1. Introduction
Domesticated carrot (
Daucus carota L. ssp.
sativus) can be separated into two genetically distinct groups: the Eastern (Asian) and Western (European and American) carrots. The Eastern carrot (
D. carota L. ssp.
sativus var.
atroburens), whose color ranges from yellow to dark purple (black carrot), accumulates anthocyanins as major pigments, whereas Western carrot varieties appear white to orange due to the accumulation of carotenoids [
1,
2]. Molecular approaches have located Central Asia as the origin for the carrot, with a rapid domestication process that spread carrots into North Africa, Anatolia, Asia and later into Europe by the 14th century [
1,
3].
Carrot production, mostly based on orange cultivars, has quadrupled during the last 45 years, reaching over 40 million tonnes worldwide, which makes carrot one of the 10 most economically important vegetable crops and the main source of pro-vitamin A worldwide [
4]. The main increase in production has been recorded in Asia, which implies that more carrot production is now cultivated in drier and warmer climates than in the past [
4]. Therefore, crop improvement should focus efforts in developing cultivars with improved abiotic stress resistance. Commercial cultivars are, overall, more productive than landraces. However, landraces are becoming significant sources of genetic variability in the seeking for genes for tolerance to abiotic and biotic stress factors [
5].
The extraordinary antioxidant activity of black carrot, four times higher on average than that of orange carrot [
5], results from its very high anthocyanin concentration in the taproot. Easter carrot germplasm displays a wide genetic diversity in terms of anthocyanin content and distribution along different taproot tissues. In this sense, several authors have analyzed anthocyanin pigments in over 30 carrot accessions (accs.) including lines from genbanks, and open-pollinated and hybrid commercial cultivars, reporting a concentration range of 0.5–250 mg/100 g FW [
6,
7,
8,
9]. In this sense, total monomeric anthocyanin concentration measured by the pH differential method has been frequently utilized, since it provides a robust spectrophotometric method applicable to different anthocyanins [
6,
8,
10]. The interest of the food industry in natural colorants replacing synthetic dyes has increased enormously over the last years, due to both rigorous legal restrictions and consumer concerns [
11,
12]. Black carrot anthocyanins are an excellent source of natural colorants (labelled E163 in Europe) due to their physiochemical properties (high pH, light, and heat stability), but also for their potential health benefits such as strong dietary antioxidants [
6,
13]. In addition to anthocyanins, black carrot taproot is a rich source of non-anthocyanin phenolic compounds, such as flavonoids [
14], which in turn correlates with the strong antioxidant activity of the taproot extract. Moreover, the onset and accumulation kinetic of anthocyanins and non-anthocyanin phenolic compounds seems to resemble in black carrot taproot, both type of compounds reaching their maximum levels at plant maturity [
8]. As non-enzymatic plant antioxidants, phenolic compounds function as scavengers of reactive oxygen species (ROS), participating in the response to environmental stress conditions [
15]. In this sense, the accumulation of phenolic compounds increases under a variety of both abiotic and biotic stresses such as heat stress, pathogen attack and UV radiation, among many others [
16,
17].
Carrot is an outcrossing insect-pollinated crop typically bred for open-pollinated cultivar production until cytoplasmic male sterility was discovered in the 1940s. From that moment on, cultivar development shifted to hybrids, which today represents the majority of large-scale production [
18]. Nowadays, Western carrots appear better adapted for commercial production and processing. Eastern carrots often tend to flower early, but may be better adapted to warmer (above 30 °C) and drier climates. In order to improve carrot cultivation traits in the context of a changing environment, the gene pool from open-pollinated Eastern carrot varieties should be incorporated into breeding programs. In the present work, eleven Eastern carrot accs. Originated from India and Middle Eastern countries, and a black carrot commercial cultivar were cultivated under a sub-optimal temperature range (up to 33 °C) and characterized in terms of germination indices, plant performance parameters, mineral nutrient contents and anthocyanin and flavonoid contents of the taproot, measured spectrophotometrically. The results of this work may contribute to expand knowledge on the genetic variation of cultivated carrot and opens up prospects for further evaluation.
