
Abiotic Stresses: Alteration of Composition and Grain Quality in Food Legumes


 ,
,  , and
, and
Abstract
:1. Introduction
2. Food Legumes
3. Economic Importance of Food Legumes
4. Grain Composition of Food Legumes
5. Nutritional and Health Benefits
6. Abiotic Stresses
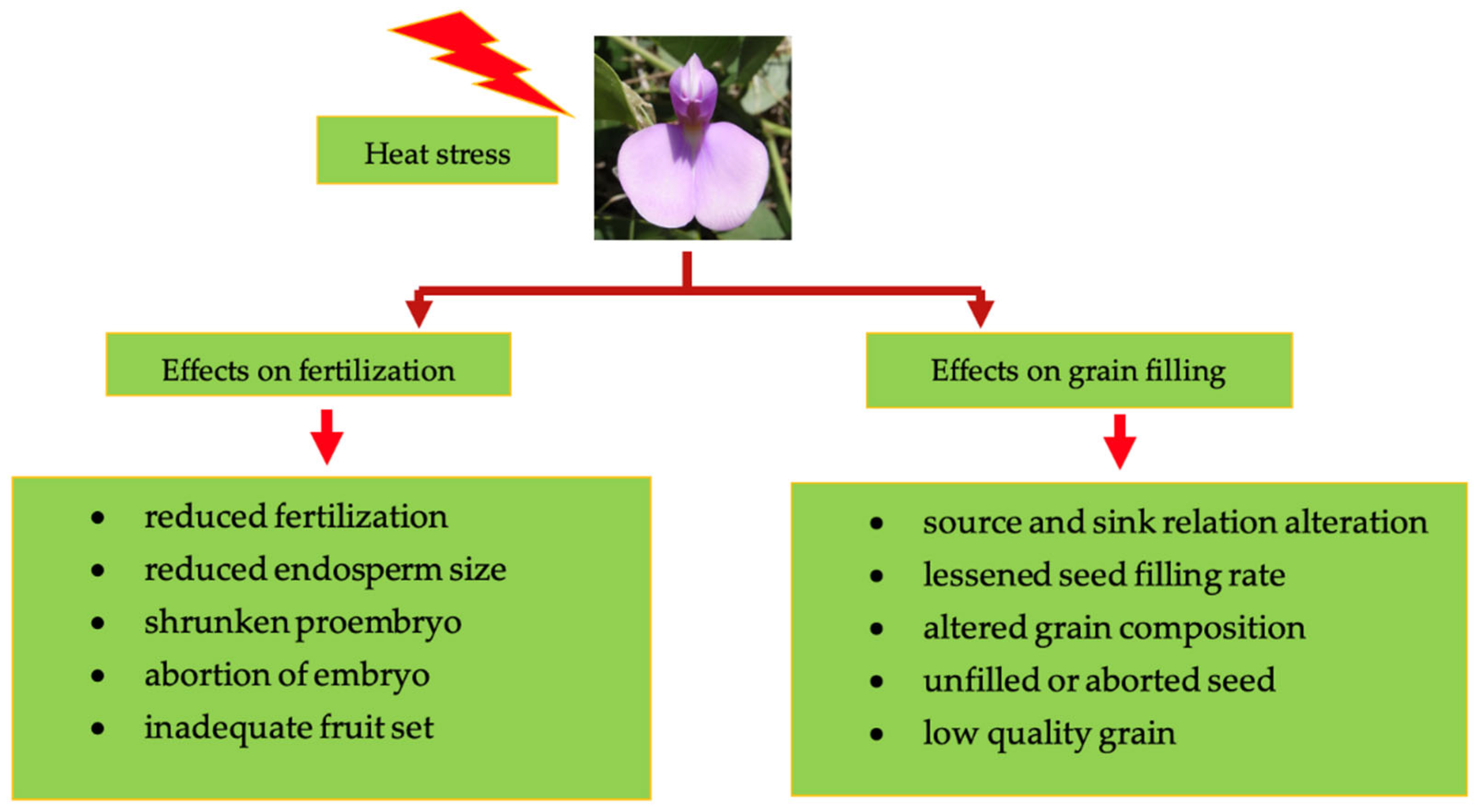
6.1. Temperature Stress
6.1.1. High Temperature
6.1.2. Low Temperature
6.2. Drought
6.3. Salinity
6.4. Heavy Metals
7. Impacts of Abiotic Stresses on Nodulation and Nitrogen Fixation
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Farooq, M.; Hussain, M.; Usman, M.; Farooq, S.; Alghamdi, S.S.; Siddique, K.H.M. Impact of Abiotic Stresses on Grain Composition and Quality in Food Legumes. J. Agric. Food Chem. 2018, 66, 8887–8897. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Romdhane, L.; Rehman, A.; Farooq, M. Adequate zinc nutrition improves the tolerance against drought and heat stresses in chickpea. Plant Physiol. Biochem. 2019, 143, 11–18. [Google Scholar] [CrossRef]
- Gobal Network against Food Crisis. Global Report on Food Crises. Food Security Information Network. 2020. Available online: https://www.sadc.int/files/8415/8818/9448/GRFC_2020_ONLINE.pdf (accessed on 22 May 2021).
- Jimenez-Lopez, J.C.; Singh, K.B.; Clemente, A.; Nelson, M.N.; Ochatt, S.; Smith, P.M.C. Editorial: Legumes for Global Food Security. Front. Plant Sci. 2020, 11, 926. [Google Scholar] [CrossRef]
- Liu, Y.; Li, J.; Zhu, Y.; Jones, A.; Rose, R.J.; Song, Y. Heat Stress in Legume Seed Setting: Effects, Causes, and Future Prospects. Front. Plant Sci. 2019, 10, 938. [Google Scholar] [CrossRef] [Green Version]
- El Sabagh, A.; Hossain, A.; Barutcular, C.; Gormus, O.; Ahmad, Z.; Hussain, S.; Islam, M.S.; Alharby, H.; Bamagoos, A.; Kumar, N.; et al. Effects of drought stress on the quality of major oilseed crops: Implications and possible mitigation strategies—A review. Appl. Ecol. Environ. Res. 2019, 17, 4019–4043. [Google Scholar] [CrossRef]
- Sehrawat, N.; Yadav, M.; Bhat, K.; Sairam, R.; Jaiwal, P. Effect of salinity stress on mungbean [Vigna radiata (L.) Wilczek] during consecutive summer and spring seasons. J. Agric. Sci. 2015, 60, 23–32. [Google Scholar] [CrossRef]
- Garg, N.; Bhandari, P. Silicon nutrition and mycorrhizal inoculations improve growth, nutrient status, K+/Na+ ratio and yield of Cicer arietinum L. genotypes under salinity stress. Plant. Growth Regul. 2016, 78, 371–387. [Google Scholar] [CrossRef]
- Lebrazi, S.; Fikri-Benbrahim, K. Rhizobium-Legume symbioses: Heavy metal effects and principal approaches for bioremediation of contaminated soil. In Legumes Soil Health Sustainable Managent; Springer: Singapore, 2018; pp. 205–233. [Google Scholar] [CrossRef]
- Ali, Q.; Malik, A. Genetic evaluation of legume species under heavy metal and biogas. Biol. Clin. Sci. Res. J. 2021, 2021, 1–6. [Google Scholar] [CrossRef]
- Considine, M.J.; Siddique, K.H.M.; Foyer, C.H. Nature’s pulse power: Legumes, food security and climate change. J. Exp. Bot. 2017, 68, 1815–1818. [Google Scholar] [CrossRef] [PubMed]
- Siddique, K.H.; Johansen, C.; Turner, N.C.; Jeuffroy, M.H.; Hashem, A.; Sakar, D.; Gan, Y.; Alghamdi, S.S. Innovations in agronomy for food legumes. A review. Agron. Sustain. Dev. 2012, 32, 45–64. [Google Scholar] [CrossRef] [Green Version]
- Stagnari, F.; Maggio, A.; Galieni, A.; Pisante, M. Multiple benefits of legumes for agriculture sustainability: An overview. Chem. Biol. Technol. Agric. 2017, 4, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Celmeli, T.; Sari, H.; Canci, H.; Sari, D.; Adak, A.; Eker, T.; Toker, C. The nutritional content of common bean (Phaseolus vulgaris L.) landraces in comparison to modern varieties. Agronomy 2018, 8, 166. [Google Scholar] [CrossRef] [Green Version]
- Jukanti, A.K.; Gaur, P.M.; Gowda, C.L.L.; Chibbar, R.N. Chickpea: Nutritional properties and its benefits. Br. J. Nutr. 2012, 108, S11–S26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouchenak, M.; Lamri-Senhadji, M. Nutritional quality of legumes, and their role in cardiometabolic risk prevention: A review. J. Med. Food. 2013, 16, 185–198. [Google Scholar] [CrossRef]
- Kamboj, R.; Nanda, V. Proximate composition, nutritional profile and health benefits of legumes—A review. Legum Res. 2018, 41, 325–332. [Google Scholar]
- Singh, B.; Singh, J.P.; Kaur, A.; Singh, N. Phenolic composition and antioxidant potential of grain legume seeds: A review. Food Res. Int. 2017, 101, 1–16. [Google Scholar] [CrossRef]
- Cowpea (Black-Eyed Pea) vs Green Bean—In-Depth Nutrition Comparison. Available online: https://foodstruct.com/compareimages/lentil-vs-cowpeas-commonblackeyes-crowder-southern-matureseeds-cooked-boiled-withoutsalt.jpg (accessed on 22 May 2021).
