
Climate Change Impacts on Cacao: Genotypic Variation in Responses of Mature Cacao to Elevated CO2 and Water Deficit



Abstract
:1. Introduction
2. Materials and Methods
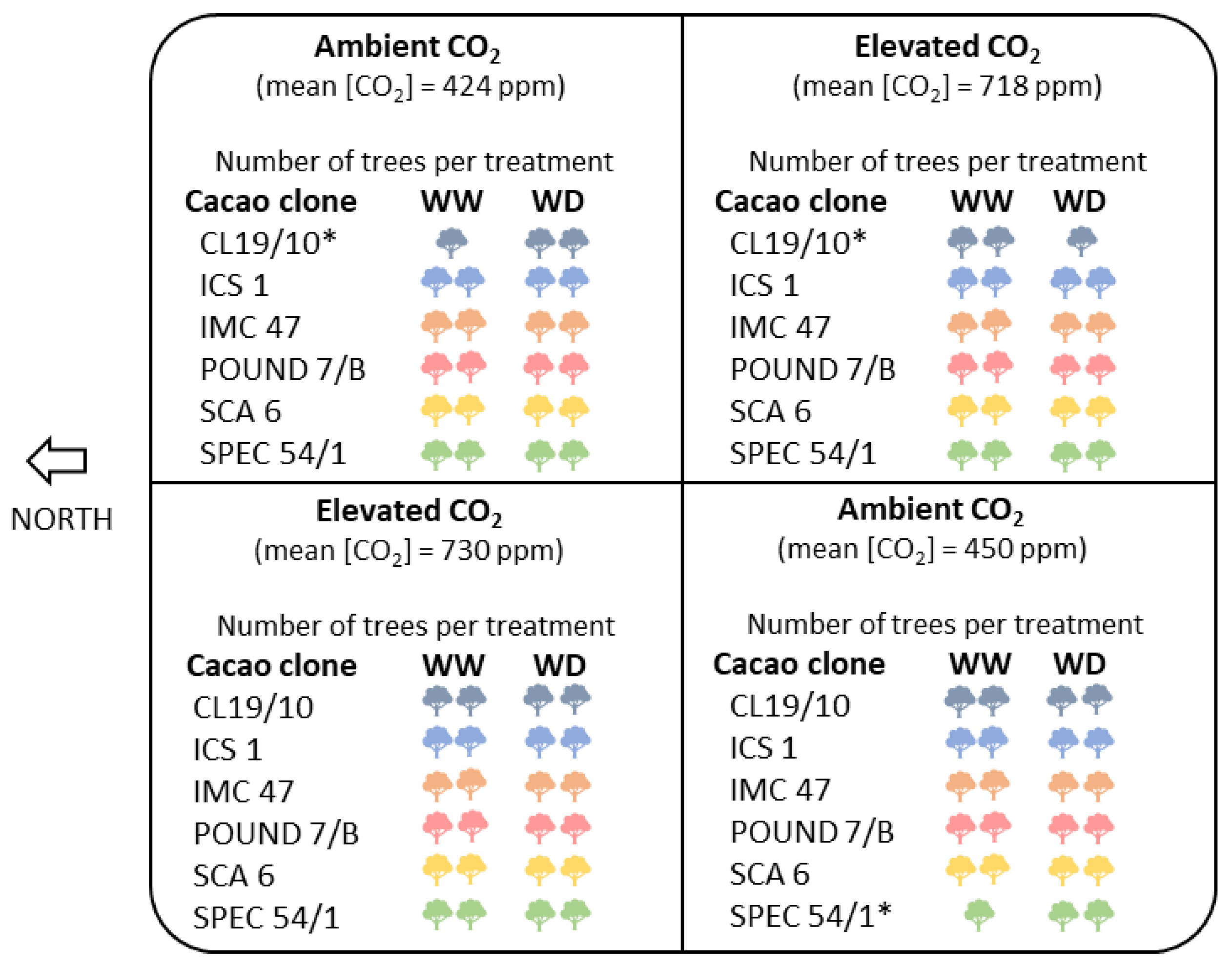
2.1. Plant Material and Growing Environment
2.2. Water Deficit Treatment
2.3. Photosynthetic Measurements
2.4. Growth Measurements
2.5. Canopy Photosynthesis Estimation
2.6. Final Biomass
2.7. Statistical Analyses
3. Results
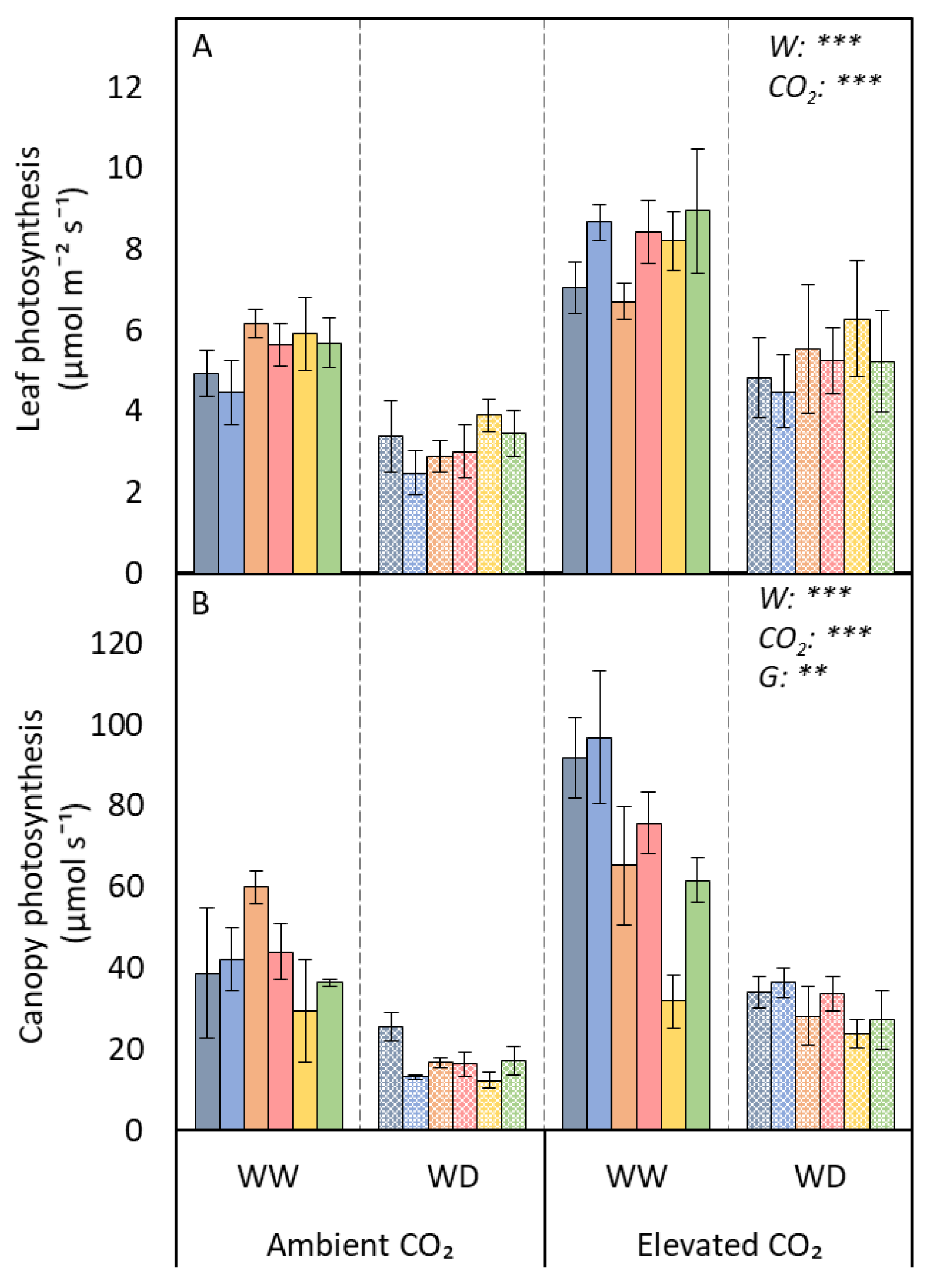
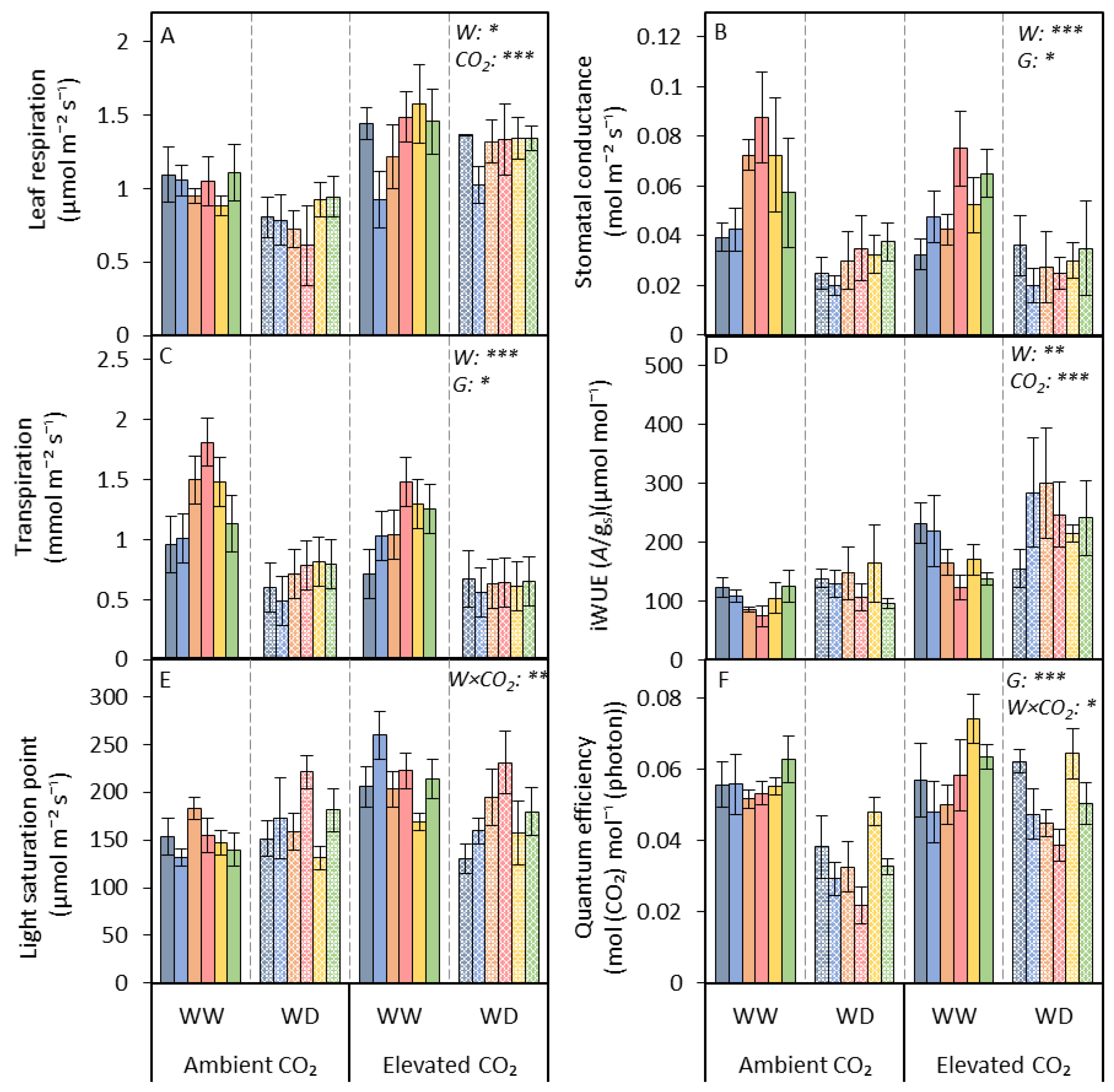
3.1. Photosynthetic Traits
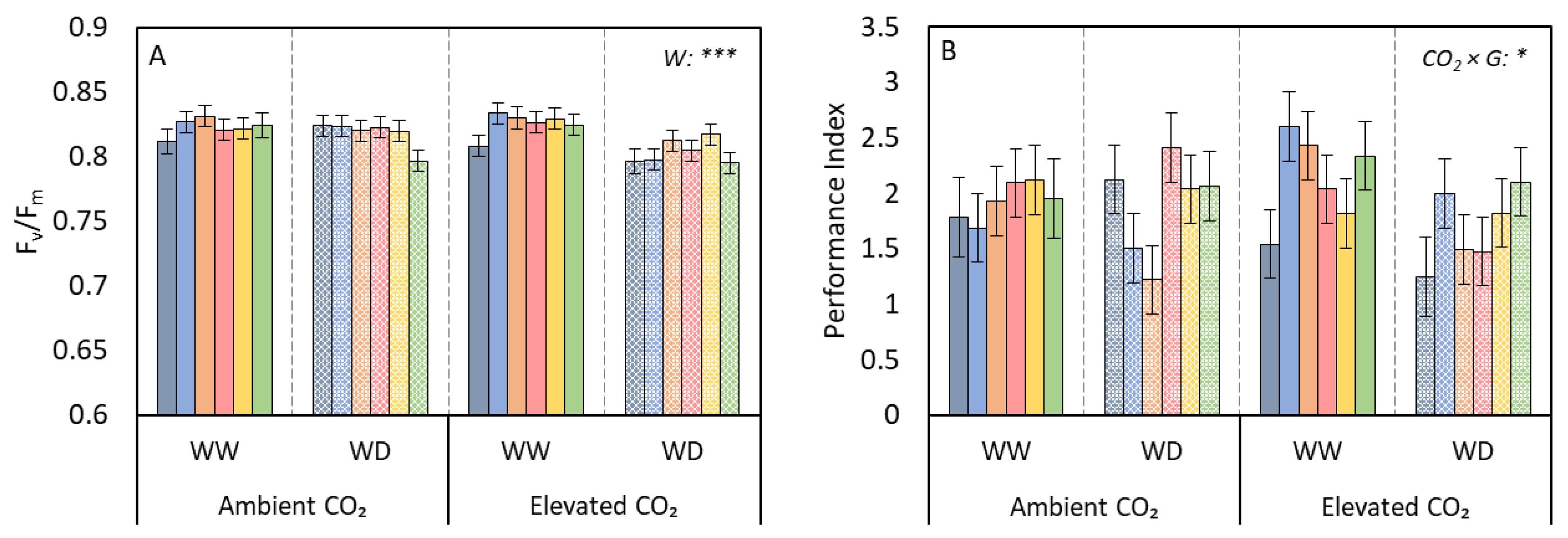
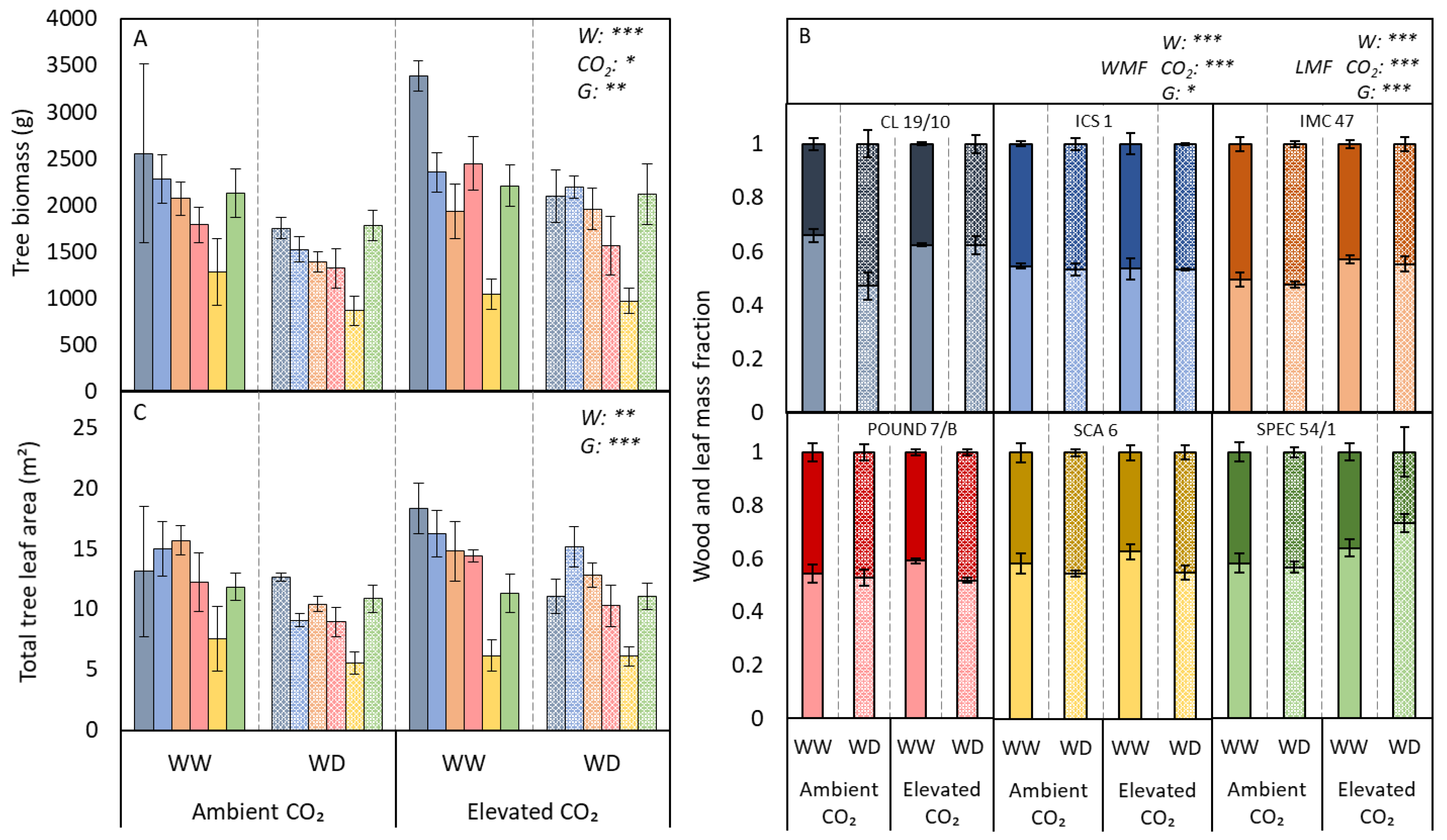
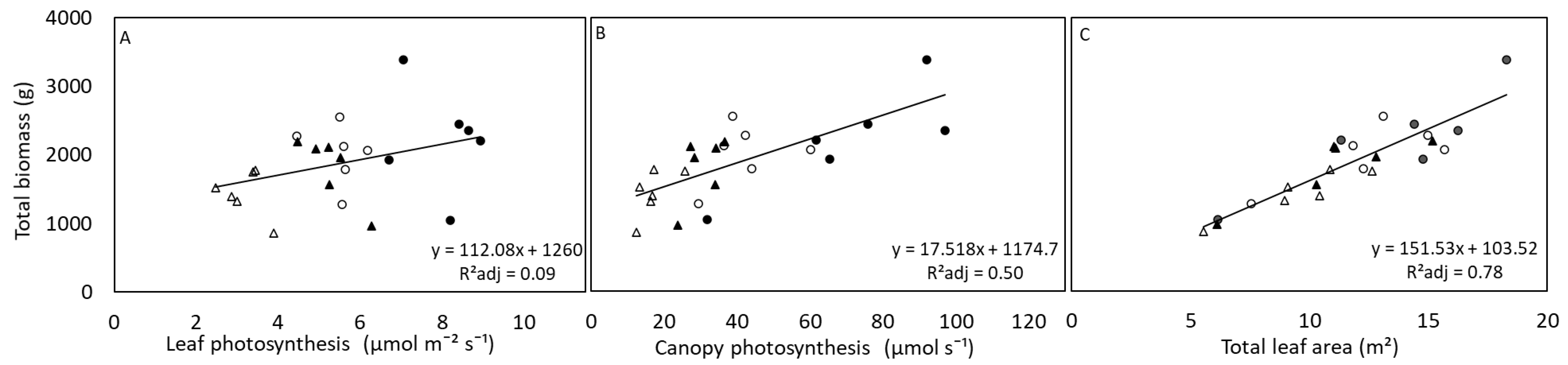
3.2. Growth and Leaf Traits
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sultan, B.; Gaetani, M. Agriculture in West Africa in the twenty-first century: Climate change and impacts scenarios, and potential for adaptation. Front. Plant Sci. 2016, 7, 1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biasutti, M. Forced Sahel rainfall trends in the CMIP5 archive. J. Geophys. Res. Atmos. 2013, 118, 1613–1623. [Google Scholar] [CrossRef] [Green Version]
- Dunning, C.M.; Black, E.; Allan, R.P. Later wet seasons with more intense rainfall over Africa under future climate change. J. Clim. 2018, 31, 9719–9738. [Google Scholar] [CrossRef] [Green Version]
- Gateau-Rey, L.; Tanner, E.V.J.; Rapidel, B.; Marelli, J.-P.; Royaert, S. Climate change could threaten cocoa production: Effects of 2015-16 El Niño-related drought on cocoa agroforests in Bahia, Brazil. PLoS ONE 2018, 13, e0200454. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, J.; Farquhar, G.D. Effects of rising temperatures and [CO2] on the physiology of tropical forest trees. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 1811–1817. [Google Scholar] [CrossRef] [Green Version]
