
Yield Comparisons between Cotton Variety Xin Nong Mian 1 and Its Transgenic ScALDH21 Lines under Different Water Deficiencies in a Desert-Oasis Ecotone

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
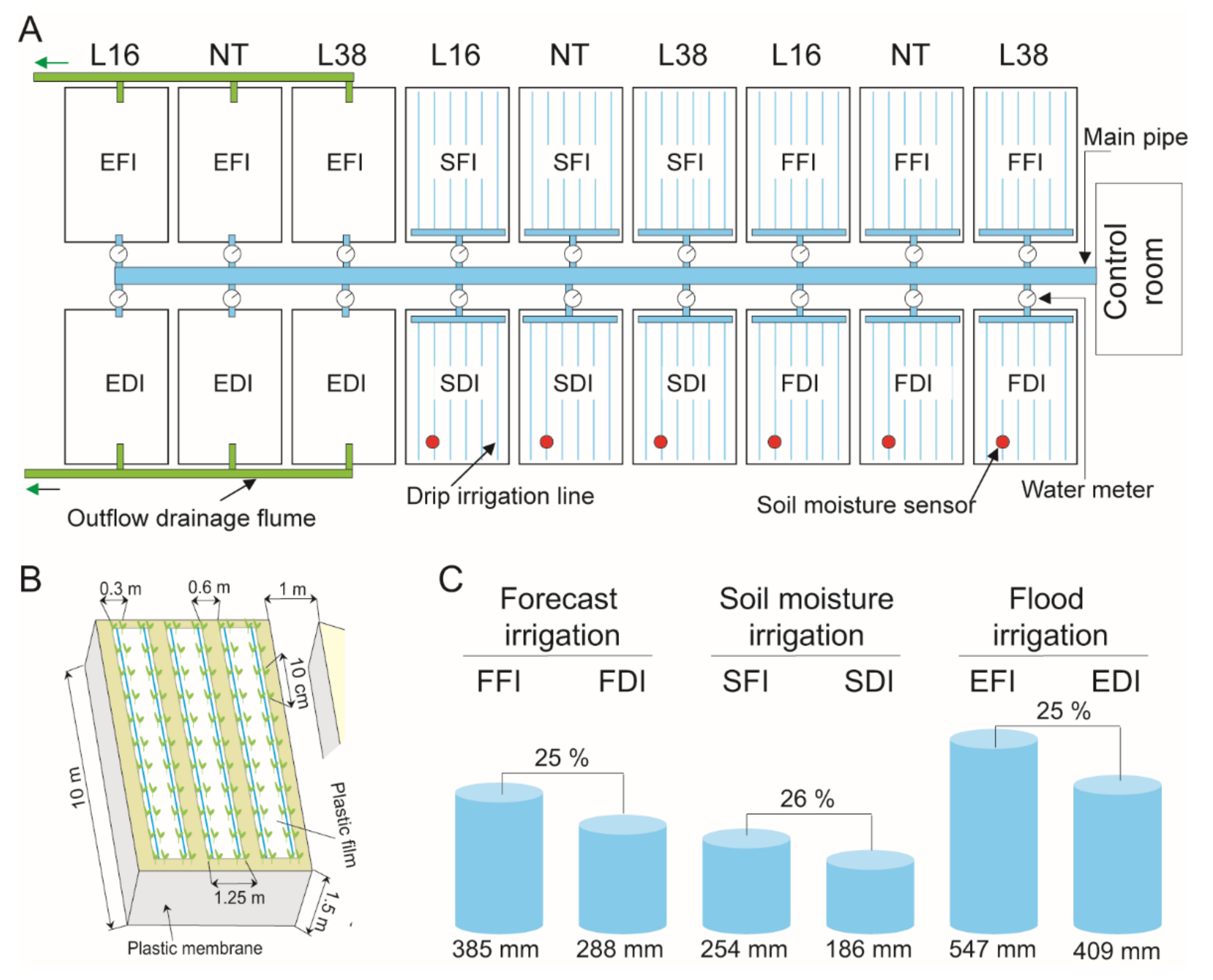
2.1. Plant Material and Field Experiment Design
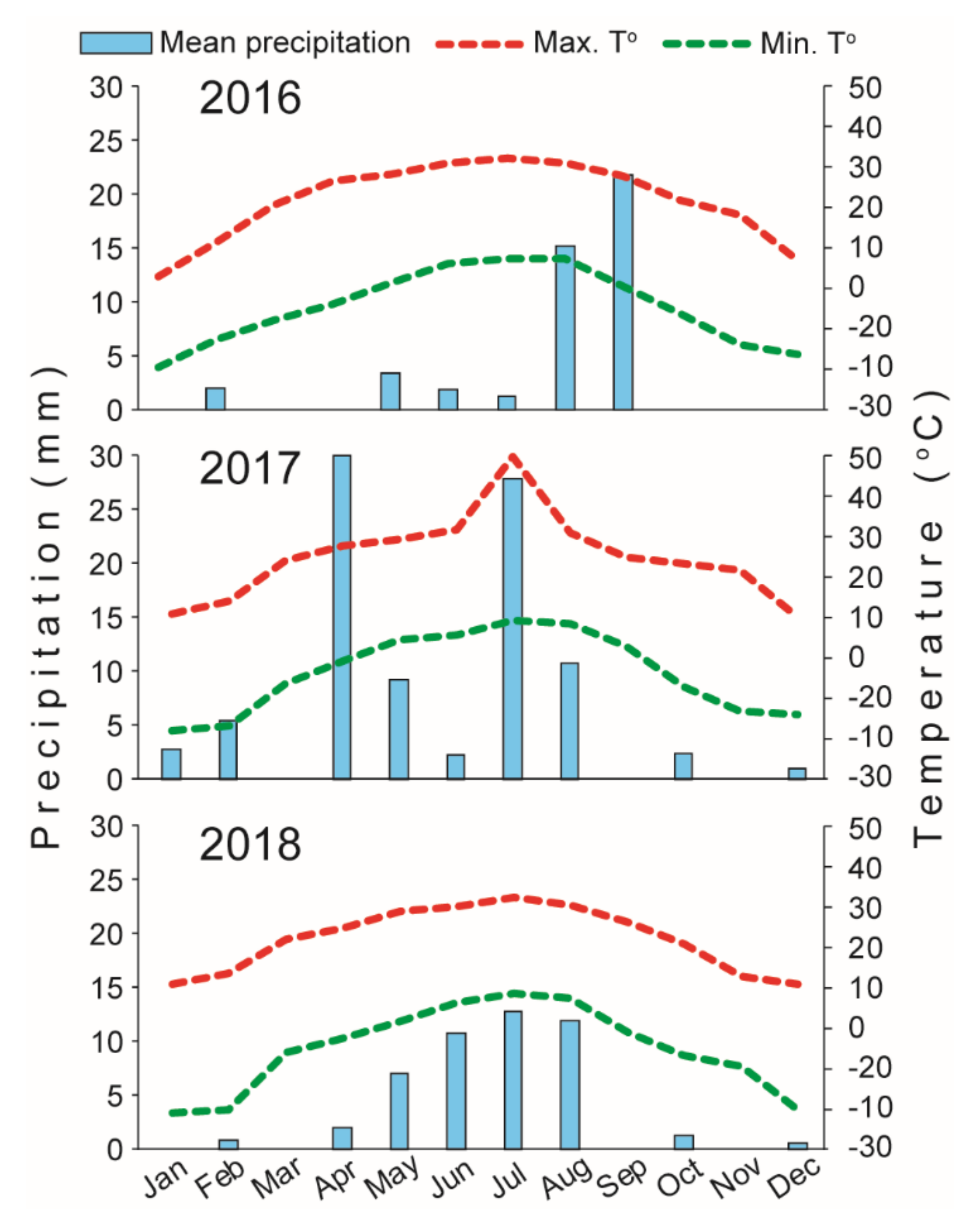
2.2. Study Sites
2.3. Physiological Measurements
2.4. Phenotypic Measurements
2.5. Statistics Analysis
3. Results
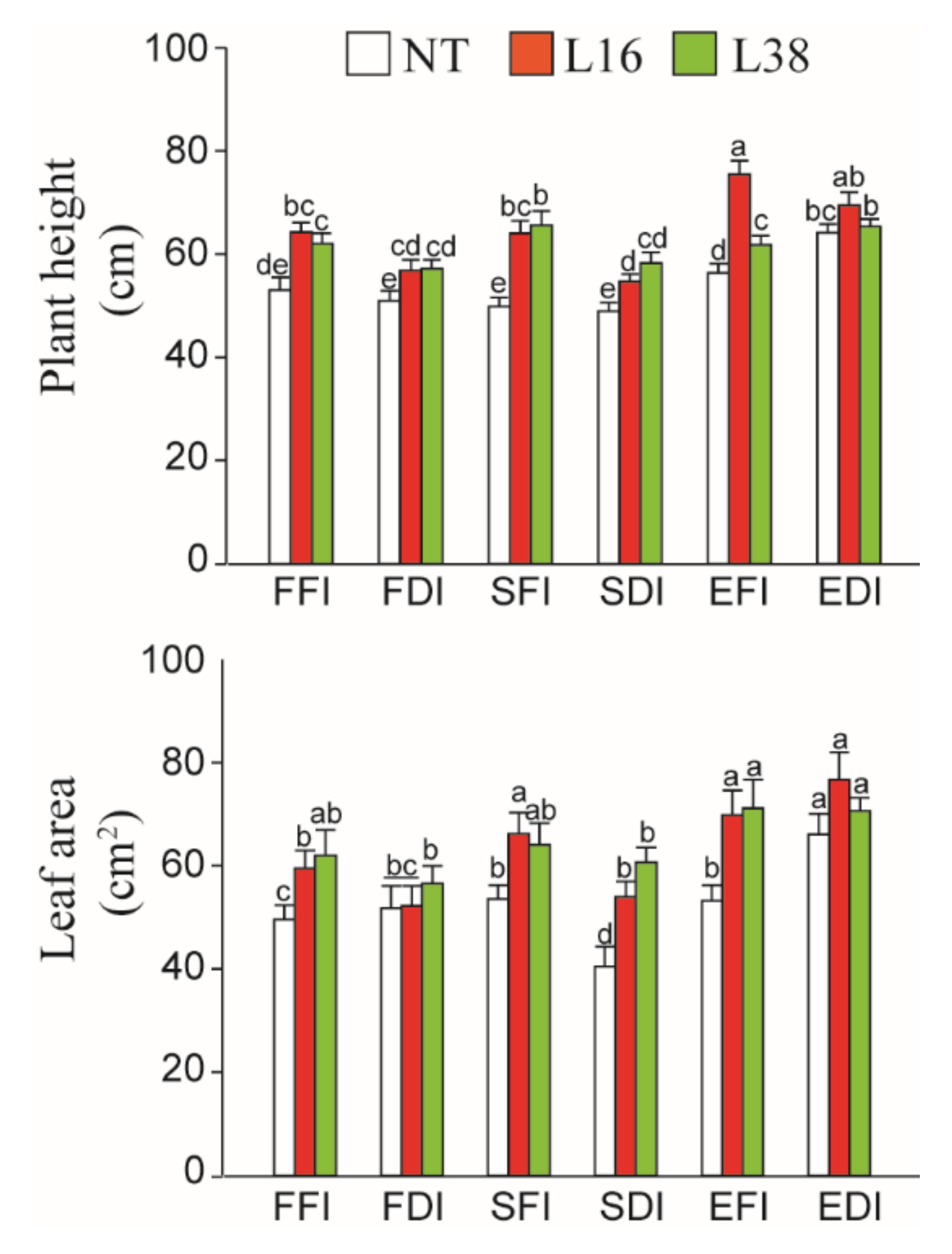
3.1. Effects of Different Water Deficiency Treatments on Plant Growth and Development
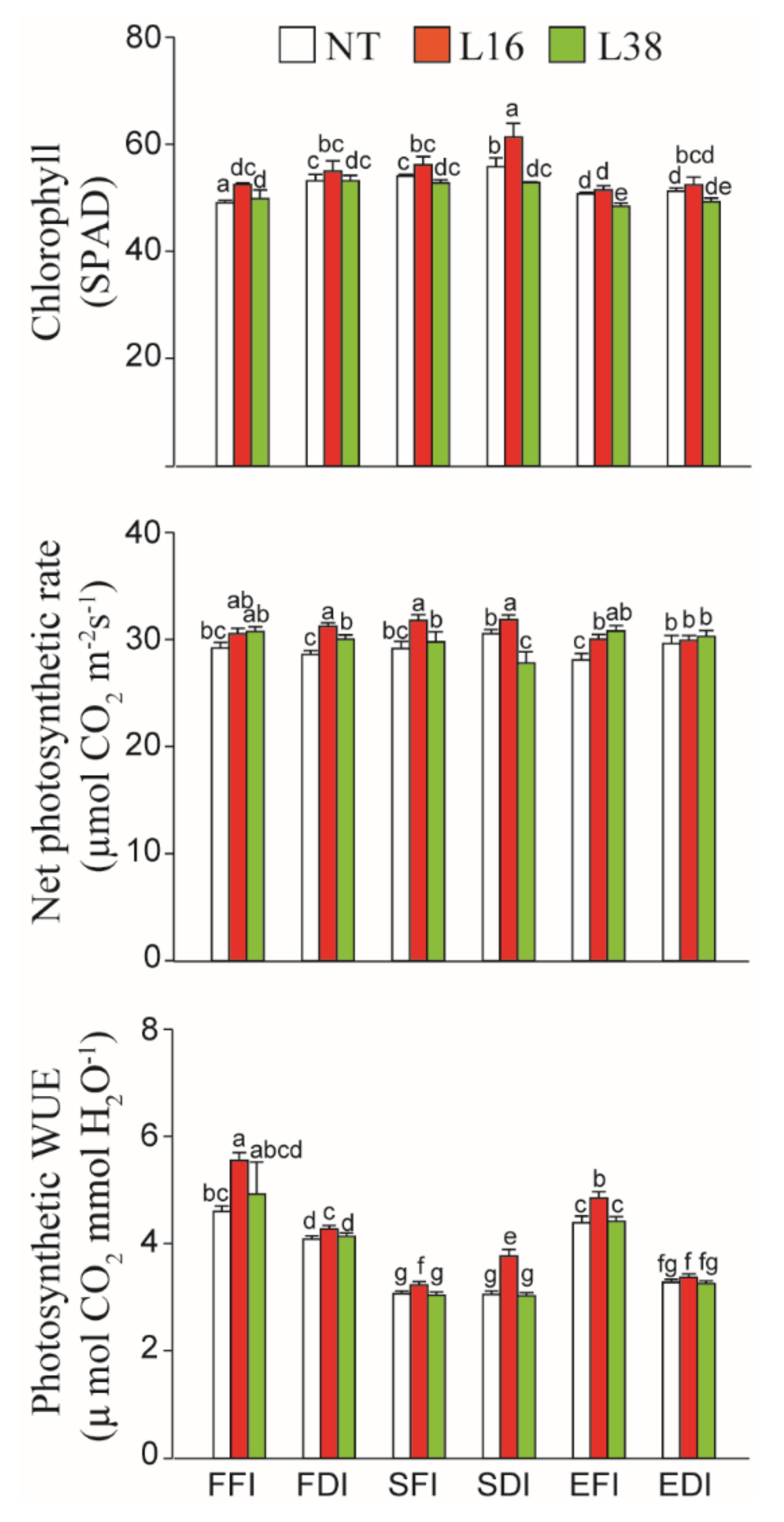
3.2. Effects of Irrigation Treatments on the Chlorophyll Content (SPAD Value) and Net Photosynthetic Rate of Cotton
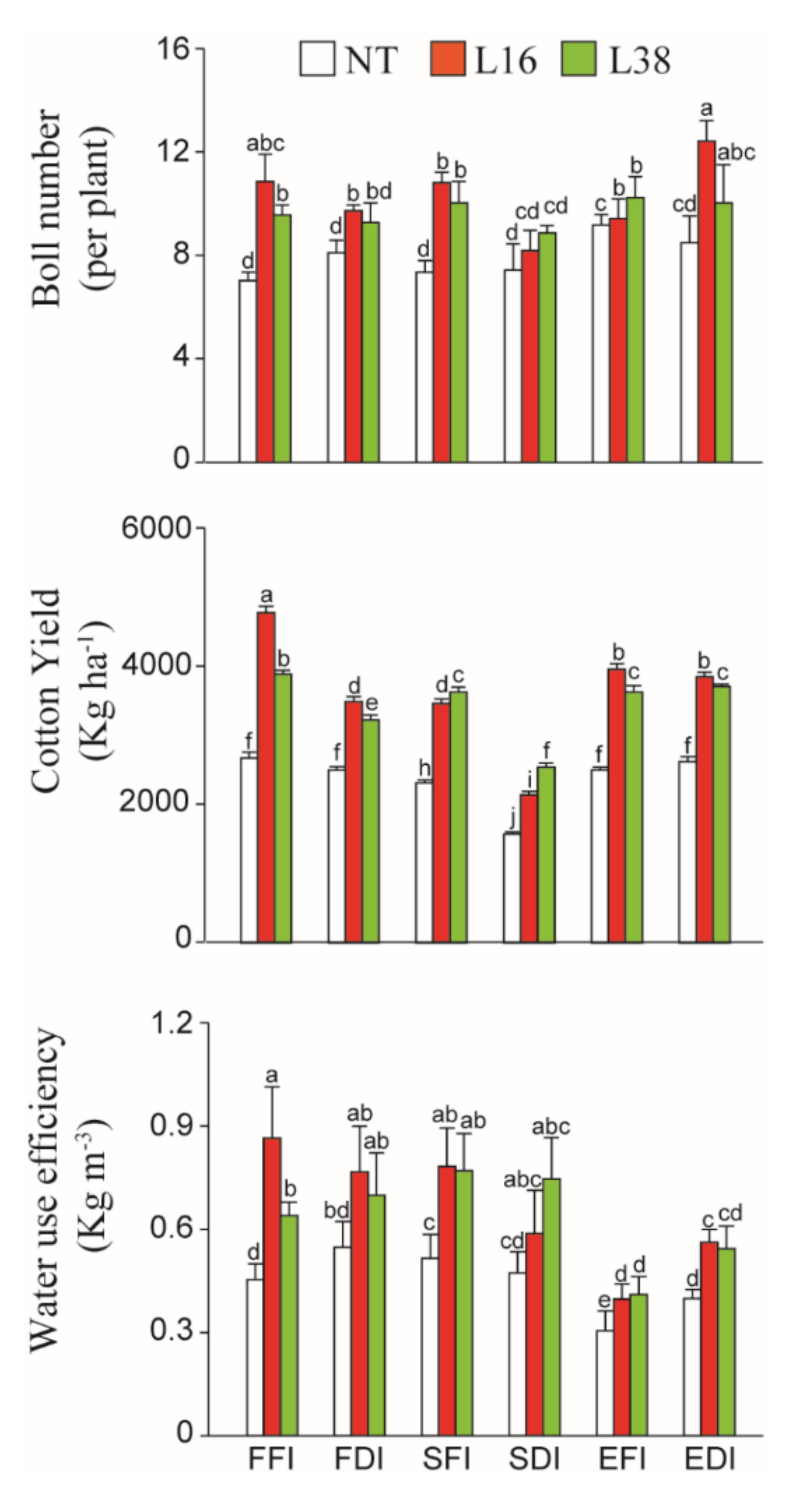
3.3. Cotton Yield Components and WUE
3.4. Effect of Irrigation Water Management on Fiber Quality
3.5. Multifactorial Analysis of Cotton Lines, Irrigation Levels and Irrigation Protocols on Cotton Performance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| NT | Non-transgenic |
| TC | Transgenic cotton |
| WUE | Water use efficiency |
| IWUE | Instantaneous water use efficiency |
| FFI | Forecasted full irrigation |
| FDI | Forecasted deficit irrigation |
| DSSIS-based | Decision support system for irrigation scheduling |
| SFI | Soil moisture sensor-based full irrigation |
| SDI | Soil moisture sensor-based deficit irrigation |
| EFI | Experience-based full irrigation |
| EDI | Experience-based deficit irrigation |
| RZWQM2 | Root zone water quality model |
| I | Irrigation protocols |
| L | Cotton lines |
| SCY | Seed cotton yield |
| POD | Peroxidase |
| MDA | Malondialdehyde |
References
- Zhu, X.L.; Sun, L.; Kuppu, S.; Hu, R.B.; Mishra, N.; Smith, J.; Esmaeili, N.; Herath, M.; Gore, M.A.; Payton, P.; et al. The yield difference between wildtype cotton and transgenic cotton that expresses IPT depends on when water-deficit stress is applied. Sci. Rep. 2018, 8, 1–11. [Google Scholar]
- Chaves, M.M.; Maroco, J.P.; Pereira, J.S. Understanding plant responses to drought-from genes to the whole plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef]
- Cooper, M.; Messina, C.D.; Podlich, D.; Totir, L.R.; Baumgarten, A.; Hausmann, N.J.; Wright, D.; Graham, G. Predicting the future of plant breeding: Complementing empirical evaluation with genetic prediction. Crop Pasture Sci. 2014, 65, 311–336. [Google Scholar] [CrossRef] [Green Version]
- James, C. Global adoption, impact and future prospects of biotech/GM crops. J. Biotechnol. 2008, 136, S213–S214. [Google Scholar] [CrossRef]
- Ronald, P.C. Lab to Farm: Applying Research on Plant Genetics and Genomics to Crop Improvement. PLoS Biol. 2014, 12. [Google Scholar] [CrossRef] [Green Version]