3. Results and Discussion
In the present study, 11 Eastern carrot accs. from different origins were cultivated and evaluated under glasshouse conditions. The geographic origin covered an area between 10 and 40 degrees of latitude, from India to Turkey (
Table 1). The commercial carrot cultivar ‘Night Bird’ F1 was used as a reference. Characterization comprised seed germination indices and leaf and taproot phenotypic traits of interest for ulterior breeding programs such as size, taproot shape, bolting tendency, leaf number, leaf area and those related to taproot antioxidant capacity and use as source of colorants (anthocyanin and flavonoid contents). Firstly, germination indices were determined in in vitro conditions (
Table 2). Germination percentage varied 61–96%; this is in the range of that observed for diverse carrot germplasm [
24], in which, overall, commercial hybrids displayed higher germination percentages than landraces and wild accessions. Moreover, seedling vigor varied 4.40–11.32, whereas speed of emergence varied between 1.52 and 3.45 (
Table 2). In this sense, accs. 2, 3 and 5 showed values above those recorded for the reference cultivar for the three indices measured (
Table 2).
Twelve-week old carrot plants showed a variable overall appearance in function of top size (
Figure 2a). On the other hand, overall, the taproot appearance consisted of the typical long conical shape with a pointy end, the most frequent shape in both Eastern and Western cultivars [
25,
26,
27], the taproot shoulder of accs. 2 and 8 being clearly wider than that of the other accs. (
Figure 2b). Coloration pattern of periderm and cross taproot sections was highly variable among accs. (
Figure 2b,c). Purple coloration was observed for the majority of accs. except for n. 2 and 9, ranging from a purple periderm and non-purple xylem and phloem (for acc. 3, 4, 8 and 10) to a predominant coloration of phloem (for acc. 5 and 7) and a solid purple coloration (for acc. 1 and 12). At the genetic level, the pathway for anthocyanin biosynthesis showed a high overlap among carrot varieties. However, different variants of anthocyanin-related genes result in tissue-specific accumulations of anthocyanins [
28], which in turn is manifested in very distinct coloration patterns. Moreover, two MYB transcription factors, DcMYB6 and 7 have been proven to be regulators of anthocyanin pigmentation in purple versus non purple carrot roots [