- Telles, A.C.; Kupski, L.; Furlong, E.B. Phenolic compound in beans as protection against mycotoxins. Food Chem. 2017, 214, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Arslan, M. Diversity for vitamin and amino acid content in grass pea (Lathyrus sativus L.). Legume Res. 2017, 40, 803–810. [Google Scholar] [CrossRef] [Green Version]
- Amarowicz, R. Legume Seeds as an Important Component of Human Diet. Foods 2020, 9, 1812. [Google Scholar] [CrossRef]
- Erbersdobler, H.F.; Barth, C.A.; Jahreis, G. Grain Legumes in the Human Nutrition Nutrient Content and Protein Quality of Pulses. Ernahr. Umsch. 2017, 64, M550–M554. [Google Scholar]
- Myers, J.R.; Kmiecik, K. Common bean: Economic importance and relevance to biological science research. In The Common Bean Genome; Springer: Cham, Switzerland, 2017; pp. 1–20. [Google Scholar]
- Eicher, C.K. The Evolution of Agricultural Education and Training: Global Insights of Relevance for Africa; No. 1099-2016-89233; Michigan State University: East Lansing, MI, USA, 2006. [Google Scholar]
- Rani, K.; Sharma, P.; Kumar, S.; Wati, L.; Kumar, R.; Gurjar, D.S.; Kumar, D. Legumes for sustainable soil and crop management. In Sustainable Management of Soil and Environment; Springer: Singapore, 2019; pp. 193–215. [Google Scholar] [CrossRef]
- Maji, S.; Das, A.; Nath, R.; Bandopadhyay, P.; Das, R.; Gupta, S. Cool Season Food Legumes in Rice Fallows: An Indian Perspective. In Agronomic Crops; Hasanuzzaman, M., Ed.; Production Technologies [Internet]; Springer: Singapore, 2019; Volume 1, pp. 561–605. [Google Scholar] [CrossRef]
- Day, L. Proteins from land plants—Potential resources for human nutrition and food security. Trends Food Sci. Technol. 2013, 32, 25–42. [Google Scholar] [CrossRef]
- USDA. National Nutrient Database for Standard Reference Release 28. 733. 2016. Available online: http://www.ars.usda.gov/Services/docs.htm?docid=8964 (accessed on 1 October 2016).
- Ge, F.K. Legumes in Cropping Systems. Angew. Chem. Int. Ed. 1967, 6, 951–952. [Google Scholar]
- Mathobo, V.M.; Silungwe, H.; Ramashia, S.E.; Anyasi, T.A. Effects of heat-moisture treatment on the thermal, functional properties and composition of cereal, legume and tuber starches—A review. J. Food Sci. Technol. 2021, 58, 412–426. [Google Scholar] [CrossRef]
- Aguilera, Y.; Estrella, I.; Benitez, V.; Esteban, R.M.; Martín-Cabrejas, M.A. Bioactive phenolic compounds and functional properties of dehydrated bean flours. Food Res. Int. 2011, 44, 774–780. [Google Scholar] [CrossRef]
- Gan, R.Y.; Deng, Z.Q.; Yan, A.X.; Shah, N.P.; Lui, W.Y.; Chan, C.L.; Corke, H. Pigmented edible bean coats as natural sources of polyphenols with antioxidant and antibacterial effects. LWT-Food Sci. Technol. García-Lafuente 2016, 73, 168–177. [Google Scholar] [CrossRef]
- Chen, P.X.; Tang, Y.; Marcone, M.F.; Pauls, P.K.; Zhang, B.; Liu, R.; Tsao, R. Characterization of free, conjugated and bound phenolics and lipophilic antioxidants in regular-and non-darkening cranberry beans (Phaseolus vulgaris L.). Food Chem. 2015, 185, 298–308. [Google Scholar] [CrossRef]
- Magalhães, S.C.; Taveira, M.; Cabrita, A.R.; Fonseca, A.J.; Valentão, P.; Andrade, P.B. European marketable grain legume seeds: Further insight into phenolic compounds profiles. Food Chem. 2017, 215, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Public Health England. The Eatwell Guide; Department of Health in Association with the Welsh Assembly Government, the Scottish Government and the Food Standards Agency in Northern Ireland L.; Public Health England: London, UK, 2016.
- Humer, E.; Schedle, K. Fermentation of food and feed: A technology for efficient utilization of macro and trace elements in monogastrics. J. Trace Elem. Med. Biol. 2016, 37, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Sirtori, C.R.; Triolo, M.; Bosisio, R.; Bondioli, A.; Calabresi, L.; De Vergori, V.; Gomaraschi, M.; Mombelli, G.; Pazzucconi, F.; Zacherl, C.; et al. Hypo-cholesterolaemic effects oflupin protein and pea protein/fibre combinations in moder-atelycholesterolaemic effects oflupin protein and pea protein/fibre combinations in moderately hypercholesterolaemic individuals. Br. J. Nutr. 2012, 107, 1176–1183. [Google Scholar] [CrossRef]
- Budhathoki, S.; Sawada, N.; Iwasaki, M.; Yamaji, T.; Goto, A.; Kotemori, A.; Ishihara, J.; Takachi, R.; Charvat, H.; Mizoue, T.; et al. Association of animal and plant protein intake with all-cause and cause-specific mortality in a Japanese cohort. JAMA Intern. Med. 2019, 179, 1509–1518. [Google Scholar] [CrossRef]
- Bean Growers Association. Bean Nutrition Overview. The Bean Institute. 2019. Available online: http://beaninstitute.com/bean-nutrition-overview/ (accessed on 22 May 2021).
- Cowpea Facts, Health Benefits and Nutritional Value. 1899. Available online: https://www.healthline.com/nutrition/black-eyed-peas-nutrition (accessed on 22 May 2021).
- Cowpeas_10 Incredible Health Benefits of Cowpeas (Beans)—Finelib. Available online: https://www.finelib.com/about/nigeria-food-produce/about-cowpea-and-nutritional-benefits/204 (accessed on 22 May 2021).
- Cornmeal Vitamins—How Many Vitamins in Cornmeal. Available online: http://www.freenutritionfacts.com/cornmeal/vitamins/ (accessed on 22 May 2021).
- Purslane Nutrition Facts and Health Benefits. Available online: http://www.nutrition-and-you.com/purslane.html (accessed on 22 May 2021).
- Pigeon Pea vs. Peanut—In-Depth Nutrition Comparison. Available online: https://foodstruct.com/compareimages/peanut-vs-pigeonpeasredgram-matureseeds-raw.jpg (accessed on 22 May 2021).
- Green Tomatoes Facts, Health Benefits and Nutritional Value. Available online: https://www.healthbenefitstimes.com/green-tomatoes/ (accessed on 22 May 2021).