- Vinocur, B.; Altman, A. Recent advances in engineering plant tolerance to abiotic stress: Achievements and limitations. Curr. Opin. Biotech. 2005, 16, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Castiglioni, P.; Warner, D.; Bensen, R.J.; Anstrom, D.C.; Harrison, J.; Stoecker, M.; Abad, M.; Kumar, G.; Salvador, S.; D’Ordine, R.; et al. Bacterial RNA chaperones confer abiotic stress tolerance in plants and improved grain yield in maize under water-limited conditions. Plant Physiol. 2008, 147, 446–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deikman, J.; Petracek, M.; Heard, J.E. Drought tolerance through biotechnology: Improving translation from the laboratory to farmers’ fields. Curr. Opin. Biotech. 2012, 23, 243–250. [Google Scholar] [CrossRef]
- Xiao, B.Z.; Chen, X.; Xiang, C.B.; Tang, N.; Zhang, Q.F.; Xiong, L.Z. Evaluation of Seven Function-Known Candidate Genes for their Effects on Improving Drought Resistance of Transgenic Rice under Field Conditions. Mol. Plant 2009, 2, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Hirayama, T.; Shinozaki, K. Research on plant abiotic stress responses in the post-genome era: Past, present and future. Plant J. 2010, 61, 1041–1052. [Google Scholar] [CrossRef]
- Hussain, S.S.; Kayani, M.A.; Amjad, M. Transcription Factors as Tools to Engineer Enhanced Drought Stress Tolerance in Plants. Biotechnol. Prog. 2011, 27, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.Z.; Li, X.L.; Jin, S.X.; Liu, X.Y.; Zhu, L.F.; Nie, Y.C.; Zhang, X.L. Overexpression of Rice NAC Gene SNAC1 Improves Drought and Salt Tolerance by Enhancing Root Development and Reducing Transpiration Rate in Transgenic Cotton. PLoS ONE 2014, 9, e86895. [Google Scholar] [CrossRef] [PubMed]
- Shen, G.X.; Wei, J.; Qiu, X.Y.; Hu, R.B.; Kuppu, S.; Auld, D.; Blumwald, E.; Gaxiola, R.; Payton, P.; Zhang, H. Co-overexpression of AVP1 and AtNHX1 in Cotton Further Improves Drought and Salt Tolerance in Transgenic Cotton Plants. Plant Mol. Biol. Rep. 2015, 33, 167–177. [Google Scholar] [CrossRef]
- Mishra, N.; Sun, L.; Zhu, X.L.; Smith, J.; Srivastava, A.P.; Yang, X.J.; Pehlivan, N.; Esmaeili, N.; Luo, H.; Shen, G.X.; et al. Overexpression of the Rice SUMO E3 Ligase Gene OsSIZ1 in Cotton Enhances Drought and Heat Tolerance, and Substantially Improves Fiber Yields in the Field under Reduced Irrigation and Rainfed Conditions. Plant Cell Physiol. 2017, 58, 735–746. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.L.; Zhang, D.Y.; Li, H.Y.; Dong, L.F.; Lan, H.Y. Ectopic overexpression of the aldehyde dehydrogenase ALDH21 from Syntrichia caninervis in tobacco confers salt and drought stress tolerance. Plant Physiol. Biochem. 2015, 95, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.L.; Zhang, D.Y.; Li, X.S.; Li, H.Y.; Zhang, D.W.; Lan, H.Y.; Wood, A.J.; Wang, J.C. Overexpression of ScALDH21 gene in cotton improves drought tolerance and growth in greenhouse and field conditions. Mol. Breed. 2016, 36, 34. [Google Scholar] [CrossRef]