- Zuidema, P.A.; Heinrich, I.; Rahman, M.; Vlam, M.; Zwartsenberg, S.A.; Sleen, P. Recent CO2 rise has modified the sensitivity of tropical tree growth to rainfall and temperature. Glob. Chang. Biol. 2020, 26, 4028–4041. [Google Scholar] [CrossRef]
- Lahive, F.; Hadley, P.; Daymond, A.J. The impact of elevated CO2 and water deficit stress on growth and photosynthesis of juvenile cacao (Theobroma cacao L.). Photosynthetica 2018, 56, 911–920. [Google Scholar] [CrossRef] [Green Version]
- Baligar, V.C.; Bunce, J.A.; Machado, R.C.R.; Elson, M.K. Photosynthetic photon flux density, carbon dioxide concentration, and vapor pressure deficit effects on photosynthesis in cacao seedlings. Photosynthetica 2008, 46, 216–221. [Google Scholar] [CrossRef]
- Baligar, V.C.; Elson, M.K.; Almeida, A.-A.F.; de Araujo, Q.R.; Ahnert, D.; He, Z. The impact of carbon dioxide concentrations and low to adequate photosynthetic photon flux density on growth, physiology and nutrient use efficiency of juvenile cacao genotypes. Agronomy 2021, 11, 397. [Google Scholar] [CrossRef]
- Avila, R.T.; de Almeida, W.L.; Costa, L.C.; Machado, K.L.G.; Barbosa, M.L.; de Souza, R.P.B.; Martino, P.B.; Juárez, M.A.T.; Marçal, D.M.S.; Martins, S.C.V.; et al. Elevated air [CO2] improves photosynthetic performance and alters biomass accumulation and partitioning in drought-stressed coffee plants. Environ. Exp. Bot. 2020, 177, 104137. [Google Scholar] [CrossRef]
