
Effect of Biofertilizers Application on Soil Biodiversity and Litter Degradation in a Commercial Apricot Orchard

 ,
, 
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Orchard Description and Treatments
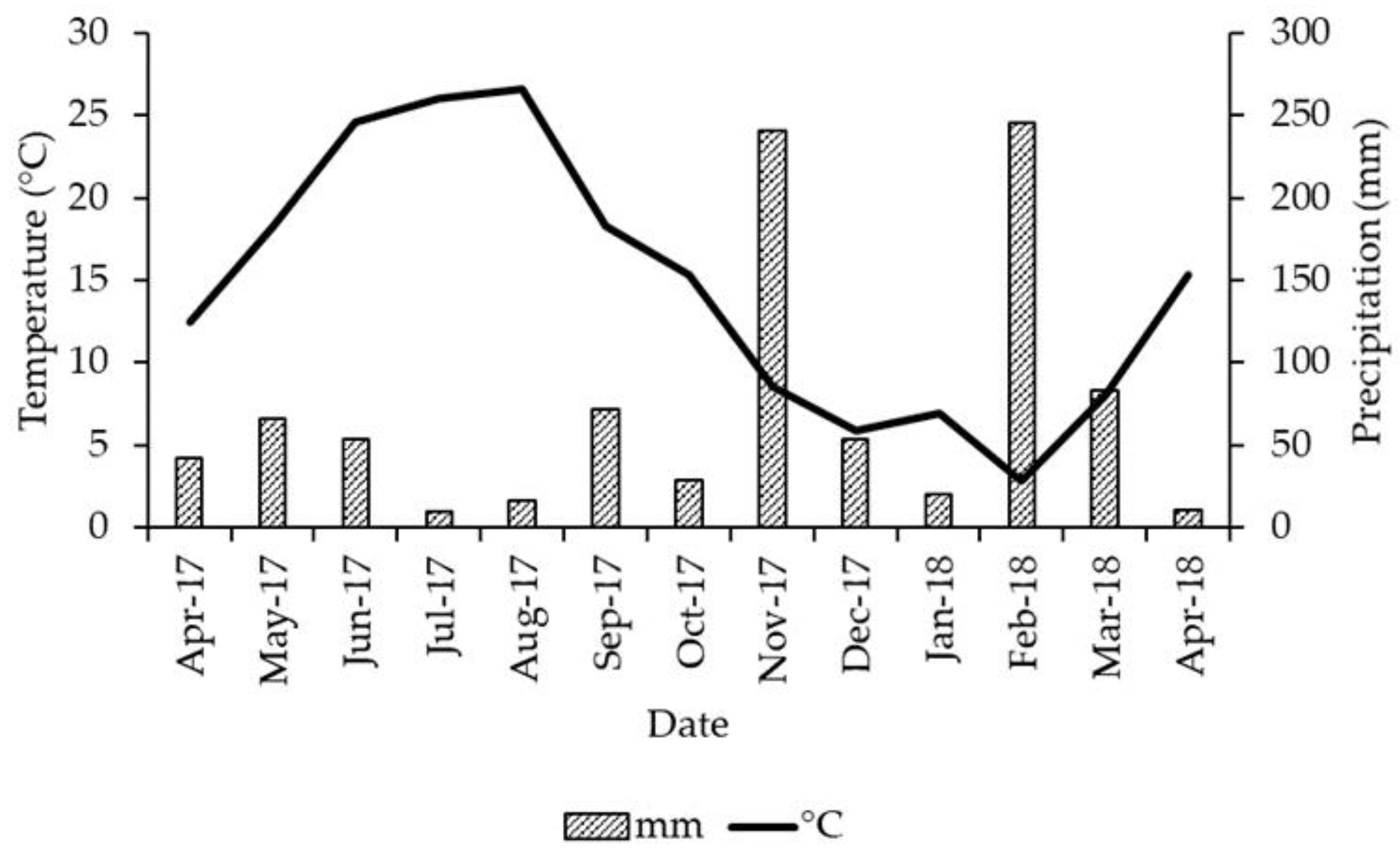
2.2. Litterbags Deposition and Sampling