- Yang, H.L.; Zhang, D.Y.; Wang, J.C.; Wood, A.J.; Zhang, Y.M. Molecular cloning of a stress-responsive aldehyde dehydrogenase gene ScALDH21 from the desiccation-tolerant moss Syntrichia caninervis and its responses to different stresses. Mol. Biol. Rep. 2012, 39, 2645–2652. [Google Scholar] [CrossRef]
- Yang, H.L.; Zhang, D.W.; Zhang, D.Y.; Bozorov, T.A.; Abdullaev, A.A.; Wood, A.J.; Wang, J.C.; Li, X.S.; Zhao, J.Y. Overexpression of ALDH21 from Syntrichia caninervis Moss in Upland Cotton Enhances Fiber Quality, Boll Component Traits, and Physiological Parameters during Deficit Irrigation. Crop Sci. 2019, 59, 553–564. [Google Scholar] [CrossRef] [Green Version]
- Mushtaq, S.; Moghaddasi, M. Evaluating the potentials of deficit irrigation as an adaptive response to climate change and environmental demand. Environ. Sci. Policy 2011, 14, 1139–1150. [Google Scholar] [CrossRef]
- Oweis, T.Y.; Farahani, H.J.; Hachum, A.Y. Evapotranspiration and water use of full and deficit irrigated cotton in the Mediterranean environment in northern Syria. Agric. Water Manag. 2011, 98, 1239–1248. [Google Scholar] [CrossRef]
- Falkenberg, N.R.; Piccinni, G.; Cothren, J.T.; Leskovar, D.I.; Rush, C.M. Remote sensing of biotic and abiotic stress for irrigation management of cotton. Agric. Water Manag. 2007, 87, 23–31. [Google Scholar] [CrossRef]
- DeTar, W.R. Yield and growth characteristics for cotton under various irrigation regimes in sandy soil. Agric. Water Manag. 2008, 95, 69–76. [Google Scholar] [CrossRef]
- Yang, C.J.; Luo, Y.; Sun, L.; Wu, N. Effect of Deficit Irrigation on the Growth, Water Use Characteristics and Yield of Cotton in Arid Northwest China. Pedosphere 2015, 25, 910–924. [Google Scholar] [CrossRef]
- Gui, D.W.; Lei, J.Q.; Zeng, F.J. Farmland management effects on the quality of surface soil during oasification in the southern rim of the Tarim Basin in Xinjiang, China. Plant Soil Environ. 2010, 56, 348–356. [Google Scholar] [CrossRef] [Green Version]
- Shareef, M.; Gui, D.W.; Zeng, F.J.; Waqas, M.; Zhang, B.; Iqbal, H. Water productivity, growth, and physiological assessment of deficit irrigated cotton on hyperarid desert-oases in northwest China. Agric. Water Manag. 2018, 206, 1–10. [Google Scholar] [CrossRef]
- Yang, Y.M.; Yang, Y.H.; Han, S.M.; Macadam, I.; Liu, D.L. Prediction of cotton yield and water demand under climate change and future adaptation measures. Agric. Water Manag. 2014, 144, 42–53. [Google Scholar] [CrossRef]
- Kuang, W.N.; Gao, X.P.; Gui, D.W.; Tenuta, M.; Flaten, D.N.; Yin, M.Y.; Zeng, F.J. Effects of fertilizer and irrigation management on nitrous oxide emission from cotton fields in an extremely arid region of northwestern China. Field Crop Res. 2018, 229, 17–26. [Google Scholar] [CrossRef]
