
Diversity of Cowpea [Vigna unguiculata (L.) Walp] Landraces in Mozambique: New Opportunities for Crop Improvement and Future Breeding Programs

 ,
,  , , ,
, , ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Morphological Characterization
2.2. Data Analysis
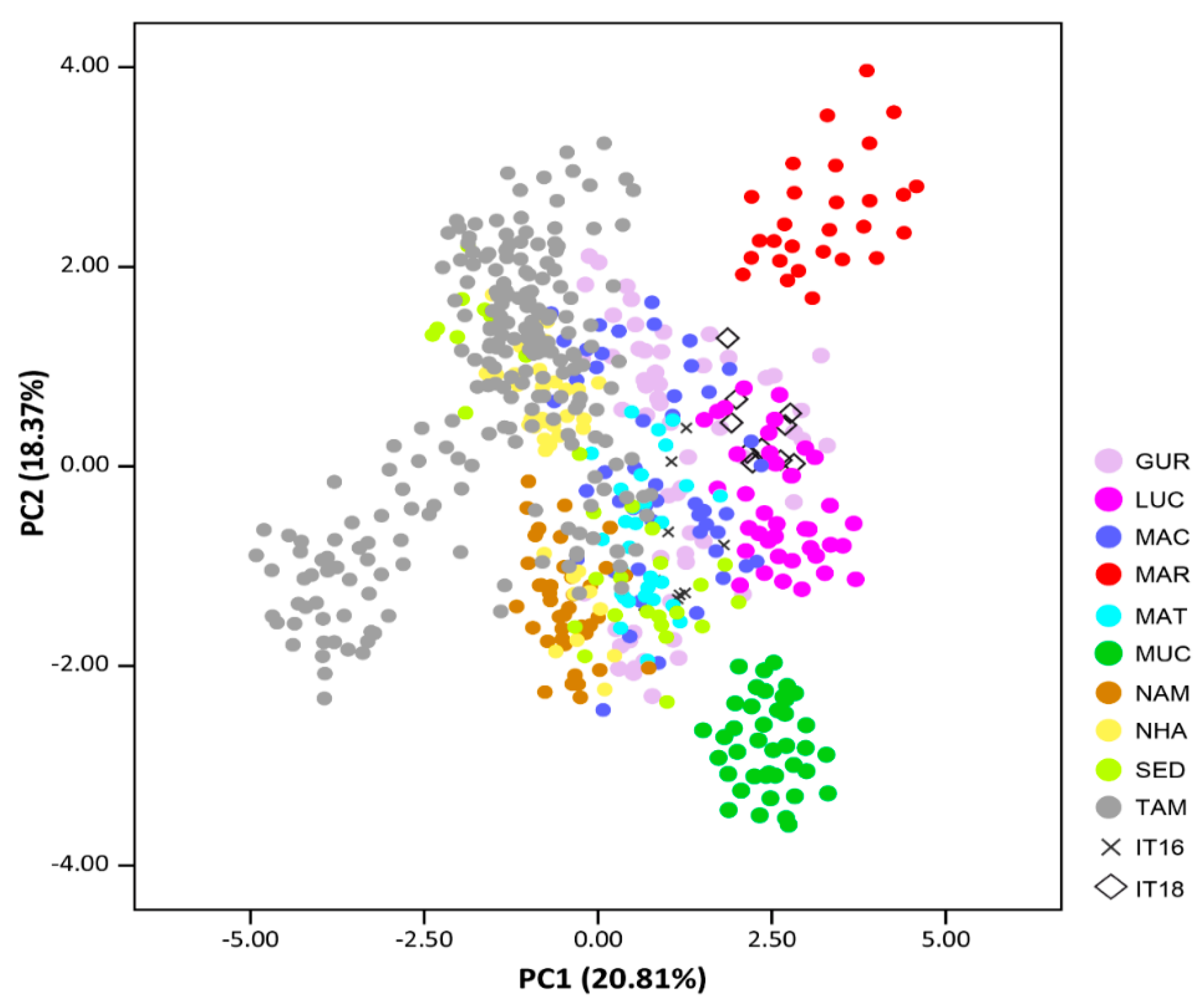
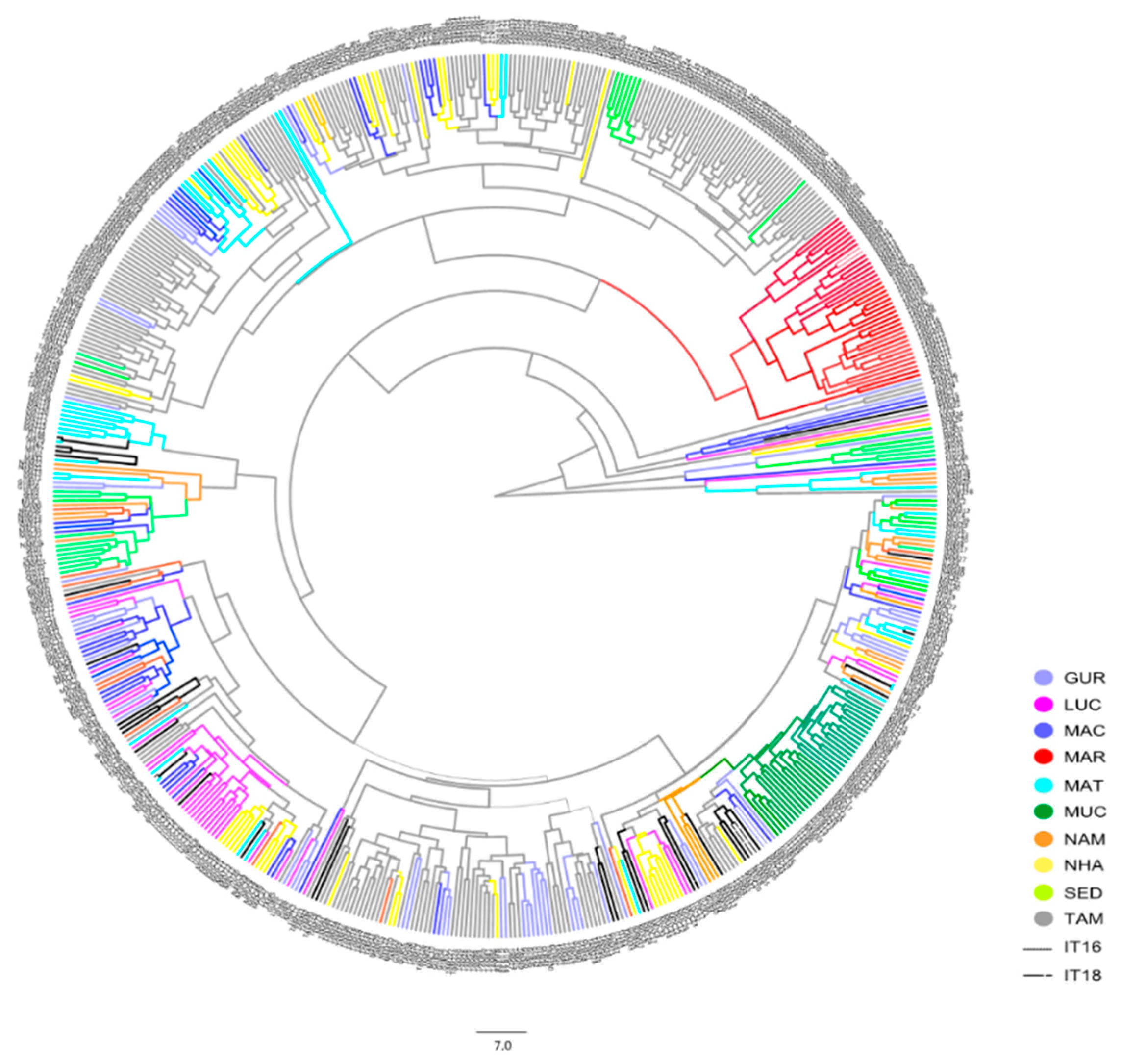
3. Results
4. Discussion
4.1. Importance of Morphological Traits in Cowpea Landraces
4.2. Diversity in Landraces and Cowpea Commercial Varieties
4.3. Guidelines for Future Breeding Programs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Boukar, O.; Belko, N.; Chamarthi, S.; Togola, A.; Batieno, J.; Owusu, E.; Haruna, M.; Diallo, S.; Umar, M.L.; Olufajo, O.; et al. Cowpea (Vigna unguiculata): Genetics, genomics and breeding. Plant Breed. 2019, 138, 415–424. [Google Scholar] [CrossRef] [Green Version]
- Chivenge, P.; Mabhaudhi, T.; Modi, A.; Mafongoya, P. The Potential Role of Neglected and Underutilised Crop Species as Future Crops under Water Scarce Conditions in Sub-Saharan Africa. Int. J. Environ. Res. Public Health 2015, 12, 5685–5711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bado, B.V.; Bationto, A.; Cescas, M. Assessment of cowpea and groundnut contributions to soil fertility and succeeding sorghum yields in the Guinean savannah zone of Burkina Faso (West Africa). Biol. Fertil. Soils 2006, 43, 71–176. [Google Scholar] [CrossRef]
- Enyiukwu, D.N.; Amadioha, A.C.; Ononuju, C.C. Nutritional Significance of Cowpea Leaves for Human Consumption. Greener Trends Food Sci. Nutr. 2018, 1, 1–10. [Google Scholar] [CrossRef]
- Jayathilake, C.; Visvanathan, R.; Deen, A.; Bangamuwage, R.; Jayawardana, B.C.; Nammi, S.; Liyanage, R. Cowpea: An overview on its nutritional facts and health benefits. J. Sci. Food Agric. 2018, 98, 4793–4806. [Google Scholar] [CrossRef]