- De Oliveira, M.F.; Marenco, R.A. Gas exchange, biomass allocation and water-use efficiency in response to elevated CO2 and drought in andiroba (Carapa surinamensis, Meliaceae). IForest 2019, 12, 61–68. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Rogers, A. The response of photosynthesis and stomatal conductance to rising [CO2]: Mechanisms and environmental interactions. Plant. Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef] [PubMed]
- Medlyn, B.E.; Barton, C.V.M.; Broadmeadow, M.S.J.; Ceulemans, R.; De Angelis, P.; Forstreuter, M.; Freeman, M.; Jackson, S.B.; Kellomäki, S.; Laitat, E.; et al. Stomatal conductance of forest species after long-term exposure to elevated CO2 concentration: A synthesis. New Phytol. 2001, 149, 247–264. [Google Scholar] [CrossRef]
- Avila, R.T.; Cardoso, A.A.; de Almeida, W.L.; Costa, L.C.; Machado, K.L.G.; Barbosa, M.L.; de Souza, R.P.B.; Oliveira, L.A.; Batista, D.S.; Martins, S.C.V.; et al. Coffee plants respond to drought and elevated [CO2] through changes in stomatal function, plant hydraulic conductance, and aquaporin expression. Environ. Exp. Bot. 2020, 177, 104148. [Google Scholar] [CrossRef]
- Daymond, A.J.; Tricker, P.J.; Hadley, P. Genotypic variation in photosynthesis in cacao is correlated with stomatal conductance and leaf nitrogen. Biol. Plant. 2011, 55, 99–104. [Google Scholar] [CrossRef]
- Daymond, A.J.; Hadley, P.; Machado, R.C.R.; Ng, E. Canopy characteristics of contrasting clones of cacao (Theobroma cacao). Exp. Agric. 2002, 38, 359–367. [Google Scholar] [CrossRef]