- Zhong, R.S.; Tian, F.Q.; Yang, P.N.; Yi, Q.X. Planting and Irrigation Methods for Cotton in Southern Xinjiang, China. Irrig. Drain. 2016, 65, 461–468. [Google Scholar] [CrossRef]
- Chen, X.; Qi, Z.; Gui, D.; Gu, Z.; Ma, L.; Zeng, F.; Li, L. Simulating impacts of climate change on cotton yield and water requirement using RZWQM2. Agric. Water Manag. 2019, 222, 231–241. [Google Scholar] [CrossRef]
- Chen, X.; Qi, Z.; Gui, D.; Gu, Z.; Ma, L.; Zeng, F.; Li, L.; Sima, M.W. A Model-Based Real-Time Decision Support System for Irrigation Scheduling to Improve Water Productivity. Agronomy 2019, 9, 686. [Google Scholar] [CrossRef] [Green Version]
- Gu, Z.; Qi, Z.M.; Ma, L.W.; Gui, D.W.; Xu, J.Z.; Fang, Q.X.; Yuan, S.Q.; Feng, G. Development of an irrigation scheduling software based on model predicted crop water stress. Comput. Electron. Agric. 2017, 143, 208–221. [Google Scholar] [CrossRef]
- Upadhyay, R.K.; Gupta, A.; Ranjan, S.; Singh, R.; Pathre, U.V.; Nath, P.; Sane, A.P. The EAR Motif Controls the Early Flowering and Senescence Phenotype Mediated by Over-Expression of SlERF36 and Is Partly Responsible for Changes in Stomatal Density and Photosynthesis. PLoS ONE 2014, 9, e101995. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Qi, Z.; Gu, Z.; Gui, D.; Zeng, F. Optimizing Irrigation Rates for Cotton Production in an Extremely Arid Area Using Rzwqm2-Simulated Water Stress. Trans. Asabe 2017, 60, 2041–2052. [Google Scholar] [CrossRef] [Green Version]
- Bozorov, T.A.; Usmanov, R.M.; Yang, H.L.; Hamdullaev, S.A.; Musayev, S.; Shavkiev, J.; Nabiev, S.; Zhang, D.Y.; Abdullaev, A.A. Effect of water deficiency on relationships between metabolism, physiology, biomass, and yield of upland cotton (Gossypium hirsutum L.). J. Arid Land 2018, 10, 441–456. [Google Scholar] [CrossRef] [Green Version]
- Mao, D.L.; Lei, J.Q.; Zeng, F.J.; Zaynulla, R.; Wang, C.; Zhou, J. Characteristics of wind erosion and deposition in oasis-desert ecotone in southern margin of Tarim Basin, China. Chin. Geogr Sci. 2014, 24, 658–673. [Google Scholar] [CrossRef] [Green Version]
- Zeng, F. The countermeasures of suntainable development in Cele Oasis Ecosystem, Xinjiang. Arid Zone Res. 1999, 16, 6. [Google Scholar]
- Zeng, F.; Lei, J.; Zhang, X. Environmental Characteristics of Desert Transitional Area and Dominant Plants Adaptation; Science Press: Beijing, China, 2010. [Google Scholar]
- Hu, H.H.; Xiong, L.Z. Genetic Engineering and Breeding of Drought-Resistant Crops. Annu. Rev. Plant Biol. 2014, 65, 715–741. [Google Scholar] [CrossRef]
- Mittler, R.; Blumwald, E. Genetic Engineering for Modern Agriculture: Challenges and Perspectives. Annu. Rev. Plant Biol. 2010, 61, 443–462. [Google Scholar] [CrossRef] [Green Version]
- Basal, H.; Dagdelen, N.; Unay, A.; Yilmaz, E. Effects of Deficit Drip Irrigation Ratios on Cotton (Gossypium hirsutum L.) Yield and Fibre Quality. J. Agron. Crop Sci. 2009, 195, 19–29. [Google Scholar] [CrossRef]
