
Root Trait Diversity in Field Grown Durum Wheat and Comparison with Seedlings

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Growth Chamber Experiment
2.3. Shovelomics Experiment
2.3.1. Experimental Conditions
2.3.2. Phenotyping
2.4. Data Analyses
3. Results
3.1. Shovelomics Experiment
3.1.1. Trait Variation
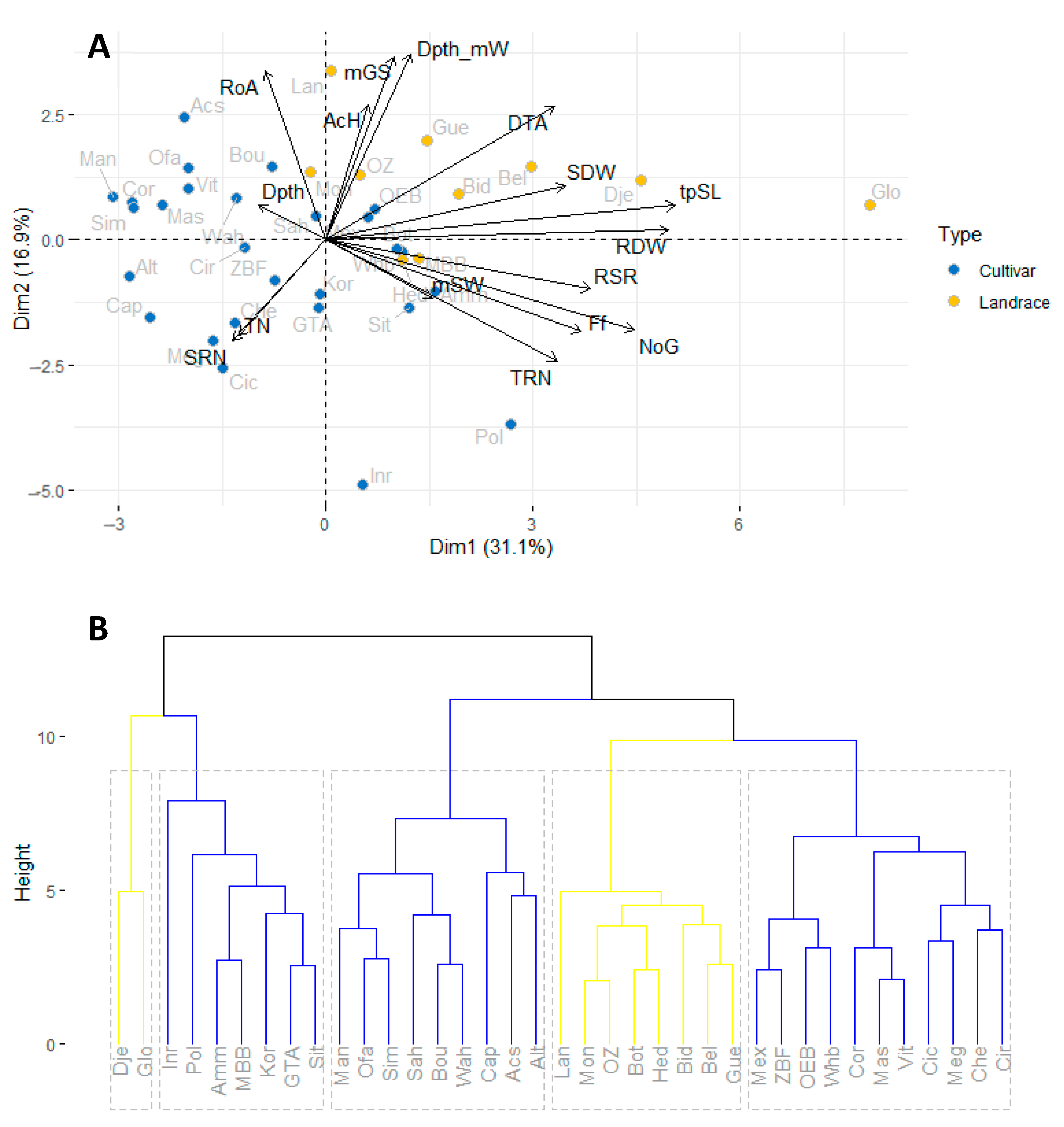
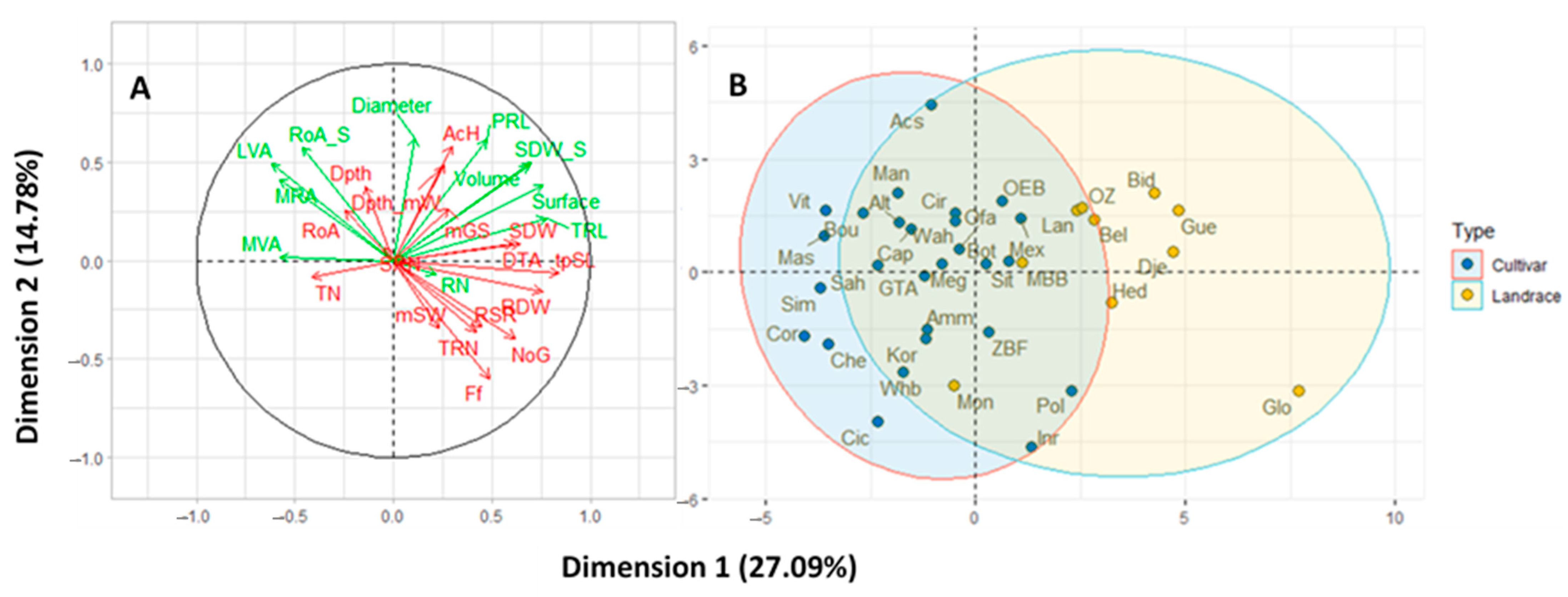
3.1.2. Relationships among Traits and Clustering of Varieties
3.2. Relationships of Traits between Experiments and Growth Stages
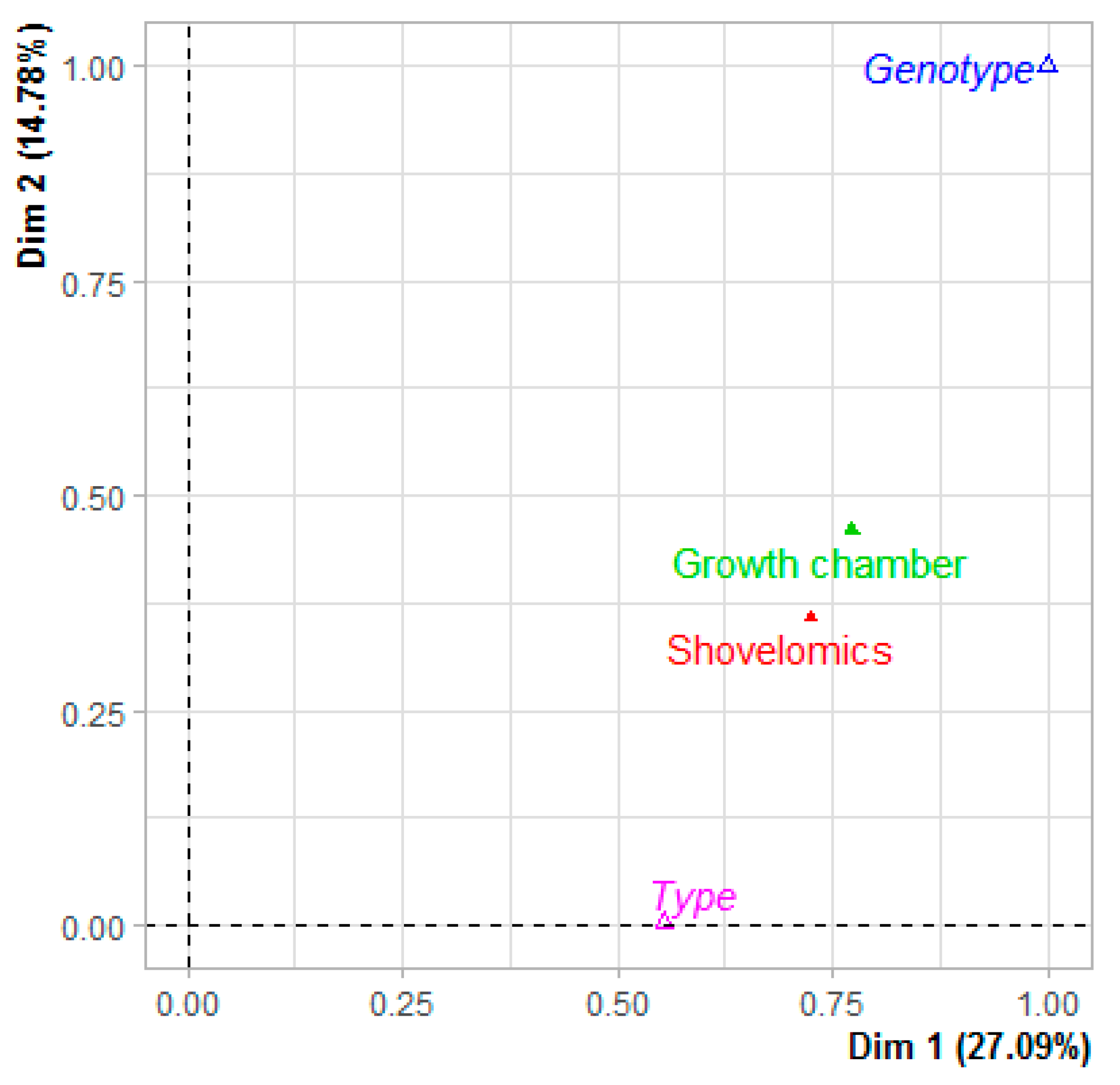
3.2.1. Multifactorial Analysis Considering the Two Systems
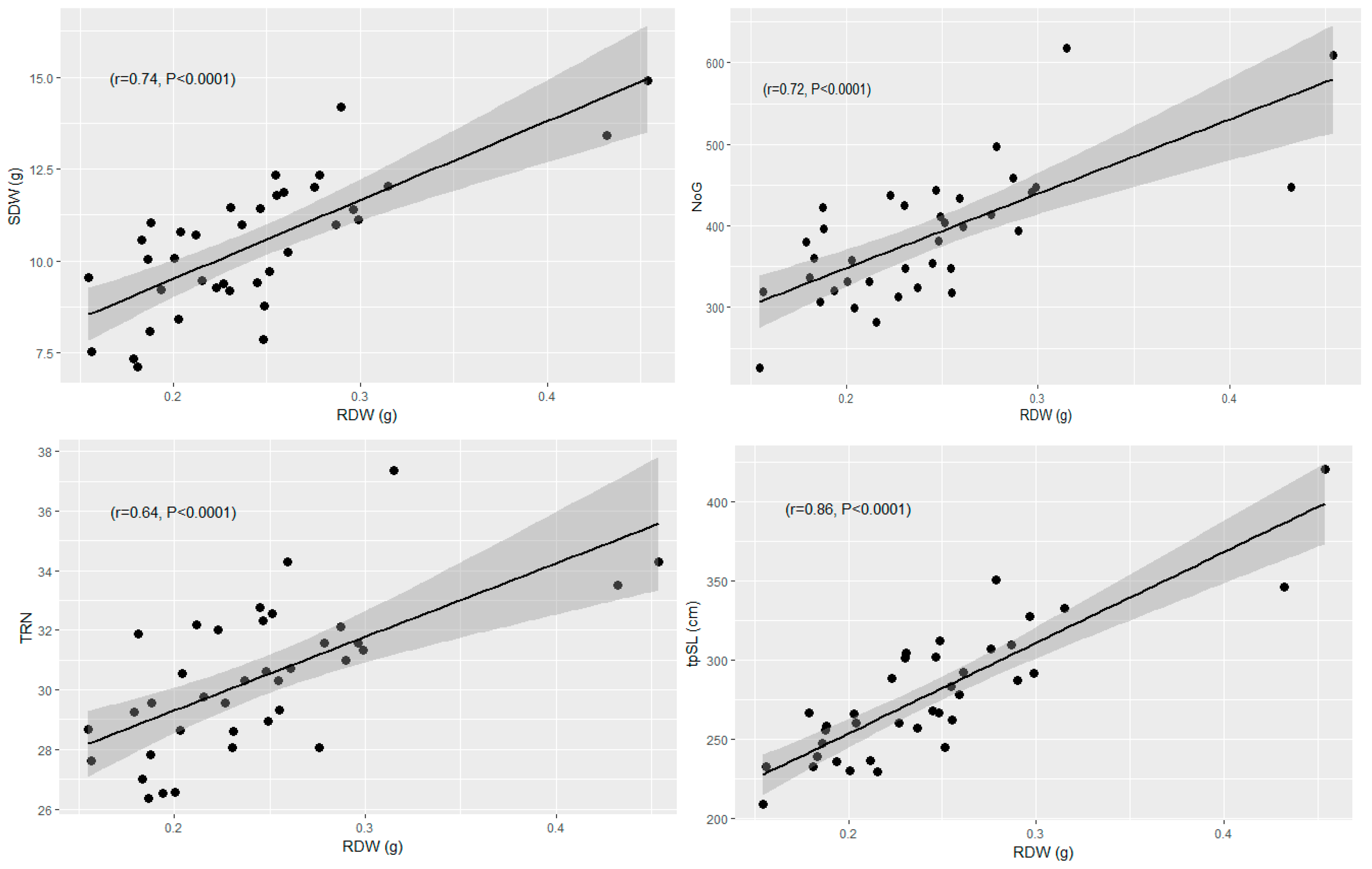
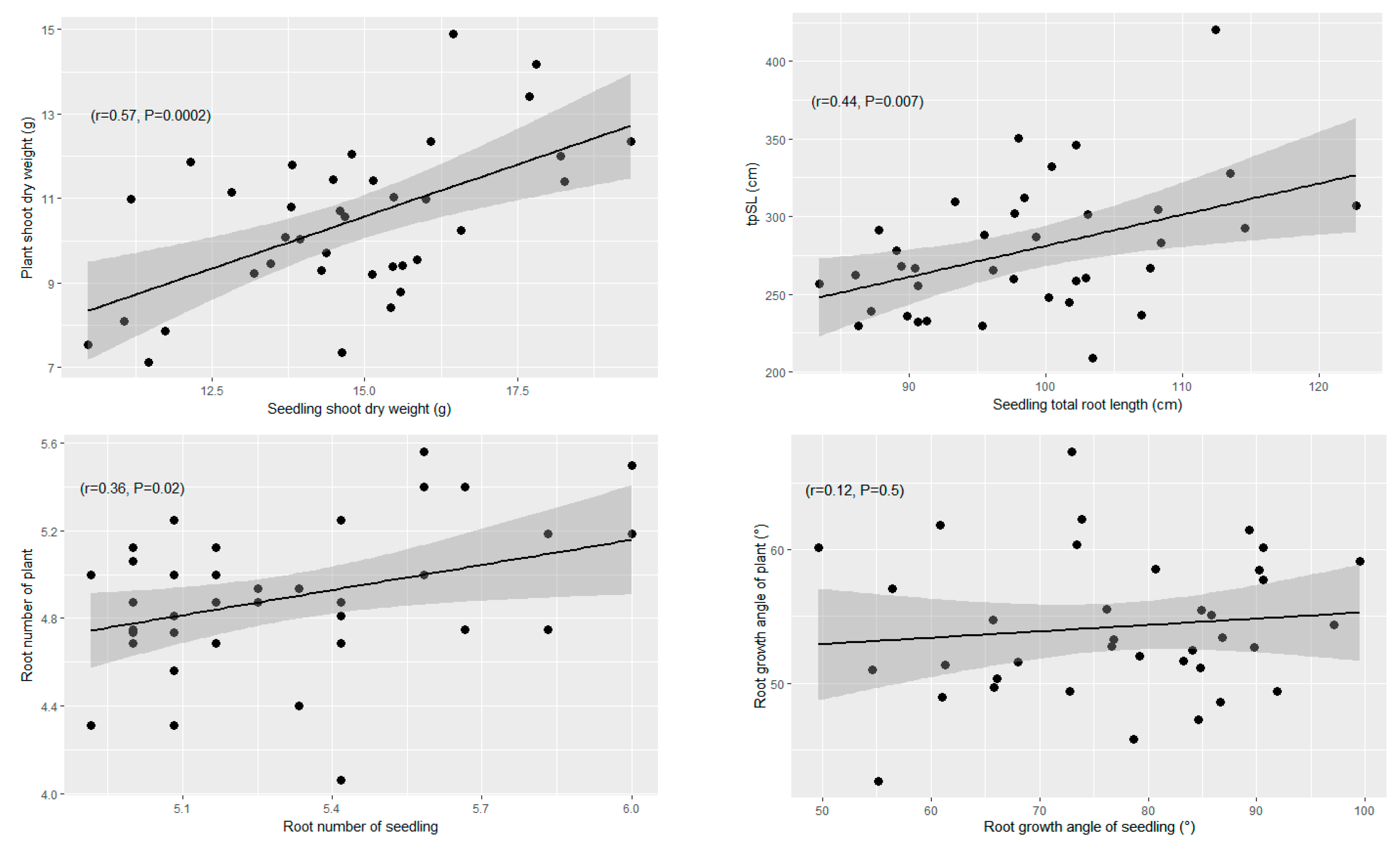
3.2.2. Correlation of Adult Traits Measured in the Field with Seedling Traits
4. Discussion
4.1. Sowing Density and Trait Variability in the Shovelomics Experiment
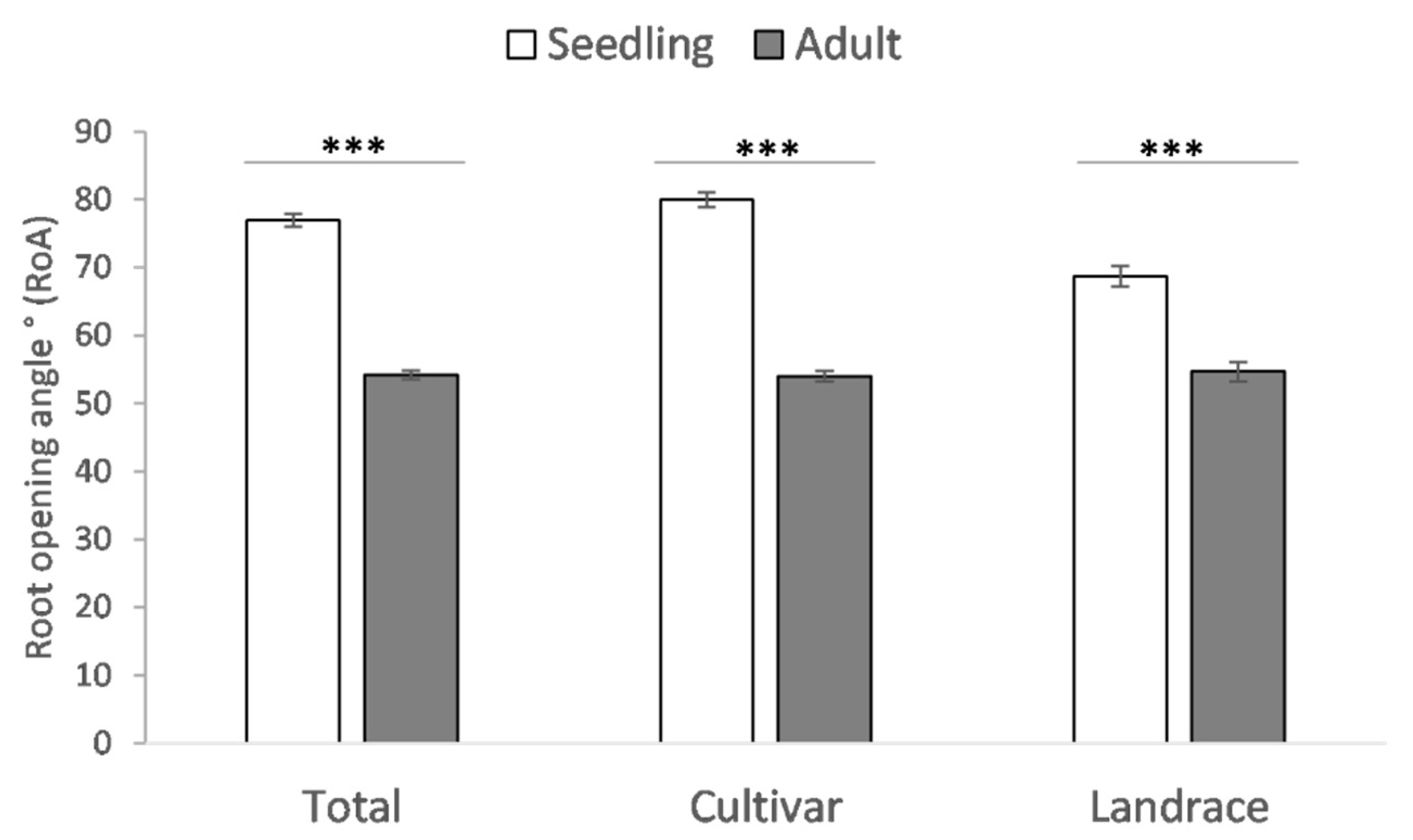
4.2. Shovelomics Revealed Different Patterns of Root Growth in Durum Wheat Cultivars
4.3. Assessment of Two Different Experimental Set-Ups to Phenotype Durum Wheat Roots
5. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Coque, M.; Gallais, A. Genomic regions involved in response to grain yield selection at high and low nitrogen fertilization in Maize. Theor. Appl. Genet. 2006, 112, 1205–1220. [Google Scholar] [CrossRef] [PubMed]
- Esau, K. Plant Anatomy, 2nd ed.; JohnWiley: New York, NY, USA, 1965; 767p. [Google Scholar]
- Kono, Y.; Yamauchi, A.; Nonoyama, T.; Tatsumi, J.; Kawamura, N. A Revised experimental system of root-soil interaction for laboratory Work. Environ. Control. Biol. 1987, 25, 141–151. [Google Scholar] [CrossRef]