- Kirigia, D.; Winkelmann, T.; Kasili, R.; Mibus, H. Development stage, storage temperature and storage duration influence phytonutrient content in cowpea (Vigna unguiculata L. Walp.). Heliyon 2018, 4. [Google Scholar] [CrossRef]
- Owade, J.O.; Abong, G.O.; Okoth, M.W.; Mwang’ombe, A.W. Trends and constraints in the production and utilization of cowpea leaves in the arid and semi-arid lands of Kenya. Open Agric. 2020, 5, 325–334. [Google Scholar] [CrossRef]
- Gomes, A.M.F.; Nhantumbo, N.; Ferreira-Pinto, M.; Massinga, R.; Ramalho, J.C.; Ribeiro-Barros, A. Breeding Elite Cowpea [Vigna unguiculata (L.) Walp] Varieties for Improved Food Security and Income in Africa: Opportunities and Challenges. In Lagume Crops; El-Esawi, M.A., Ed.; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef] [Green Version]
- Hall, A. Breeding for adaptation to drought and heat in cowpea. Eur. J. Agron. 2004, 21, 447–454. [Google Scholar] [CrossRef]
- Horn, L.; Shimelis, H.; Laing, M. Participatory appraisal of production constraints, preferred traits and farming system of cowpea in the northern Namibia: Implications for breeding. Legume Res. Int. J. 2015, 38, 691–700. [Google Scholar] [CrossRef] [Green Version]
- Horn, L.N.; Shimelis, H. Production constraints and breeding approaches for cowpea improvement for drought prone agro-ecologies in Sub-Saharan Africa. Ann. Agric. Sci. 2020, 65, 83–91. [Google Scholar] [CrossRef]
- FAO. Cowpea: Post-Harvest Operations; FAO: Italy, Rome, 2004; Available online: http://www.fao.org/3/a-au994e.pdf (accessed on 3 November 2019).
- Timko, M.P.; Singh, B.B. Cowpea, a Multifunctional Legume. In Genomics of Tropical Crop Plants; Moore, P.H., Ming, R., Eds.; Springer: New York, NY, USA, 2008; pp. 227–258. [Google Scholar] [CrossRef]
