
Lignocellulosic Biomass Production and Persistence of Perennial Grass Species Grown in Mediterranean Marginal Lands


Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Experiment Set-Up
2.2. Determinations
2.3. Biomass Quality
2.4. Statistical Analysis
3. Results
3.1. Soil and Meteorological Data
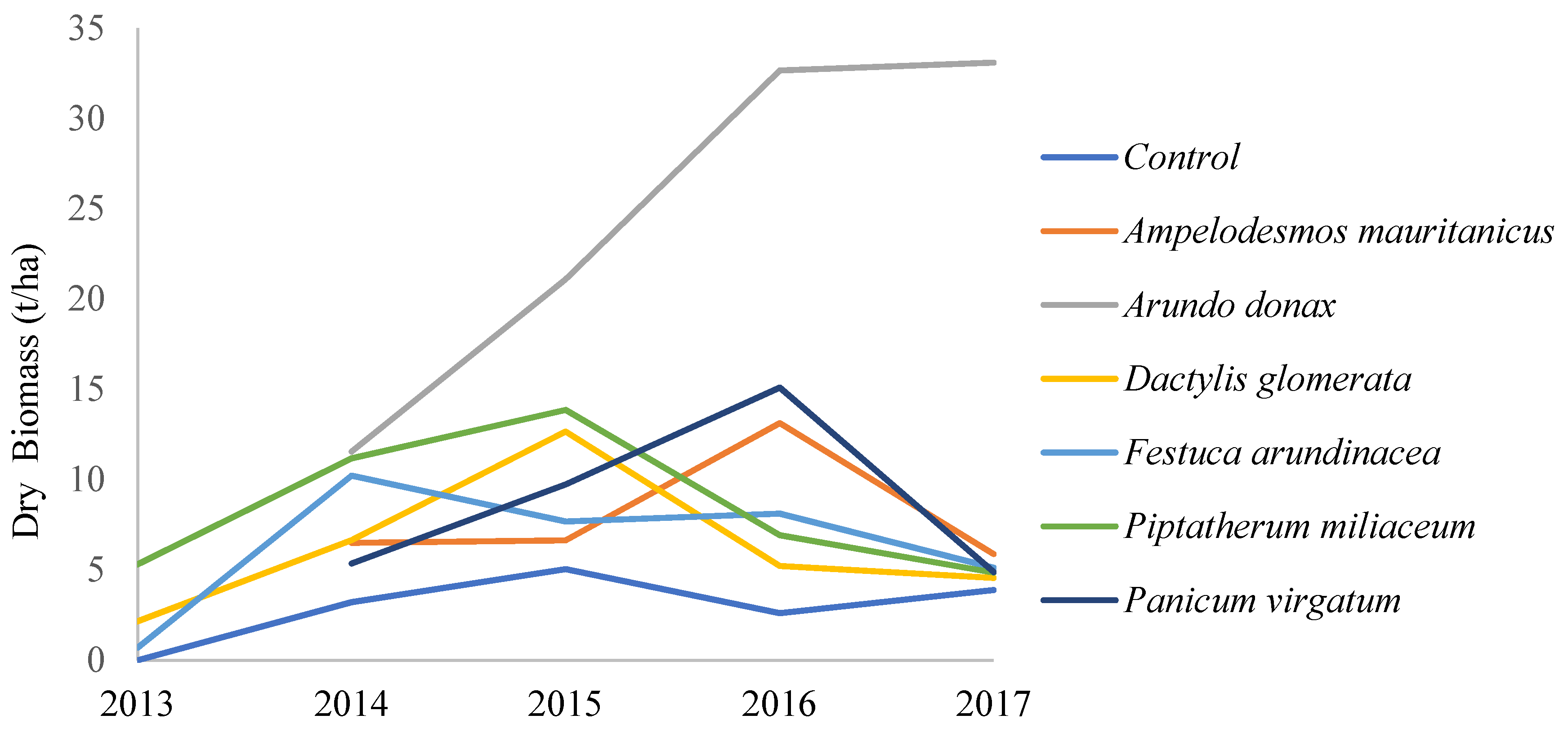
3.2. Biomass Production
3.3. Perennialism
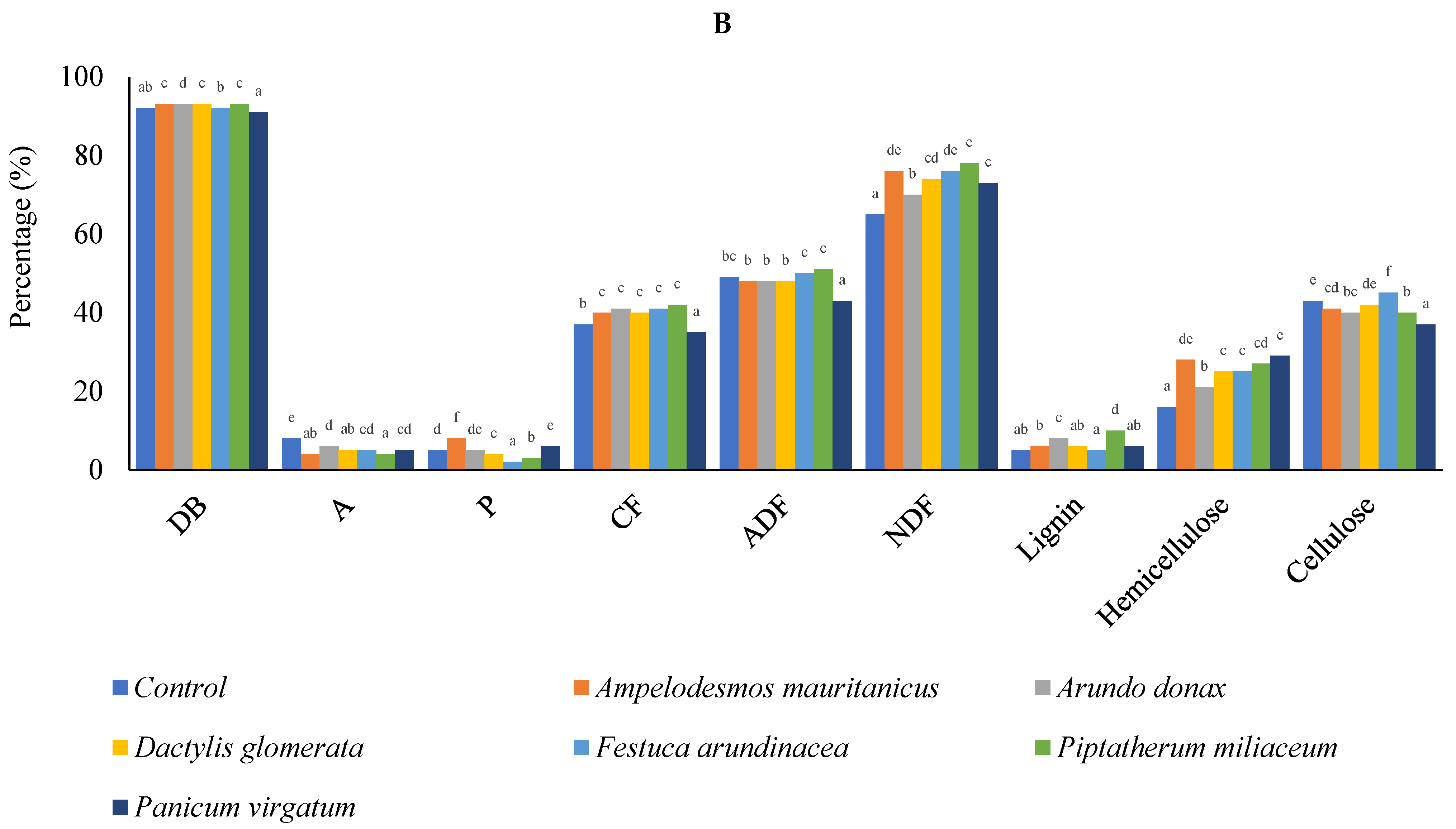
3.4. Biomass Quality
4. Discussion
4.1. Biomass Production and Perennialism
4.2. Biomass Quality and Potential Energy Production
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Peñuelas, J.; Sardans, J.; Walsh, B.J.; Ciais, P.; Janssens, I.A.; Riahi, K.; Rydzak, F.; Obersteiner, M. Posibles escenarios energéticos con vistas al cumplimiento del Acuerdo de París. Ecosistemas 2017, 26, 103–105. [Google Scholar] [CrossRef] [Green Version]
- Edrisi, S.A.; Abhilash, P. Exploring marginal and degraded lands for biomass and bioenergy production: An Indian scenario. Renew. Sustain. Energy Rev. 2016, 54, 1537–1551. [Google Scholar] [CrossRef]
- Renewables 2014. Global Status Report-2014. 2014. Available online: http://wwwren21net/ (accessed on 24 April 2021).
