
Genetic Diversity and Relationships of Terebinth (Pistacia terebinthus L.) Genotypes Growing Wild in Turkey

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. DNA Extraction
2.3. In Silico Polymorphic SSR-PCR Reactions
2.4. Molecular Data Analysis
2.4.1. Genetic Diversity
2.4.2. Genetic Relationship
3. Results
3.1. Polymorphism Levels of SSR Loci
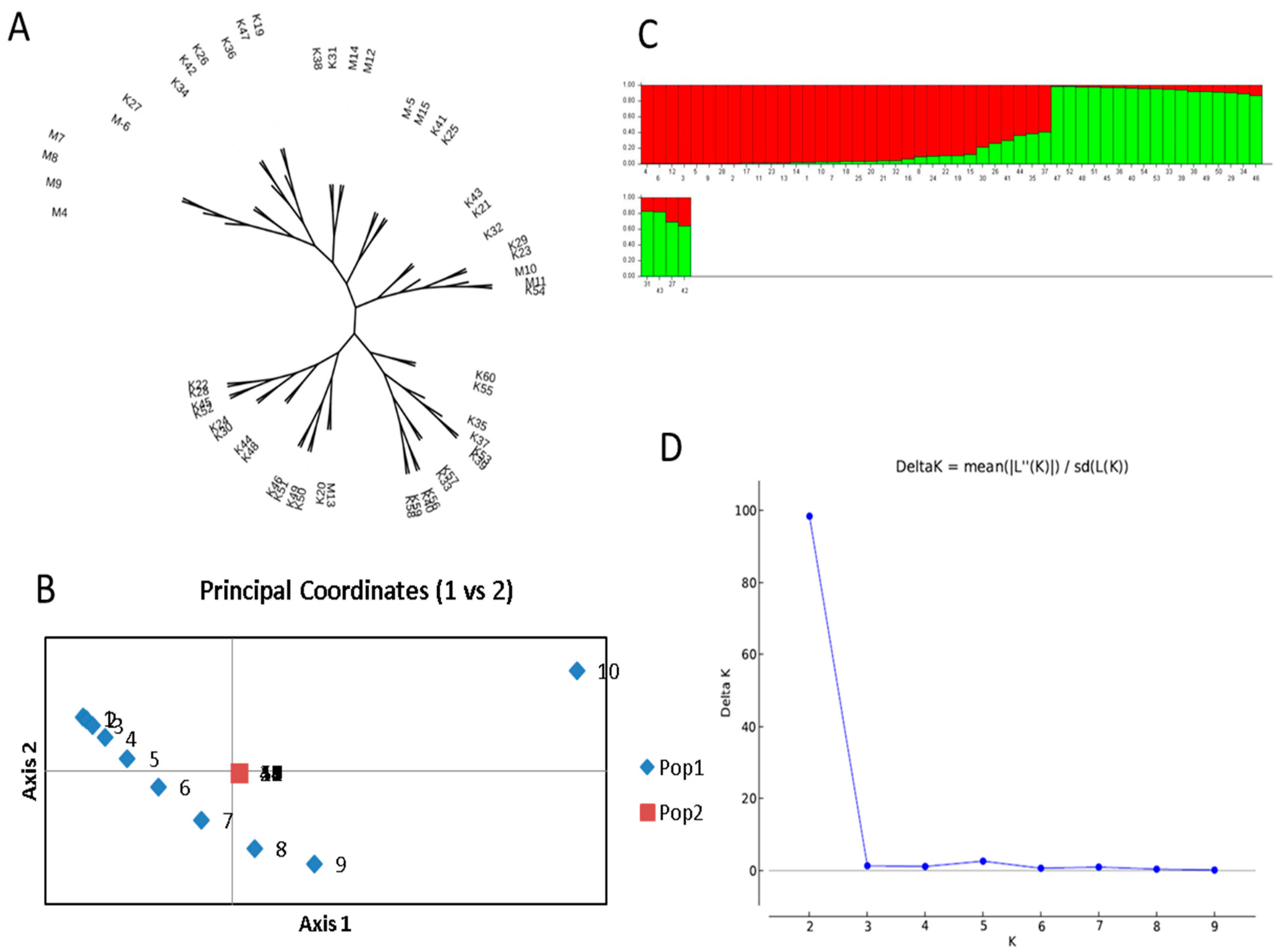
3.2. Genetic Relationships among Terebinth Genotypes
4. Discussion
4.1. In Silico Polymorphic SSR
4.2. Genetic Relationships among Terebinth Genotypes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kafkas, S. Phylogeny Evolution and Biodiversity in the Genus Pistacia (Anacardiaceae). In Plant Genome Biodiversity and Evolution; Sharma, A.K., Sharma, A., Eds.; CRC Press: Plymouth, NJ, USA, 2006; Volume 1, pp. 526–557. [Google Scholar]
- Kozhoridze, G.; Orlovsky, N.; Blumberg, D.G.; Golan-Goldhirsh, A.; Orlovsky, L. Geographic distribution and migration pathways of Pistacia-present, past and future. Ecography 2015, 38, 1141–1154. [Google Scholar] [CrossRef]
- Tunalioglu, R.; Taskaya, B. Antepfıstıgı Tarımsal Ekonomi Araştırma Enstitüsü. TEAE Bakıs 2003, 5, 1–4. [Google Scholar]
- Kuru, C.; Ozsabuncuoglu, I.H. Yabani Pistacia Turlerinin Asılanmasında Sorunlar ve Cozum Yolları. Presented at the Türkiye I, Antepfıstigi Sempozyumu, Pistachio Research Institute, Gaziantep, Turkey, 11–12 September 1990. [Google Scholar]
- Kafkas, S. Phylogenetic analysis of the genus Pistacia by AFLP markers. Plant Syst. Evol. 2006, 262, 113–124. [Google Scholar] [CrossRef]
- Ozbek, S.; Ayfer, M. An hermaphrodite Pistacia found in the vicinity of Antep, Turkey. Proc. Am. Soc. Hortic. Sci. 1958, 72, 240–241. [Google Scholar]
- Crane, J.C. Hermaphroditism in Pistacia. Calif. Agric. 1974, 28, 3–4. [Google Scholar]
- Kafkas, S.; Perl-Treves, R. Morphological and molecular phylogeny of Pistacia species in Turkey. Theor. Appl. Genet. 2001, 102, 908–915. [Google Scholar] [CrossRef]
- Zohary, M. A Monographical Study of the Genus Pistacia. Palest. J. Bot. 1952, 5, 187–228. [Google Scholar]
- Yaltirik, F. Anacardiaceae. In Flora of Turkey; Davis, P.H., Ed.; Edinburgh University Press: Edinburgh, UK, 1967; Volume 2, pp. 544–548. [Google Scholar]
- Ayfer, M.; Okay, Y.; Erdogan, V. Pistachio rootstocks and their propagation. Presented at the Türkiye I, Antepfistigi Sempozyumu Bildirileri, Gaziantep, Turkey, 11–12 September 1990. [Google Scholar]