- Daymond, A.J.; Hadley, P.; Machado, R.C.R.; Ng, E. Genetic variability in partitioning to the yield component of cacao (Theobroma cacao L.). Hortscience 2002, 37, 799–801. [Google Scholar] [CrossRef] [Green Version]
- Daymond, A.J.; Hadley, P. The effects of temperature and light integral on early vegetative growth and chlorophyll fluorescence of four contrasting genotypes of cacao (Theobroma cacao). Ann. Appl. Biol. 2004, 145, 257–262. [Google Scholar] [CrossRef]
- Daymond, A.J.; Hadley, P. Differential effects of temperature on fruit development and bean quality of contrasting genotypes of cacao (Theobroma cacao). Ann. Appl. Biol. 2008, 2, 080527111818499. [Google Scholar] [CrossRef]
- De Almeida, A.A.F.; Brito, R.C.T.; Aguilar, M.A.G.; Valle, R.R. Some water relations aspects of Theobroma cacao clones. In Proceedings of the 13th International Cocoa Research Conference, Kota Kinabalu, Malaysia, 9–14 October 2000; pp. 349–363. [Google Scholar]
- Deng, X.; Joly, R.J.; Hahn, D.T. Effects of plant water deficit on the daily carbon balance of leaves of cacao seedlings. Physiol. Plant. 1989, 77, 407–412. [Google Scholar] [CrossRef]
- Balasimha, D.; Daniel, E.; Bhat, P.G. Influence of environmental factors on photosynthesis in cocoa trees. Agric. For. Meteorol. 1991, 55, 15–21. [Google Scholar] [CrossRef]
- Dos Santos, E.A.; de Almeida, A.-A.F.; da Silva Branco, M.C.; dos Santos, I.C.; Ahnert, D.; Baligar, V.C.; Valle, R.R. Path analysis of phenotypic traits in young cacao plants under drought conditions. PLoS ONE 2018, 13, e0191847. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, I.C.; de Almeida, A.