2.3. Soil Microbial Analysis
2.4. Amplicon Sequencing Data Processing
2.5. Litterbags Chemical Analysis
2.6. Litterbags Thermal Analysis
2.7. Litterbag NIR Analysis
2.8. Statistical Analysis
3. Results
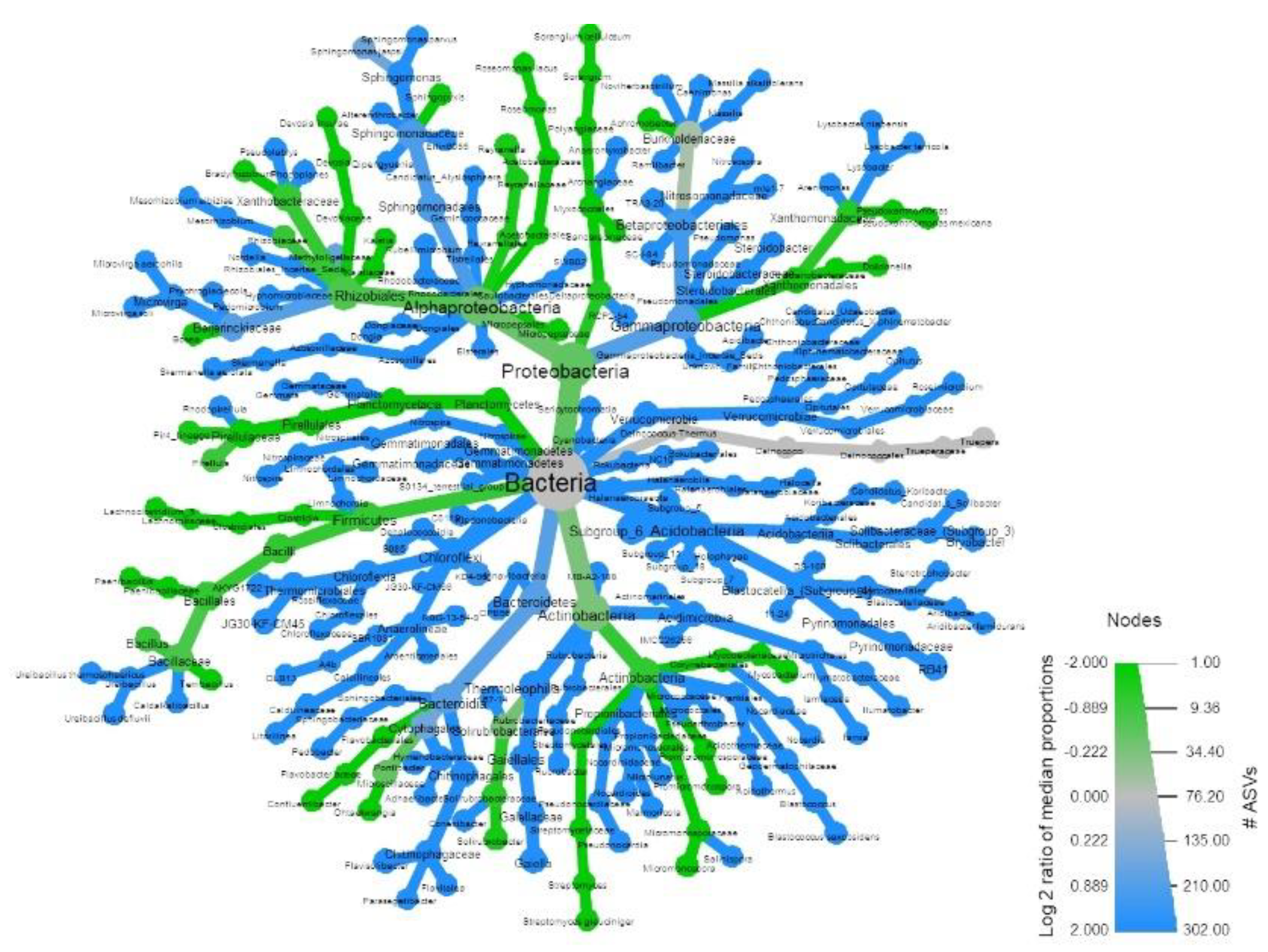