- Perrino, E.V.; Tomaselli, V.; Costa, R.; Pavone, P. Conservation status of habitats (Directive 92/43 EEC) of coastal and low hill belts in a Mediterranean biodiversity hot spot (Gargano—Italy). Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2013, 147, 1006–1028. [Google Scholar] [CrossRef]
- Yüksel, A.K.; Şat, I.G.; Yüksel, M. The effect of terebinth (Pistacia terebinthus L.) coffee addition on the chemical and physical characteristics, colour values, organic acid profiles, mineral compositions and sensory properties of ice creams. J. Food Sci. Technol. 2015, 52, 8023–8031. [Google Scholar] [CrossRef] [Green Version]
- Koxholt, M.; Eisenmann, B.; Hinrichs, J. Effect of the fat globule sizes on the meltdown of ice cream. J. Dairy Sci. 2001, 84, 31–37. [Google Scholar] [CrossRef]
- Granger, C.; Leger, A.; Barey, P.; Langendorff, V.; Cansell, M. Influence of formulation on the structural networks in ice cream. Int. Dairy J. 2005, 15, 255–262. [Google Scholar] [CrossRef]
- Bonsignore, L.; Cottiglia, F.; Loy, G. Antibacterial activity of Pistacia lentiscus L. aerial parts. Fitoterapia 1998, 69, 537–538. [Google Scholar]
- Ahmad, R.; Ferguson, L.; Southwick, S.M. Analysis of Pistachio rootstocks by SSR and SRAP molecular markers. J. Hortic. Sci. Biotechnol. 2005, 80, 382–386. [Google Scholar] [CrossRef]
- Scarano, M.-T.; Abbate, L.; Ferrante, S.; Lucretti, S.; Tusa, N. ISSR-PCR technique: A useful method for characterizing new allotetraploid somatic hybrids of mandarin. Plant Cell Rep. 2002, 20, 1162–1166. [Google Scholar] [CrossRef]
- Cadée, N. Genetic and environmental effects on morphology and fluctuating asymmetry in nestling barn swallows. J. Evol. Biol. 2000, 13, 359–370. [Google Scholar] [CrossRef]
- Bucheyeki, T.; Gwanama, C.; Mgonja, M.; Chisi, M.; Folkertsma, R.; Mutegi, R. Genetic variability characterisation of Tanzania sorghum landraces based on simple sequence repeats (ssrs) molecular and morphological markers. Afr. Crop Sci. J. 2010, 17, 71–86. [Google Scholar] [CrossRef]
- Güney, M.; Kafkas, S.; Keles, H.; Aras, S.; Ercişli, S. Characterization of hawthorn (Crataegus spp.) genotypes by SSR markers. Physiol. Mol. Biol. Plants 2018, 24, 1221–1230. [Google Scholar] [CrossRef]
- Kafkas, S. Advances in Breeding of Pistachio; Burleigh Dodds Science Publishing: Cambridge, UK, 2019; pp. 389–430. [Google Scholar]
- Tautz, D. Hypervariability of simple sequences as a general source for polymorphic DNA markers. Nucleic Acids Res. 1989, 17, 6463–6471. [Google Scholar] [CrossRef]
- Akkaya, M.S.; A Bhagwat, A.; Cregan, P.B. Length polymorphisms of simple sequence repeat DNA in soybean. Genetics 1992, 132, 1131–1139. [Google Scholar] [CrossRef]
