 137.73 b | 100 | 6.35 |
| Elpida F5-1 | 897.47 b | 83 | 2.62 | 5.82 b | 74 | 2.53 | 157.80 a | 115 | 5.68 |
| Elpida F5-2 | 992.29 b | 92 | 2.35 | 6.82 b | 87 | 2.29 | 147.13 b | 107 | 5.81 |
| Elpida F5-3 | 1043.31 a | 97 | 2.56 | 6.94 b | 88 | 2.73 | 154.17 a | 112 | 4.26 |
| Elpida F5-4 | 1114.65 a | 103 | 2.95 | 6.59 b | 84 | 2.84 | 177.64 a | 129 | 3.58 |
| Elpida F5-5 | 949.00 b | 88 | 1.85 | 5.75 b | 73 | 2.13 | 165.66 a | 120 | 3.30 |
| Elpida F5-6 | 1308.94 a | 121 | 2.63 | 7.63 a | 97 | 2.59 | 173.96 a | 126 | 8.04 |
| Elpida F5-7 | 901.47 b | 84 | 1.77 | 6.60 b | 84 | 2.09 | 138.57 b | 101 | 2.94 |
| Elpida F5-8 | 1023.58 a | 95 | 1.83 | 6.08 b | 77 | 1.74 | 174.97 a | 127 | 3.80 |
| Iron F1 | 771.33 b | 100 | 1.68 | 5.33 b | 100 | 1.70 | 149.90 b | 100 | 3.97 |
| Iron HS6-2 | 879.39 b | 114 | 3.34 | 6.61 b | 124 | 4.95 | 133.47 b | 89 | 4.48 |
| Iron HS6-3 | 967.53 b | 125 | 2.72 | 6.76 b | 127 | 3.86 | 141.54 b | 94 | 4.55 |
| Sahara F1 | 1511.70 a | 100 | 2.76 | 9.22 a | 100 | 2.62 | 164.12 a | 100 | 6.54 |
| Sahara HS6-1 | 735.81 b | 49 | 1.82 | 6.25 b | 68 | 1.62 | 125.34 b | 76 | 3.78 |
| Sahara HS6-2 | 1368.43 a | 91 | 3.23 | 9.93 a | 107 | 2.79 | 140.72 b | 86 | 7.03 |
| Makedonia | 915.61 b | — | 2.11 | 5.72 b | — | 2.36 | 160.14 b | — | 4.04 |
| Entry | Total Yield in Low Inputs | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| g/Plant | Fruit Number/Plant | g/Fruit | |||||||
| V/D | /s | V/D | /s | V/D | /s | ||||
| Formula F1 | 3432.35 a * | 100 | 3.58 | 18.00 b | 100 | 4.12 | 190.02 a | 100 | 5.59 |
| Formula F5-1 | 3394.56 a | 99 | 4.58 | 20.00 b | 111 | 4.68 | 171.95 a | 90 | 5.39 |
| Formula F5-2 | 3395.53 a | 99 | 4.39 | 20.07 b | 111 | 5.46 | 171.52 a | 90 | 4.68 |
| Formula F5-3 | 3240.13 a | 94 | 5.18 | 17.93 b | 100 | 4.20 | 185.11 a | 97 | 5.41 |
| Formula F5-4 | 3207.50 a | 93 | 3.96 | 19.50 b | 108 | 3.65 | 168.78 a | 89 | 5.03 |
| Formula F5-5 | 2851.93 b | 83 | 2.24 | 17.07 b | 95 | 2.64 | 164.03 a | 86 | 4.71 |
| Formula F5-6 | 3311.13 a | 96 | 3.26 | 21.56 a | 120 | 4.48 | 152.63 a | 80 | 5.44 |
| Formula F5-7 | 2689.64 b | 78 | 2.33 | 17.93 b | 100 | 2.98 | 147.58 a | 78 | 4.39 |
| Formula F5-8 | 3538.75 a | 103 | 5.87 | 21.81 a | 121 | 4.38 | 167.15 a | 88 | 4.99 |
| Elpida F1 | 3717.76 a | 100 | 5.14 | 23.94 a | 100 | 5.19 | 156.95 a | 100 | 6.59 |
| Elpida F5-1 | 3821.18 a | 103 | 4.58 | 24.76 a | 103 | 5.35 | 155.65 a | 99 | 6.33 |
| Elpida F5-2 | 3483.47 a | 94 | 4.94 | 22.18 a | 93 | 4.07 | 159.84 a | 102 | 7.51 |
| Elpida F5-3 | 3911.50 a | 105 | 4.88 | 24.44 a | 102 | 4.50 | 162.95 a | 104 | 5.58 |