- Thomas, J.M.G.; Boote, K.J.; Allen, L.H., Jr.; Gallo-Meagher, M.; Davis, J.M. Elevated temperature and carbon dioxide effects on soybean seed composition and transcript abundance. Crop Sci. 2003, 43, 1548–1557. [Google Scholar] [CrossRef]
- Devirian, T.A.; Volpe, S.L. The physiological effects of dietary boron. Crit. Rev. Food Sci. Nutr. 2003, 43, 219–231. [Google Scholar] [CrossRef]
- Qados, A.M.S.A. Effect of salt stress on plant growth and metabolism of bean plant Vicia faba (L.). J. Saudi Soc. Agri. Sci. 2011, 10, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.X.; Bozzo, G.G.; Freixas-Coutin, J.A.; Marcone, M.F.; Pauls, P.K.; Tang, Y.; Zhang, B.; Liu, R.; Tsao, R. Free and conjugated phenolic compounds and their antioxidant activities in regular and nondarkening cranberry bean (Phaseolus vulgaris L.) seed coats. J. Funct. Foods 2015, 18, 1047–1056. [Google Scholar] [CrossRef]
- Toker, C.; Yadav, S.S. Legumes cultivars for stress environments. In Climate Change and Management of Cool Season Grain Legume Crops; Yadav, S.S., McNeil, D.L., Redden, R., Patil, S.A., Eds.; Springer: Dordrecht, The Netherland, 2010; pp. 351–376. [Google Scholar] [CrossRef]
- Singh, K.B. Chickpea breeding. In The Chickpea; Saxena, E., Ed.; CAB International: Wallingford, CT, USA, 1987; pp. 127–162. [Google Scholar]
- Yadav, S.S.; Redden, R.; McNeil, D.L.; Patil, S.A. Climate Change and Management of Cool Season Grain Legume Crops; Springer Science & Business Media: New York, NY, USA, 2010; pp. 1–460. [Google Scholar]
- Bhandari, K.; Sharma, K.D.; Hanumantha Rao, B.; Siddique, K.H.M.; Gaur, P.; Agrawal, S.K.; Nair, R.M.; Nayyar, H. Temperature sensitivity of food legumes: A physiological insight. Acta Physiol. Plant 2017, 39, 68. [Google Scholar] [CrossRef] [Green Version]
- Sita, K.; Sehgal, A.; HanumanthaRao, B.; Nair, R.M.; Vara Prasad, P.V.; Kumar, S.; Gaur, P.M.; Farooq, M.; Siddique, K.H.; Varshney, R.K.; et al. Food legumes and rising temperatures: Effects, adaptive functional mechanisms specific to reproductive growth stage and strategies to improve heat tolerance. Front. Plant Sci. 2017, 8, 1658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farooq, M.; Nadeem, F.; Gogoi, N.; Ullah, A.; Alghamdi, S.S.; Nayyar, H.; Siddique, K.H. Heat stress in grain legumes during reproductive and grain-filling phases. Crop Pasture Sci. 2017, 68, 985–1005. [Google Scholar] [CrossRef]
- Redden, R.J.; Hatfield, J.L.; Vara Prasad, P.V.; Ebert, A.W.; Yadav, S.S.; O’Leary, G.J. Temperature, climate change, and global food security. Temp. Plant Dev. 2014, 8, 181–202. [Google Scholar]
- Sehgal, A.; Sita, K.; Kumar, J.; Kumar, S.; Singh, S.; Siddique, K.H.M.; Nayyar, H. Effects of drought, heat and their interaction on the growth, yield and photosynthetic function of lentil (Lens culinaris Medikus) genotypes varying in heat and drought sensitivity. Front. Plant Sci. 2017, 8, 1776. [Google Scholar] [CrossRef] [Green Version]
- Sita, K.; Sehgal, A.; Bhandari, K.; Kumar, J.; Kumar, S.; Singh, S.; Siddique, K.H.; Nayyar, H. Impact of heat stress during seed filling on seed quality and seed yield in lentil (Lens culinaris Medikus) genotypes. J. Sci. Food Agric. 2018, 98, 5134–5141. [Google Scholar] [CrossRef]
- Abeysingha, G.L. The Effect of Auxins on Seed Yield Parameters in Wheat, Pea and Canola Grown under Controlled Environment and Western Canadian Field Conditions. Master’s Thesis, University of Alberta, Edmonton, AB, Canada, 2015. [Google Scholar] [CrossRef]
- Djanaguiraman, M.; Vara Prasad, P.V.; Boyle, D.L.; Schapaugh, W.T. Soybean pollen anatomy, viability and pod set under high temperature stress. J. Agron. Sci. 2013, 199, 171–177. [Google Scholar] [CrossRef]
- Gaur, P.M.; Samineni, S.; Krishnamurthy, L.; Varshney, R.K.; Kumar, S.G.; Ghanem, M.E.; Beebe, S.E.; Rao, I.M.; Chaturvedi, S.K.; Basu, P.S.; et al. High temperature tolerance in grain legumes. Legume Perspect. 2014, 7, 23–24. [Google Scholar]
- Bindumadhava, H.; Nair, R.M.; Nayyar, H.; Riley, J.J.; Easdown, W. Mungbean production under a changing climate-insights from growth physiology. Mysore. J. Agric. Sci. 2017, 51, 21–26. [Google Scholar]
- Bishop, J.; Potts, S.G.; Jones, H.E. Susceptibility of faba bean (Vicia faba L.) to heat stress during floral development and anthesis. J. Agron. Crop Sci. 2016, 202, 508–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirsath and Bhosale Agro India Ltd. Modern Agrotechniques for Cultivation of Black Gram/Urdbean (Vigna mungo L.); Shirsath and Bhosale Agro India Ltd.: Ahmednagar, India, 2017. [Google Scholar]
- Vaz Patto, M.C.; Amarowicz, R.; Aryee, A.N.A.; Boye, J.I.; Chung, H.J.; Martín-Cabrejas, M.A.; Domoney, C. Achievements and challenges in improving the; nutritional quality of food legumes. Crit. Rev. Plant Sci. 2015, 34, 105–143. [Google Scholar] [CrossRef]
- Kaushal, N.; Awasthi, R.; Gupta, K.; Gaur, P.; Siddique, K.H.M.; Nayyar, H. Heat stress induced reproductive failures in chickpea (Cicer arietinum) are associated with impaired sucrose metabolism in leaves and anthers. Funct. Plant Biol. 2013, 40, 1334–1349. [Google Scholar] [CrossRef] [PubMed]
- Sharma, L.; Priya, M.; Bindumadhava, H.; Nair, R.M.; Nayyar, H. Influence of high temperature stress on growth, phenology and yield performance of mungbean [Vigna radiata (L.) Wilczek] under managed growth conditions. Sci. Hortic. 2016, 213, 379–391. [Google Scholar] [CrossRef]
- Sadeghipour, O. The influence of water stress on biomass and harvest index in three mung 460 bean (Vigna radiata L. (Wilczek)) cultivars. Asian J. Plant Sci. 2009, 8, 245–249. [Google Scholar] [CrossRef]
- Thomas, J.M.G.; Prasad, P.V.V.; Boote, K.J.; Allen, L.H., Jr. Seed composition, seedling emergence and early seedling vigor of red kidney bean seed produced at elevated temperature and carbon dioxide. J. Agron. Crop Sci. 2009, 195, 148–156. [Google Scholar] [CrossRef]
- Bhandari, K.; Siddique, K.H.; Turner, N.C.; Kaur, J.; Singh, S.; Agrawal, S.K.; Nayyar, H. Heat stress at reproductive stage disrupts leaf carbohydrate metabolism, impairs reproductive function, and severely reduces seed yield in lentil. J. Crop Improv. 2016, 30, 118–151. [Google Scholar] [CrossRef]
- Awasthi, R.; Kaushal, N.; Vadez, V.; Turner, N.C.; Berger, J.; Siddique, K.H.; Nayyar, H. Individual and combined effects of transient drought and heat stress on carbon assimilation and seed filling in chickpea. Funct. Plant Biol. 2014, 41, 1148–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araújo, S.S.; Beebe, S.; Crespi, M.; Delbreil, B.; González, E.M.; Gruber, V.; Vaz Patto, M.C. Abiotic stress responses in legumes: Strategies used to cope with environmental challenges. Crit. Rev. Plant Sci. 2015, 34, 237–280. [Google Scholar] [CrossRef]
- Sehgal, A.; Sita, K.; Siddique, K.H.M.; Kumar, R.; Bhogireddy, S.; Varshney, R.K.; HanumanthaRao, B.; Nair, R.M.; Prasad, P.V.; Nayyar, H. Drought or/and heat-stress effects on seed filling in food crops: Impacts on functional biochemistry, seed yields, and nutritional quality. Front. Plant Sci. 2018, 9, 1705. [Google Scholar] [CrossRef] [Green Version]
- Croser, J.S.; Clarke, H.J.; Siddique, K.H.M.; Khan, T.N. Low temperature stress: Implications for chickpea (Cicer arietinum L.) improvement. Crit. Rev. Plant Sci. 2003, 22, 185–219. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Al-Khaishany, M.Y.; Al-Qutami, M.A.; Al-Whaibi, M.H.; Grover, A.; Ali, H.M.; Al-Wahibi, M.S. Morphological and physiological characterization of different genotypes of faba bean under heat stress. Saudi J. Biol. Sci. 2015, 22, 656–663. [Google Scholar] [CrossRef] [Green Version]
- Heidarvand, L.; Amiri, R.M.; Naghavi, M.R.; Farayedi, Y.; Sadeghzadeh, B.; Alizadeh, K. Physiological and morphological characteristics of chickpea accessions under low temperature stress. Russ. J. Plant Physiol. 2011, 58, 157–163. [Google Scholar] [CrossRef]
- Shunmugam, A.S.K.; Kannan, U.; Jiang, Y.; Daba, K.A.; Gorim, L.Y. Physiology Based Approaches for Breeding of Next-Generation Food Legumes. Plants 2018, 7, 72. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Nayyar, H.; Bhanwara, R.K.; Upadhyaya, H.D. Chilling Stress Effects on Reproductive Biology of Chickpea. J. Sat. Agric. Res. 2010, 8, 1–14. Available online: http://oar.icrisat.org/id/eprint/5374 (accessed on 22 May 2021).