- Tóth, G.; Gáspári, Z.; Jurka, J. Microsatellites in Different Eukaryotic Genomes: Survey and Analysis. Genome Res. 2000, 10, 967–981. [Google Scholar] [CrossRef] [Green Version]
- Zaloğlu, S.; Kafkas, S.; Doğan, Y.; Güney, M. Development and characterization of SSR markers from pistachio (Pistacia vera L.) and their transferability to eight Pistacia species. Sci. Hortic. 2015, 189, 94–103. [Google Scholar] [CrossRef]
- Hormaza, J. Molecular characterization and similarity relationships among apricot (Prunus armeniaca L.) genotypes using simple sequence repeats. Theor. Appl. Genet. 2002, 104, 321–328. [Google Scholar] [CrossRef]
- Potts, S.M.; Han, Y.; Khan, M.A.; Kushad, M.M.; Rayburn, A.L.; Korban, S.S. Genetic Diversity and Characterization of a Core Collection of Malus Germplasm Using Simple Sequence Repeats (SSRs). Plant Mol. Biol. Rep. 2012, 30, 827–837. [Google Scholar] [CrossRef]
- Fan, L.; Zhang, M.-Y.; Liu, Q.-Z.; Li, L.T.; Song, Y.; Wang, L.-F.; Zhang, S.-L.; Wu, J. Transferability of Newly Developed Pear SSR Markers to Other Rosaceae Species. Plant Mol. Biol. Rep. 2013, 31, 1271–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Topçu, H.; Ikhsan, A.S.; Sütyemez, M.; Çoban, N.; Güney, M.; Kafkas, S. Development of 185 polymorphic simple sequence repeat (SSR) markers from walnut (Juglans regia L.). Sci. Hortic. 2015, 194, 160–167. [Google Scholar] [CrossRef]
- Güney, M.; Kafkas, S.; Koç, A.; Aras, S.; Keleş, H. Characterization of quince (Cydonia oblonga Mill.) accessions by simple sequence repeat markers. Turk. J. Agric. For. 2019, 43, 69–79. [Google Scholar] [CrossRef]
- Hoffman, J.I.; Nichols, H.J. A novel approach for mining polymorphic microsatellite markers in silico. PLoS ONE 2011, 6, e23283. [Google Scholar] [CrossRef]
- Vukosavljev, M.; Esselink, G.D.; van’t Westende, W.P.C.; Cox, P.; Visser, R.G.F.; Arens, P.; Smulders, M.J.M. Efficient development of highly polymorphic microsatellite markers based on polymorphic repeats in transcriptome sequences of multiple individuals. Mol. Ecol. Resour. 2015, 15, 17–27. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A Rapid isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Khodaeiaminjan, M.; Kafkas, S.; Motalebipour, E.Z.; Çoban, N. In silico polymorphic novel SSR marker development and the first SSR-based genetic linkage map in pistachio. Tree Genet. Genomes 2018, 14, 45. [Google Scholar] [CrossRef]