- Xiong, H.; Shi, A.; Mou, B.; Qin, J.; Motes, D.; Lu, W.; Ma, J.; Weng, Y.; Yang, W.; Wu, D. Genetic Diversity and Population Structure of Cowpea (Vigna unguiculata L. Walp). PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [Green Version]
- Ehlers, J.D.; Hall, A.E. Cowpea (Vigna unguiculata L. Walp.). Field Crops Res. 1997, 53, 187–204. [Google Scholar] [CrossRef]
- Kouam, E.B.; Pasquet, R.S.; Campagne, P.; Tignegre, J.-B.; Thoen, K.; Gaudin, R.; Ouedraogo, J.T.; Salifu, A.B.; Muluvi, G.M.; Gepts, P. Genetic structure and mating system of wild cowpea populations in West Africa. BMC Plant Biol. 2012, 12, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, A.M.F.; Draper, D.; Talhinhas, P.; Santos, P.B.; Simões, F.; Nhantumbo, N.; Massinga, R.; Ramalho, J.C.; Marques, I.; Ribeiro-Barros, A.I. Genetic Diversity among Cowpea (Vigna unguiculata (L.) Walp.) Landraces Suggests Central Mozambique as an Important Hotspot of Variation. Agronomy 2020, 10, 1893. [Google Scholar] [CrossRef]
- Gomes, A.M.F.; Rodrigues, A.P.; António, C.; Rodrigues, A.M.; Leitão, A.E.; Batista-Santos, P.; Nhantumbo, N.; Massinga, R.; Ribeiro-Barros, A.I.; Ramalho, J.C. Drought response of cowpea (Vigna unguiculata (L.) Walp.) landraces at leaf physiological and metabolite profile levels. Environ. Exp. Bot. 2020, 175, 104060. [Google Scholar] [CrossRef]
- Ficiciyan, A.; Loos, J.; Sievers-Glotzbach, S.; Tscharntke, T. More than Yield: Ecosystem Services of Traditional versus Modern Crop Varieties Revisited. Sustainability 2018, 10, 2834. [Google Scholar] [CrossRef] [Green Version]
- International Board for Plant Genetic Resources. Cowpea Descriptors; IBPGR: Rome, Italy, 1983. [Google Scholar]
- IBM Corp. IBM SPSS Statistics for Windows; Version 26.0; IBM Corp: Armonk, NY, USA, 2019. [Google Scholar]
- Menssen, M.; Linde, M.; Otunga Omondi, E.; Abukutsa-Onyango, M.; Dinssa, F.F.; Winkelmann, T. Genetic and morphological diversity of cowpea (Vigna unguiculata (L.) Walp.) entries from East Africa. Sci. Hortic. 2017, 226, 268–276. [Google Scholar] [CrossRef]