29,
30], as well as regulators of anthocyanin glycosylation and acylation [
30]. In addition, intra-population genetic diversity of Eastern carrot accs. has been found to be higher than that of the Western carrot, whereas higher allelic richness and variability of landraces has also been observed with respect to F1 hybrids [
31], which would explain the color variability within specimens of a same acc. found in this study, especially for accs. 6 and 10. In turn, this suggests that the diversity present in carrot landraces could potentially support carrot breeding efforts in terms of coloration.
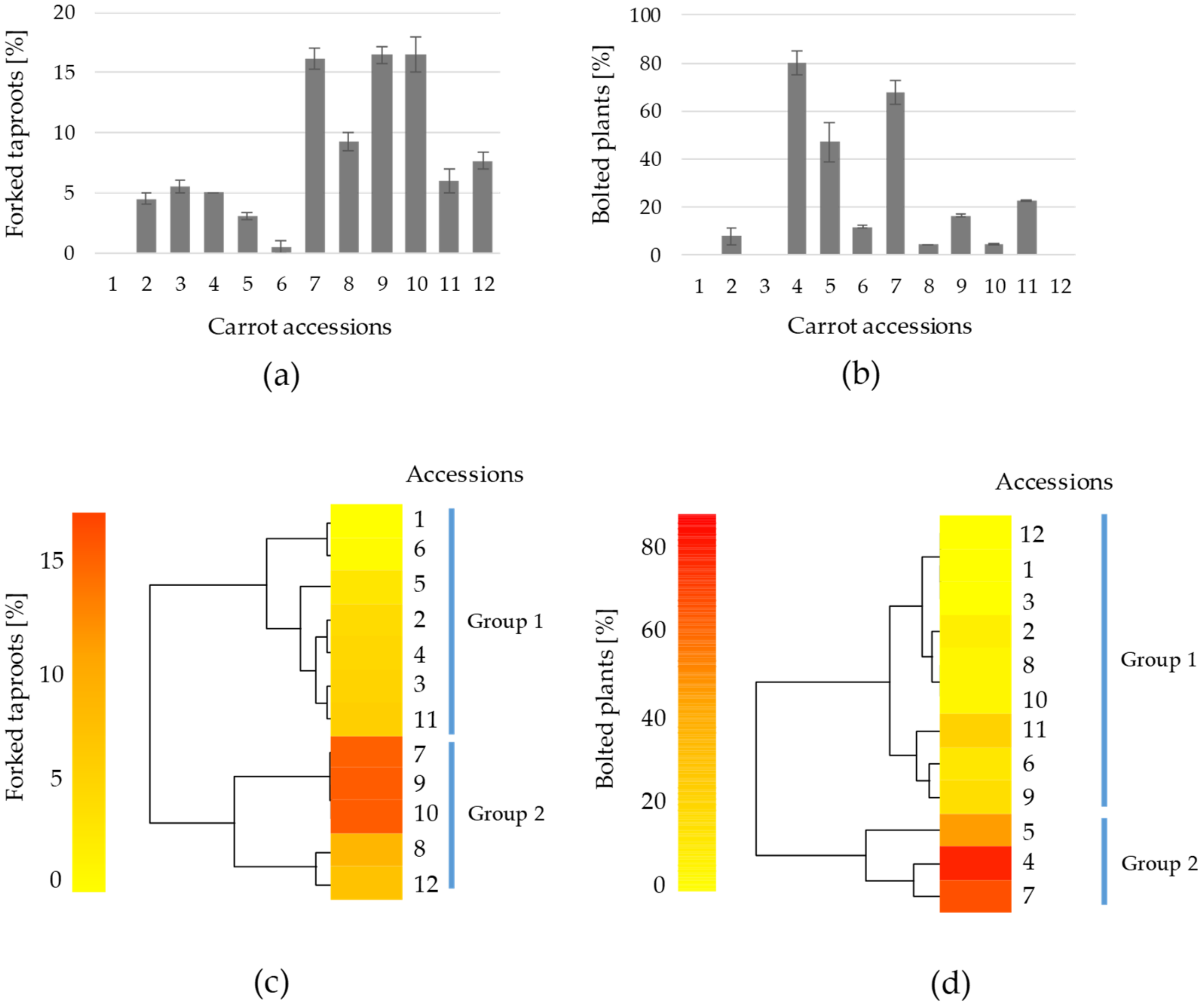
Deformed taproot shape and premature bolting are important constrains for carrot cultivation, marketability and carrot breeding [
18]. In this work, concerning taproot shape abnormalities, excluding the commercial cultivar as it did not display any forking, the proportion of forked taproots ranged from 3 to 18%, the accs. 7, 9 and 10 showing the highest values (
Figure 3a). Again, higher genetic diversity within Eastern carrot accs. may explain the variability on taproot forking found in this study [
31], which can rely on differences on optimal plant spacing and nutrient requirements among accs. [
32]. As a cool-season vegetable, carrot is normally classified as a biennial species, requiring vernalisation for flowering induction. However, cultivars and landraces adapted to warmer climates—as is the case of the accs. of this study—may need less vernalisation time, and therefore may behave as early flowering or annual [
18]. In this work, all accs. except n. 1 (reference cultivar), 3 and 12 showed a certain percentage of bolting specimens (
Figure 3b). Accs. 4, 5 and 7 were found to bolt severely, having 80, 47 and 68%, respectively, of bolting plants (
Figure 3b). Since premature bolting is an undesirable trait, bolting plants were discarded and not considered for further measurements and analysis. This lead to the reduction of available specimens for accs. 4 and 7 to 16 and 24, respectively, whereas for the rest of accs. the number of plants used for further measurements ranged from 40 to 67. Complementarily, a heatmap dendogram was elaborated to visualize the preponderance of both traits among the accessions (
Figure 3c,d); herein, based on similarities, two main groups were distinguished: group 1 for low and group 2 for high forking (
Figure 3c) and bolting (
Figure 3d).