- Yamauchi, A.; Kono, Y.; Tatsumi, J. Quantitave analysis on root system structures of upland rice and maize. Jpn. J. Crop Sci. 1987, 56, 608–617. [Google Scholar] [CrossRef] [Green Version]
- Lijima, M.; Kono, Y.; Yamauchi, A.; Pardales, J.R. Effects of soil compaction on the development of rice and maize root systems. Environ. Exp. Bot. 1991, 31, 333–342. [Google Scholar] [CrossRef]
- Shewry, P.R.; Hey, S.J. The contribution of wheat to human diet and health. Food Energy Secur. 2015, 4, 178–202. [Google Scholar] [CrossRef]
- Siddique, K.H.M.; Belford, R.K.; Tennant, D. Root:shoot ratios of old and modern, tall and semi-dwarf wheats in a mediterranean environment. Plant Soil 1990, 121, 89–98. [Google Scholar] [CrossRef]
- Belaid, A. Durum wheat in WANA: Production, trade, and gains from technological change. In Durum Wheat Improvement in the Mediterranean Region: New Challenges; Options Méditerranéennes: Série, A. Séminaires Méditerranéens, n. 40; Royo, C., Nachit, M., Di Fonzo, N., Araus, J.L., Eds.; CIHEAM: Zaragoza, Spain, 2000; pp. 35–49. [Google Scholar]
- Mohammadi, R.; Sadeghzadeh, D.; Armion, M.; Amri, A. Evaluation of durum wheat experimental lines under different climate and water regime conditions of Iran. Crop Pasture Sci. 2011, 62, 137–151. [Google Scholar] [CrossRef]
- Bassi, F.M.; Sanchez-Garcia, M. Adaptation and stability analysis of ICARDA durum wheat elites across 18 countries. Crop Sci. 2017, 57, 2419–2430. [Google Scholar] [CrossRef]
- Christensen, J.H.; Hewitson, B.; Busuioc, A.; Chen, A.; Gao, X.; Held, I.; Jones, R.; Kolli, R.K.; Kwon, W.T.; Laprise, R.; et al. Regional climate projections. In Climate Change 2007: The Physical Science Basis; Contributions of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 847–940. [Google Scholar]