-A.F.; Anhert, D.; da Conceição, A.S.; Pirovani, C.P.; Pires, J.L.; Valle, R.R.; Baligar, V.C. Molecular, physiological and biochemical responses of Theobroma cacao L. genotypes to soil water deficit. PLoS ONE 2014, 9, e115746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ávila-Lovera, E.; Coronel, I.; Jaimez, R.; Urich, R.; Pereyra, G.; Araque, O.; Chacón, I.; Tezara, W. Ecophysiological traits of adult trees of Criollo cocoa cultivars (Theobroma cacao L.) from a germplasm bank in Venezuela. Exp. Agric. 2016, 52, 137–153. [Google Scholar] [CrossRef]
- Ofori, A.; Padi, F.K.; Acheampong, K.; Lowor, S. Genetic variation and relationship of traits related to drought tolerance in cocoa (Theobroma cacao L.) under shade and no-shade conditions in Ghana. Euphytica 2015, 201, 411–421. [Google Scholar] [CrossRef]
- Farrell, A.D.; Rhiney, K.; Eitzinger, A.; Umaharan, P. Climate adaptation in a minor crop species: Is the cocoa breeding network prepared for climate change? Agroecol. Sustain. Food Syst. 2018, 42, 812–833. [Google Scholar] [CrossRef]
- Lahive, F.; Hadley, P.; Daymond, A.J. The physiological responses of cacao to the environment and the implications for climate change resilience. A review. Agron. Sustain. Dev. 2019, 39. [Google Scholar] [CrossRef] [Green Version]
- Lachenaud, P.; Paulin, D.; Ducamp, M.; Thevenin, J.-M. Twenty years of agronomic evaluation of wild cocoa trees (Theobroma cacao L.) from French Guiana. Sci. Hortic. 2007, 113, 313–321. [Google Scholar] [CrossRef]
- End, M.J. A Study of the Effects of the Photo-Thermal Environment on Fruit and Seed Growth and Development in Theobroma cacao L.; University of Reading: Reading, UK, 1990. [Google Scholar]