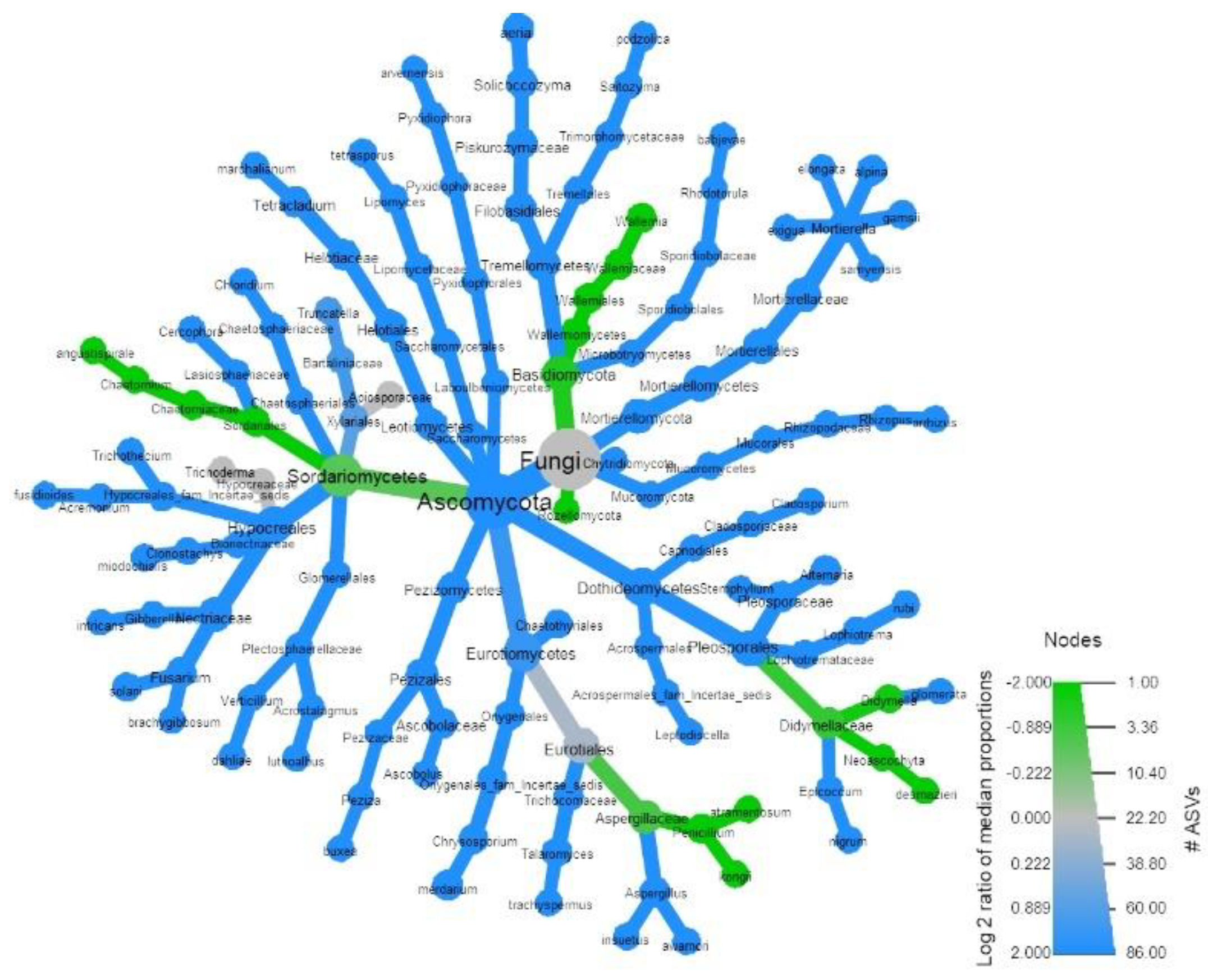
3.1. Soil Microbial Analysis
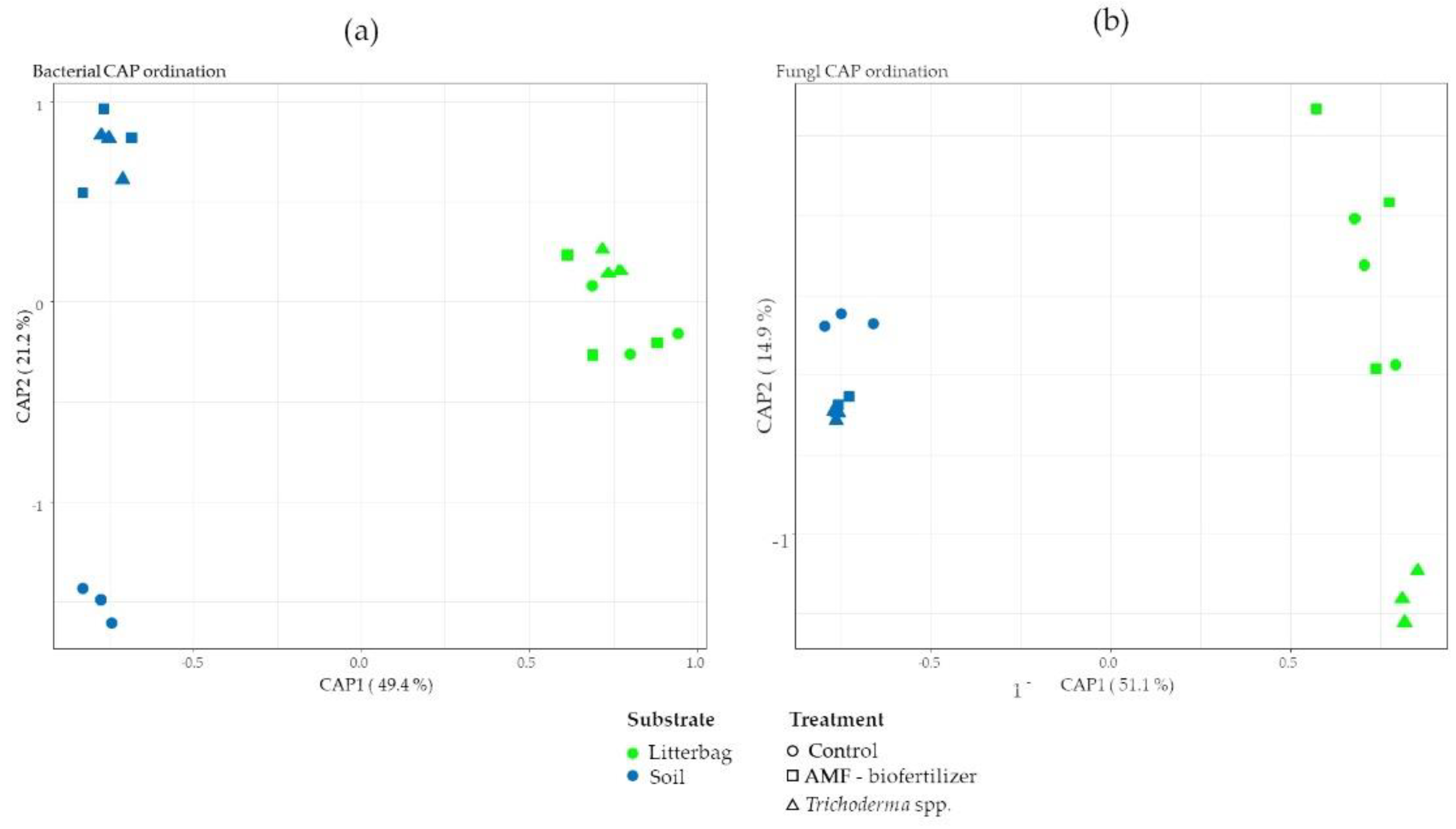
3.2. Soil Microbial Community Structure