- Bengough, A.G.; Gordon, D.C.; Al-Menaie, H.; Ellis, R.P.; Allan, D.; Keith, R.; Thomas, W.T.B.; Forster, B.P. Gel observation chamber for rapid screening of root traits in cereal seedlings. Plant Soil 2004, 262, 63–70. [Google Scholar] [CrossRef]
- Nagel, K.A.; Putz, A.; Gilmer, F.; Heinz, K.; Fischbach, A.; Pfeifer, J.; Faget, M.; Blossfeld, S.; Ernst, M.; Dimaki, C.; et al. GROWSCREEN-Rhizo is a novel phenotyping robot enabling simultaneous measurements of root and shoot growth for plants grown in soil-filled rhizotrons. Funct. Plant Biol. 2012, 39, 891. [Google Scholar] [CrossRef] [Green Version]
- Richard, C.; Hickey, L.T.; Fletcher, S.; Jennings, R.; Chenu, K.; Christopher, J.T. High-throughput phenotyping of seminal root traits in wheat. Plant Methods 2015, 11, 13. [Google Scholar] [CrossRef] [Green Version]
- York, L.M.; Slack, S.; Bennett, M.J.; Foulkes, M.J. Wheat Shovelomics I: A Field phenotyping approach for characterising the structure and function of root systems in tillering species. bioRxiv 2018. [CrossRef]
- Friedli, C.N.; Abiven, S.; Fossati, D.; Hund, A. Modern wheat semi-dwarfs root deep on demand: Response of rooting depth to drought in a set of swiss era wheats covering 100 years of breeding. Euphytica 2019, 215, 85. [Google Scholar] [CrossRef] [Green Version]
- Manschadi, A.M.; Christopher, J.; deVoil, P.; Hammer, G.L. The role of root architectural traits in adaptation of wheat to water-limited environments. Funct. Plant Biol. 2006, 33, 823–837. [Google Scholar] [CrossRef] [Green Version]
- Christopher, J.; Christopher, M.; Jennings, R.; Jones, S.; Fletcher, S.; Borrell, A.; Manschadi, A.M.; Jordan, D.; Mace, E.; Hammer, G. QTL for root angle and number in a population developed from bread wheats (Triticum Aestivum) with contrasting adaptation to water-limited environments. Theor. Appl. Genet. 2013, 126, 1563–1574. [Google Scholar] [CrossRef]