- Maqbool, A.; Shafiq, S.; Lake, L. Radiant frost tolerance in pulse crops—A review. Euphytica 2010, 172, 1–12. [Google Scholar] [CrossRef]
- Peix, A.; Ramırez-Bahena, M.H.; Velazquez, E.; Bedmar, E.J. Bacterial associations with legumes. Crit. Rev. Plant Sci. 2015, 34, 17–42. [Google Scholar] [CrossRef]
- Farooq, M.; Gogoi, N.; Barthakur, S.; Baroowa, B.; Bharadwaj, N.; Alghamdi, S.S. Drought Stress in Grain Legumes during Reproduction and Grain Filling. J. Agron. Crop Sci. 2017, 203, 81–102. [Google Scholar] [CrossRef]
- Yagoob, H.; Yagoob, M. The effects of water deficit stress on protein yield of mung bean genotypes. Peak. J. Agri. Sci. 2014, 2, 30–35. [Google Scholar]
- Ghanbari, A.A.; Shakiba, M.R.; Toorchi, M.; Choukan, R. Nitrogen changes in the leaves and accumulation of some minerals in the seeds of red, white and chitti beans (Phaseolus vulgaris) under water deficit conditions. Aus. J. Crop Sci. 2013, 7, 706–712. [Google Scholar]
- Ashrafi, V.; Pourbozorg, H.; Kor, N.M.; Ajirloo, A.R.; Shamsizadeh, M.; Shaaban, M. Study on seed protein and protein profile pattern of chickpea (Cicer arietinum L.) by SDS-PAGE under drought stress and fertilization. Int. J. Life Sci. 2015, 9, 87–90. [Google Scholar] [CrossRef] [Green Version]
- Ardakani, L.G.; Farajee, H.; Kelidari, A. The effect of water stress on grain yield and protein of spotted bean (Phaseolus vulgaris L.), cultivar Talash. Int. J. Adv. Biol. Biomed. Res. 2013, 1, 940–949. [Google Scholar]
- Baroowa, B.; Gogoi, N. Biochemical changes in black gram and green gram genotypes after imposition of drought stress. J. Food Legume 2014, 27, 350–353. [Google Scholar]
- Maleki, A.; Naderi, A.; Naseri, R.; Fathi, A.; Bahamin, S.; Maleki, R. Physiological performance of soybean cultivars under drought stress. Bull. Environ. Pharmacol. Life Sci. 2013, 2, 38–44. [Google Scholar]
- Kyei-boahen, S.; Savala, C.E.N.; Chikoye, D.; Abaidoo, R.; Kyei-boahen, S. Growth and yield responses of cowpea to inoculation and phosphorus fertilization in different environments. Front. Plant Sci. 2017, 8, 646. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.; Jin, J.; Jiang, S.; Ning, S.; Liu, L. Quantitative response of soybean development and yield to drought stress during different growth stages in the Huaibei Plain, China. Agronomy 2018, 8, 97. [Google Scholar] [CrossRef] [Green Version]
- Varshney, R.K.; Thudi, M.; Nayak, S.N.; Gaur, P.M.; Kashiwagi, J.; Krishnamurthy, L.; Jaganathan, D.; Koppolu, J.; Bohra, A.; Tripathi, S. Genetic dissection of drought tolerance in chickpea (Cicer arietinum L.). Theor. Appl. Genet. 2014, 127, 445–462. [Google Scholar] [CrossRef]
- Kazai, P.; Noulas, C.; Khah, E.; Vlachostergios, D. Yield and seed quality parameters of common bean cultivars grown under water and heat stress field conditions. AIMS Agric. Food 2019, 4, 285–302. [Google Scholar] [CrossRef]
- Khan, R.; Srivastava, R.; Abdin, M.Z.; Manzoor, N.M. Effect of soil contamination with heavy metals on soybean seed oil quality. Eur. Food Res. Technol. 2013, 236, 707–714. [Google Scholar] [CrossRef]
- Allahmoradi, P.; Mansourifar, C.; Saiedi, M.; Jalali Honarmand, S. Effect of different water deficiency levels on some antioxidants at di erent growth stages of lentil (Lens culinaris L.). Adv. Environ. Biol. 2013, 7, 535–543. [Google Scholar]
- Bellaloui, N.; Mengistu, A.; Kassem, M.A. Effects of genetics and environment on fatty acid stability in soybean seed. Food Nutr. Sci. 2013, 4, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Rozrokh, M.; Sabaghpour, S.H.; Armin, M.; Asgharipour, M. The effects of drought stress on some biochemical traits in twenty genotypes of chickpea. Eur. J. Exp. Biol. 2012, 2, 1980–1987. [Google Scholar]
- Ahmad, A.; Selim, M.M.; Alderfasi, A.A.; Afzal, M. Effect of drought stress on mungbean (Vigna radiata L.) under arid climatic conditions of Saudi Arabia. Ecosyst. Sustain. Dev. 2015, 192, 185–193. [Google Scholar]
- Farooq, M.; Hussain, M.; Wakeel, A.; Siddique, K.H.M. Salt stress in maize: Effects, resistance mechanisms and management. A review. Agron. Sustain. Dev. 2015, 35, 461–481. [Google Scholar] [CrossRef] [Green Version]
- El Sayed, H.E.S.A. Influence of NaCl and Na2SO4 treatments on growth development of broad bean (Vicia faba L.). Plant J. Life Sci. 2011, 5, 513–523. [Google Scholar]
- Farooq, M.; Gogoi, N.; Hussain, M.; Barthakur, S.; Paul, S.; Bharadwaj, N.; Migdadi, H.M.; Alghamdi, S.S.; Siddique, K.H. Effects, tolerance mechanisms and management of salt stress in grain legumes. Plant Physiol. Biochem. 2017, 118, 199–217. [Google Scholar] [CrossRef]
- Talei, D.; Kadir, M.A.; Yusop, M.K.; Abdullah, M.P.; Valdiani, A. Salinity effects on macro and micronutrients uptake in medicinal plant King of Bitters (Andrographis paniculata Nees.). Plant OMICS 2012, 5, 271–278. [Google Scholar]
- Torabian, S.; Farhangi-Abriz, S.; Rathjen, J. Biochar and lignite affect H+-ATPase and H+-PPase activities in root tonoplast and nutrient contents of mungbean under salt stress. Plant Physiol. Biochem. 2018, 129, 141–149. [Google Scholar] [CrossRef]
- Nadeem, M.; Li, J.; Yahya, M.; Wang, M.; Ali, A.; Cheng, A.; Wang, X.; Ma, C. Grain legumes and fear of salt stress: Focus on mechanisms and management strategies. Int. J. Mol. Sci. 2019, 20, 799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, H.A.; Siddique, K.H.M.; Colmer, T.D. Vegetative and reproductive growth of salt stressed chickpea are carbon-limited: Sucrose infusion at the reproductive stage improves salt tolerance. J. Exp. Bot. 2017, 68, 2001–2011. [Google Scholar] [CrossRef]
- Zhou, R.; Hyldgaard, B.; Yu, X.; Rosenqvist, E.; Ugarte, R.M.; Yu, S.; Wu, Z.; Ottosen, C.O.; Zhao, T. Phenotyping of faba beans (Vicia faba L.) under cold and heat stresses using chlorophyll fluorescence. Euphytica 2018, 214, 1–13. [Google Scholar] [CrossRef]
- Amira, M.S.; Qados, A. Effect of arginine on growth, nutrient composition, yield and nutritional value of mung bean plants grown under salinity stress. Nat. Sci. 2010, 8, 30–42. [Google Scholar]
- Sehrawat, N.; Yadav, M.; Sharma, A.K.; Kumar, V.; Bhat, K.V. Salt stress and mungbean [Vigna radiata (L.) Wilczek]: Effects, physiological perspective and management practices for alleviating salinity. Arch. Agron. Soil Sci. 2019, 65, 1287–1301. [Google Scholar] [CrossRef]
- Ghassemi-Golezani, K.; Nikpour-Rashidabad, N.; Zehtab-Salmasi, S. Effect of salinity on yield and yield components of pinto bean cultivars. Int. J. Plant Anim. Environ. Sci. 2012, 2, 47–51. [Google Scholar]
- Ahmad, M.; Zahir, Z.A.; Asghar, H.N.; Asghar, M. Inducing salt tolerance in mung bean through coinoculation with rhizobia and plant-growth-promoting rhizobacteria containing 1- aminocyclopropane-1-carboxylate deaminase. Can. J. Microbiol. 2011, 57, 578–589. [Google Scholar] [CrossRef]
- Khan, M.S.A.; Karim, M.A.; Haque, M.M.; Islam, M.M.; Karim, A.J.M.S.; Mian, M.A.K. Influence of salt and water stress on growth and yield of soybean genotypes. Trop. Agric. Sci. 2016, 39, 167–180. [Google Scholar]
- Narula, S.; Anand, R.C.; Dudeja, S.S. Beneficial traits of endophytic bacteria from field pea nodules and plant growth promotion of field pea. J. Food. Legume 2013, 26, 73–79. [Google Scholar]