- Lee, O.N.; Park, H.Y. Assessment of genetic diversity in cultivated radishes (Raphanus sativus) by agronomic traits and SSR markers. Sci. Hortic. 2017, 223, 19–30. [Google Scholar] [CrossRef]
- Arteaga, S.; Yabor, L.; Torres, J.; Solbes, E.; Muñoz, E.; Díez, M.J.; Vicente, O.; Boscaiu, M. Morphological and Agronomic Characterization of Spanish Landraces of Phaseolus vulgaris L. Agriculture 2019, 9, 149. [Google Scholar] [CrossRef] [Green Version]
- Ouaja, M.; Bahri, B.A.; Aouini, L.; Ferjaoui, S.; Medini, M.; Marcel, T.C.; Hamza, S. Morphological characterization and genetic diversity analysis of Tunisian durum wheat (Triticum turgidum var. durum) accessions. BMC Genom. Data 2021, 22, 3. [Google Scholar] [CrossRef]
- Mafakheri, K.; Bihamta, M.R.; Abbasi, A.R. Assessment of genetic diversity in cowpea (Vigna unguiculata L.) germplasm using morphological and molecular characterisation. Cogent Food Agric. 2017, 3. [Google Scholar] [CrossRef]
- Njonjo, M.W.; Muthomi, J.W.; Mwang’ombe, A.W. Production Practices, Postharvest Handling, and Quality of Cowpea Seed Used by Farmers in Makueni and Taita Taveta Counties in Kenya. Int. J. Agron. 2019, 2019, 1607535. [Google Scholar] [CrossRef] [Green Version]
- Doumbia, I.Z.; Akromah, R.; Asibuo, J.Y. Comparative study of cowpea germplasm diversity from Ghana and Mali using morrphological characteristics. J. Plant Breed. Genet. 2013, 1, 139–147. [Google Scholar]
- Cobbinah, F.A.; Addo-Quaye, A.A.; Asante, I.K. Characterization, evaluation and selection of cowpea (Vigna unguiculata (L.) walp) accessions with desirable traits from eight regions of Ghana. J. Agric. Biol. Sci. 2011, 6, 21–32. [Google Scholar]
- Vafaee, Y.; Ghaderi, N.; Khadivi, A. Morphological variation and marker-fruit trait associations in a collection of grape (Vitis vinifera L.). Sci. Hortic. 2017, 225, 771–782. [Google Scholar] [CrossRef]