- Saqib, A.; Tabbssum, M.R.; Rashid, U.; Ibrahim, M.; Gill, S.S.; Mehmood, M.A. Marine macro algae Ulva: A Potential feed-stock for bio-ethanol and biogas production. Asian J. Agric. Biol. 2013, 1, 155–163. [Google Scholar]
- Sanderson, M.A.; Adler, P.R. Perennial forages as second generation bioenergy crops. Int. J. Mol. Sci. 2008, 9, 768–788. [Google Scholar] [CrossRef] [Green Version]
- Simmons, B.A.; Loque, D.; Blanch, H.W. Next generation biomass feedstocks for biofuel production. Genome Biol. 2008, 9, 242.1–242.6. [Google Scholar] [CrossRef] [PubMed]
- Naik, S.N.; Goud, V.V.; Rout, P.K.; Dalai, A.K. Production of first and second generation biofuels: A comprehensive review. Renew. Sustain. Energy Rev. 2010, 14, 578–597. [Google Scholar] [CrossRef]
- Smith, S.L.; Thelen, K.D.; MacDonald, S.J. Yield and quality analyses of bioenergy crops grown on a regulatory brownfield. Biomass Bioenergy 2013, 49, 123–130. [Google Scholar] [CrossRef]
- Kang, S.; Post, W.M.; Nichols, J.A.; Wang, D.; West, T.O.; Bandaru, V.; Izaurralde, R.C. Marginal Lands: Concept, Assessment and Management. J. Agric. Sci. 2013, 5, 129–139. [Google Scholar] [CrossRef] [Green Version]
- Skevas, T.; Swinton, S.M.; Hayden, N.J. What type of landowner would supply marginal land for energy crops? Biomass Bioenergy 2014, 67, 252–259. [Google Scholar] [CrossRef]
- Tilman, D.; Hill, J.; Lehman, C. Carbon-negative biofuels from low-input highdiversity grassland biomass. Science 2006, 314, 1598–1600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelfand, I.; Sahajpal, R.; Zhang, X.; Izaurralde, R.C.; Gross, K.L.; Robertson, G.P. Sustainable bioenergy production from marginal lands in the US Midwest. Nature 2013, 493, 514–517. [Google Scholar] [CrossRef] [PubMed]
- Milbrandt, A.R.; Heimiller, D.M.; Perry, A.D.; Field, C.B. Renewable energy potential on marginal lands in the United States. Renew. Sustain. Energy Rev. 2014, 29, 473–481. [Google Scholar] [CrossRef]
- Mehmood, M.A.; Ibrahim, M.; Rashid, U.; Nawaz, M.; Ali, S.; Hussain, A.; Gull, M. Biomass production for bioenergy using marginal lands. Sustain. Prod. Consum. 2017, 9, 3–21. [Google Scholar] [CrossRef]
- Meehan, T.D.; Hurlbert, A.H.; Gratton, C. Bird communities in future bioenergy landscapes of the uppermidwest. Proc. Natl Acad. Sci. USA 2010, 107, 18533–18538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, G.P.; Hamilton, S.K.; Del Grosso, S.J.; Parton, W.J. The biogeochemistry of bioenergy landscapes: Carbon, nitrogen, and water considerations. Ecol. Appl. 2011, 21, 1055–1067. [Google Scholar] [CrossRef]
- Werling, B.P.; Dickson, T.L.; Isaacs, R.; Gaines, H.; Gratton, C.; Gross, K.L.; Liere, H.; Malmstrom, C.M.; Meehan, T.D.; Ruan, L.; et al. Perennial grasslands enhance biodiversity and multiple ecosystem services in bioenergy landscapes. Proc. Natl. Acad. Sci. USA 2014, 111, 1652–1657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez, E.; Scordia, D.; Lino, G.; Arias, C.; Cosentino, S.L.; Nogués, S. Salinity and water stress effects on biomass production in different Arundo donax L. clones. Bioenerg. Res. 2015, 8, 1461–1479. [Google Scholar] [CrossRef] [Green Version]