- Hossain, M.A.; Hasanuzzaman, M.; Fujita, M. Up-regulation of antioxidant and glyoxalase systems by exogenous glycinebetaine and proline in mung bean confer tolerance to cadmium stress. Physiol. Mol. Biol. Plants 2010, 16, 259–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pokhrel, D.; Bhandari, B.S.; Viraraghavan, T. Arsenic contamination of groundwater in the Terai region of Nepal: An overview of health concerns and treatment options. Environ. Inter. 2009, 35, 157–161. [Google Scholar] [CrossRef]
- Shi, G.; Liu, C.; Cai, Q.; Liu, Q.; Hou, C. Cadmium Ac- cumulation and Tolerance of Two Safflower Cultivars in Relation to Photosynthesis and Antioxidantive Enzymes. Bull. Environ. Contam. Toxicol. 2010, 85, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Asati, A.; Pichhode, M.; Nikhil, K. Effect of heavy metals on plants: An overview. Int. J. Appl. Innov. Eng. Manag. 2016, 5, 56–66. [Google Scholar]
- Bae, J.; Benoit, D.L.; Watson, A.K. Effect of heavy metals on seed germination and seedling growth of common ragweed and roadside ground cover legumes. Environ. Pollut. 2016, 213, 112–118. [Google Scholar] [CrossRef]
- Bahmani, R.; Bihamta, M.R.; Habibi, D.; Forozesh, P.; Ahmadvand, S. Effect of cadmium chloride on growth parameters of different bean genotypes (Phaseolus vulgaris L.). ARPN J. Agri. Biol. Sci. 2012, 7, 35–41. [Google Scholar]
- Dewan, M.M.; Dhingra, H.R. Cadmium partitioning and seed quality in two varities of pea and their hybrid as influenced by rhizopheric cadmium. Indian J. Plant Physiol. 2004, 9, 15–20. [Google Scholar]
- Malik, J.A.; Goel, S.; Sandhir, R.; Nayyar, H. Uptake and distribution of arsenic in chick pea:Effects on seed yield and seed composition. Commun. Soil Sci. Plant Anal. 2011, 42, 1728–1738. [Google Scholar] [CrossRef]
- Wei, J.; Cen, K. Contamination and health risk assessment of heavy metals in cereals, legumes, and their products: A case study based on the dietary structure of the residents of Beijing, China. J. Clean. Prod. 2020, 260, 121001. [Google Scholar] [CrossRef]
- Valentine, A.J.; Benedito, V.A.; Kang, Y. Legume nitrogen fixation and soil abiotic stress: From physiology to genomics and beyond. In Nitrogen Metabolism in Plants in the Post-Genomic Era; Annual Plant Reviews Book Series; Wiley Online Library: Hoboken, NJ, USA, 2018; Volume 42, pp. 207–248. [Google Scholar]
- Kantar, F.; Shivakumar, B.G.; Arrese-Igor, C.; Hafeez, F.Y.; González, E.M.; Imran, A.; Larrainzar, E. Efficient biological nitrogen fixation under warming climates. In Climate Change and Management of Cool Season Grain Legume Crops; Springer: Dordrecht, The Netherland, 2010; pp. 283–306. [Google Scholar]
- Abdel-Wahab, A.; Abdel-Muhsin, A.M.A.; Ali, E.; Suleiman, S.; Ahmed, S.; Walliker, D.; Babiker, H.A. Dynamics of gametocytes among Plasmodium falciparum clones in natural infections in an area of highly seasonal transmission. J. Infect. Dis. 2002, 12, 1838–1842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abd-Alla, M.H.; Nafady, N.A.; Bashandy, S.R.; Hassan, A.A. Mitigation of effect of salt stress on the nodulation, nitrogen fixation and growth of chickpea (Cicer arietinum L.) by triple microbial inoculation. Rhizosphere 2019, 10, 100148. [Google Scholar] [CrossRef]
- Abd-Alla, M.H.; Issa, A.A.; Ohyama, T. Impact of harsh environmental conditions on nodule formation and dinitrogen fixation of legumes. Adv. Biol. Ecol. Nitrogen Fixat. 2014, 9, 1. [Google Scholar]
- Singh, S.; Parihar, P.; Singh, R.; Singh, V.P.; Prasad, S. Heavy metal tolerance in plants: Role of transcriptomics, proteomics, metabolomics, and ionomics. Front. Plant Sci. 2016, 6, 1143. [Google Scholar] [CrossRef] [Green Version]
- Wani, P.; Khan, M. Bioremediaiton of lead by a plant growth promoting Rhizobium species RL9. Bacteriol. J. 2012, 2, 66–78. [Google Scholar] [CrossRef] [Green Version]
- Meena, R.S.; Yadav, R.S.; Meena, H.; Kumar, S.; Meena, Y.K.; Singh, A. Towards the current need to enhance legume productivity and soil sustainability worldwide: A book review. J. Clean. Prod. 2015, 104, 513–515. [Google Scholar] [CrossRef]


{kind=link}
| Sl. No | Picture | Common Name | Scientific Name | Major Use | Sources of Images [Accessed on 22 May 2021] |
|---|---|---|---|---|---|
| 1. |  | Soybean | Glycine max | Mainly used for soybean oil. Additionally used as food products such as soymilk, soy sauce, some beverages and whipped toppings, soy-fortified pastas, breakfast cereals and bars [14]. | https://zh-prod-1cc738ca-7d3b-4a72-b792-20bd8d8fa069.storage.googleapis.com/s3fs-public/styles/max_650×650/public/2020-08/soybeans.jpg?itok=DuPfsOBn (accessed on 22 May 2021) |
| 2. |  | Chickpea | Cicer arietinum | Used as a dry pulse and also as a green vegetable [14,15]. | https://agtfoods.co.za/wp-content/uploads/2018/06/Desi-Chickpea_600x600_1.jpg (accessed on 22 May 2021) |
| 3. |  | Pea | Pisum sativum | Used both fresh and dried. Peas are rich in protein, cholesterol-free and have good amounts of dietary fiber [14]. | https://www.allergicliving.com/wp-content/uploads/2019/09/Green-peas.jpg (accessed on 22 May 2021) |
| 4. |  | Groundnut | Arachis hypogaea | Groundnuts contains high level of monounsaturated and polyunsaturated fatty acids that may keep the heart healthy by maintaining lower blood cholesterol levels [16]. | https://www.nutstop.com/how-peanuts-grow/ (accessed on 22 May 2021) |
| 5. |  | Red lentil | Lens culinaris | Red lentils contain plenty of protein and fiber; thus, it is called the meat of poor people. It is a healthier choice for the heart instead of processed meat [14]. | https://jiraphaserviceltd.com/wp-content/uploads/2020/10/7RLT1-1.jpg (accessed on 22 May 2021) |
| 6. |  | Green lentil | Lens culinaris | It is a rich source of protein and a good supplement for meat. It reduces the risk of heart diseases [17]. | https://www.grainstar.com.au/wp-content/uploads/2018/05/richlea-lentils.jpg (accessed on 22 May 2021) |
| 7. |  | Brown lentil | Lens culinaris | Brown lentils are a good source of nutrients and its low calorie content and high fiber helps healthy digestion [16]. | https://fthmb.tqn.com/2JdDB8mU_-S9vxjJ6bik4O763sg=/960x0/filters:no_upscale()/170460595-56a30edc5f9b58b7d0d03627.jpg (accessed on 22 May 2021) |
| 8. |  | Black lentil | Lens culinaris | Black lentils are the most flavorful lentils and are quite different from other lentils, used in salads and soups [17]. | https://agtfoods.co.za/wp-content/uploads/2018/06/Black-Lentils_3.jpg (accessed on 22 May 2021) |
| 9. |  | Mung bean | Vigna radiata | Contains essential amino acids and antioxidants that help to neutralize free radicals, thus working against chronic inflammation, heart disease, cancers and other diseases [14,18]. | https://www.espaceagro.com/_AFFAIRE/188567.jpg (accessed on 22 May 2021) |
| 10. |  | Black eyed bean | Vigna unguiculata | Improves digestion and its adequate iron content helps to prevent anemia. It is rich in potassium that helps to maintain lower blood pressure [14,19]. | https://agtfoods.co.za/wp-content/uploads/2018/06/Black-Eyed-Beans_600 x600_1.jpg (accessed on 22 May 2021) |
| 11. |  | Fayot bean | Phaseolus vulgaris | It is a source of dietary fiber that helps to prevent cholesterol absorption and increase the fat elimination [14]. | https://www.onlyfoods.net/different-types-of-beans.html (accessed on 22 May 2021) |
| 12. |  | Navy bean | Phaseolus vulgaris | This bean has anticancer potential. It also helps to lower diabetes risk and greater gut health [14]. | https://www.thedailymeal.com/sites/default/files/slideshows/1670994/2173040/21-navy_beans-ThinkstockPhotos-494876324.jpg (accessed on 22 May 2021) |