- Wani, A.A.; Zargar, S.A.; Malik, A.H.; Kashtwari, M.; Nazir, M.; Khuroo, A.A.; Ahmad, F.; Dar, T.A. Assessment of variability in morphological characters of apricot germplasm of Kashmir, India. Sci. Hortic. 2017, 225, 630–637. [Google Scholar] [CrossRef]
- Ghalmi, N.; Malice, M.; Jacquemin, J.-M.; Ounane, S.; Mekliche, L.; Baudoin, J.-P. Morphological and molecular diversity within Algerian cowpea (Vigna unguiculata (L.) Walp.) landraces. Genet. Resour. Crop Evol. 2009, 57, 371–386. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.-C.; Wu, Y.-Q.; Ma, L.; Chen, Y.-J.; Ji, X. Genetic and morphological divergence among three closely related Phrynocephalus species (Agamidae). BMC Evol. Biol. 2019, 19, 114. [Google Scholar] [CrossRef] [Green Version]
- Lam, D.T.; Ichitani, K.; Henry, R.J.; Ishikawa, R. Molecular and Morphological Divergence of Australian Wild Rice. Plants 2020, 9, 224. [Google Scholar] [CrossRef] [Green Version]
- Galindo, J.; Carvalho, J.; Sotelo, G.; Duvetorp, M.; Costa, D.; Kemppainen, P.; Panova, M.; Kaliontzopoulou, A.; Johannesson, K.; Faria, R. Genetic and morphological divergence between Littorina fabalis ecotypes in Northern Europe. J. Evol. Biol. 2021, 34, 97–113. [Google Scholar] [CrossRef]
- Fatokun, C.; Girma, G.; Abberton, M.; Gedil, M.; Unachukwu, N.; Oyatomi, O.; Yusuf, M.; Rabbi, I.; Boukar, O. Genetic diversity and population structure of a mini-core subset from the world cowpea (Vigna unguiculata (L.) Walp.) germplasm collection. Sci. Rep. 2018, 8, 16035. [Google Scholar] [CrossRef] [Green Version]
- Huynh, B.-L.; Close, T.J.; Roberts, P.A.; Hu, Z.; Wanamaker, S.; Lucas, M.R.; Chiulele, R.; Cissé, N.; David, A.; Hearne, S.; et al. Gene Pools and the Genetic Architecture of Domesticated Cowpea. Plant Genome 2013, 6. [Google Scholar] [CrossRef] [Green Version]