- Wood, G.A.R. Environment. In Cacao; Wood, G.A.R., Lass, R.A., Eds.; Longman Group Limited: London, UK; New York, NY, USA, 1985; pp. 38–79. ISBN 9780470698983. [Google Scholar]
- Prioul, J.L.; Chartier, P. Partitioning of transfer and carboxylation components of intracellularresistance to photosynthetic CO₂ fixation: A critical analysis of the methods used. Ann. Bot. 1977, 41, 789–800. [Google Scholar] [CrossRef]
- Salisbury, E.J. On the causes and ecological significance of stomatal frequency, with special reference to woodland flora. Philos. Trans. R. Soc. B 1927, 216, 1–65. [Google Scholar]
- Fleischer, K.; Rammig, A.; De Kauwe, M.G.; Walker, A.P.; Domingues, T.F.; Fuchslueger, L.; Garcia, S.; Goll, D.S.; Grandis, A.; Jiang, M.; et al. Amazon forest response to CO2 fertilization dependent on plant phosphorus acquisition. Nat. Geosci. 2019, 12, 736–741. [Google Scholar] [CrossRef]
- Centritto, M.; Loreto, F.; Chartzoulakis, K. The use of low [CO2] to estimate diffusional and non-diffusional limitations of photosynthetic capacity of salt-stressed olive saplings. Plant Cell Environ. 2003, 26, 585–594. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Loreto, F.; Cornic, G.; Sharkey, T.D. Diffusive and metabolic limitations to photosynthesis under drought and salinity in C3 plants. Plant Biol. 2004, 6, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Flexas, J.; Ribas-Carbó, M.; Diaz-Espejo, A.; Galmés, J.; Medrano, H. Mesophyll conductance to CO2: Current knowledge and future prospects. Plant. Cell Environ. 2008, 31, 602–621. [Google Scholar] [CrossRef] [PubMed]