- Zonta, J.H.; Brandao, Z.N.; Rodrigues, J.I.D.; Sofiatti, V. Cotton Response to Water Deficits at Different Growth Stages. Rev Caatinga 2017, 30, 980–990. [Google Scholar] [CrossRef] [Green Version]
- Gase, K.; Weinhold, A.; Bozorov, T.; Schuck, S.; Baldwin, I.T. Efficient screening of transgenic plant lines for ecological research. Mol. Ecol. Resour. 2011, 11, 890–902. [Google Scholar] [CrossRef] [PubMed]
- Schwachtje, J.; Kutschbach, S.; Baldwin, I.T. Reverse Genetics in Ecological Research. PLoS ONE 2008, 3, e1543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.Q.; He, C.X.; Wang, J.; Mao, Z.H.; Holaday, S.A.; Allen, R.D.; Zhang, H. Overexpression of the Arabidopsis 14-3-3 protein GF14 lambda in cotton leads to a “Stay-Green” phenotype and improves stress tolerance under moderate drought conditions. Plant Cell Physiol. 2004, 45, 1007–1014. [Google Scholar] [CrossRef] [PubMed]
- Ke, Y.; Yuan, M.; Liu, H.; Hui, S.; Qin, X.; Chen, J.; Zhang, Q.; Li, X.; Xiao, J.; Zhang, Q.; et al. The versatile functions of OsALDH2B1 provide a genic basis for growth–defense trade-offs in rice. Proc. Natl. Acad. Sci. USA 2020, 117, 3867–3873. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Gutteridge, J.M.C. Protection against oxidants in biological systems. In The Superoxide Theory of Oxygen Toxicity; Clarendon Press: Oxford, UK, 1989; pp. 86–187. [Google Scholar]
- Bolwell, G.P. Role of active oxygen species and NO in plant defence responses. Curr. Opin. Plant Biol. 1999, 2, 287–294. [Google Scholar] [CrossRef]
- Kotchoni, S.O.; Kuhns, C.; Ditzer, A.; Kirch, H.H.; Bartels, D. Over-expression of different aldehyde dehydrogenase genes in Arabidopsis thaliana confers tolerance to abiotic stress and protects plants against lipid peroxidation and oxidative stress. Plant Cell Environ. 2006, 29, 1033–1048. [Google Scholar] [CrossRef]
- Hozain, M.; Abdelmageed, H.; Lee, J.; Kang, M.; Fokar, M.; Allen, R.D.; Holaday, A.S. Expression of AtSAP5 in cotton up-regulates putative stress-responsive genes and improves the tolerance to rapidly developing water deficit and moderate heat stress. J. Plant Physiol. 2012, 169, 1261–1270. [Google Scholar] [CrossRef]
- Shi, Y.H.; Zhu, S.W.; Mao, X.Z.; Feng, J.X.; Qin, Y.M.; Zhang, L.; Cheng, J.; Wei, L.P.; Wang, Z.Y.; Zhu, Y.X. Transcriptome profiling, molecular biological, and physiological studies reveal a major role for ethylene in cotton fiber cell elongation. Plant Cell 2006, 18, 651–664. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| I | L | 2017 | 2018 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Seed Yield (kg ha−1) | Lint % | SCY (g Plant−1) | Lint Yield (kg ha−1) | Seed Yield (kg ha−1) | Lint % | SCY (g Plant−1) | Lint Yield (kg ha−1) | ||
| FFI | NT | 1610 ± 77e | 41.59 ± 0.19abc | 19.60 ± 0.48g | 1119 ± 53e | 1748 ± 141gh | 41.92 ± 0.88bc | 32.02 ± 1.66i | 1246 ± 84gh |