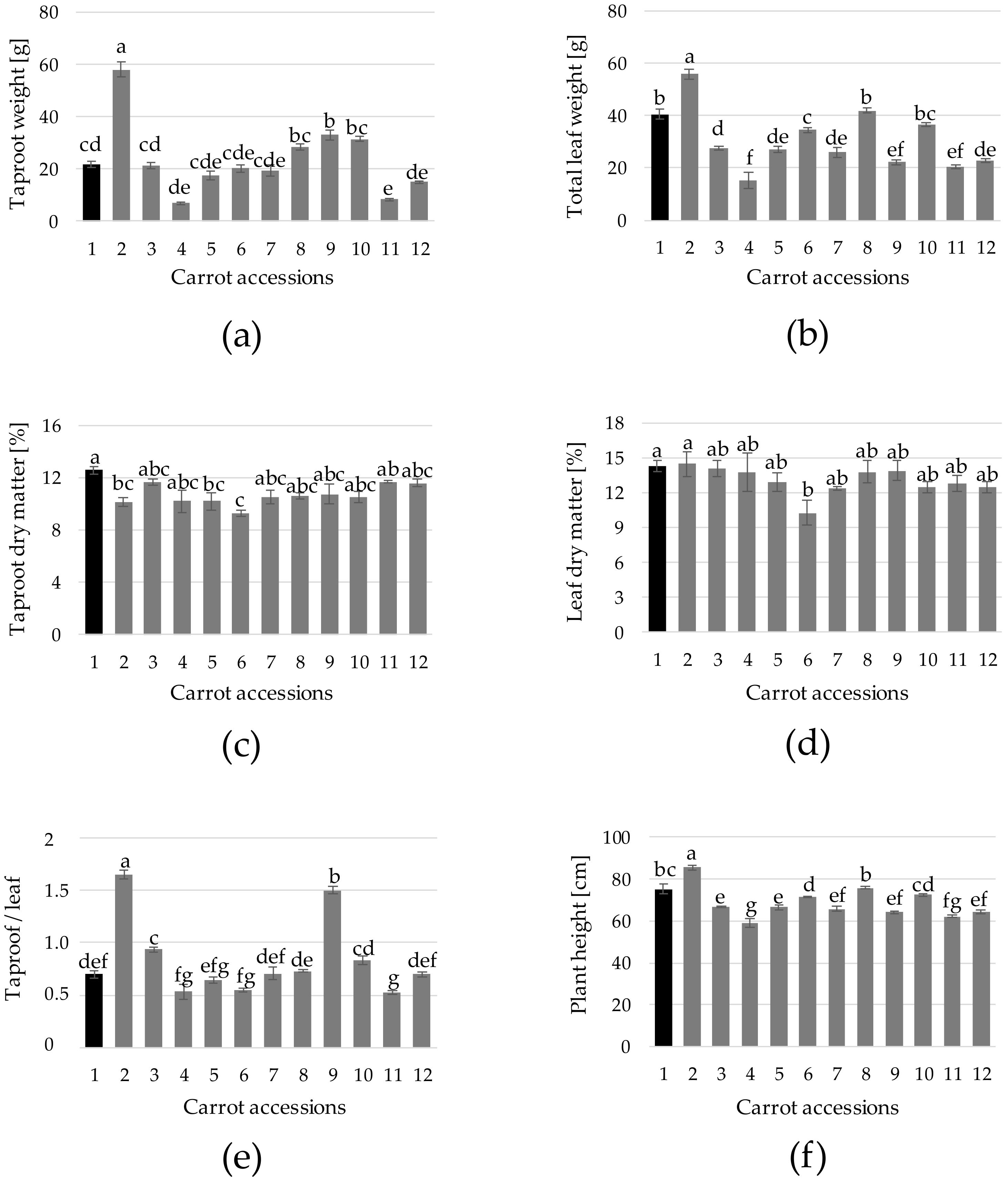
Taproot weight is a key factor determining productivity [
18]. In the present study, the weight of the reference cultivar (21.9 g) was considerably lower than that of black carrot grown in field experiments [
6,
8], although similar to that obtained in glasshouse trials under comparable environmental and fertirrigation conditions [
9] (
Figure 4a). Moreover, temperatures registered during the growing period reached 33 °C and averaged 24.4 °C, whereas ca. 15% of the time, temperature was above 30 °C. These are beyond the suitable temperatures for conventional carrot growth [
33,
34], which may partially explain the limited taproot growth observed. Compared with the reference cultivar, taproot weight was statistically higher in two accs. (n. 2 and 9) and lower in a single acc. (n. 11), whereas no statistical differences were observed in the rest of accs. (
Figure 4a). Remarkably, the weight of acc. 2 was over 160% superior than that of the reference (
Figure 4a). Total leaf weight was, to some extent, correlated with taproot weight, although in this case only acc. 2 displayed higher weight values (38%) than the reference cultivar (
Figure 4b). Taproot dry matter varied from 9.3% (acc. 6) to 12.6% (reference cultivar) (
Figure 4c), which was in the range of the values observed in black carrot [
8] and commercial orange carrot cultivars [
35]. On the other hand, leaf dry matter levels were higher than those of taproot dry matter, ranging from 10.3% (acc. 6) to 14.5% (acc. 2) (
Figure 4d). The root/leaf ratio, measured as the ratio of the dry weight of the root to the one of the top, represents a key plant-adaptive mechanism that reflects biomass allocation [
36]. In this study, taproot/leaf ratio varied notably among samples, accs. 2, 3 and 9 showing values higher than that of the reference cultivar. The values registered are in the range of those observed by others authors [
37] for Western carrots for the same growth period (12 weeks). The difference among accs. may be tightly linked to the genetic background, although it has also been found that plants accumulate more root biomass under more stressful, low-nutrient and poor climatic conditions [
36]. Concerning plant height, acc. 6 displayed the highest mean value (85.6 cm), followed by the reference cultivar (75.3) and accs. 8 (75.8 cm) and 10 (72.5), whereas the rest of accs. displayed values significantly lower than that of the reference cultivar (
Figure 4f).