- Elbersen, B.; Van Eupen, M.; Mantel, S.; Alexopoulou, E.; Bai, Z.; Boogard, H.; Zanetti, F. Mapping Marginal land potentially available for industrial crops in Europe. In Proceedings of the EUBCE 2018—26th European Biomass Conference & Exhibition, Copenhagen, Denmark, 14–17 May 2018. [Google Scholar]
- Nunn, C.; Hastings, A.F.S.J.; Kalinina, O.; Özgüven, M.; Schüle, H.; Tarakanov, I.G.; Van Der Weijde, T.; Anisimov, A.A.; Iqbal, Y.; Kiesel, A.; et al. Environmental influences on the growing season duration and ripening of diverse miscanthus germplasm grown in six countries. Front. Plant Sci. 2017, 8, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Scordia, D.; Testa, G.; Copani, V.; Patane, C.; Cosentino, S.L. Lignocellulosic biomass production of Mediterranean wild accessions (Oryzopsis miliacea, Cymbopogon hirtus, Sorghum halepense and Saccharum spontaneum) in a semi-arid environment. Field Crops Res. 2017, 214, 56–65. [Google Scholar] [CrossRef]
- Monti, A.; Di Virgilio, N.; Venturi, G. Mineral composition and ash content of six major energy crops. Biomass Bioenergy 2008, 32, 216–223. [Google Scholar] [CrossRef] [Green Version]
- Monti, A.; Zanetti, F.; Scordia, D.; Testa, G.; Cosentino, S.L. What to harvest when? Autumn, winter, annual and biennial harvesting of giant reed, miscanthus and switchgrass in northern and southern Mediterranean area. Ind. Crops Prod. 2015, 75, 129–134. [Google Scholar] [CrossRef]
- MAPA. Métodos Oficiales de Análisis de Suelos y Aguas para Riegos; Tomo III; Servicio de Publicaciones del Ministerio de Agricultura, Pesca y Alimentación: Madrid, Spain, 1994. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Method for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Scordia, D.; van den Berg, D.; van Sleen, P.; Alexopoulou, E.; Cosentino, S.L. Are herbaceous perennial grasses suitable feedstock for thermochemical conversion pathways? Ind. Crops Prod. 2016, 91, 350–357. [Google Scholar] [CrossRef]
- Scordia, D.; Zanetti, F.; Varga, S.S.; Alexopoulou, E.; Cavallaro, V.; Monti, A.; Copani, V.; Cosentino, S.L. New insights into the propagation methods of switchgrass, miscanthus and giant reed. Bioenerg. Res. 2015, 8, 1480–1491. [Google Scholar] [CrossRef]
- Webster, R.J.; Driever, S.; Kromdijk, J.; McGrath, J.; Leakey, A.; Siebke, K.; Demetriades-Shah, T.; Bonnage, S.; Peloe, T.; Lawson, T.; et al. High C3 photosynthetic capacity and high intrinsic water use efficiency underlies the high productivity of the bioenergy grass Arundo donax. Sci. Rep. 2016, 6, 20694. [Google Scholar] [CrossRef]
- Cosentino, S.L.; Copani, V.; D’Agosta, G.M.; Sanzone, E.; Mantineo, M. First results on evaluation of Arundo donax L. clones collected in Southern Italy. Ind. Crops Prod. 2006, 23, 212–222. [Google Scholar] [CrossRef]
- Angelini, L.G.; Ceccarini, L.; Nassi o Di Nasso, N.; Bonari, E. Comparison of Arundo donax L. and Miscanthus x giganteus in a long-term field experiment in Central Italy: Analysis of productive characteristics and energy balance. Biomass Bioenergry 2009, 33, 635–643. [Google Scholar] [CrossRef]
- Alexopoulou, E.; Zanetti, F.; Scordia, D.; Zegada-Lizarazu, W.; Chriustou, M.; Testa, G.; Cosentino, S.L.; Monti, A. Long-term yields of switchgrass, giant reed, and miscanthus in the Mediterranean basin. Bioenergy Res. 2015, 8, 1492–1499. [Google Scholar] [CrossRef]