| 13. |  | Red bean | Vigna umbellata | Protects the body from free radical damage that helps in controlling blood sugar levels [16]. | http://productkg.com/sites/default/files/tomatnaya-fasoltalas_0.jpg (accessed on 22 May 2021) |
| 14. |  | Red kidney bean | Phaseolus vulgaris | Red kidney beans are full of folate (vitamin B9) and fiber, which helps to promote cardiovascular health [18]. | https://www.foodsafetynews.com/files/2020/07/dreamstime_red-kidney-bean-lectins.jpg (accessed on 22 May 2021) |
| 15. |  | White kidney bean (Cannellini) | Phaseolus vulgaris | It helps in blocking the carbs from being absorbed and metabolized in the human body [18]. | https://ixivixi.com/wp-content/uploads/2015/07/White-Kidney-Bean-Extract-for-Weight-Loss-1.jpg (accessed on 22 May 2021) |
| 16. |  | Pinto bean | Phaseolus vulgaris | Contains a good amount of vitamin B1 that helps to convert food into energy. Additionally, it contains many antioxidants such as polyphenols and flavonoids [16,17]. | https://cdn.shopify.com/s/files/1/1834/0943/products/bean-pinto_569fa089-dddd-41b4-856d-3db099771330_800x.png?v=1505218437 (accessed on 22 May 2021) |
| 17. |  | Cranberry bean | Phaseolus vulgaris | It is good for the heart as it contains various powerful minerals and enzymes that help to lower bad cholesterol [18]. | https://www.mexicanplease.com/wp-content/uploads/2017/03/cranberry-beans-spread-onto-cutting-with-solids.jpg (accessed on 22 May 2021) |
| 18. |  | Adzuki bean | Vigna angularis | It helps to balance sugar level and reduces the risk of diabetes. Improves the strength of bones [20]. | https://www.suttonsbaytrading.com/wp-content/uploads/2013/06/adzuki-beans.jpg (accessed on 22 May 2021) |
| 19. |  | Faba bean (Broad bean) | Vicia faba | It helps to prevent birth defects as it is incredibly nutritious and an excellent source of soluble fiber, protein, manganese, copper folate and many other micronutrients [16]. | http://storage.googleapis.com/powop-assets/kew_profiles/KPPCONT_085134_fullsize.jpg (accessed on 22 May 2021) |
| 20. |  | Lima bean | Phaseolus lunatus | Helps to prevent chronic disease, diseases associated with digestion and stimulates blood circulation [14]. | https://judiesblog.files.wordpress.com/2010/10/img_5993.jpg (accessed on 22 May 2021) |
| 21. |  | Grass pea | Lathyrus sativus | Grass pea seeds are used as a common staple food in many countries of Asia and Africa [21]. | https://cdn.shopify.com/s/files/1/2333/6781/products/[email protected]?v=1578338252 (accessed on 22 May 2021) |
| 22. |  | Lupin bean | Lupinus albus | It contains antioxidants that promote proper digestion and keep intestines healthy. Additionally, aids in weight loss, provides essential vitamins and minerals and reduces high blood pressure [18,22]. | https://www.firstforwomen.com/wp-content/uploads/sites/2/2019/01/what-are-lupin-beans-benefits.jpg?w=715 (accessed on 22 May 2021) |
| 23. |  | Common bean | Phaseolus vulgaris | Beans are an excellent source of protein, lower in calories and saturated fat than some other protein sources such as meat [16,17]. | https://garden.org/pics/2018-09-14/Alicemac/b92064.jpg (accessed on 22 May 2021) |
| 24. |  | Runner bean | P. multiflorus | The dried pods of runner bean have diuretic properties that help to cure urinary tract infections and reduce weight [18,23]. | http://pebbleandfern.ca/wp-content/uploads/2015/03/runnerbean.jpg (accessed on 22 May 2021) |
| 25. |  | Hyacinthbean | Lablab purpureus | It contains complex carbohydrates and a good amount of zinc, which is useful to lose weight and prevent cancer, respectively, as Zn prevents cells mutating and assists cell division [16,22]. | https://www.healthbenefitstimes.com/9/gallery/hyacinth-beans/Pods-of-Hyacinth-beans.jpg (accessed on 22 May 2021) |
| 26. |  | Rice bean | V. umbellata | It helps to cure diseases such as edema and increases digestibility [17,23]. | https://www.feedipedia.org/sites/default/files/images/vigna_umbellata_seeds.jpg (accessed on 22 May 2021) |
| 27. |  | Black gram | V. mungo | It helps in boosting energy, protecting cardiovascular health, improving immunity, maintaining skin health, building strong bones, managing diabetes and strengthening the nervous system [16,18]. | https://cdn.shopify.com/s/files/1/2600/9462/products/kaliblack_large.jpg?v=1516478931 (accessed on 22 May 2021) |
| 28. |  | Pigeon pea | Cajanus cajan | It helps to manage blood pressure, boost heart health and prevent anemia, thus strengthening the immune system [18,20,22]. | https://www.specialtyproduce.com/sppics/11653.png (accessed on 22 May 2021) |
| 29. |  | Velvet bean | Mucuna pruriens | Supplies L-Dopa that turns into dopamine, which helps to improve mood, mental clarity, sense of well-being, better sleep and brain function. Additionally, it helps to combat Parkinson’s disease and depression [18]. | https://www.healthbenefitstimes.com/9/gallery/velvet-bean/Pods-of-Velvet-beans.jpg (accessed on 22 May 2021) |
| 30. |  | Winged bean | Psophocarpus tetragonolobus | This bean is rich in vitamin C and vitamin A, which help in strengthening the immune system and supporting the body against any possible infections and diseases [16,22,23]. | https://i.pinimg.com/originals/21/29/fa/2129fa4b595f818e23046dd8aae4b290.png (accessed on 22 May 2021) |
| 31. |  | Sword bean | Canavalia gladiata | It enhances the function of the nervous system, prevents bone resorption and inhibits bone turn over [16,17,18]. | https://i.etsystatic.com/7772783/r/il/442d44/1253663464/il_fullxfull.1253663464_bodq.jpg (accessed on 22 May 2021) |
| 32. |  | Jack bean | Canavalia. ensiformis | It is a fiber-rich bean that helps in removing toxins and waste products in the gut. Helps in preventing constipation and abdominal distention. The Vitamin C present in this bean helps in defending the body against disease-causing microorganisms such as bacteria and viruses [23]. | https://i.etsystatic.com/15567684/r/il/fffdef/2833501891/il_794xN.2833501891_8key.jpg (accessed on 22 May 2021) |
| Food Legumes | Protein % | Carbohydrate% | Lipid% |
|---|---|---|---|
| Chickpea | 21 | 62.95 | 6.04 |
| Groundnut | 26 | 16.13 | 49.24 |
| Lentil | 25 | 63.35 | 1.06 |
| Black gram | 25 | 60 | 1.64 |
| Mung bean | 24 | 62.62 | 1.15 |
| Soybean | 40 | 6.4 | 21.3 |
| Pea | 25 | 51.3 | 1.2 |
| Pigeon pea | 22 | 62.78 | 1.49 |
| Cowpea | 24 | 35.5 | 0.91 |
| Faba bean | 29 | 44.7 | 1.4 |
| White lupin | 38 | 0.0 | 10.0 |
| Adzuki bean | 20 | 62.90 | 0.53 |
| Navy bean | 22 | 60.75 | 1.50 |
| Lima bean | 21 | 63.38 | 0.69 |
| Legume | Phenolic Compounds | Quantity (µg/g) | Found in | References |
|---|---|---|---|---|
| Lentil | Hydroxybenzoics | 5.69 | Seed | [18,32,33] |
| Dihydroxybenzoic acid | 3.68 | |||
| p-hydroxybenzoic acid | 1.48 | |||
| Protocatechuic acid | 0.36 | |||
| Protocatechuic aldehyde | 0.13 | |||
| 2,3,4-trihydroxybenzoic acid | 16.9–29.2 | |||
| Gallic acid | 90.9–136.8 | |||
| Vanillic acid | 0.59–3.22 | |||
| Hydroxycinnamics | 3.76 | |||
| Trans-p-coumaroyl malic acid | 10.02 | |||
| Trans-p-coumaroyl glycolic acid | 2.88 | |||
| Trans-p-coumaric acid | 5.74 | |||
| Sinapic acid | 1099–2217 | |||
| chlorogenic acid | 159–213 | |||
| Green lentil | Trans-p-coumaric acid | 37.3 | Seed | [18,32] |
| Trans-p-coumaric acid derivative | 6.4 | |||