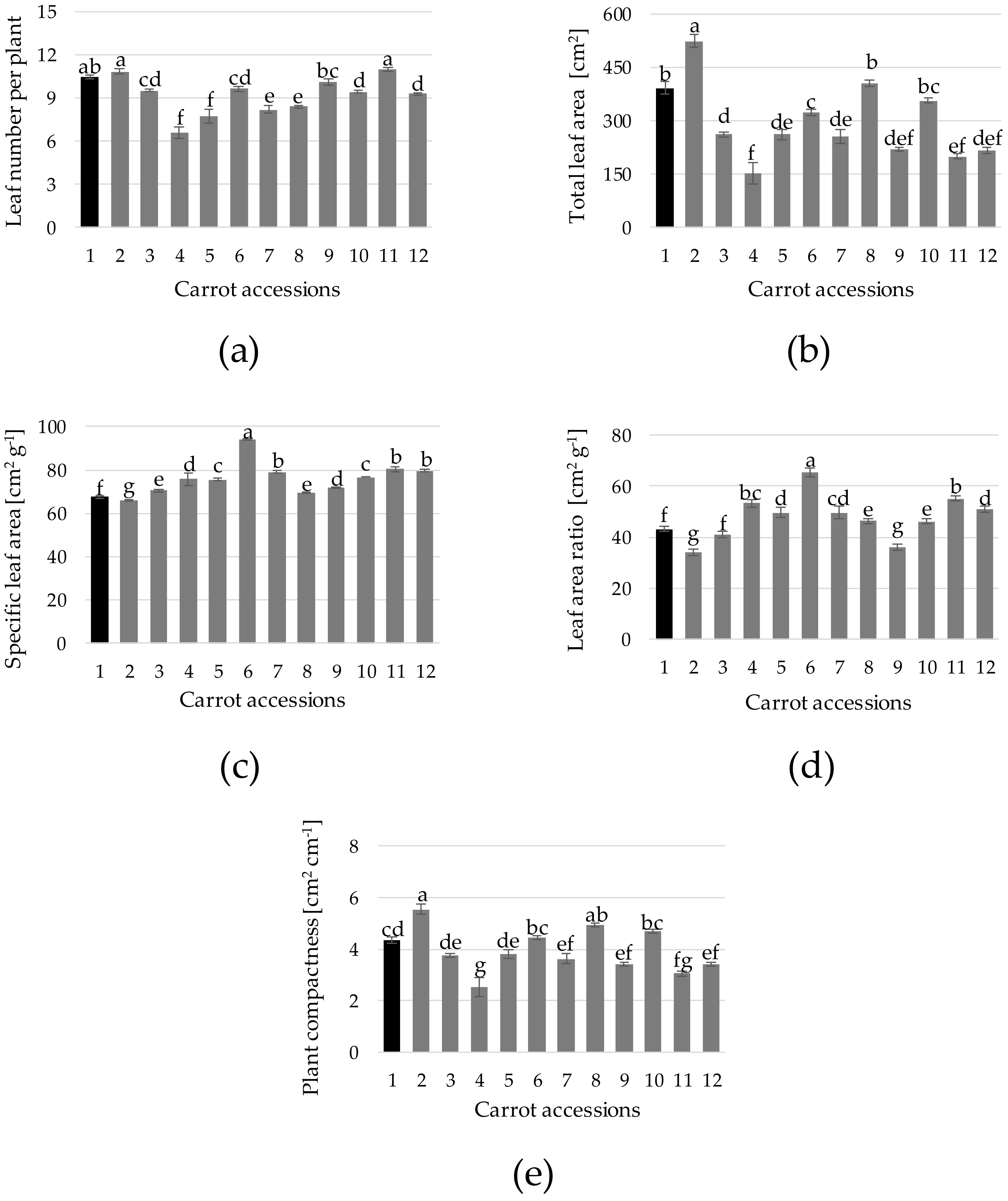
Leaf number and area, and leaf area-related indices have been reported as some of the traits best indicating potential crop yield [
38]. The count of leaf number (6.6 to 11.0 leaves per plant) and total leaf area (154 to 524 cm
2 per plant) provided values in the range of that found in field-grown carrot [
39,
40] (
Figure 5a,b). Specific leaf area (SLA) is defined as the ratio between total leaf area and total leaf dry weight—in other words, the amount of leaf area needed for each unit of biomass produced [
41,
42]. Carrot SLA varied from 94 cm
2 g
−1 (acc. 6) to 66 cm
2 g
−1 (acc. 2) and 68 cm
2 g
−1 (reference cultivar) (
Figure 5c). This may indicate a higher efficiency of accs. 1 and 2 in producing biomass. On the other hand, leaf area ratio (LAR), defined as the ratio of leaf area and total plant weigh showed a great variation (
Figure 5d); the accs. with the lowest levels were n. 2, 9, 3 and 1, providing values of 34, 36, 41 and 43 cm
2 g
−1, respectively. These results highlight accs. 1 and 2 as the most efficient in producing leaf biomass, while accs. 2 and 9 would produce plant biomass most efficiently (
Figure 5d). The variation among accs. of values for plant compactness (
Figure 5e), defined as the ratio between the total leaf area and the plant height, resembles the values for total leaf area. Moreover, overall, values for plant compactness, SLA, total leaf area and total leaf weight showed some correlation, which indicates that weight and length differences can be associated with leaf area as the main variable determining productivity [
38]. In this sense, a large leaf area may provide better utilization of diminishing growth resources [
37].
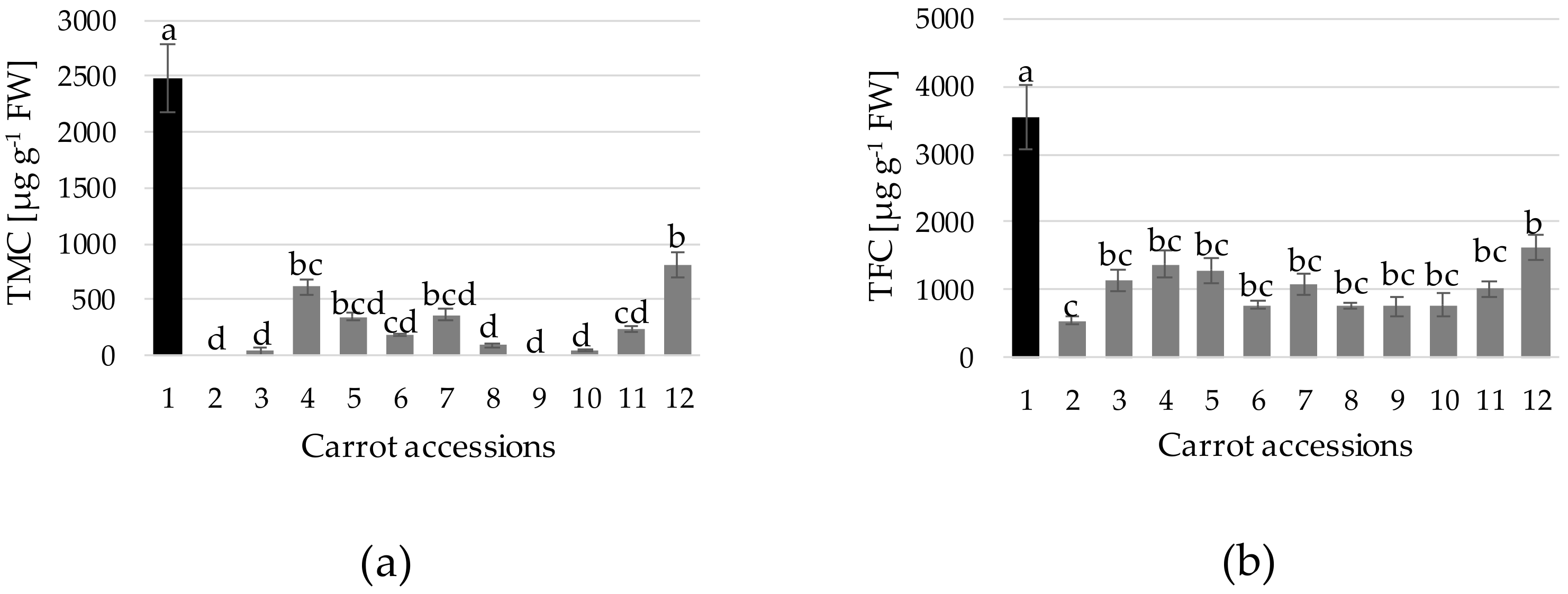
Anthocyanins and flavonoids are important components of the secondary metabolism of Eastern carrot accs., found to be highly correlated with antioxidant capacity [