| L16 | 3221 ± 153a | 41.81 ± 0.14abc | 41.00 ± 1.00a | 2239 ± 107a | 3535 ± 52a | 43.94 ± 0.93abc | 47.72 ± 0.15c | 2478 ± 55a | |
| L38 | 1982 ± 94cd | 43.57 ± 0.17ab | 28.52 ± 0.70e | 1378 ± 66cd | 2674 ± 13c | 43.75 ± 1.54abc | 41.94 ± 0.03def | 1956 ± 25b | |
| FDI | NT | 1610 ± 77e | 41.04 ± 1.21abc | 24.23 ± 0.59f | 1119 ± 53e | 1765 ± 60gh | 40.07 ± 2.43c | 40.61 ± 0.29efg | 1262 ± 11gh |
| L16 | 2106 ± 100bc | 44.96 ± 0.35a | 28.10 ± 0.69e | 1464 ± 70bc | 2902 ± 53b | 42.61 ± 0.57bc | 56.69 ± 0.26a | 2021 ± 38b | |
| L38 | 2230 ± 106bc | 42.18 ± 0.58abc | 41.00 ± 1.00a | 1550 ± 74bc | 2403 ± 15cd | 41.19 ± 0.45c | 55.99 ± 0.37ab | 1468 ± 14f | |
| SFI | NT | 1239 ± 59fg | 41.25 ± 0.35abc | 24.12 ± 0.59f | 861 ± 41fg | 1579 ± 20h | 40.65 ± 1.34c | 27.54 ± 0.05j | 1114 ± 13h |
| L16 | 1735 ± 83de | 41.49 ± 0.97abc | 33.76 ± 0.82c | 1205 ± 57de | 2366 ± 84cd | 43.17 ± 2.20abc | 39.43 ± 0.54gh | 1624 ± 24cde | |
| L38 | 2354 ± 112b | 44.29 ± 2.04a | 40.57 ± 0.99a | 1636 ± 78b | 2293 ± 41d | 47.81 ± 2.24a | 46.55 ± 0.20c | 1606 ± 55def | |
| SDI | NT | 805 ± 38h | 39.47 ± 0.99c | 12.45 ± 0.30i | 560 ± 27h | 1004 ± 12i | 40.04 ± 0.65c | 18.49 ± 0.20l | 734 ± 15i |
| L16 | 1611 ± 77e | 41.75 ± 0.15abc | 25.63 ± 0.63f | 1119 ± 53e | 1625 ± 29gh | 39.21 ± 2.88c | 21.20 ± 0.02k | 1144 ± 9h | |
| L38 | 1487 ± 71ef | 41.70 ± 0.02abc | 31.14 ± 0.76d | 1033 ± 49ef | 1823 ± 49fg | 41.74 ± 1.43bc | 27.48 ± 0.59j | 1311 ± 44g | |
| EFI | NT | 991 ± 47gh | 39.70 ± 0.49bc | 16.57 ± 0.40h | 689 ± 74gh | 2430 ± 92cd | 39.37 ± 2.71c | 37.69 ± 0.97h | 1737 ± 45cd |
| L16 | 2230 ± 106bc | 43.09 ± 0.19abc | 36.53 ± 0.89b | 1550 ± 33bc | 2179 ± 25e | 42.05 ± 0.77bc | 43.21 ± 0.27d | 1591 ± 11ef | |
| L38 | 1982 ± 94cd | 43.79 ± 0.03a | 36.85 ± 0.90b | 1378 ± 66cd | 2552 ± 40c | 43.29 ± 0.38abc | 40.18 ± 0.20fg | 1916 ± 36b | |
| EDI | NT | 867 ± 47h | 39.60 ± 0.14bc | 16.49 ± 0.40h | 603 ± 29h | 1572 ± 179h | 38.73 ± 2.43c | 30.61 ± 2.38i | 1159 ± 110h |
| L16 | 1982 ± 94cd | 42.86 ± 0.51abc | 28.28 ± 0.69e | 1378 ± 66cd | 2196 ± 51de | 41.65 ± 1.75bc | 42.58 ± 0.34de | 1523 ± 26ef | |
| L38 | 991 ± 41g | 41.18 ± 1.28abc | 20.00 ± 0.49g | 689 ± 33gh | 2007 ± 10ef | 38.96 ± 1.92c | 53.82 ± 0.18b | 1769 ± 17c | |
| 2016 | 2017 | 2018 | Average ± SD | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Irrigation | L16 | L38 | Mean | L16 | L38 | Mean | L16 | L38 | Mean | |
| FFI | 100 | 23.0 | 61.5 | 95.6 | 51.6 | 73.6 | 18.0 | 57.2 | 37.6 | 57.0 ± 18.3 |
| FDI | 30.7 | 38.5 | 34.6 | 61. | 36.5 | 48.7 | 29.5 | 64.1 | 46.8 | 43.4 ± 7.7 |
| SFI | 40 | 90.0 | 65.0 | 49.8 | 45.7 | 47.7 | 59.1 | 39.1 | 49.1 | 54.0 ± 9.6 |
| SDI | 100 | 84.6 | 92.3 | 57.4 | 77.5 | 67.5 | 69.4 | 112.1 | 90.2 | 83.5 ± 14.0 |
| EFI | 125 | 100.0 | 112.5 | 19.1 | 13.1 | 16.1 | 74.8 | 57.7 | 66.3 | 65.0 ± 48.2 |