- Parkhurst, D.F. Diffusion of CO2 and other gases inside leaves. New Phytol. 1994, 126, 449–479. [Google Scholar] [CrossRef] [PubMed]
- Alvim, P.D.T. Cacao. In Ecophysiology of Tropical Crops; De Alvim, P.T., Kozlowski, T.T., Eds.; Academy Press: New York, NY, USA, 1977; pp. 279–313. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Ambient CO2 | Elevated CO2 | |||
|---|---|---|---|---|---|
| Well-Watered | Water Deficit | Well-Watered | Water Deficit | ||
| CL 19/10 | Stomatal density (stomata mm−2) | 840.60 (±58.93) | 930.20 (±50.94) | 798.00 (±50.94) | 724.20 (±58.93) |
| Stomatal index (%) | 14.29 (±0.68) | 14.87 (±0.59) | 15.27 (±0.59) | 14.16 (±0.68) | |
| Flush interval (days) | 29 (n =1) | 76 (±18) | 49 (±26) | 52 (±15) | |
| ICS 1 | Stomatal density (stomata mm−2) | 798.20 (±50.94) | 742.1 (±50.94) | 858.1 (±50.94) | 760.2 (±50.94) |
| Stomatal index (%) | 12.27 (±0.59) | 12.7 (±0.59) | 13.11 (±13.11) | 12.84 (±0.59) | |
| Flush interval (days) | 26 (±2) | 55 (±21) | 43 (±2) | 77 (±10) | |
| IMC 47 | Stomatal density (stomata mm−2) | 910.6 (±50.94) | 965.2 (±50.94) | 856.1 (±50.94) | 942.6 (±50.94) |
| Stomatal index (%) | 15.51 (±0.59) | 14.91 (±0.59) | 15.01 (±0.59) | 16.31 (±0.59) | |
| Flush interval (days) | 38 (±3) | 51 (±4) | 34 (±9) | 43 (±6) | |
| POUND 7/B | Stomatal density (stomata mm−2) | 919.3 (±50.94) | 967.5 (±50.94) | 911.5 (±50.94) | 1051.0 (±50.94) |
| Stomatal index (%) | 15.17 (±0.59) | 15.24 (±0.59) | 15.63 (±0.59) | 16.8 (±0.59) | |
| Flush interval (days) | 58 (±8) | 63 (±6) | 56 (±15) | 69 (±15) | |