43]. In this sense, high ROS-scavenging capacity is a common trait of different flavonoids, attributed to the high reactivity of their hydroxyl groups to ROS [
44]. Eastern carrot germplasm displays a wide genetic diversity in terms of anthocyanin content and distribution along different taproot tissues [
6,
7,
8,
9]. Anthocyanin content is directly associated with the color intensity and the extent of taproot tissue covered with purple; in this sense, in the majority of purple genetic accs., anthocyanins are mainly located in the outer root epidermal layer [
45]. In this work, from the 12 accs. evaluated, the reference cultivar showed the highest TMC (2482 µg g
−1 FW), at values comparable to those reported for commercial varieties [
6,
7,
8,
9], followed at some distance by acc. 12. (807 µg g
−1 FW) and then by n. 4, 7, 5, 11, 6, 8, 10 and 3, whereas levels were undetected for accs. 2 and 9 (
Figure 6a). This is associated with the coloration observed in the taproot sections (
Figure 2). Concerning TFC, a certain correlation with TMC was observed (
Figure 6b). However, from the difference between TFC and TMC, it was estimated a higher proportion of non-anthocyanin flavonoids for the accs. displaying lower TMC. Black carrot genotypes display a high proportion of mono-acylated anthocyanins, which confers physio-chemical stability to food color products over their shelf-life [
28]. Nevertheless, a signification variation on the ratio of acylated to non-acylated anthocyanins can be found in the carrot germplasm, which is also modulated in function of the environmental conditions [
28]. Therefore, further characterization on the accs. of interest in field experiments will be needed.
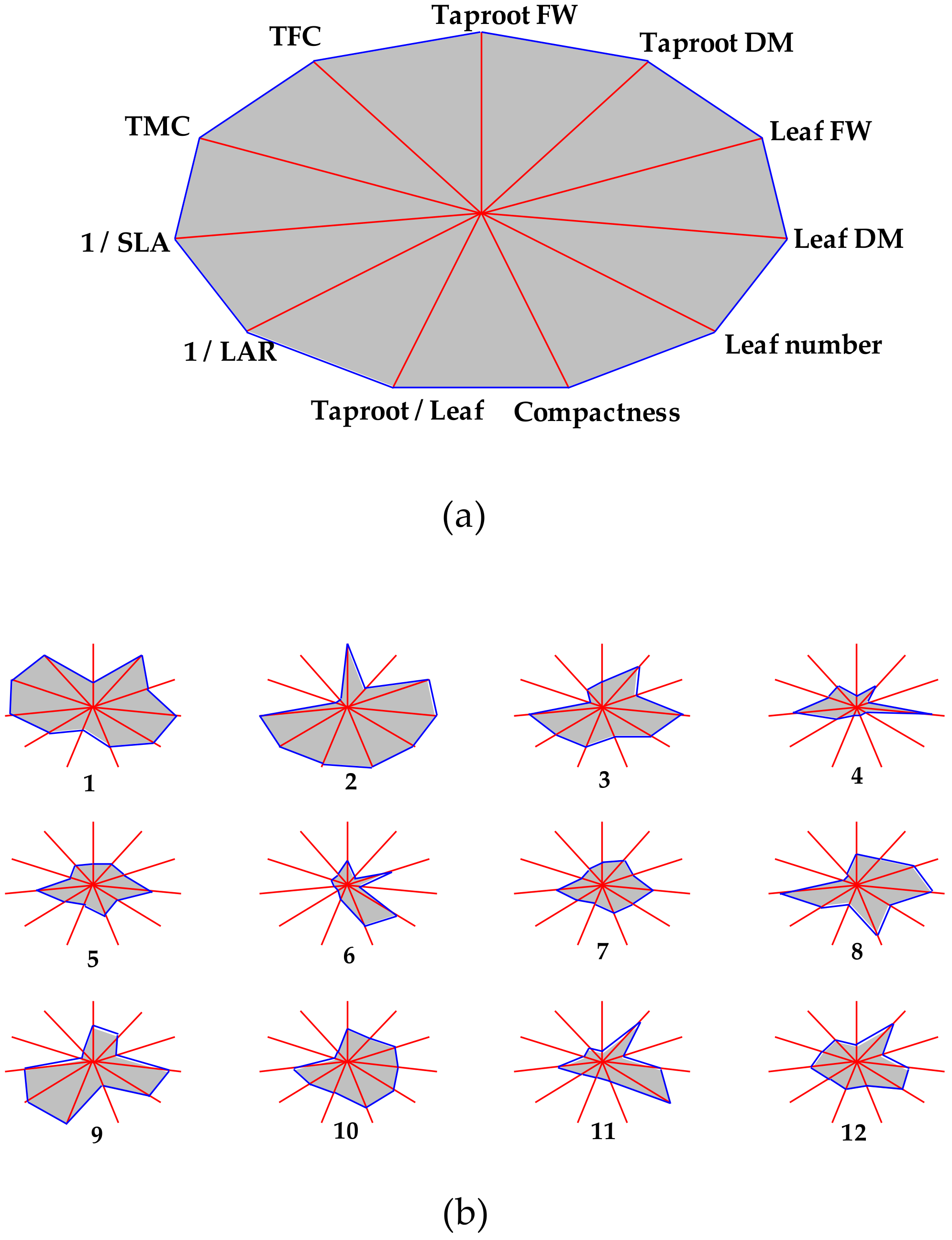
In order to provide a clearer overview of the performance of each acc., a stars and rays graph was elaborated using the main agronomical traits and indices measured so far (