| Elpida F5-4 | 3832.06 a | 103 | 6.55 | 21.53 a | 90 | 4.81 | 182.00 a | 116 | 5.73 |
| Elpida F5-5 | 2622.13 b | 71 | 3.02 | 16.50 b | 69 | 3.62 | 159.83 a | 102 | 4.14 |
| Elpida F5-6 | 3114.56 a | 84 | 5.04 | 18.94 b | 79 | 5.28 | 166.89 a | 106 | 5.70 |
| Elpida F5-7 | 3398.07 a | 91 | 4.20 | 21.00 a | 88 | 4.52 | 164.61 a | 105 | 4.94 |
| Elpida F5-8 | 3332.67 a | 90 | 3.27 | 18.67 b | 78 | 6.73 | 176.66 a | 113 | 4.72 |
| Iron F1 | 2891.00 b | 100 | 3.19 | 17.61 b | 100 | 3.64 | 164.04 a | 100 | 5.04 |
| Iron HS6-2 | 2834.06 b | 98 | 3.53 | 19.39 b | 110 | 4.54 | 146.87 a | 90 | 5.19 |
| Iron HS6-3 | 2665.71 b | 92 | 3.26 | 17.12 b | 97 | 5.67 | 154.50 a | 94 | 4.83 |
| Sahara F1 | 3509.92 a | 100 | 4.67 | 22.12 a | 100 | 5.12 | 159.85 a | 100 | 5.65 |
| Sahara HS6-1 | 2371.31 b | 68 | 5.41 | 18.44 b | 83 | 5.10 | 129.80 a | 81 | 7.40 |
| Sahara HS6-2 | 3673.50 a | 104 | 4.04 | 24.21 a | 109 | 5.61 | 152.46 a | 95 | 5.56 |
| Makedonia | 2484.44 b | — | 3.32 | 16.33 b | — | 3.66 | 153.56 a | — | 4.78 |
| Entry | Fruit Quality Traits | |||
|---|---|---|---|---|
| Total Soluble Solids (°Brix) | Total Solids (%) | |||
| Vig/Dep | Vig/Dep | |||
| Formula F1 | 5.20 c * | 100 | 5.68 c | 100 |
| Formula F5-1 | 6.50 a | 125 | 6.22 b | 110 |
| Formula F5-2 | 4.77 d | 92 | 5.84 c | 103 |
| Formula F5-3 | 5.50 c | 106 | 6.17 b | 109 |
| Formula F5-4 | 4.77 d | 92 | 5.37 d | 95 |
| Formula F5-5 | 4.87 d | 94 | 5.58 c | 98 |
| Formula F5-6 | 5.48 c | 105 | 6.45 b | 114 |
| Formula F5-7 | 5.67 c | 109 | 6.66 a | 117 |
| Formula F5-8 | 5.38 c | 103 | 6.34 b | 112 |
| Elpida F1 | 5.92 b | 100 | 6.72 a | 100 |
| Elpida F5-1 | 5.07 d | 86 | 5.83 c | 87 |
| Elpida F5-2 | 4.97 d | 84 | 5.95 c | 89 |
| Elpida F5-3 | 4.42 e | 75 | 5.34 d | 79 |
| Elpida F5-4 | 4.64 d | 78 | 5.84 c | 87 |
| Elpida F5-5 | 5.27 c | 89 | 5.95 c | 89 |
| Elpida F5-6 | 5.40 c | 91 | 5.98 c | 89 |
| Elpida F5-7 | 5.24 c | 89 | 5.94 c | 88 |
| Elpida F5-8 | 5.38 c | 91 | 5.99 c | 89 |
| Iron F1 | 5.63 c | 100 | 6.21 b | 100 |
| Iron HS6-2 | 5.47 c | 97 | 6.05 c | 97 |
| Iron HS6-3 | 5.77 c | 102 | 6.56 a | 106 |
| Sahara F1 | 4.35 e | 100 | 5.89 c | 100 |
| Sahara HS6-1 | 3.90 e | 90 | 4.58 d | 78 |
| Sahara HS6-2 | 4.45 e | 102 | 6.04 c | 103 |
| Makedonia | 6.37 a | — | 6.96 a | — |
| Evaluation | High Inputs | Low Inputs | Sum | Ranking | High Inputs | Low Inputs | Sum |
|---|---|---|---|---|---|---|---|
| 1. Formula F1 | 83 | 88 | 171 | 13. Elpida F5-3 | 100 | 100 | 200 |
| 2. Formula F5-1 | 61 | 87 | 148 | 14. Elpida F5-4 | 92 | 98 | 190 |