| Trans-ferulic acid | 10.1 | |||
| Pinto bean | Hydroxybenzoics, | 84.92 | Seed | [18,32,33] |
| Salicyclic acid | 44.89 | |||
| Vanillic acid | 17.01 | |||
| P-hydroxybenzoic acid | 12.20 | |||
| P-hydroxyphenyl acetic acid | 8.42 | |||
| Protocatechuic acid | 2.40 | |||
| Hydroxycinnamic acids | 36.31 | |||
| Trans-ferulic acid | 11.80 | |||
| Cannellini bean | Hydroxybenzoics, | 21.66 | Seed | [32,33] |
| Vanillic acid | 10.71 | |||
| P-hydroxyphenyl acetic acid | 6.92 | |||
| P-hydroxybenzoic acid | 4.30 | |||
| Hydroxycinnamic acids | 23.52 | |||
| Trans-ferulic acid | 8.95 | |||
| Adzuki bean | Protocatechuic acid | 67.6 | Crude extract | [18,20,32] |
| Protocatechuic aldehyde | 7.71 | |||
| Trans-p-coumaric acid | 31.3 | Seed | [18,32,33] | |
| Trans-p-coumaroyl malic acid | 4.57 | |||
| Cowpea | Gallic | 27 | Seed coat | [32] |
| Protocatechuic | 18.9 | |||
| P-hydroxybenzoic | 5.81 | |||
| Ferulic | 26.25 | Seed | ||
| Coumaric acid | 1.25 | |||
| Cranberry beans | Protocatechuic | 217 | Seed coat | [32,34] |
| P-hydroxybenzoic acid | 239 | |||
| Chickpea | P-hydroxybenzoic acid | 19.2 to 60.5 | Seed | [18,32,35] |
| Syringic acid | 45.9 | |||
| Gentisic acid | 8.1 to 26.0 | |||
| Pea | Protocatechuic acid | 12.1 to 163.5 | Seed | [32,35] |
| P-hydroxybenzoic acid | 45.4 to 101.7 | |||
| soybean | Benzoic acids | 57 | Seed | [32,33] |
| Protocatechuic acids | 44 | |||
| Ferulic acid | 95 | |||
| kidney | P-hydroxybenzoic | 10.33 | Bean | [32] |
| 10 | Sprout |
| Vitamins and Minerals | Soybean (Per 100 g Seed) | Chickpea (Per 100 g Seed) | Pea (Per 100 g Seed) | Grass Pea (Per 100 g Seed) | Cowpea (Per 100 g Seed) | Pigeon Pea (Per 100 g Seed) | Groundnut (Per 100 g Seed) | Lentil (Per 100 g Seed) | Mung Bean (Per 100 g Seed) | Faba Bean (Per 100 g Seed) | Lupin (Per 100 g Seed) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| α tocopherol | 6.5 mg | 2.24 mg | 0.11 mg | - | - | - | - | - | - | 0.08 mg | 1.1 mg |
| γ tocopherol | 23.0 mg | 10.68 mg | 5.0 mg | - | - | - | - | - | - | - | 15.3 mg |
| Vitamin B1 | 1.0 mg | 0.477 mg | 0.7 mg | 0.37–0.54 mg | 0.345 mg | 0.643 mg | 0.64 mg | 0.87 mg | 0.621 mg | 0.55 mg | 0.32 mg |
| Vitamin B2 | 0.46 mg | 0.212 mg | 0.27 mg | 0.18–0.27 mg | 0.094 mg | 0.187 mg | 0.135 mg | 0.21 mg | 0.233 mg | 0.23 mg | 0.59 mg |
| Vitamin B3 | - | 1.541 mg | - | 1.23–2.02 mg | - | 2.96 mg | 12.06 mg | 2.6 mg | 2.251 mg | - | - |
| Vitamin B5 | - | 1.588 mg | - | 1.44–2.24 mg | 0.703 mg | 1.26 mg | 1.76 mg | 2.14 mg | - | - | - |
| Vitamin B6 | 1.1 mg | 0.55 mg | 0.12 mg | 0.49–0.66 mg | 0.171 mg | 0.283 mg | 0.348 mg | 0.5 mg | 0.382 mg | 0.37 mg | 0.4 mg |
| β-Carotene | - | 40.00 mg | - | 24.08–41.01 μg | - | - | - | - | 68 μg | - | - |
| Vitamin K | - | 9.00 mg | - | - | 1.7 µg | - | - | 5.0 µg | - | - | - |
| Calcium | 0.21 g | 160 mg | 0.05 g | 0.97–1.03 g | - | 130 mg | 92 mg | 35 mg | 132 mg | 0.14 g | 0.24 g |
| Potassium | 1.8 g | 875.0 mg | 1 g | 8.75–9.2 g | 475 mg | 1392 mg | 705 mg | 677 mg | 1246 mg | 1.2 g | 1.1 g |
| Magnesium | 0.22 g | 138 mg | 0.12 g | 1.14–1.24 g | 91 mg | 183 mg | 168 mg | 47 mg | 189 mg | 0.15 g | 0.13 g |
| Phosphorus | - | 366.0 mg | - | 4.68–5.13 g | 267 mg | - | 376 mg | 281 mg | 367 mg | - | - |
| Iron | 8.0 mg | 5.0 mg | 5.2 mg | 1.33–1.53 mg | 4.29 mg | 5.23 mg | 4.58 mg | 6.51 mg | 6.74 mg | 6.7 mg | 5.4 mg |
| Copper | 1.2 mg | 0.847 mg | 0.66 mg | 6.98–7.95 g | 0.458 mg | 1.057 mg | 1.144 mg | 0.75 mg | 0.941 μg | 1.1 mg | 0.6 mg |
| Zinc | 4.2 mg | 4.1 mg | 3.2 mg | 4.35 mg | 2.21 mg | 2.76 mg | 3.27 mg | 3.27 mg | 2.68 mg | 4.1 mg | 5.1 mg |
| Selenium | 19 µg | - | 1.6 µg | - | - | - | - | 0.1 µg | 8.2 μg | 2 µg | 4.7 µg |
| Food Legumes | Threshold Temp. | Growth Stage | Heat Stress (Day/Night) | Effects | References |
|---|---|---|---|---|---|
| Lentil | 15–30 | Reproductive stage | 38/23 | Reduced electron flow during photosynthesis | [56,57,58,59] |
| Peanut | 30–35 | Vegetative development | 38/22 | Decreased pollen production, impaired photosystem II | [56,57] |
| 23 | Anthesis | ||||
| Pod and grain yield | |||||
| Pea | 15–25 | Vegetative growth | 30/25 | Reduced photosynthetic activity; impeded electron donation by OEC (Oxygen-Evolving Center) of PS II; reduced oxygen evolution and photochemical energy storage; shutting of PSI reaction center | [57,59,60] |
| Chickpea | 15–30 | Growth | 35/16 | Impaired RuBisCO and sucrose metabolism in leaves; disrupted PSII; damaged structure and functioning of related enzymes and proteins; decreased stigma receptivity | [56,59] |
| 25 | Reproductive growth | ||||
| Pigeon pea | 18–30 | Flowering | 45/40 | Damaged PSII | [56,59] |
| Cowpea | 18–28 | Flowering | 36/27 | Tapetal cells degeneration and anther indehiscence | [56,57,59] |
| Soybean | 26 | Reproductive | 38/30 | Abscission of flower, reduced reproductive development; pollen germination, pollen tube growth and yield; shrunken pollen; damaged PSII; reduced chlorophyll content and photosynthesis; decreased Fv/Fm | [56,57,58,59,60,61] |
| 23 | Post-anthesis | 35 | |||
| 30.2 | Pollen germination | 35 | |||
| 36.1 | Pollen tube growth | 38/30 | |||
| Common bean | 20–24 | Flowering | 32/27 | Carbon assimilation limited and NADPH supply reduced; reduced photosynthetic rate | [59,62] |
| Mung bean | 28–35 | Flowering | >40/25 | Efficiency of photosynthesis impaired; reduced sucrose in leaves due to decreased sucrose synthesizing enzymes and RuBisCO activity | [57,59,63] |
| Pod development | |||||
| Broad bean | 25–35 | Flowering | 42 | Reduced photosynthesis | [57,59,64] |
| Black gram | 25–35 | Flowering | 35 | Reduced photosynthesis | [65] |
| Lupin | 20–30 | Flowering | 38 | Cytokinin level reduced in seed leading to diminished seed cell numbers and growth rates of seed, reduced seed growth and development processes | [57,59] |
| Food Legumes | Temperature | Grain Composition | Increase % (+) or Decrease % (−) over Control | References | |
|---|---|---|---|---|---|
| Control (Day/Night) | Heat Stress (Day/Night) | ||||
| Soybean | 15/30 °C | 40/30 °C | Oleic acid | +104% | [1,66] |
| Linolenic acid | −48.6 | ||||
| 18/13 °C | 33/28 °C | Oil content | +37% | ||
| 18/13 °C | 33/28 °C | Sucrose | −56% | ||
| Peanut | 20/14 °C | 32/26 °C | Total sugars | −24.5% | [1,66] |
| Starch | −53% | ||||
| Protein | −19.6% | ||||
| 20/14 °C | 26/20 °C | Oil content | +20% | ||
| Oleic acid | +24% | ||||
| Chickpea | 25 | 35/16 | Soluble proteins | +20% | [1,66,68,76] |
| 25 | ˃32/20 °C | Sucrose content | −9% | ||
| Kidney bean | 28/18 °C | 34/24 °C | Oil content | −22.7% | [1,66,71] |
| Food Legumes | Cold Stress | Effects |
|---|---|---|
| Soybean | 1 °C for 4, 6 and 8 h | Early vegetative phase damage, impaired microsporogenesis and megasporogenesis, loss of pollen germination, inhibition of pollen tube growth, abnormal pod formation and seed filling [54] and alteration in starch, protein, fat and fiber composition [78] |
| Pea | 3 °C | Early vegetative phase damage, reduction in embryogenesis and poor seed quality [54] |
| Chickpea | ˂10 °C; −10 °C for 15–30 min | Early vegetative phase damage, impaired microsporogenesis and megasporogenesis, pollen viability loss, loss of pollen germination, stigma receptivity loss, abnormal pod formation [79] and seed filling [54] and alteration in starch, protein, fat and fiber composition [78] |