Figure 7). In summary, the further the intersection from the polygon center between each axis (trait) and the polygon perimeter, the higher the trait magnitude, which helps visualize the differences among accs. For example, acc. 2 is evidenced as the one fulfilling most of the agronomic requirements, except for TMC and TFC, of which magnitudes are low. On the contrary, the reference cultivar displays high magnitudes for TMC and TFC and intermediate/high values for the rest of traits. On the other hand, accs. such as 4–7 show small magnitudes for most of the traits (
Figure 7b), which point the minor potential of these accs. for breeding purposes.
Mineral composition of carrot taproot has been evaluated for Eastern cultivars [
46,
47], whereas for purple cultivars the literature is scarce [
48]. Among the many minerals of the taproot, relatively high amounts of K, Mg, Ca, Na and Fe have been reported, with potassium as the most abundant one [
46,
47]. On the other hand, the content of Fe, Na and Mg was highly dependent on the carrot variety [
47]. In this study, among the macronutrients analyzed, for all the varieties studied K was the most abundant element in the taproot followed by Na and Ca (
Table 3). This is in agreement with previous literature [
46,
47]. Carrot leaves are rich in several minerals such as Na, P, K, Ca, Mg, Mn, Zn and Fe [
49]. Herein, the nutrient composition of black carrot leaves was reported for the first time; interestingly, K, Ca and Mg were much abundant in leaf than in taproot tissue (
Table 3), highlighting black carrot leaves as a potential nutrient source. Na was the element of which levels varied the most among accs. for both taproot and leaf tissues; this would be in line with previous studies, where Na content in carrot taproot was highly dependent on the fertilization and growing practices [
50,
51]. Micronutrients are essential to the cell function and as such are extensively involved in primary and secondary metabolisms. Alterations in optimal micronutrients concentrations may, therefore, directly or indirectly impair plant metabolism and increase susceptibility towards environmental stresses [