| 3. Formula F5-2 | 61 | 87 | 148 | 10. Elpida F1 | 93 | 95 | 188 |
| 4. Formula F5-3 | 44 | 83 | 127 | 18. Elpida F5-8 | 100 | 85 | 185 |
| 5. Formula F5-4 | 69 | 82 | 151 | 11. Elpida F5-1 | 85 | 98 | 182 |
| 6. Formula F5-5 | 58 | 73 | 131 | 17. Elpida F5-7 | 94 | 87 | 181 |
| 7. Formula F5-6 | 68 | 85 | 153 | 1. Formula F1 | 83 | 88 | 171 |
| 8. Formula F5-7 | 60 | 69 | 128 | 12. Elpida F5-2 | 77 | 89 | 166 |
| 9. Formula F5-8 | 61 | 90 | 151 | 22. Sahara F1 | 69 | 90 | 158 |
| 10. Elpida F1 | 93 | 95 | 188 | 16. Elpida F5-6 | 76 | 80 | 156 |
| 11. Elpida F5-1 | 85 | 98 | 182 | 7. Formula F5-6 | 68 | 85 | 153 |
| 12. Elpida F5-2 | 77 | 89 | 166 | 24. Sahara HS6-2 | 58 | 94 | 152 |
| 13. Elpida F5-3 | 100 | 100 | 200 | 9. Formula F5-8 | 61 | 90 | 151 |
| 14. Elpida F5-4 | 92 | 98 | 190 | 5. Formula F5-4 | 69 | 82 | 151 |
| 15. Elpida F5-5 | 78 | 67 | 145 | 19. Iron F1 | 75 | 74 | 149 |
| 16. Elpida F5-6 | 76 | 80 | 156 | 3. Formula F5-2 | 61 | 87 | 148 |
| 17. Elpida F5-7 | 94 | 87 | 181 | 2. Formula F5-1 | 61 | 87 | 148 |
| 18. Elpida F5-8 | 100 | 85 | 185 | 15. Elpida F5-5 | 78 | 67 | 145 |
| 19. Iron F1 | 75 | 74 | 149 | 6. Formula F5-5 | 58 | 73 | 131 |
| 20. Iron HS6-2 | 58 | 72 | 130 | 20. Iron HS6-2 | 58 | 72 | 130 |
| 21. Iron HS6-3 | 55 | 68 | 123 | 8. Formula F5-7 | 60 | 69 | 128 |
| 22. Sahara F1 | 69 | 90 | 158 | 4. Formula F5-3 | 44 | 83 | 127 |
| 23. Sahara HS6-1 | 58 | 61 | 118 | 21. Iron HS6-3 | 55 | 68 | 123 |
| 24. Sahara HS6-2 | 58 | 94 | 152 | 23. Sahara HS6-1 | 58 | 61 | 118 |
| 25. Makedonia | 40 | 64 | 104 | 25. Makedonia | 40 | 64 | 104 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avdikos, I.D.; Tagiakas, R.; Mylonas, I.; Xynias, I.N.; Mavromatis, A.G. Assessment of Tomato Recombinant Lines in Conventional and Organic Farming Systems for Productivity and Fruit Quality Traits. Agronomy 2021, 11, 129. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11010129
Avdikos ID, Tagiakas R, Mylonas I, Xynias IN, Mavromatis AG. Assessment of Tomato Recombinant Lines in Conventional and Organic Farming Systems for Productivity and Fruit Quality Traits. Agronomy. 2021; 11(1):129. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11010129
Chicago/Turabian StyleAvdikos, Ilias D., Rafail Tagiakas, Ioannis Mylonas, Ioannis N. Xynias, and Athanasios G. Mavromatis. 2021. "Assessment of Tomato Recombinant Lines in Conventional and Organic Farming Systems for Productivity and Fruit Quality Traits" Agronomy 11, no. 1: 129. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11010129