- Gregory, P.J.; Bengough, A.G.; Grinev, D.; Schmidt, S.; Thomas, W.B.T.B.; Wojciechowski, T.; Young, I.M. Root phenomics of crops: Opportunities and challenges. Funct. Plant Biol. 2009, 36, 922. [Google Scholar] [CrossRef] [PubMed]
- Asif, M.; Kamran, A. Plant breeding for water-limited environments. Crop Sci. 2011, 51, 2911–2912. [Google Scholar] [CrossRef]
- Wasson, A.P.; Richards, R.A.; Chatrath, R.; Misra, S.C.; Prasad, S.V.S.; Rebetzke, G.J.; Kirkegaard, J.A.; Christopher, J.; Watt, M. Traits and selection strategies to improve root systems and water uptake in water-limited wheat crops. J. Exp. Bot. 2012, 63, 3485–3498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borrell, A.K.; Mullet, J.E.; George-Jaeggli, B.; van Oosterom, E.J.; Hammer, G.L.; Klein, P.E.; Jordan, D.R. Drought adaptation of stay-green sorghum is associated with canopy development, leaf anatomy, root growth, and water uptake. J. Exp. Bot. 2014, 65, 6251–6263. [Google Scholar] [CrossRef]
- Nakamoto, T.; Shimoda, K.; Matsuzaki, A. Elongation angle of nodal roots and its possible relation to spatial root distribution in maize and foxtail millet. Jpn. J. Crop Sci. 1991, 60, 543–549. [Google Scholar] [CrossRef] [Green Version]
- Oyanagi, A.; Nakamoto, T.; Wada, M. Relationship between root growth angle of seedlings and vertical distribution of roots in the field in wheat cultivars. Jpn. J. Crop Sci. 1993, 62, 565–570. [Google Scholar] [CrossRef] [Green Version]
- Oyanagi, A. Gravitropic response growth angle and vertical distribution of roots of wheat (Triticum aestivum L.). Plant Soil 1994, 165, 323–326. [Google Scholar] [CrossRef]