| SCA 6 | Stomatal density (stomata mm−2) | 911.4 (±50.94) | 942.6 (±50.94) | 835.8 (±50.94) | 870.5 (±50.94) |
| Stomatal index (%) | 15.7 (±0.59) | 15.54 (±0.59) | 15.11 (±0.59) | 15.02 (±0.59) | |
| Flush interval (days) | 42 (±6) | 48 (±11) | 37 (±9) | 40 (±10) | |
| SPEC 54/1 | Stomatal density (stomata mm−2) | 714.7 (±58.94) | 835.3 (±50.94) | 741.1 (±50.94) | 847.2 (±50.94) |
| Stomatal index (%) | 12.04 (±0.68) | 13.59 (±0.59) | 12.72 (±0.59) | 13.63 (±0.59) | |
| Flush interval (days) | 44 (±6) | 63 (±6) | 53 (±12) | 60 (±10) | |
| Water treatment | CO2 treatment | Genotype | Interaction | ||
| Stomatal density | ns | ns | *** | ns | |
| Stomatal index | ns | ns | *** | ns | |
| Flush Interval | *** | ns | ns | ns | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lahive, F.; Handley, L.R.; Hadley, P.; Daymond, A.J. Climate Change Impacts on Cacao: Genotypic Variation in Responses of Mature Cacao to Elevated CO2 and Water Deficit. Agronomy 2021, 11, 818. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11050818
Lahive F, Handley LR, Hadley P, Daymond AJ. Climate Change Impacts on Cacao: Genotypic Variation in Responses of Mature Cacao to Elevated CO2 and Water Deficit. Agronomy. 2021; 11(5):818. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11050818
Chicago/Turabian StyleLahive, Fiona, Liam R. Handley, Paul Hadley, and Andrew J. Daymond. 2021. "Climate Change Impacts on Cacao: Genotypic Variation in Responses of Mature Cacao to Elevated CO2 and Water Deficit" Agronomy 11, no. 5: 818. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11050818