| EDI | 128.5 | 14.3 | 71.4 | 42.7 | 33.4 | 38.1 | 47.3 | 27.1 | 37.2 | 49.0 ± 19.5 |
| Average | 87.4 | 58.4 | 72.9 | 54.3 | 43.0 | 48.6 | 49.7 | 59.6 | 54.5 | 58.7 ± 12.9 |
| Source of Variation | F and P | Plant Height | Leaf Area | Photo-Synthesis | IWUE | Chloro-Phyll Content | Boll Number per Plant | Cotton Yield | WUE |
|---|---|---|---|---|---|---|---|---|---|
| Irrigation schedule | F | 40.204 | 29.167 | 4.246 | 34.191 | 22.495 | 5.229 | 15.555 | 22.829 |
| P | <0.001 | <0.001 | <0.05 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| Cotton lines | F | 52.508 | 25.101 | 10.391 | 4.193 | 39.762 | 12.780 | 15.591 | 19.122 |
| P | <0.001 | <0.001 | <0.001 | <0.05 | <0.001 | <0.001 | <0.001 | <0.001 | |
| Irrigation level | F | 11.564 | 2.234 | 1.296 | 3.153 | 21.138 | 4.434 | 7.918 | 0.464 |
| P | <0.001 | 0.137 | 0.256 | 0.077 | <0.001 | <0.05 | <0.001 | 0.497 | |
| IS × CL | F | 4.941 | 1.442 | 0.843 | 0.422 | 2.896 | 0.730 | 0.973 | 2.127 |
| P | <0.001 | 0.222 | 0.499 | 0.793 | <0.05 | 0.572 | 0.424 | 0.080 | |
| IS × IL | F | 13.117 | 9.702 | 1.088 | 4.534 | 4.190 | 11.155 | 6.976 | 4.243 |
| P | <0.001 | <0.001 | 0.338 | <0.05 | <0.05 | <0.001 | <0.001 | <0.05 | |
| CL × IL | F | 9.428 | 0.964 | 4.904 | 0.089 | 1.357 | 0.455 | 1.327 | 1.086 |
| P | <0.001 | 0.383 | <0.001 | 0.915 | 0.232 | 0.635 | 0.268 | 0.340 | |
| IS × CL × IL | F | 2.475 | 1.993 | 0.942 | 1.502 | 22.495 | 2.579 | 0.498 | 0.677 |
| P | <0.05 | 0.097 | 0.440 | 0.201 | <0.001 | <0.05 | 0.737 | 0.609 |
| IS | Line | Fiber Length (mm) | Fiber Uniformity (%) | Micronaire | Fiber Elongation (%) | Fiber Strength (cN. tex−1) | Line | Fiber Length (mm) | Fiber Uniformity (%) | Micronaire | Fiber Elongation (%) | Fiber Strength (cN. tex−1) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2017 | 2018 | |||||||||||
| FFI | NT | 30.3 ± 0.6cd | 86.6 ± 1.1abc | 5.1 ± 0.0ab | 6.5 ± 0.4abc | 26.75 ± 0.9c | NT | 28.2 ± 0.8 | 84.4 ± 0.9c | 4.9 ± 0.3 | 8.5 ± 0.1bc | 31.3 ± 1.6 |
| L16 | 32.8 ± 0.3a | 88.9 ± 0.0a | 5.3 ± 0.1a | 6.6 ± 0.1ab | 30.65 ± 0.5ab | L16 | 30.3 ± 0.6 | 86.5 ± 0.7ab | 4.9 ± 0.6 | 8.6 ± 0.3bc | 32.8 ± 1.4 | |
| L38 | 30.2 ± 0.0cd | 87.2 ± 1.4abc | 5.4 ± 0.2a | 5.9 ± 0.0bc | 29.50 ± 0.8ab | L38 | 28.6 ± 0.3 | 85.8 ± 0.2b | 5.4 ± 0.1 | 9.0 ± 0.1b | 30.3 ± 0.9 | |
| FDI | NT | 29.7 ± 0.3cd | 85.4 ± 0.0c | 5.2 ± 0.2a | 6.3 ± 0.4abc | 28.45 ± 0.5ab | NT | 27.3 ± 1.1 | 84.4 ± 0.5c | 5.0 ± 0.4 | 8.8 ± 0.2b | 30.6 ± 3.4 |
| L16 | 32.1 ± 0.2ab | 87.8 ± 0.2abc | 5.3 ± 0.2a | 6.4 ± 0.3abc | 30.70 ± 0.9ab | L16 | 29.5 ± 0.8 | 86.1 ± 0.6b | 5.1 ± 0.1 | 10.23 ± 1.0a | 30.6 ± 1.2 | |
| L38 | 31.0 ± 0.6bc | 87.9 ± 0.4ab | 5.0 ± 0.0ab | 6.3 ± 0.3abc | 29.50 ± 0.1ab | L38 | 27.7 ± 0.5 | 85.3 ± 0.1bc | 4.8 ± 0.1 | 10.1 ± 0.5a | 31.5 ± 0.9 | |
| SFI | NT | 30.0 ± 0.8cd | 86.3 ± 0.8bc | 4.6 ± 0.4ab | 5.8 ± 0.2bc | 29.45 ± 0.7ab | NT | 29.2 ± 1.1 | 85.6 ± 1.0bc | 5.3 ± 0.3 | 8.0 ± 0.1c | 29.9 ± 1.5 |