- FAO/IPGRI. Review and Development of Indicators for Genetic Diversity, Genetic Erosion and Genetic Vulnerability (GDEV): Summary Report of a Joint FAO/IPGRI. Workshop; FAO/IPGRI: Rome, Italy, 2002. [Google Scholar]
- Weng, Y.; Ravelombola, W.; Yang, W.; Qin, J.; Zhou, W.; Wang, Y.; Mou, B.; Shi, A. Screening of Seed Soluble Sugar Content in Cowpea (Vigna unguiculata (L.) Walp). Am. J. Plant Sci. 2018, 9, 1455–1466. [Google Scholar] [CrossRef] [Green Version]
- Santos, A.D.; Ceccon, G.; Davide, L.M.C.; Correa, A.M.; Alves, V.B. Correlations and path analysis of yield components in cowpea. Crop Breed. Appl. Biotechnol. 2014, 14, 82–87. [Google Scholar] [CrossRef] [Green Version]
- Udensi, O.; Ikpeme, E.V.; Edu, E.A.; Ekpe, D.E. Relationship studies in cowpea (Vigna unguiculata L. Walp) landraces grown under humid lowland condition. Int. J. Agric. Res. 2012, 7, 33–45. [Google Scholar] [CrossRef]
- Iseki, K.; Olaleye, O.; Ishikawa, H. Intra-plant variation in seed weight and seed protein content of cowpea. Plant Prod. Sci. 2020, 23, 103–113. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
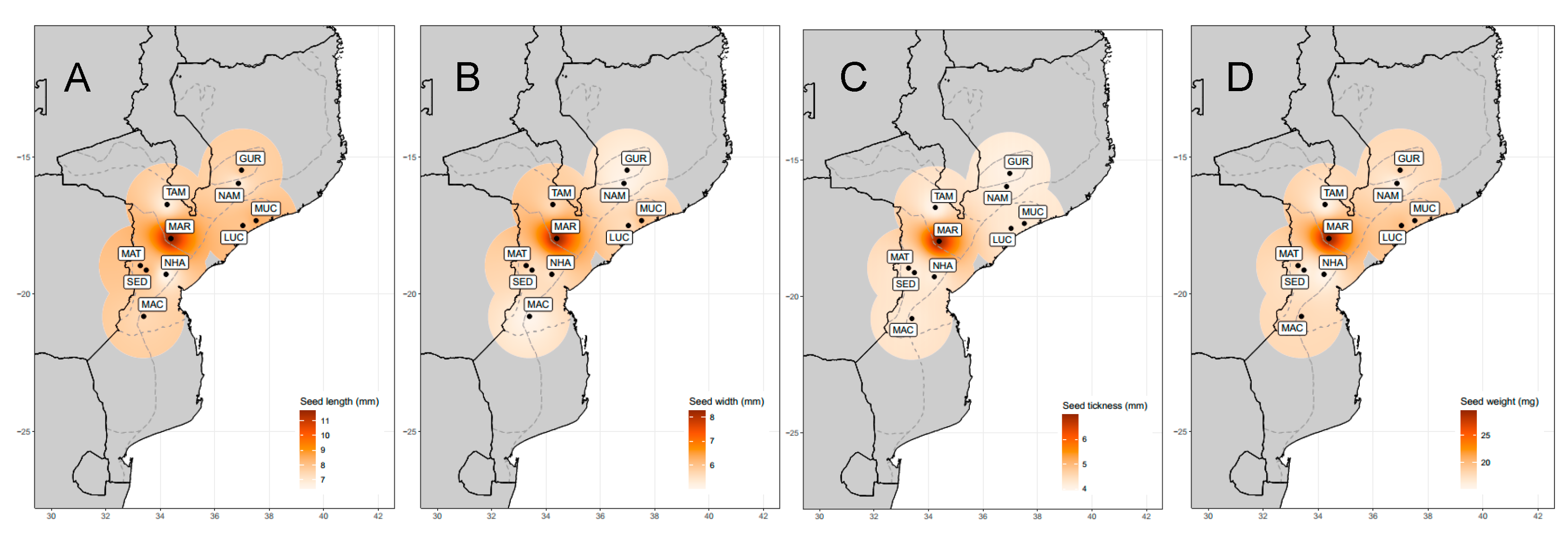
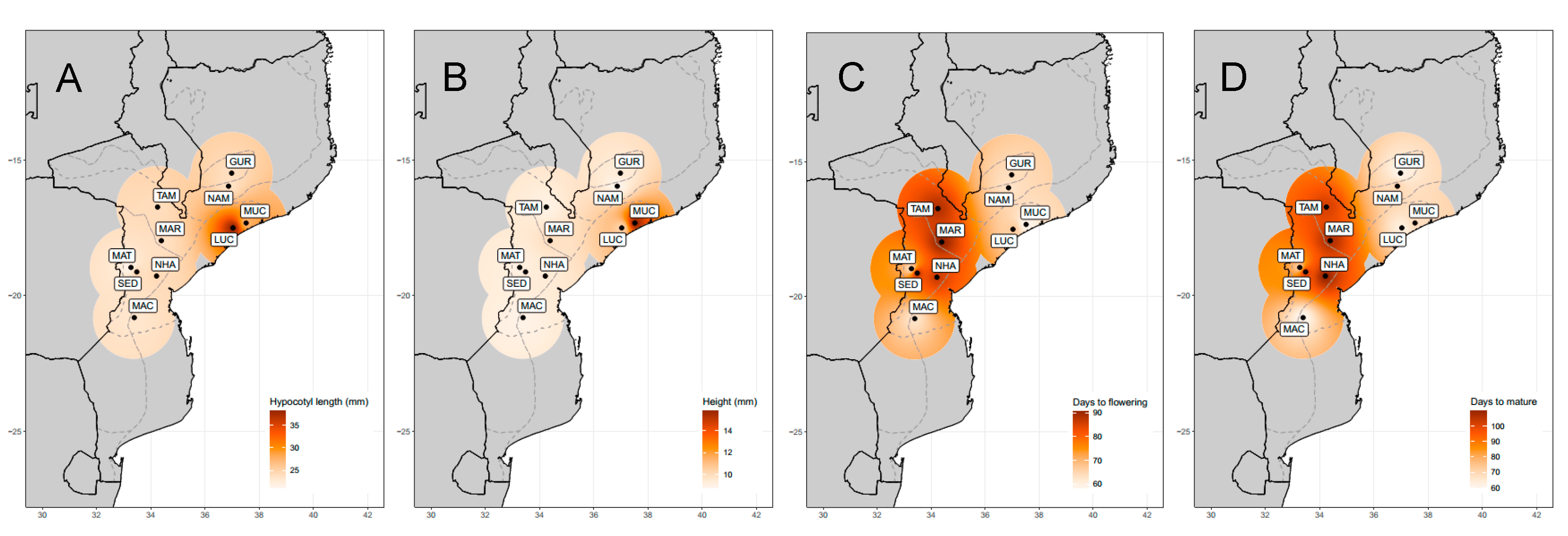
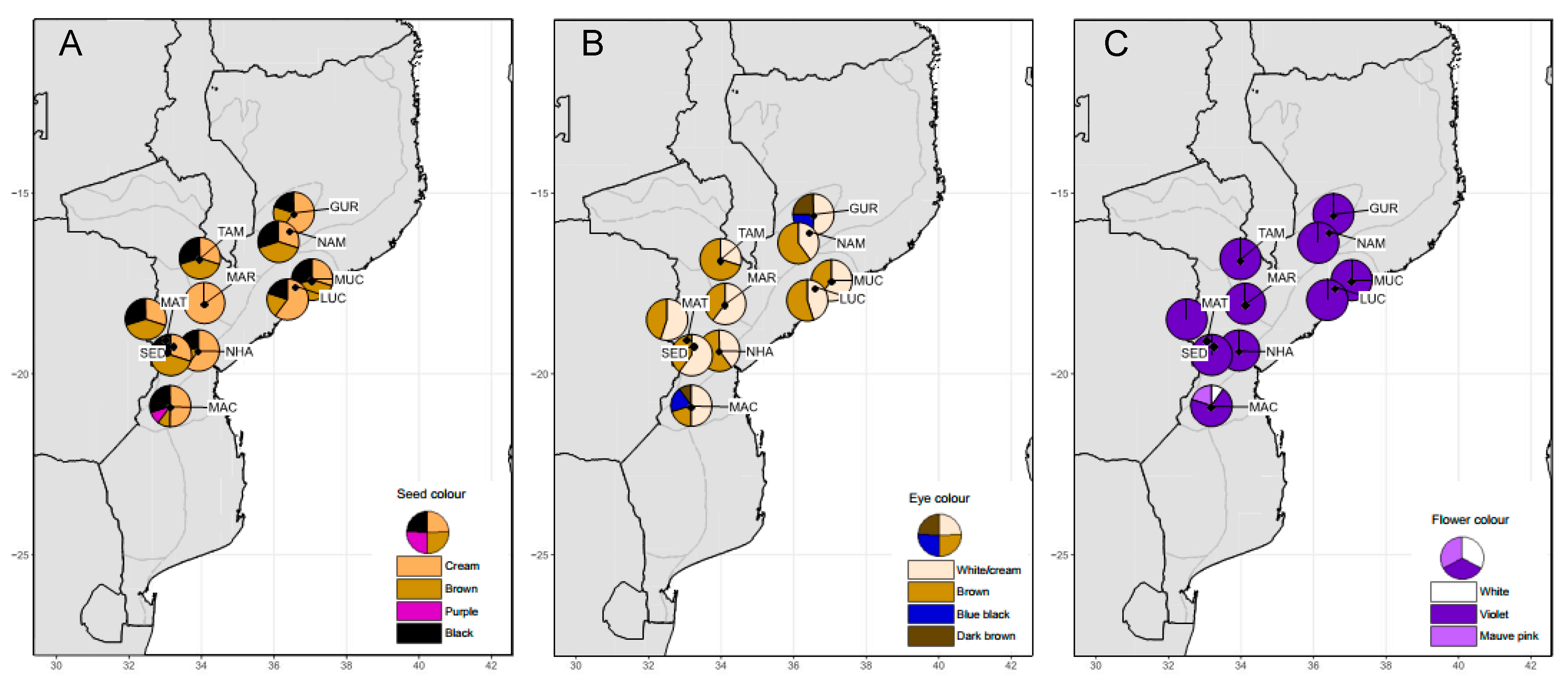
| Origin | Abbreviation | Province | AEZ | N | Latitude | Longitude |
|---|---|---|---|---|---|---|
| Gurue | GUR | North Zambezia | R10 | 6 | −15.4714 | 36.9809 |
| Namarroi | NAM | North Zambezia | R7 | 4 | −15.9539 | 36.8658 |
| Muchela | MUC | Central Zambezia | R7 | 4 | −17.3111 | 37.5147 |
| Lucas Branco | LUC | South Zambezia | R7 | 4 | −17.4919 | 37.0289 |
| Nhamatanda | NHA | Central Sofala | R4 | 4 | −19.2692 | 34.2128 |
| Maringue | MAR | Central Sofala | R5 | 3 | −17.9644 | 34.3906 |
| Tambara | TAM | North Manica | R6 | 23 | −15.1258 | 32.0558 |