- Acuña, T.L.B.; Wade, L.J. Genotype×environment interactions for root depth of wheat. Field Crops Res. 2012, 137, 117–125. [Google Scholar] [CrossRef]
- Christopher, J.T.; Manschadi, A.M.; Hammer, G.L.; Borrell, A.K. Developmental and physiological traits associated with high yield and stay-green phenotype in wheat. Aust. J. Agric. Res. 2008, 59, 354–364. [Google Scholar] [CrossRef]
- Hamada, A.; Nitta, M.; Nasuda, S.; Kato, K.; Fujita, M.; Matsunaka, H.; Okumoto, Y. Novel qtls for growth angle of seminal roots in wheat (Triticum aestivum L.). Plant Soil 2012, 354, 395–405. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, M.P.; Pierre, C.S.; Saad, A.S.I.; Vargas, M.; Condon, A.G. Evaluating potential genetic gains in wheat associated with stress-adaptive trait expression in elite genetic resources under drought and heat stress. Crop Sci. 2007, 47, S-172–S-189. [Google Scholar] [CrossRef]
- Alahmad, S.; El Hassouni, K.; Bassi, F.M.; Dinglasan, E.; Youssef, C.; Quarry, G.; Aksoy, A.; Mazzucotelli, E.; Juhász, A.; Able, J.A.; et al. A Major root architecture qtl responding to water limitation in durum wheat. Front. Plant Sci. 2019, 10, 436. [Google Scholar] [CrossRef] [Green Version]
- Ehdaie, B.; Waines, J.G. Genetic variation, heritability and path-analysis in landraces of bread wheat from southwestern Iran. Euphytica 1989, 41, 183–190. [Google Scholar] [CrossRef]
- Harlan, J. Crops and Man, 2nd ed.; American Society of Agronomy and Crop Science Society of America: Madison, WI, USA, 1992. [Google Scholar]
- Jaradat, A. Wheat Landraces: Genetic Resources for Sustenance and Sustainability. USDA-ARS 2011. pp. 1–20. Available online: https://www.ars.usda.gov/ARSUserFiles/50600000/products-wheat/AAJ-Wheat%20Landraces.pdf (accessed on 8 December 2021).