| L16 | 32.5 ± 0.2a | 88.1 ± 0.1ab | 4.8 ± 0.4ab | 6.3 ± 0.1abc | 33.20 ± 0.1a | L16 | 30.0 ± 0.3 | 86.2 ± 0.2b | 4.5 ± 0.2 | 10.2 ± 0.2a | 32.0 ± 1.0 | |
| L38 | 30.2 ± 0.3cd | 87.5 ± 1.1abc | 5.0 ± 0.1ab | 7.0 ± 0.0a | 30.30 ± 0.0ab | L38 | 28.4 ± 0.4 | 85.1 ± 0.7c | 5.4 ± 0.2 | 8.4 ± 0.5bc | 28.4 ± 0.3 | |
| SDI | NT | 29.0 ± 0.4d | 86.1 ± 0.0bc | 4.5 ± 0.0ab | 6.1 ± 0.1bc | 29.85 ± 2.2ab | NT | 29.0 ± 0.2 | 86.1 ± 0.0b | 4.5 ± 0.0 | 6.1 ± 0.9d | 29.8 ± 1.2 |
| L16 | 32.8 ± 0.1a | 87.9 ± 0.2ab | 4.3 ± 0.0ab | 6.3 ± 0.3abc | 34.05 ± 1.1 a | L16 | 29.6 ± 1.4 | 85.3 ± 0.5bc | 4.7 ± 0.4 | 8.7 ± 0.6b | 28.2 ± 0.8 | |
| L38 | 29.7 ± 0.3cd | 87.0 ± 1.1abc | 4.9 ± 0.0ab | 6.3 ± 0.1abc | 32.15 ± 1.8ab | L38 | 29.3 ± 0.6 | 86.2 ± 0.2b | 5.1 ± 0.1 | 8.9 ± 0.1b | 29.1 ± 0.8 | |
| EFI | NT | 29.7 ± 0.5cd | 87.6 ± 0.6abc | 4.3 ± 0.3ab | 6.1 ± 0.1bc | 30.10 ± 0.8ab | NT | 28.4 ± 0.7 | 85.1 ± 0.5c | 5.2 ± 0.2 | 9.4 ± 0.2b | 28.5 ± 0.5 |
| L16 | 32.8 ± 0.4a | 87.2 ± 0.9abc | 4.4 ± 0.4ab | 6.5 ± 0.3abc | 31.75 ± 1.2ab | L16 | 31.0 ± 0.5 | 87.7 ± 0.6a | 5.3 ± 0.0 | 9.0 ± 0.3b | 32.5 ± 0.9 | |
| L38 | 30.1 ± 0.3cd | 87.1 ± 0.4abc | 4.3 ± 0.0ab | 5.7 ± 0.1c | 29.10 ± 0.7ab | L38 | 29.0 ± 0.8 | 86.6 ± 0.6ab | 4.9 ± 0.4 | 9.2 ± 0.2b | 29.3 ± 0.9 | |
| EDI | NT | 30.6 ± 0.5c | 87.3 ± 0.6abc | 4.0 ± 0.0b | 5.7 ± 0.1c | 29.70 ± 1.0ab | NT | 29.2 ± 0.2 | 86.9 ± 0.3ab | 5.2 ± 0.3 | 9.7 ± 0.3ab | 32.1 ± 1.1 |
| L16 | 32.4 ± 0.4a | 87.4 ± 0.4abc | 4.4 ± 0.1ab | 6.3 ± 0.1abc | 31.35 ± 1.2ab | L16 | 30.3 ± 0.6 | 87.4 ± 0.2a | 4.3 ± 0.4 | 10.7 ± 0.2a | 30.6 ± 0.5 | |
| L38 | 30.7 ± 0.4c | 86.8 ± 0.6abc | 4.7 ± 0.2ab | 6.6 ± 0.3ab | 32.85 ± 0.3a | L38 | 28.3 ± 0.5 | 85.8 ± 0.6b | 5.4 ± 0.1 | 9.9 ± 0.2ab | 31.7 ± 1.5 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, H.; Bozorov, T.A.; Chen, X.; Zhang, D.; Wang, J.; Li, X.; Gui, D.; Qi, Z.; Zhang, D. Yield Comparisons between Cotton Variety Xin Nong Mian 1 and Its Transgenic ScALDH21 Lines under Different Water Deficiencies in a Desert-Oasis Ecotone. Agronomy 2021, 11, 1019. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11051019
Yang H, Bozorov TA, Chen X, Zhang D, Wang J, Li X, Gui D, Qi Z, Zhang D. Yield Comparisons between Cotton Variety Xin Nong Mian 1 and Its Transgenic ScALDH21 Lines under Different Water Deficiencies in a Desert-Oasis Ecotone. Agronomy. 2021; 11(5):1019. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11051019
Chicago/Turabian StyleYang, Honglan, Tohir A. Bozorov, Xiaoping Chen, Dawei Zhang, Jiancheng Wang, Xiaoshuang Li, Dongwei Gui, Zhiming Qi, and Daoyuan Zhang. 2021. "Yield Comparisons between Cotton Variety Xin Nong Mian 1 and Its Transgenic ScALDH21 Lines under Different Water Deficiencies in a Desert-Oasis Ecotone" Agronomy 11, no. 5: 1019. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11051019