- Still, C.J.; Berry, J.A.; Collatz, J.; DeFries, R.S. Global distribution of C3 and C4 vegetation: Carbon cycle implications. Glob. Biogeochem. Cycles 2003, 17, 1–14. [Google Scholar] [CrossRef]
- Zhu, X.G.; Chang, T.G.; Song, Q.F.; Finnan, J.; Barth, S.; Martensson, L.M.; Jones, M.B. A system approach guiding future biomass crop development on marginal land. In Perennial Biomass Crops for a Resource-Constrained World; Barth, S., Murphy-Bokern, D., Kalinina, O., Taylor, G., Jones, M., Eds.; Springer International: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Scarlat, N.; Dallemand, J.F.; Monforti-Ferrario, F.; Nita, V. The role of biomass and bioenergy in a future bioeconomy: Policies and facts. J. Environ. Dev. 2015, 15, 3–34. [Google Scholar] [CrossRef]
- Kludze, H.; Deen, B.; Dutta, A. Impact of agronomic treatments on fuel characteristics of herbaceous biomass for combustion. Fuel Process. Technol. 2013, 109, 96–102. [Google Scholar] [CrossRef]
- Zhu, L.; Zhong, Z. Effects of cellulose, hemicellulose and lignin on biomass pyrolysis kinetics. Korean J. Chem. Eng. 2020, 37, 1660–1668. [Google Scholar] [CrossRef]
- Baraza, E.; Tauler, M.; Romero-Munar, A.; Cifre, J.; Gulías, J. Mycorrhiza-based biofertilizer application to improve the quality of Arundo donax L. plantlets. In Perennial Biomass Crops for a Resource-Constrained World; Barth, S., Murphy-Bokern, D., Kalinina, O., Taylor, G., Jones, M., Eds.; Springer International: Berlin/Heidelberg, Germany, 2016. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Property | |
|---|---|
| Sand(g/kg) | 261 |
| Silt (g/kg) | 465 |
| Clay (g/kg) | 274 |
| Texture | Loam clayey |
| Organic Carbon(g/kg) | 26.1 |
| N (total)(g/kg) | 2.24 |
| P (total) (mg/kg) | 1010 |
| P (organic) (mg/kg) | 332 |
| P (Olsen) (mg/kg) | 52.5 |
| CCE (g/kg) | 310 |
| Active Lime (g/kg) | 60 |
| Electrical Conductivity (1:5; 25 °C) | 168 |
| pH H2O | 8.3 |
| Cation Exchange Capacity (cmol/kg) | 15.9 |
| Hm −0.3 bar (EG, g/kg) | 125 |
| Hm −0.3 bar (g/kg) | 194 |
| FB 1 (t/ha) | DB 2 (t/ha) | |
|---|---|---|
| Control | 6.60 ± 1.457 a,3 | 2.95 ± 0.717 a |
| Ampelodesmos mauritanicus | 16.34 ± 1.629 bc | 8.03 ± 0.802 bc |
| Arundo donax | 45.11 ± 1.629 d | 24.59 ± 0.802 d |
| Dactylis glomerata | 13.47 ± 0.841 b | 6.25 ± 0.414 b |
| Festuca arundinacea | 16.64 ± 1.457 bc | 6.37 ± 0.717 b |
| Piptatherum miliaceum | 18.11 ± 1.030 c | 8.42 ± 0.507 c |
| Panicum virgatum | 16.64 ± 1.629 bc | 8.77 ± 0.802 c |
| 1 | 3.16 ± 1.371 a | 2.04 ± 0.675 a |
| 2 | 18.01 ± 1.124 c | 7.81 ± 0.553 b |
| 3 | 25.84 ± 1.124 d | 10.96 ± 0.553 d |
| 4 | 22.44 ± 1.124 c | 11.96 ± 0.553 c |
| 5 | 15.70 ± 1.124 b | 8.90 ± 0.553 b |
| R2 | 0.852 | 0.839 |
| P-value (Species) | 0.000 | 0.000 |
| P-value (Cycle) | 0.000 | 0.000 |
| P-value (Species × Cycle) | 0.000 | 0.000 |
| Survival (%) | |
|---|---|
| Ampelodesmos mauritanicus | 98.8 ± 0.56 1 |
| Arundo donax | 100.0 ± 0.00 |
| Dactylis glomerata | 85.1 ± 2.32 |
| Festuca arundinacea | 81.1 ± 5.33 |
| Piptatherum miliaceum | 82.7 ± 3.14 |
| Panicum virgatum | 95.0 ± 2.66 |
| 1 | 98.0 ± 0.90 2 |
| 2 | 97.9 ± 0.90 |