| Sede Nova | SED | North Manica | R6 | 3 | −19.1164 | 33.4833 |
| Matsinho | MAT | Central Manica | R4 | 3 | −18.9511 | 33.2686 |
| Machaze | MAC | South Manica | R3 | 5 | −20.2456 | 34.1697 |
| IT16 | Commercial cultivar | - | 1 | - | - | |
| IT18 | Commercial cultivar | - | 1 | - | - |
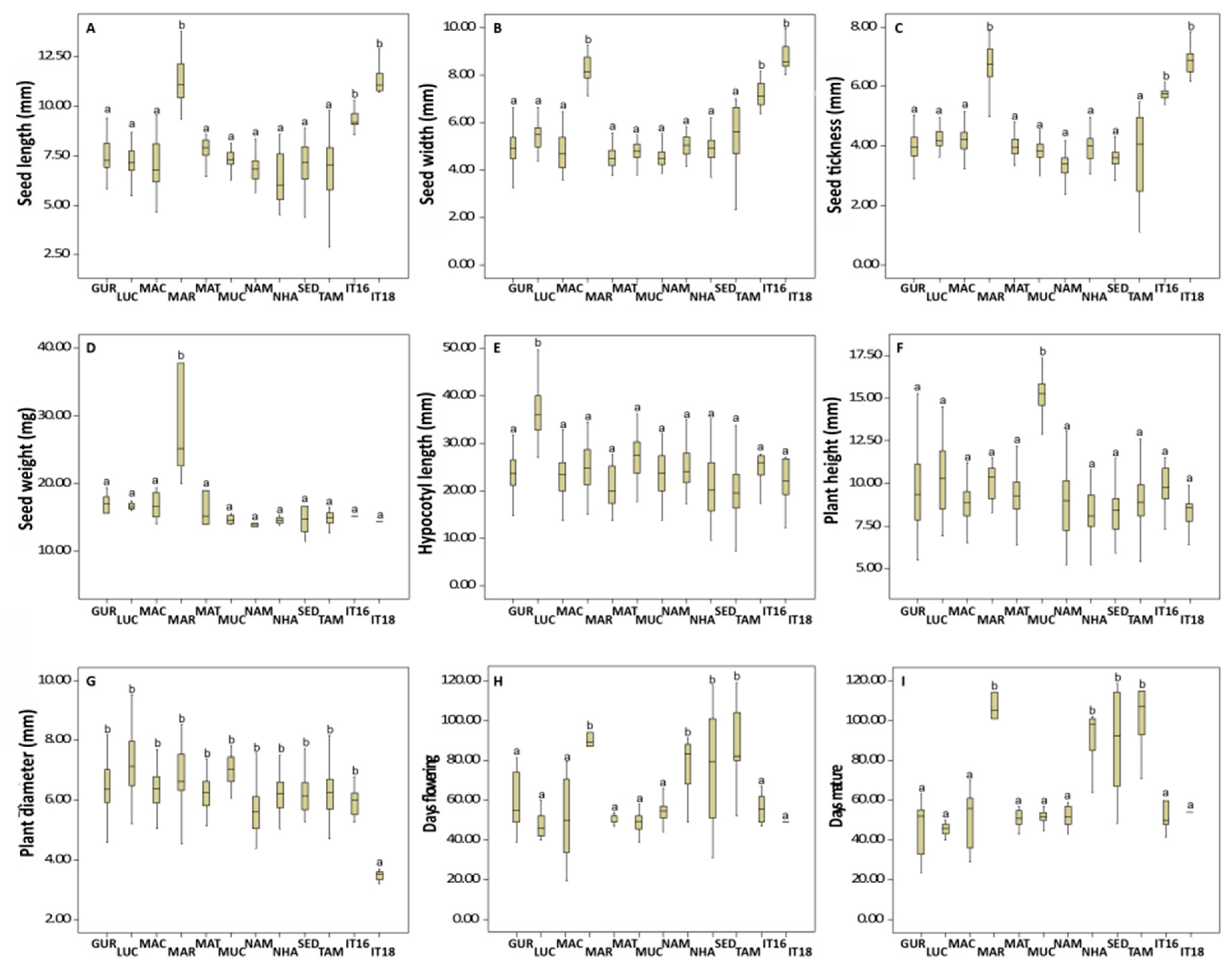
| Quantitative Traits | Mean ± SE | F Values |
|---|---|---|
| Seed length (mm) | 7.80 ± 1.74 (2.90–13.10) | 64.316 *** |
| Seed width (mm) | 6.50 ± 1.37 (1.65–9.99) | 31.960 *** |
| Seed thickness (mm) | 4.63 ± 1.69 (0.23–7.66) | 18.886 *** |
| Seed weight (mg) | 14.94 ± 4.64 (9.96–37.39) | 114.918 *** |
| Hypocotyl length (mm) | 22.57 ± 5.94 (2.01–43.07) | 13.216 *** |
| Terminal leaflet length (mm) | 112.92 ± 23.47 (9.03–201.31) | 9.273 |
| Terminal leaflet width (mm) | 72.44 ± 19.62 (4.41–145.79) | 18.075 |
| Number main branches | 15.58 ± 5.27 (4.00–28.00) | 8.049 |
| Number nodes | 11.84 ± 3.11 (4.00–24.00) | 8.428 |
| Stipule width (mm) | 5.37 ± 1.11 (2.14–8.96) | 1.521 |
| Stipule length (mm) | 14.48 ± 3.37 (5.73–24.48) | 1.288 |
| Height (mm) | 9.25 ± 1.83 (4.06–16.80) | 14.209 *** |
| Plant diameter (mm) | 4.28 ± 0.96 (2.01–9.92) | 10.756 |
| Stem diameter (mm) | 12.66 ± 3.15 (4.95–23.14) | 10.909 *** |
| Days to flowering | 71.40 ± 22.40 53.00–119.00) | 61.151 *** |
| Days to first mature pods | 86.86 ± 20.72 (62.00–115.00) | 49.827 *** |
| Qualitative Traits | Mode | Minimum and Maximum Range | χ2 |
| Seed shape | Ovoid | Kidney to rhomboid | 71.328 |
| Seed color | Cream | Cream to black | 195.902 *** |
| Texta texture | Smooth | Smooth to rough | 7.656 |
| Eye pattern | Absent | Absent to very small | 62.131 |
| Eye color | White/cream | White/cream to dark brown | 388.689 *** |
| Leaf color | Pale green | Pale green to dark green | 73.862 |
| Leaf texture | Cariaceous | Cariaceous to membranous | 96.961 |