| Broad bean | 5 °C for 24 h | Early vegetative phase damage and poor seed quality [54] |
| Food Legumes | Drought Stress at Growth Stages | Effects | References |
|---|---|---|---|
| Lentil | Pod development and reproductive phase | Yield reduction by 70 and 24%, respectively | [87] |
| Chickpea | Reproductive phase, anthesis and late ripening | Yield loss by 49–54, 27–40 and 49–54%, respectivelyGrain protein, sodium, potassium and calcium content reduced by 41, 33, 25 and 7%, respectively | [88,89] |
| Soybean | Reproductive phase, pod set and Seed filling | Oil content of grain reduced by 3% and protein content increased by 5% Loss of grain yield by 46–71, 45–50 and 42%, respectively | [90,91,92] |
| Common bean | Reproductive, flowering and Pod filling stage | Sucrose and starch content reduced in grain by 29–47 and 18–20% Yield loss by 58–87, 49 and 40%, respectively | [93] |
| Mung bean | Reproductive and vegetative stage | Grain protein content increased by 8 and 3%, respectively Yield reduction by 26% | [94] |
| Faba bean | Grain filling | Carbohydrate, fat and protein content increased by 4, 5 and 3–9%, respectively Grain yield loss by 68% | [83] |
| Spotted bean | Reproductive stage | Protein content of grain increased by 6% | [87] |
| Black gram (Mash bean) | Flowering and reproductive | Loss of grain yield by 31–57 and 26%, respectively | [95] |
| Cowpea | Reproductive and pod filling | Yield loss by 34–66 and 29%, respectively | [92] |
| Pigeon pea | Reproductive phase and flowering | Grain yield loss by 40–55 and 42–57% | [83] |
| Lupins | 15 days after anthesis | Reduction in soluble sugar, crude fiber and starch in grain by 18, 11 and 43%, respectively | [83] |
| Food Legumes | Concentration of Salt | Impacts |
|---|---|---|
| Soybean | NaCl 3, 6 and 9 dS m−1 | Grain protein reduction by 29, 60 and 79%, respectively |
| NaCl 9 dS m−1 | Oil content of grain reduced by 77% | |
| 7 dS m−1 in loam soil and 6.3 dS m−1 clay soil | Yield loss around 46% | |
| Chickpea | 3 and 3.8 dS m−1 | Loss of grain yield by 50 and 69%, respectively |
| 50 and 100 mM | Sodium increased by 200 and 271%, respectively | |
| 50 and 100 mM | Potassium decreased by 79.09 and 72.72%, respectively | |
| 2 and 9 dS m−1 | Sodium increased by 79.80% and Potassium increased by 0.58% | |
| NaCl 40 mM | Increase in sodium, 51.03%; potassium, 40.31%; and chloride, 58.41% | |
| Lentil (cv. 6796) | 3.1 and 2 dS m−1 | Grain yield loss found to be 100 and 14%, respectively |
| Mung bean | 4500 and 6000 ppm | Reduction in grain protein content of 11 and 20%, respectively |
| Reduction in total soluble sugars of 29 and 32%, respectively | ||
| Reduction in total amino acids of 19 and 21%, respectively | ||
| Nitrogen content in grain decreased by 37 and 24%, respectively | ||
| Grain phosphorus content decreased by 30 and 20%, respectively | ||
| Reduction in grain potassium content by 13 and 8%, respectively | ||
| 250 mM NaCl | ≥80–100% yield loss | |
| Mungbean (cv. Pusavishal) | 50 mM NaCl | Yield loss by 41% |
| Faba bean | 6.6 dS m−1 in loam soil | Total yield loss around 50% |
| 5.6 dS m−1 in clay soil | Yield loss by 52% | |
| 50 and 100 mM | Total carbohydrates of grain reduced by 9.97 and 33.40%, respectively | |
| Decrease in grain potassium content of 3.30 and 11.57%, respectively | ||
| Increase in sodium content of around 12.5 and 62.5%, respectively | ||
| Magnesium content reduction in grain by 28.57% in both salt concentration | ||
| Pinto bean (cv. Talash) | 8 and 12 dS m−1 | Reduction in grain yield by 26 and 41% |
| Heavy Metals | Food Legumes | Level of Metals in Soil or Growth Media | Effects | References |
|---|---|---|---|---|
| Cadmium (Cd) | Groundnut | - | Xerophytic anatomical features and reduction in grain quality | [1,9,10] |
| Common bean | 5 μg mL−1 | Changes in lipid composition and alteration in the structural component of thylakoid membrane | [117,118] | |
| Pea | 50 μM CdCl2 | Chloroplast damage, reduction in grain filling rate | [119] | |
| 2.5 mM | Decrease in starch content of seeds | |||
| Chickpea | 23 mg kg−1 | Decrease in grain protein by 22% | [1,9,120] | |
| Green gram | 24 mg kg−1 | Grain protein reduction by 8% | [1,9] | |
| Soybean | 0.1, 0.5 and 1.0 mM | Reduction in grain oil by 23, 28 and 33%, respectively | [1,9,97,117] | |
| Pigeon pea | 56 and 112 mg L−1 | Reduced photosynthesis up to 50% | [1] | |
| Lead (Pb) | Grass pea | 25, 50, 100, 200 and 300 ppm | Chromosomal abnormalities | [1,120] |
| Chickpea | 195 and 390 mg kg−1 | Grain proteins increase by 3 and 6%, respectively | [1,9] | |
| Soybean | - | Inhibited growth | [98,117] | |
| Chromium (Cr) | Common bean | 500 ppm | Reduced seed germination up to 48% | [117] |
| Chickpea | 67.5 and 135 mg kg−1 | Grain protein increased by 3% and decreased by 2%, respectively | [1,9,120] | |
| Green gram | 68 and 136 mg kg−1 | Increase in grain protein by 7 and 11%, respectively | [1,9] | |
| Mercury (Hg) | Black gram | 20 ppm | Reduced 50% seed germination potential, contamination in the entire food chain | [1] |
| Mung bean | [117,120] | |||
| Pea | [1] | |||
| Lentil | [1,120] | |||
| Soybean | 0.1, 0.5 and 1.0 mM | Grain oil reduction by 38, 58 and 68%, respectively | [1,97,117] | |
| Copper (Cu) | Pea | (50 and 75 µM) | Changes in the ultra-structure of chloroplasts, swelling of starch grains in the stroma | [1,117,120] |
| Chickpea | 66.9 and 143.8 mg kg−1 | Reduced grain protein of 9 and 18%, respectively | [1,116] | |
| Cowpea | 5 ppm | Adversely affected the germination process | ||
| Green gram | 334.5 and 669 mg kg−1 | Grain protein reduced by 4 and 5%, respectively | [1] | |
| Nickel (Ni) | Chickpea | 50, 100, 200 and 400 ppm | Reduction in seed germination and seedling growth | [1,116] |
| 290.1 and 580.2 mg kg−1 | Reduced grain protein by 2 and 16%, respectively | |||
| Cowpea | 5 ppm | adversely influenced the germination process | [1] | |
| Pigeon pea | 1.0 mM | 32% reduction in net photosynthesis, decrease enzyme activity | [1,10] | |
| Zinc (Zn) | Cowpea | 5 ppm | Adversely influenced the germination process | [1] |
| Chickpea | 4890 and 9780 mg kg−1 | Increased grain protein by 10 and 19%, respectively | [1,116,120] | |
| Arsenic (As) | Peas | 12.5–73.3 mg of sodium arsenate kg−1 | Caused interference in mineral nutrient balance | [1,114,116] |
| Chickpea | 5 mg kg−1 | Considerable inhibition in seed reserves accumulation such as starch, proteins, sugars and minerals, reduced the quality of seeds | [1,114,121] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarkar, S.; Khatun, M.; Era, F.M.; Islam, A.K.M.M.; Anwar, M.P.; Danish, S.; Datta, R.; Islam, A.K.M.A. Abiotic Stresses: Alteration of Composition and Grain Quality in Food Legumes. Agronomy 2021, 11, 2238. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11112238
Sarkar S, Khatun M, Era FM, Islam AKMM, Anwar MP, Danish S, Datta R, Islam AKMA. Abiotic Stresses: Alteration of Composition and Grain Quality in Food Legumes. Agronomy. 2021; 11(11):2238. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11112238
Chicago/Turabian StyleSarkar, Sumi, Marium Khatun, Farzana Mustafa Era, A. K. M. Mominul Islam, Md. Parvez Anwar, Subhan Danish, Rahul Datta, and A. K. M. Aminul Islam. 2021. "Abiotic Stresses: Alteration of Composition and Grain Quality in Food Legumes" Agronomy 11, no. 11: 2238. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11112238