52]. In this study, as for the macronutrients, some variation on the levels of the different micronutrients has been found among accs. (
Table 4 and
Table 5). Remarkably, acc. 8 displayed statistically higher levels than the reference cultivar for most of analyzed micronutrients, whereas acc. 11 showed an opposite trend (
Table 4 and
Table 5). Whether this variation among accs. may imply an improved stress tolerance or not, it is matter requiring further research. Overall, the yield potential of each cultivar may influence nutrient demand, as reported [
35], and therefore may be reflected in mineral concentrations of both taproot and leaf tissues. In this sense, broader trials should be conducted to know fertilization needs accurately.
A Pearson’s correlation matrix was elaborated in order to associate variations in macronutrient contents, DM, TMC and TFC at the taproot level (
Table 6). For each variable, data for all accs. were grouped and treated jointly. As a result, except for the interaction P/S, the rest of macronutrient pairs provided significant and positive correlation coefficients, which highlights the shared underlying physiological mechanisms implied in nutrient uptake and the potential linked mechanisms in maintaining nutrient stoichiometry [
53]. Since the increase in accumulation of all these five macronutrients may precede DM accumulation, a correlation between these variables can be of interest as predictors in growth models [
54]. Furthermore, our results showed a negative correlation between DM and Ca and Na contents. On the other hand, DM highly correlated with TMC and TFC. This may be due to a time-dependent accumulation of DM [
37] and phenolic compounds [
8] during carrot taproot growth (
Table 6).


 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}