| Leaf marking | Present | Absent to present | 98.059 |
| Terminal leaflet shape | Sub-hastate | Globose to hastate | 39.834 |
| Growth habitat | Indeterminate, spreading not climbing | Indeterminate to determinate | 62.393 |
| Twinning | None | None to pronounced | 44.426 |
| Plant pigmentation | Moderate | None to solid | 43.921 |
| Plant hairiness | Short appressed hairs | Short to pubescent hairs) | 21.715 |
| Plant vigor | Very vigorous | Non-vigorous to very vigorous | 47.598 |
| Pest incidence | Non-infested | Non-infested to low infestation | 78.901 |
| Rust incidence | Non-susceptible | Non to high susceptible | 90.931 |
| Flowering pigment | Pigmented margins | None to completed pigmented | 26.282 |
| Flower color | Violet | White to mauve-pink | 457.075 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomes, A.M.F.; Draper, D.; Nhantumbo, N.; Massinga, R.; Ramalho, J.C.; Marques, I.; Ribeiro-Barros, A.I. Diversity of Cowpea [Vigna unguiculata (L.) Walp] Landraces in Mozambique: New Opportunities for Crop Improvement and Future Breeding Programs. Agronomy 2021, 11, 991. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11050991
Gomes AMF, Draper D, Nhantumbo N, Massinga R, Ramalho JC, Marques I, Ribeiro-Barros AI. Diversity of Cowpea [Vigna unguiculata (L.) Walp] Landraces in Mozambique: New Opportunities for Crop Improvement and Future Breeding Programs. Agronomy. 2021; 11(5):991. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11050991
Chicago/Turabian StyleGomes, Ana Maria Figueira, David Draper, Nascimento Nhantumbo, Rafael Massinga, José C. Ramalho, Isabel Marques, and Ana I. Ribeiro-Barros. 2021. "Diversity of Cowpea [Vigna unguiculata (L.) Walp] Landraces in Mozambique: New Opportunities for Crop Improvement and Future Breeding Programs" Agronomy 11, no. 5: 991. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11050991