- Motalebipour, E.Z.; Gozel, H.; Khodaeiaminjan, M.; Kafkas, S. SSR-based genetic linkage map construction in pistachio using an interspecific F1 population and QTL analysis for leaf and shoot traits. Mol. Breed. 2018, 38, 134. [Google Scholar] [CrossRef]
- Schuelke, M. An economic method for the fluorescent labeling of PCR fragments. Nat. Biotechnol. 2000, 18, 233–234. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.; Smouse, P.E. Genalex 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Liu, K.; Muse, S.V. PowerMarker: Integrated Analysis Environment for Genetic Marker Data. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K.; Battistuzzi, F.U. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar]
- Vendramin, E.; Dettori, M.; Verde, I.; Micali, S.; Giovinazzi, J.; Mardi, M.; Avanzato, D.; Quarta, R. Molecular characterization of Pistacia genus by microsatellite markers. Acta Hortic. 2009, 55–62. [Google Scholar] [CrossRef]
- Topçu, H.; Çoban, N.; Kafkas, S. Novel microsatellite markers in Pistacia vera L. and their transferability across the genus Pistacia. Sci. Hortic. 2016, 198, 91–97. [Google Scholar] [CrossRef]
- Motalebipour, E.Z.; Kafkas, S.; Khodaeiaminjan, M.; Çoban, N.; Gözel, H. Genome survey of pistachio (Pistacia vera L.) by next generation sequencing: Development of novel SSR markers and genetic diversity in Pistacia species. BMC Genom. 2016, 17, 998. [Google Scholar] [CrossRef] [Green Version]
- Karcι, H.; Paizila, A.; Topçu, H.; Ilikcioglu, E.; Kafkas, S. Transcriptome sequencing and development of novel genic SSR markers from Pistacia vera L. Front. Genet. 2020, 11, 1021. [Google Scholar] [CrossRef] [PubMed]
- Parfitt, D.E.; Badenes, M.L. Phylogeny of the genus Pistacia as determined from analysis of the chloroplast genome. Proc. Natl. Acad. Sci. USA 1997, 94, 7987–7992. [Google Scholar] [CrossRef] [Green Version]
- Engler, A. Burseraceae et Anacardiaceae. In Monopgraphiae Phaneragamarum; De-Candolle, A.C., Ed.; Lubrecht and Cramer Ltd.: Port Jervis, NY, USA, 1883; pp. 284–293. [Google Scholar]
- Golan-Goldhirsh, A.; Barazani, O.; Wang, Z.S.; Khadka, D.K.; Saunders, J.A.; Kostiukovsky, V.; Rowland, L.J. Genetic relationships among Mediterranean Pistacia species evaluated by RAPD and AFLP markers. Plant Syst. Evol. 2004, 246, 9–18. [Google Scholar] [CrossRef]
- Kafkas, S.; Perl-Traves, R.; Kaska, N. Unusual Pistacia atlantica Desf. (Anacardiaceae) monoecious sex type in the Yunt mountains of the Manisa province of Turkey. Israil J. Plant. Sci. 2000, 48, 277–280. [Google Scholar] [CrossRef]