- Zeven, A.C. Traditional maintenance breeding of landraces: 1. Data by crop. Euphytica 2000, 116, 65–85. [Google Scholar] [CrossRef]
- Zeven, A.C. Traditional maintenance breeding of landraces: 2. Practical and theoretical considerations on maintenance of variation of landraces by farmers and gardeners. Euphytica 2002, 123, 147–158. [Google Scholar] [CrossRef]
- Jaradat, A.A. Wheat landraces: A mini review. Emir. J. Food Agric. 2013, 25, 20–29. [Google Scholar] [CrossRef]
- Ceccarelli, S.; Acevedo, E.; Grando, S. Breeding for yield stability in unpredictable environments: Single traits, interaction between traits, and architecture of genotypes. Euphytica 1991, 56, 169–185. [Google Scholar] [CrossRef]
- Blum, A. Crop responses to drought and the interpretation of adaptation. In Drought Tolerance in Higher Plants: Genetical, Physiological and Molecular Biological Analysis; Belhassen, E., Ed.; Springer: Dordrecht, The Netherlands, 1997; pp. 57–70. ISBN 978-94-017-1299-6. [Google Scholar]
- Borlaug, N.E. Sixty-two years of fighting hunger: Personal recollections. Euphytica 2007, 157, 287–297. [Google Scholar] [CrossRef]
- Waines, J.G.; Ehdaie, B. Domestication and crop physiology: Roots of green-revolution wheat. Ann. Bot. 2007, 100, 991–998. [Google Scholar] [CrossRef] [Green Version]
- Wojciechowski, T.; Gooding, M.J.; Ramsay, L.; Gregory, P.J. The effects of dwarfing genes on seedling root growth of wheat. J. Exp. Bot. 2009, 60, 2565–2573. [Google Scholar] [CrossRef]
- Elazab, A.; Serret, M.D.; Araus, J.L. Interactive effect of water and nitrogen regimes on plant growth, root traits and water status of old and modern durum wheat genotypes. Planta 2016, 244, 125–144. [Google Scholar] [CrossRef] [PubMed]
- Aziz, M.M.; Palta, J.A.; Siddique, K.H.M.; Sadras, V.O. Five decades of selection for yield reduced root length density and increased nitrogen uptake per unit root length in australian wheat varieties. Plant Soil 2017, 413, 181–192. [Google Scholar] [CrossRef]
- Hurd, E.A. Phenotype and drought tolerance in wheat. Agric. Meteorol. 1974, 14, 39–55. [Google Scholar] [CrossRef]
- Chloupek, O.; Forster, B.P.; Thomas, W.T.B. The effect of semi-dwarf genes on root system size in field-grown barley. Theor. Appl. Genet. 2006, 112, 779–786. [Google Scholar] [CrossRef]
- Lupton, F.G.H.; Oliver, R.H.; Ellis, F.B.; Barnes, B.T.; Howse, K.R.; Welbank, P.J.; Taylor, P.J. Root and shoot growth of semi-dwarf and taller winter wheats. Ann. Appl. Biol. 1974, 77, 129–144. [Google Scholar] [CrossRef]
- Cholick, F.A.; Welsh, J.R.; Cole, C.V. Rooting patterns of semi-dwarf and tall winter wheat cultivars under dryland field conditions1. Crop Sci. 1977, 17, 637–639. [Google Scholar] [CrossRef]
- González, J.M.; Friero, E.; Selfa, L.; Froilán, S.; Jouve, N. A Comparative study of root system architecture in seedlings of Brachypodium Spp. using three plant growth supports. Cereal Res. Commun. 2016, 44, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Blum, A. Plant Breeding for Water-Limited Environments; Springer: New York, NY, USA, 2011. [Google Scholar]
- Subira, J.; Ammar, K.; Álvaro, F.; García del Moral, L.F.; Dreisigacker, S.; Royo, C. Changes in durum wheat root and aerial biomass caused by the introduction of the Rht-B1b dwarfing allele and their effects on yield formation. Plant Soil 2016, 403, 291–304. [Google Scholar] [CrossRef] [Green Version]
- Grzesiak, M.T.; Hordyńska, N.; Maksymowicz, A.; Grzesiak, S.; Szechyńska-Hebda, M. Variation among spring wheat (Triticum aestivum L.) genotypes in response to the drought stress. II-Root system structure. Plants 2019, 8, 584. [Google Scholar] [CrossRef] [Green Version]
- Wasson, A.P.; Rebetzke, G.J.; Kirkegaard, J.A.; Christopher, J.; Richards, R.A.; Watt, M. Soil coring at multiple field environments can directly quantify variation in deep root traits to select wheat genotypes for breeding. J. Exp. Bot. 2014, 65, 6231–6249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trachsel, S.; Kaeppler, S.M.; Brown, K.M.; Lynch, J.P. Shovelomics: High throughput phenotyping of maize (Zea mays L.) root architecture in the field. Plant Soil 2011, 341, 75–87. [Google Scholar] [CrossRef]
- Maccaferri, M.; El-Feki, W.; Nazemi, G.; Salvi, S.; Canè, M.A.; Colalongo, M.C.; Stefanelli, S.; Tuberosa, R. Prioritizing quantitative trait loci for root system architecture in tetraploid wheat. J. Exp. Bot. 2016, 67, 1161–1178. [Google Scholar] [CrossRef]
- Colombi, T.; Kirchgessner, N.; Le Marié, C.A.; York, L.M.; Lynch, J.P.; Hund, A. Next generation shovelomics: Set up a tent and REST. Plant Soil 2015, 388, 1–20. [Google Scholar] [CrossRef]
- Boudiar, R.; González, J.M.; Mekhlouf, A.; Casas, A.M.; Igartua, E. Durum wheat seminal root traits within modern and landrace germplasm in Algeria. Agronomy 2020, 10, 713. [Google Scholar] [CrossRef]
- Lobet, G.; Pagès, L.; Draye, X. A novel image-analysis toolbox enabling quantitative analysis of root system architecture. Plant Physiol. 2011, 157, 29–39. [Google Scholar] [CrossRef] [Green Version]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A Decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH image to imagej: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Payne, R.W.; Murray, D.A.; Harding, S.A.; Baird, D.B.; Soutar, D.M. GenStat for Windows Introduction, 12th ed.; VSN International: Hemel Hempstead, UK, 2009. [Google Scholar]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Kassambara, A.; Mundt, F. Extract and Visualize the Results of Multivariate Data Analyses. Available online: https://rpkgs.datanovia.com/factoextra/index.html (accessed on 9 December 2020).