| 3 | 94.4 ± 1.49 |
| 4 | 86.2 ± 1.90 |
| 5 | 66.2 ± 4.10 |
| Stems (nº of Stems) | |
|---|---|
| Ampelodesmos mauritanicus | 87.35 ± 2.886 e,1 |
| Arundo donax | 10.65 ± 2.886 a |
| Dactylis glomerata | 38.19 ± 1.700 b |
| Festuca arundinacea | 50.29 ± 2.707 c |
| Piptatherum miliaceum | 37.29 ± 2.220 b |
| Panicum virgatum | 77.88 ± 2.886 d |
| 1 | 22.23 ± 2.605 a |
| 2 | 45.99 ± 2.115 c |
| 3 | 44.21 ± 2.306 b |
| 4 | 45.44 ± 2.115 b |
| 5 | 76.26 ± 2.537 d |
| R2 | 0.760 |
| P (Species) | 0.000 |
| P (Cycle) | 0.000 |
| P (Species × Cycle) | 0.000 |
| Species | DB 1 | A 2 (g/100 g DB) | CF 3 (g/100 g DB) | ADF 4 (g/100 g DB) | NDF 5 (g/100 g DB) | P 6 (g/100 g DB) | Cellulose (g/100 g DB) | Hemicellulose (g/100 g DB) | Lignin (g/100 g DB) |
|---|---|---|---|---|---|---|---|---|---|
| Control | 92.03 ± 0.198 d.7 | 8.76 ± 0.118 f | 37.40 ± 0.563 a | 49.34 ± 0.502 b | 65.49 ± 0.492 a | 5.40 ± 0.202 c | 43.36 ± 0.438 e | 16.14 ± 0.696 a | 5.98 b ± 0.289 |
| Ampelodesmos mauritanicus | 93.42 ± 0.140 f | 5.78 ± 0.083 b | 39.36 ± 0.398 b | 46.56 ± 0.355 a | 74.81 ± 0.348 d | 8.08 ± 0.143 e | 40.48 ± 0.309 bc | 28.25 ± 0.492 de | 6.08 b ± 0.204 |
| Arundo donax | 93.77 ± 0.140 f | 6.21 ± 0.083 c | 43.70 ± 0.398 d | 51.60 ± 0.355 c | 72.90 ± 0.348 c | 4.64 ± 0.143 b | 42.38 ± 0.309 d | 21.31 ± 0.492 b | 9.21 d ± 0.204 |
| Dactylis glomerata | 90.92 ± 0.081 c | 6.96 ± 0.048 d | 37.67 ± 0.230 a | 45.70 ± 0.205 a | 69.33 ± 0.201 b | 6.10 ± 0.083 d | 39.70 ± 0.179 b | 23.63 ± 0.284 c | 6.00 b ± 0.118 |
| Festuca arundinacea | 89.72 ± 0.140 a | 7.36 ± 0.083 e | 37.43 ± 0.398 a | 45.80 ± 0.355 a | 70.23 ± 0.348 b | 3.63 ± 0.143 a | 41.33 ± 0.309 c | 24.43 ± 0.492 c | 4.48 a ± 0.204 |
| Piptatherum miliaceum | 90.27 ± 0.099 b | 5.62 ± 0.059 b | 38.87 ± 0.281 b | 46.60 ± 0.251 a | 74.25 ± 0.246 d | 5.67 ± 0.101 c | 37.96 ± 0.219 a | 27.65 ± 0.348 d | 8.64 d ± 0.144 |
| Panicum virgatum | 92.96 ± 0.140 e | 5.21 ± 0.083 a | 41.26 ± 0.398 c | 49.84 ± 0.355 b | 79.09 ± 0.348 e | 4.02 ± 0.143 a | 42.54 ± 0.309 de | 29.25 ± 0.492 e | 7.30 c ± 0.204 |
| R2 | 0.983 | 0.988 | 0.950 | 0.967 | 0.982 | 0.973 | 0.958 | 0.911 | 0.946 |
| P (Species) | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 |
| P (Cycle) | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.001 | 0.000 |
| P (Species × Cycle) | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sacristán, D.; Cifre, J.; Llompart, M.; Jaume, J.; Gulias, J. Lignocellulosic Biomass Production and Persistence of Perennial Grass Species Grown in Mediterranean Marginal Lands. Agronomy 2021, 11, 2060. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11102060
Sacristán D, Cifre J, Llompart M, Jaume J, Gulias J. Lignocellulosic Biomass Production and Persistence of Perennial Grass Species Grown in Mediterranean Marginal Lands. Agronomy. 2021; 11(10):2060. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11102060
Chicago/Turabian StyleSacristán, Daniel, Josep Cifre, Miquel Llompart, Jaume Jaume, and Javier Gulias. 2021. "Lignocellulosic Biomass Production and Persistence of Perennial Grass Species Grown in Mediterranean Marginal Lands" Agronomy 11, no. 10: 2060. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11102060