- Avanzato, D. Una forma monoica di Pistacia terebinthus found scoperta in Bulgaria. Frutticoltura 2003, 10, 51–53. [Google Scholar]
- Ercisli, S.; Esitken, A.; Cangi, R.; Şahin, F. Adventitious root formation of kiwifruit in relation to sampling date, IBA and Agrobacterium rubi inoculation. Plant Growth Regul. 2003, 41, 133–137. [Google Scholar] [CrossRef]
- Dogan, H.; Ercisli, S.; Temim, E.; Hadziabulic, A.; Tosun, M.; Yilmaz, S.O.; Zia-Ul-Haq, M. Diversity of chemical content and biological activity in flower buds of a wide number of wild grown caper (Capparis ovate Desf.) genotypes from Turkey. Comptes Rendus Acad. Bulg. Sci. 2014, 67, 1593–1600. [Google Scholar]
- Dogan, H.; Ercisli, S.; Jurikova, T.; Temim, E.; Leto, A.; Hadziabulic, A.; Tosun, M.; Narmanlioglu, H.K.; Zia-Ul-Haq, M. Physicochemical and antioxidant characteristics of fruits of cape gooseberry (Physalis peruviana L.) from Turkey. Oxid. Commun. 2014, 37, 1005–1014. [Google Scholar]
- Kaškonienė, V.; Bimbiraitė-Survilienė, K.; Kaškonas, P.; Tiso, N.; Česonienė, L.; Daubaras, R.; Maruška, A.S. Changes in the biochemical compounds of Vacciniummyrtillus, Vacciniumvitis-idaea, and forest litter collected from various forest types. Turk. J. Agric. For. 2020, 44, 557–566. [Google Scholar] [CrossRef]
- Ozkan, G.; Ercisli, S.; Sagbas, H.I.; Ilhan, G. Diversity on Fruits of Wild Grown European Cranberrybush from Coruh Valley in Turkey. Erwerbs Obstbau 2020, 62, 275–279. [Google Scholar] [CrossRef]
- Sagbas, H.; Ilhan, G.; Ercisli, S.; Anjum, M.; Holubec, V. Characterization of Oleaster-Leafed Pear (Pyrus elaeagrifolia Pall. subsp. elaeagrifolia) Fruits in Turkey. Agronomy 2021, 11, 430. [Google Scholar] [CrossRef]
- Ercisli, S.; Esitken, A. Fruit characteristics of native rose hip (Rosa spp.) selections from the Erzurum province of Turkey. N. Z. J. Crop Hortic. Sci. 2004, 32, 51–53. [Google Scholar] [CrossRef] [Green Version]
- Maxted, N.; Ford-Lloyd, B.V.; Jury, S.; Kell, S.; Scholten, M. Towards a definition of a crop wild relative. Biodivers. Conserv. 2006, 15, 2673–2685. [Google Scholar] [CrossRef]
- McCouch, S.; Baute, G.J.; Bradeen, J.; Bramel, P.; Bretting, P.K.; Buckler, E.; Burke, J.M.; Charest, D.; Cloutier, S.; Cole, G.; et al. Feeding the future. Nat. Cell Biol. 2013, 499, 23–24. [Google Scholar] [CrossRef]
- Perrino, E.V.; Perrino, P. Crop wild relatives: Know how past and present to improve future research, conservation and utilization strategies, especially in Italy: A review. Genet. Resour. Crop. Evol. 2020, 67, 1067–1105. [Google Scholar] [CrossRef]