- Alvarado, G.; Rodríguez, F.M.; Pacheco, A.; Burgueño, J.; Crossa, J.; Vargas, M.; Pérez-Rodríguez, P.; Lopes-Cruz, M.A. META-R: A software to analyze data from multi-environment plant breeding trials. Crop J. 2020, 8, 745–756. [Google Scholar] [CrossRef]
- Fradgley, N.; Evans, G.; Biernaskie, J.M.; Cockram, J.; Marr, E.C.; Oliver, A.G.; Ober, E.; Jones, H. Effects of breeding history and crop management on the root architecture of wheat. Plant Soil 2020, 452, 587–600. [Google Scholar] [CrossRef] [PubMed]
- Hecht, V.L.; Temperton, V.M.; Nagel, K.A.; Rascher, U.; Pude, R.; Postma, J.A. Plant density modifies root system architecture in spring barley (Hordeum vulgare L.) through a change in nodal root number. Plant Soil 2019, 439, 179–200. [Google Scholar] [CrossRef]
- Soleymani, A.; Shahrajabian, M.H.; Naranjani, L. Determination of the suitable planting date and plant density for different cultivars of barley (Hordeum vulgare L.) in Fars. Afr. J. Plant Sci. 2011, 5, 284–286. [Google Scholar]
- Harper, J. Population Biology of Plants; Academic Press: London, UK, 1977. [Google Scholar]
- Darwinkel, A. Patterns of tillering and grain production of winter wheat at a wide range of plant densities. Neth. J. Agric. Sci. 1978, 26, 383–398. [Google Scholar] [CrossRef]
- Khalil, S.K.; Wahab, A.; Amanulla, A.; Khan, A.Z. Variation in leaf traits, yield and yield components of Faba bean in response to planting dates and densities. Egypt. Acad. J. Biol. Sci. H Bot. 2011, 2, 35–43. [Google Scholar] [CrossRef]
- Moosavi, S.; Seghatoleslami, M.; Moazeni, A. Effect of planting date and plant density on morphplogical traits, LAI and forage corn (Sc. 370) yield in second cultivation. Int. Res. J. Appl. Basic Sci. 2012, 3, 57–63. [Google Scholar]
- Olsen, J.; Weiner, J. The influence of Triticum aestivum density, sowing pattern and nitrogen fertilization on leaf area index and its spatial variation. Basic Appl. Ecol. 2007, 8, 252–257. [Google Scholar] [CrossRef]
- Hecht, V.L.; Temperton, V.M.; Nagel, K.A.; Rascher, U.; Postma, J.A. Sowing density: A neglected factor fundamentally affecting root distribution and biomass allocation of field grown spring barley (Hordeum vulgare L.). Front. Plant Sci. 2016, 7, 944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Marié, C.A.; York, L.M.; Strigens, A.; Malosetti, M.; Camp, K.-H.; Giuliani, S.; Lynch, J.P.; Hund, A. Shovelomics root traits assessed on the EURoot maize panel are highly heritable across environments but show low genotype-by-nitrogen interaction. Euphytica 2019, 215, 173. [Google Scholar] [CrossRef] [Green Version]
- Kramer, P.J. Plant and Soil Water Relationships: A Modern Synthesis; McGraw-Hill Book Company: New York, NY, USA, 1969. [Google Scholar]
- Jackson, R.B.; Sperry, J.S.; Dawson, T.E. Root water uptake and transport: Using physiological processes in global predictions. Trends Plant Sci. 2000, 5, 482–488. [Google Scholar] [CrossRef]
- El Hassouni, K.; Alahmad, S.; Belkadi, B.; Filali-Maltouf, A.; Hickey, L.T.; Bassi, F.M. Root system architecture and its association with yield under different water regimes in durum wheat. Crop Sci. 2018, 58, 2331–2346. [Google Scholar] [CrossRef] [Green Version]
- Palta, J.A.; Chen, X.; Milroy, S.P.; Rebetzke, G.J.; Dreccer, M.F.; Watt, M. Large root systems: Are they useful in adapting wheat to dry environments? Funct. Plant Biol. 2011, 38, 347–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirkegaard, J.A.; Lilley, J.M.; Howe, G.N.; Graham, J.M. Impact of subsoil water use on wheat yield. Aust. J. Agric. Res. 2007, 58, 303–315. [Google Scholar] [CrossRef]
- Passioura, J.B. Roots and drought resistance. Agric. Water Manag. 1983, 7, 265–280. [Google Scholar] [CrossRef]
- Tuberosa, R.; Sanguineti, M.C.; Landi, P.; Michela Giuliani, M.; Salvi, S.; Conti, S. Identification of QTLs for root characteristics in maize grown in hydroponics and analysis of their overlap with QTLs for grain yield in the field at two water regimes. Plant Mol. Biol. 2002, 48, 697–712. [Google Scholar] [CrossRef]
- Watt, M.; Moosavi, S.; Cunningham, S.C.; Kirkegaard, J.A.; Rebetzke, G.J.; Richards, R.A. A rapid, controlled-environment seedling root screen for wheat correlates well with rooting depths at vegetative, but not reproductive, stages at two field sites. Ann. Bot. 2013, 112, 447–455. [Google Scholar] [CrossRef] [Green Version]