- Gecer, M.K.; Kan, T.; Gundogdu, M.; Ercisli, S.; Ilhan, G.; Sagbas, H.I. Physicochemical characteristics of wild and cultivated apricots (Prunus armeniaca L.) from Aras valley in Turkey. Genet. Resour. Crop Evol. 2020, 67, 935–945. [Google Scholar] [CrossRef]
- Gundogdu, M.; Ozrenk, K.; Ercisli, S.; Kan, T.; Kodad, O.; Hegedus, A. Organic acids, sugars, vitamin C content and some pomological characteristics of eleven hawthorn species (Crataegus spp.) from Turkey. Biol. Res. 2014, 47, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| No. | Genotypes | Sex | No. | Genotypes | Sex |
|---|---|---|---|---|---|
| 1 | *M4 | M | 28 | K34 | F |
| 2 | *M-5 | F | 29 | K35 | F |
| 3 | *M-6 | M | 30 | K36 | M |
| 4 | *M7 | M | 31 | K37 | M |
| 5 | *M8 | M | 32 | K38 | F |
| 6 | *M9 | M | 33 | K39 | M |
| 7 | *M10 | F | 34 | K40 | F |
| 8 | *M11 | F | 35 | K41 | M |
| 9 | *M12 | F | 36 | K42 | F |
| 10 | *M13 | F | 37 | K43 | M |
| 11 | *M14 | F | 38 | K44 | F |
| 12 | *M15 | F | 39 | K45 | M |
| 13 | K19 | M | 40 | K46 | F |
| 14 | K20 | F | 41 | K47 | M |
| 15 | K21 | F | 42 | K48 | M |
| 16 | K22 | F | 43 | K49 | M |
| 17 | K23 | M | 44 | K50 | M |
| 18 | K24 | M | 45 | K51 | F |
| 19 | K25 | M | 46 | K52 | M |
| 20 | K26 | M | 47 | K53 | M |
| 21 | K27 | M | 48 | K54 | M |
| 22 | K28 | F | 49 | K55 | F |
| 23 | K29 | M | 50 | K56 | F |
| 24 | K30 | M | 51 | K57 | F |
| 25 | K31 | F | 52 | K58 | F |
| 26 | K32 | M | 53 | K59 | F |
| 27 | K33 | F | 54 | K60 | F |
| No | Origin and Reference | Acronyms | Marker Types | No. of Tested Loci | No. of Non-Amplified Loci | Polymorphic | Polymorphic Rate (%) |
|---|---|---|---|---|---|---|---|
| 1 | Khodaeiaminjan et al. [20] | CUPSiOh, CUPSiPa, CUPOhBa, CUPSiBa | In silico polymorphic SSRs | 45 | 7 | 38 | 84.44 |
| 2 | Motalebipour et al. [21] | CUPVSiirt | SSRs | 5 | 3 | 2 | 40.00 |
| Total | 50 | 10 | 40 |
| No. | SSR Loci | Allele Range | Sequence (5′–3′) | Sequence (5′–3′) | Na | Ne | Ho | He | PIC |
|---|---|---|---|---|---|---|---|---|---|
| 1 | CUPSiOh2688 | 122–160 | GACTACTGTGCCCACATGACA | GGATCGTCAGAGAAGACGTTG | 13 | 4.00 | 0.72 | 0.75 | 0.72 |
| 2 | CUPSiPa1067 | 125–141 | GTAAGAATCCCGAGGAAGCAC | GGCAAGCATTGTAAGTTGCTC | 7 | 1.98 | 0.50 | 0.50 | 0.47 |
| 3 | CUPOhBa2293 | 154–181 | TGAAGCCTTGGAAACATAGGA | TTTGGTGGAAGAAATTGTGGT | 9 | 1.98 | 0.45 | 0.49 | 0.48 |
| 4 | CUPOhPa2145 | 101–113 | GGAACTGGGTTTGGCACTACT | GGCAGTTGTTTGTTGATTGATT | 5 | 2.42 | 0.50 | 0.59 | 0.52 |
| 5 | CUPOhPa4477 | 147–157 | TGGATTCTTTGGATCCATTTT | ATAGCTGTGCACGTCTTTGGT | 4 | 2.31 | 0.58 | 0.57 | 0.48 |
| 6 | CUPSiPa185 | 149–169 | CATTGTCAATTTATGATCACTAGAGC | CTCCCCCTTCTCATTTTTGAT | 5 | 3.04 | 0.70 | 0.67 | 0.61 |
| 7 | CUPSiPa2316 | 151–192 | TGATGGAACTCCCTGTCAAAC | TGGTGATTGTTATTCCCATCC | 17 | 7.41 | 0.87 | 0.87 | 0.85 |
| 8 | CUPSiPa2184 | 125–131 | CATGTTAAGCAATGAAATGTGGA | GCATGTAACATCCATACAGAACTTG | 4 | 1.75 | 0.28 | 0.43 | 0.39 |
| 9 | CUPOhBa4756 | 139–182 | GGTTGGGATAAGTAACGGTGA | GGCGAGCAATCAATCAGTAAG | 19 | 8.81 | 0.98 | 0.89 | 0.88 |
| 10 | CUPOhPa3954 | 105–143 | TTGAATGTTGGCTAATGC | TGAAGACTTTAAAAAGGGCGATT | 17 | 9.44 | 0.81 | 0.89 | 0.89 |
| 11 | CUPSiOh2751 | 142–160 | TGAGAGCAATAAATGCAATCAAC | TGGGTGTAGGAGACTTGTGGA | 10 | 3.96 | 0.68 | 0.75 | 0.72 |
| 12 | CUPOhPa1967 | 146–178 | TGAATCATTCTCCCCAAACTC | CGACAATGAAAGTCTTCTGACG | 9 | 3.96 | 0.42 | 0.75 | 0.71 |
| 13 | CUPOhPa4736 | 140–177 | TTGGGAGAAGTAACGGTGACA | GGCGAGCAATCAATCAGTAAG | 19 | 7.85 | 0.92 | 0.87 | 0.86 |
| 14 | CUPSiOh2517 | 118–132 | CCAGAATTTGTTGGAAGTTGC | TTATCTCACATGAGGCAAAAT | 7 | 4.03 | 0.80 | 0.75 | 0.71 |
| 15 | CUPSiOh4567 | 225–261 | GTAAGAATCCCGAGGAAGCAC | GGCAAGCATTGTAAGTTGCTC | 12 | 7.49 | 0.85 | 0.87 | 0.85 |
| 16 | CUPSiBa4072 | 107–145 | GCAAACTCAATGAGGTAAAGCCTG | AATTAAGTCCTTGGACAGTGTGGG | 21 | 12.38 | 0.88 | 0.92 | 0.91 |
| 17 | CUPSiPa732 | 160–211 | GCACAAAAGTGGTTCTAACAAT | TTCCATGTTCTGTATGGTCAAGA | 25 | 10.35 | 0.45 | 0.90 | 0.90 |
| 18 | CUPSiBa3091 | 217–252 | CATTGATATCAGCACGACCTGTTC | TTTCAACAACCATATCCAAACTTAAACA | 9 | 2.93 | 0.66 | 0.66 | 0.63 |
| 19 | CUPSiPa3238 | 160–186 | GGGACCATATGGGTTGATAAAA | GACCCCTGTTATACTTAATTAAAGGTC | 15 | 8.34 | 0.85 | 0.88 | 0.87 |
| 20 | CUPOhPa4338 | 113–119 | AACAAAAGCATAAAACACCTCGT | GAAAATATCAAAGGTTTCTTGAGCA | 4 | 2.93 | 0.52 | 0.66 | 0.59 |
| 21 | CUPSiPa813 | 101–125 | GGGAGCTGTGAGGTTATTCCT | TCTGTGCAACACCAAAACAAA | 8 | 2.22 | 0.53 | 0.55 | 0.52 |
| 22 | CUPOhBa1030 | 120–148 | TGCGAGGGGTAAACATAATGA | CCCTCGCAATATAAATAGAAAGTC | 10 | 5.23 | 0.93 | 0.81 | 0.79 |
| 23 | CUPOhBa4344 | 157–189 | TTCGCCTTTGAAATTCATACC | CTCGTTTGACACGTGCATTTA | 6 | 1.56 | 0.42 | 0.36 | 0.34 |
| 24 | *CUPVSiirt625 | 150–186 | AATTACCAGCTTAGGCCCATC | TGCAAGGTACGAGACTTTTGG | 14 | 8.39 | 1.00 | 0.88 | 0.87 |
| 25 | *CUPVSiirt1326 | 184–214 | AGAGAGGGAGAAAACGCTTCA | GTCGCATGGAGATTGACTTTG | 11 | 5.39 | 1.00 | 0.81 | 0.79 |
| 26 | CUPSiOh4281 | 112–141 | TTCCCAATTTCTGTGACTAAAGC | AAAAGAATTCCCGTGTGATTTT | 17 | 9.00 | 0.94 | 0.89 | 0.88 |
| 27 | CUPOhBa281 | 114–148 | ATTTCTGAAGAAGCAGCAGCA | TCTCTGGATTTTCTGGTGTCG | 21 | 8.69 | 0.61 | 0.88 | 0.88 |