- Rich, S.M.; Christopher, J.; Richards, R.; Watt, M. Root phenotypes of young wheat plants grown in controlled environments show inconsistent correlation with mature root traits in the field. J. Exp. Bot. 2020, 71, 4751–4762. [Google Scholar] [CrossRef] [PubMed]
- York, L.M.; Lynch, J.P. Intensive field phenotyping of maize (Zea mays L.) root crowns identifies phenes and phene integration associated with plant growth and nitrogen acquisition. J. Exp. Bot. 2015, 66, 5493–5505. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Mean | CV% | SD | h2 | ANOVA | ||
|---|---|---|---|---|---|---|---|
| Genotype | Type | Genotype within Type | |||||
| Field measured traits | |||||||
| DTA (days) | 122.2 | 3.5 | 4.4 | 0.93 | *** | *** | *** |
| SDW (g) | 10.15 | 31.4 | 3.1 | 0.65 | *** | *** | *** |
| TN | 2.7 | 25.9 | 0.7 | 0.44 | * | *** | ns |
| RDW (g) | 0.240 | 44.2 | 0.1 | 0.66 | *** | *** | *** |
| RSR | 0.024 | 35.2 | 0.01 | 0.53 | *** | ns | *** |
| Laboratory and image analysis traits | |||||||
| SRN | 4.9 | 17.6 | 0.8 | 0.29 | * | ns | * |
| TRN | 30.1 | 20.5 | 6.0 | 0.37 | * | ns | ns |
| RoA (°) | 54.2 | 18.0 | 9.5 | 0.23 | ns | ns | ns |
| AcH (cm2) | 93.5 | 37.9 | 35.5 | 0.31 | *** | ns | ** |
| Ff | 0.20 | 36.7 | 0.07 | 0.37 | *** | ** | *** |
| tpSL (cm) | 278.0 | 28.8 | 80.0 | 0.55 | *** | *** | * |
| Dpth (cm) | 16.49 | 20.74 | 3.42 | 0.46 | ** | ns | ** |
| Dpth_mW (cm) | 6.10 | 27.86 | 1.70 | 0.00 | ns | ns | ns |
| NoG | 388.46 | 32.89 | 127.77 | 0.51 | ** | * | ** |
| mGS (cm2) | 0.0018 | 56.4 | 0.001 | 0.27 | ns | ** | ns |
| mSW (cm) | 0.0603 | 27.3 | 0.016 | 0.00 | ns | ns | ns |
| Modern Cultivars | Landraces | |||||||
|---|---|---|---|---|---|---|---|---|
| Traits | Min | Max | Mean | CV% | Min | Max | Mean | CV% |
| Field measured traits | ||||||||
| DTA (days) | 111.0 | 130.0 | 120.3 b | 2.8 | 124.0 | 134.0 | 127.3 a | 1.6 |
| SDW (g) | 4.40 | 15.23 | 9.73 b | 20.8 | 8.53 | 21.00 | 12.28 a | 18.9 |
| TN | 1.75 | 5.00 | 2.8 a | 20.0 | 2.00 | 3.50 | 2.5 b | 16.5 |
| RDW (g) | 0.08 | 0.53 | 0.221 b | 32.6 | 0.105 | 0.599 | 0.301 a | 34.3 |
| RSR | 0.01 | 0.04 | 0.023 a | 27.2 | 0.012 | 0.040 | 0.024 a | 26.6 |
| Laboratory and image analysis traits | ||||||||
| SRN | 3.33 | 5.75 | 4.9 a | 10.6 | 3.25 | 6.00 | 4.9 a | 12.5 |
| TRN | 16.33 | 42.00 | 30.1 a | 15.0 | 22.67 | 44.25 | 31.0 a | 13.5 |
| RoA (°) | 29.19 | 88.97 | 54.0 a | 17.6 | 36.82 | 77.14 | 54.7 a | 17.5 |
| AcH (cm2) | 43.77 | 159.79 | 91.6 a | 24.9 | 56.53 | 137.02 | 98.5 a | 17.7 |
| Ff | 0.11 | 0.31 | 0.19 b | 23.9 | 0.13 | 0.34 | 0.21 a | 18.2 |
| tpSL (cm) | 120.62 | 394.39 | 263.5 b | 20.6 | 168.16 | 472.19 | 317.9 a | 21.2 |
| Dpth (cm) | 10.07 | 34.51 | 16.7 a | 22.53 | 10.97 | 20.17 | 15.92 a | 13.6 |
| Dpth_mW (cm) | 1.83 | 11.81 | 6.0 a | 27.75 | 3.46 | 9.66 | 6.46 a | 27.7 |
| NoG | 111.75 | 788.50 | 376.6 b | 34.3 | 151.50 | 710.50 | 421.5 a | 28.3 |
| mGS (cm2) | 0.0005 | 0.0051 | 0.0018 b | 41.3 | 0.0013 | 0.0034 | 0.0022 a | 23.8 |
| mSW (cm) | 0.0461 | 0.1149 | 0.0604 a | 16.5 | 0.0516 | 0.0746 | 0.0607 a | 9.3 |
| Source of Variation | df | Sums of Squares | Mean Squares | Variance Ratio | F pr |
|---|---|---|---|---|---|
| Growth stage (GS) | 1 | 57,345 | 57,345 | 275.3 | <0.001 |
| Genotype | 36 | 56,508 | 1570 | 7.5 | <0.001 |
| Type | 1 | 7965 | 7965 | 38.2 | <0.001 |
| Within type | 35 | 48,543 | 1387 | 6.7 | <0.001 |
| GS × Genotype | 36 | 19,197 | 533 | 2.6 | <0.001 |
| GS × Type | 1 | 3104 | 3104 | 14.9 | <0.001 |
| GS × Within type | 35 | 16,094 | 460 | 2.2 | <0.001 |
| Residual | 518 | 107,882 | 208 | ||
| Total | 591 | 240,933 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boudiar, R.; Cabeza, A.; Fernández-Calleja, M.; Pérez-Torres, A.; Casas, A.M.; González, J.M.; Mekhlouf, A.; Igartua, E. Root Trait Diversity in Field Grown Durum Wheat and Comparison with Seedlings. Agronomy 2021, 11, 2545. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11122545
Boudiar R, Cabeza A, Fernández-Calleja M, Pérez-Torres A, Casas AM, González JM, Mekhlouf A, Igartua E. Root Trait Diversity in Field Grown Durum Wheat and Comparison with Seedlings. Agronomy. 2021; 11(12):2545. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11122545
Chicago/Turabian StyleBoudiar, Ridha, Alejandra Cabeza, Miriam Fernández-Calleja, Antonio Pérez-Torres, Ana M. Casas, Juan M. González, Abdelhamid Mekhlouf, and Ernesto Igartua. 2021. "Root Trait Diversity in Field Grown Durum Wheat and Comparison with Seedlings" Agronomy 11, no. 12: 2545. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11122545