| 28 | CUPOhPa2243 | 152–186 | TTTGCTAGTGCAAGCTCCTTT | TTTTGGTCACTAGATCAAACAAGG | 8 | 4.27 | 0.78 | 0.77 | 0.73 |
| 29 | CUPOhPa2632 | 149–162 | GGCAAATCCGTAGCCATAAAT | GGGAGGAAGATCCATAAGAAGA | 3 | 2.04 | 0.46 | 0.51 | 0.46 |
| 30 | CUPOhPa4270 | 137–141 | CAAGTATGTGAATGTTCATGG | AGTGGACCATTGAGCAAACC | 3 | 2.12 | 0.58 | 0.53 | 0.42 |
| 31 | CUPSiPa4413 | 130–150 | CATGTTGAGTTTGCTGATGGA | CGAAACGCTAAGACCTTCTCA | 10 | 2.78 | 0.56 | 0.64 | 0.59 |
| 32 | CUPSiPa4275 | 145–184 | GGACCTCTCATTTCAAACAGAA | CACCGGCCACATGTACTTTTA | 19 | 8.78 | 0.94 | 0.89 | 0.88 |
| 33 | CUPSiPa124 | 152–176 | TTCTCAAACGTCTTCATGCCTA | GTGCAGCTTGCTCTTTGTTTC | 5 | 2.52 | 0.48 | 0.60 | 0.52 |
| 34 | CUPOhPa4043 | 158–173 | GGAGAAGCGGATAGGAGAAAG | TCCACAGGCATCAACAGAAAT | 7 | 3.97 | 0.80 | 0.75 | 0.71 |
| 35 | CUPSiPa518 | 158–177 | GACATAATTGAATGATTTGG | CCAAGCCACATAAACTAGCAA | 6 | 2.41 | 0.69 | 0.59 | 0.55 |
| 36 | CUPSiOh2976 | 145–179 | CCTGGTCTGATTCTCTCGTTC | CACAGGAGCAGATGTTCCTACA | 15 | 4.51 | 0.61 | 0.78 | 0.76 |
| 37 | CUPSiPa1654 | 149–179 | GATAAGGCAACCACTGAGCAT | ATGCATTTCCAAGAAAGCAGA | 14 | 6.53 | 0.91 | 0.85 | 0.83 |
| 38 | CUPSiOh1518 | 100–127 | TGGTATTAACCGTTCCACATGA | AGCACTCCTGAAGTGGGAGTA | 7 | 2.71 | 0.52 | 0.63 | 0.57 |
| 39 | CUPOhBa4362 | 100–126 | ACAAGCAGGACATCAAAGGTG | CCATGCTCAACAATCATGAAG | 11 | 5.86 | 0.57 | 0.83 | 0.81 |
| 40 | CUPOhBa546 | 107–115 | ATTGGAATTGGAGAGACATGGA | AACACTCTGTTTCGCTATTTTCAC | 4 | 1.61 | 0.42 | 0.38 | 0.35 |
| Total | - | - | 430 | - | - | - | |||
| Min | - | - | 3 | 1.56 | 0.28 | 0.36 | 0.34 | ||
| Mean | - | - | 11 | 4.95 | 0.68 | 0.71 | 0.68 | ||
| Max | - | - | 25 | 12.38 | 1.00 | 0.92 | 0.91 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guney, M.; Kafkas, S.; Zarifikhosroshahi, M.; Gundesli, M.A.; Ercisli, S.; Holubec, V. Genetic Diversity and Relationships of Terebinth (Pistacia terebinthus L.) Genotypes Growing Wild in Turkey. Agronomy 2021, 11, 671. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11040671
Guney M, Kafkas S, Zarifikhosroshahi M, Gundesli MA, Ercisli S, Holubec V. Genetic Diversity and Relationships of Terebinth (Pistacia terebinthus L.) Genotypes Growing Wild in Turkey. Agronomy. 2021; 11(4):671. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11040671
Chicago/Turabian StyleGuney, Murat, Salih Kafkas, Mozhgan Zarifikhosroshahi, Muhammet Ali Gundesli, Sezai Ercisli, and Vojtěch Holubec. 2021. "Genetic Diversity and Relationships of Terebinth (Pistacia terebinthus L.) Genotypes Growing Wild in Turkey" Agronomy 11, no. 4: 671. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11040671