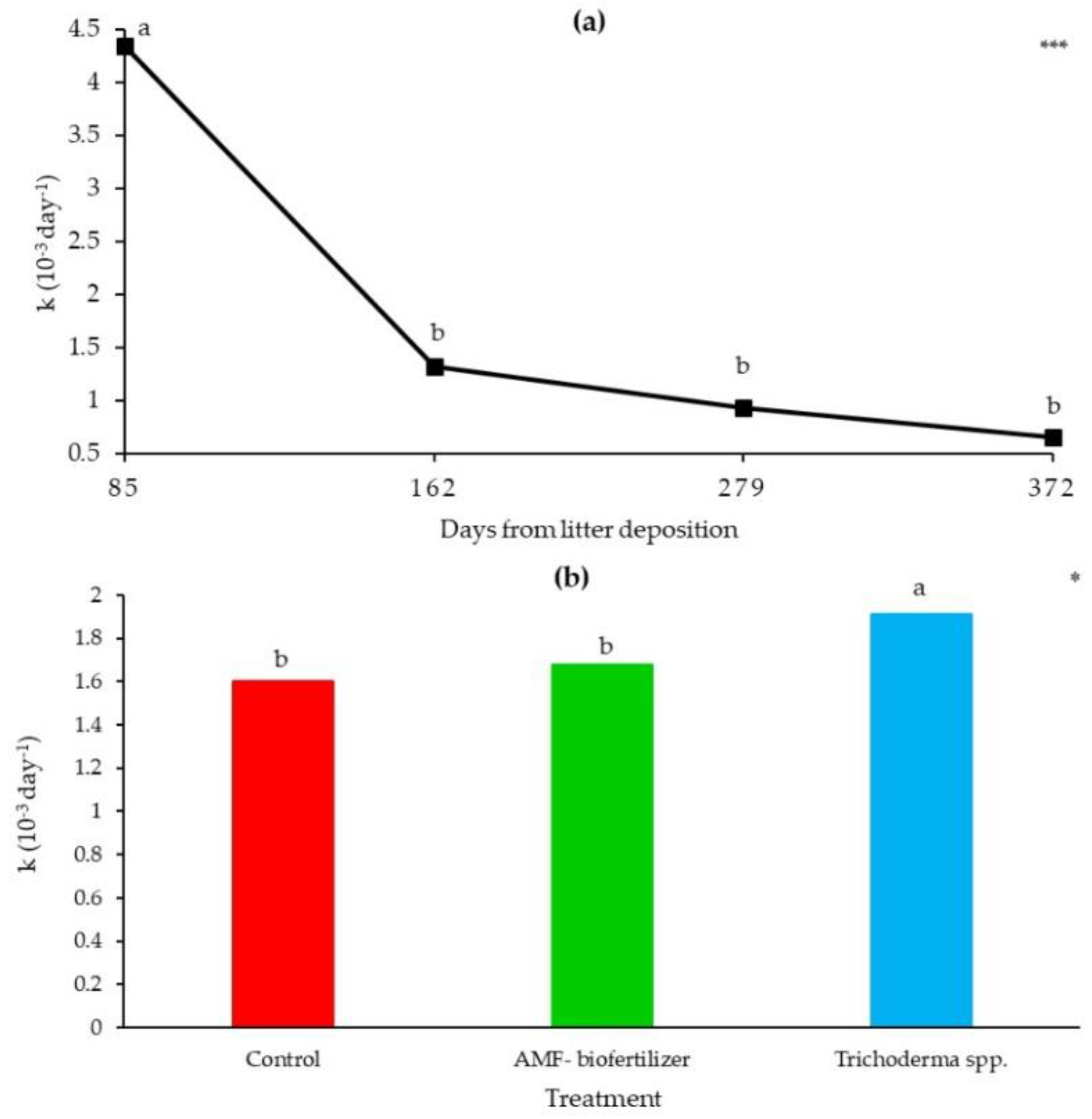
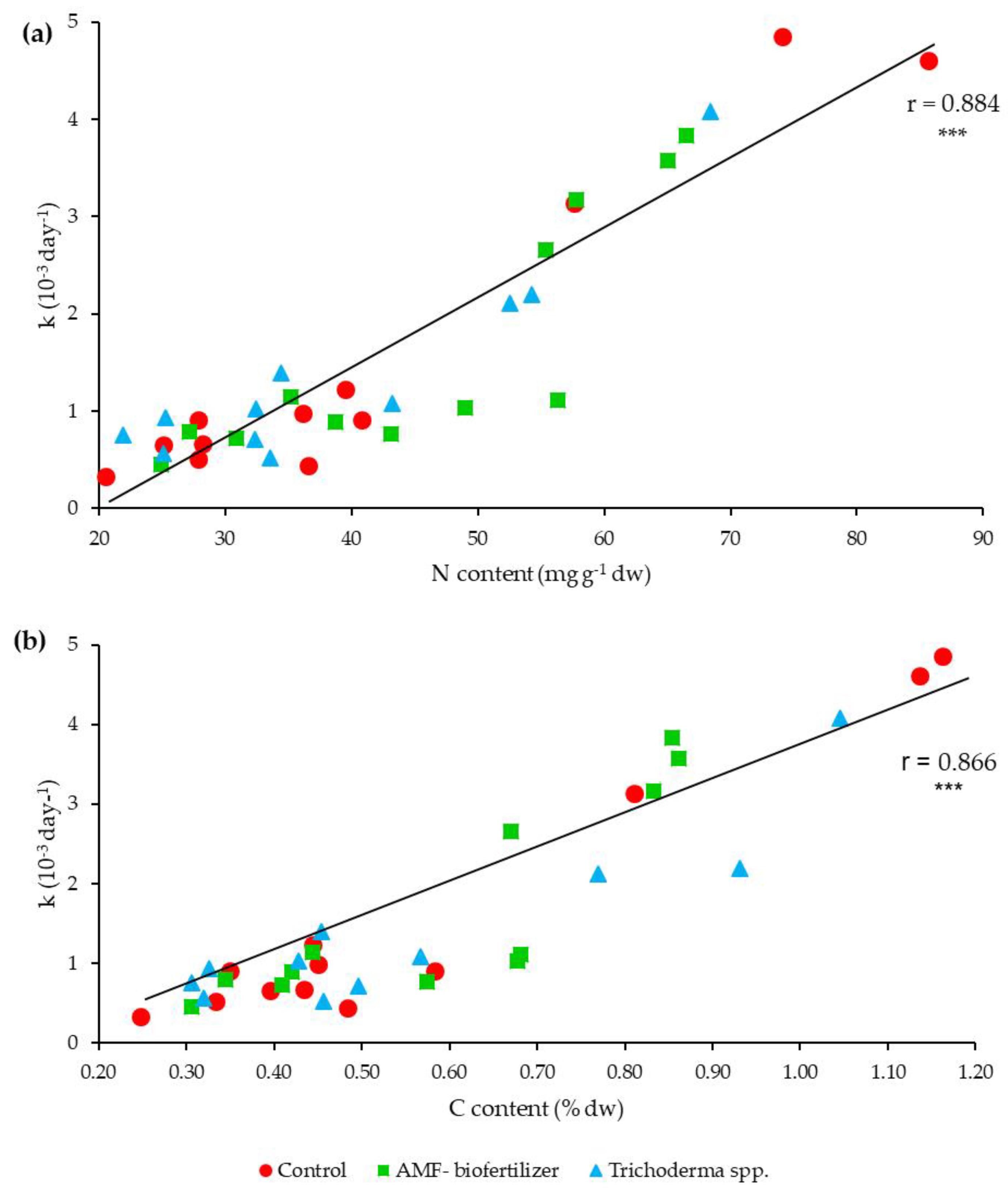
3.3. Litterbags Degradation and Thermal Analysis
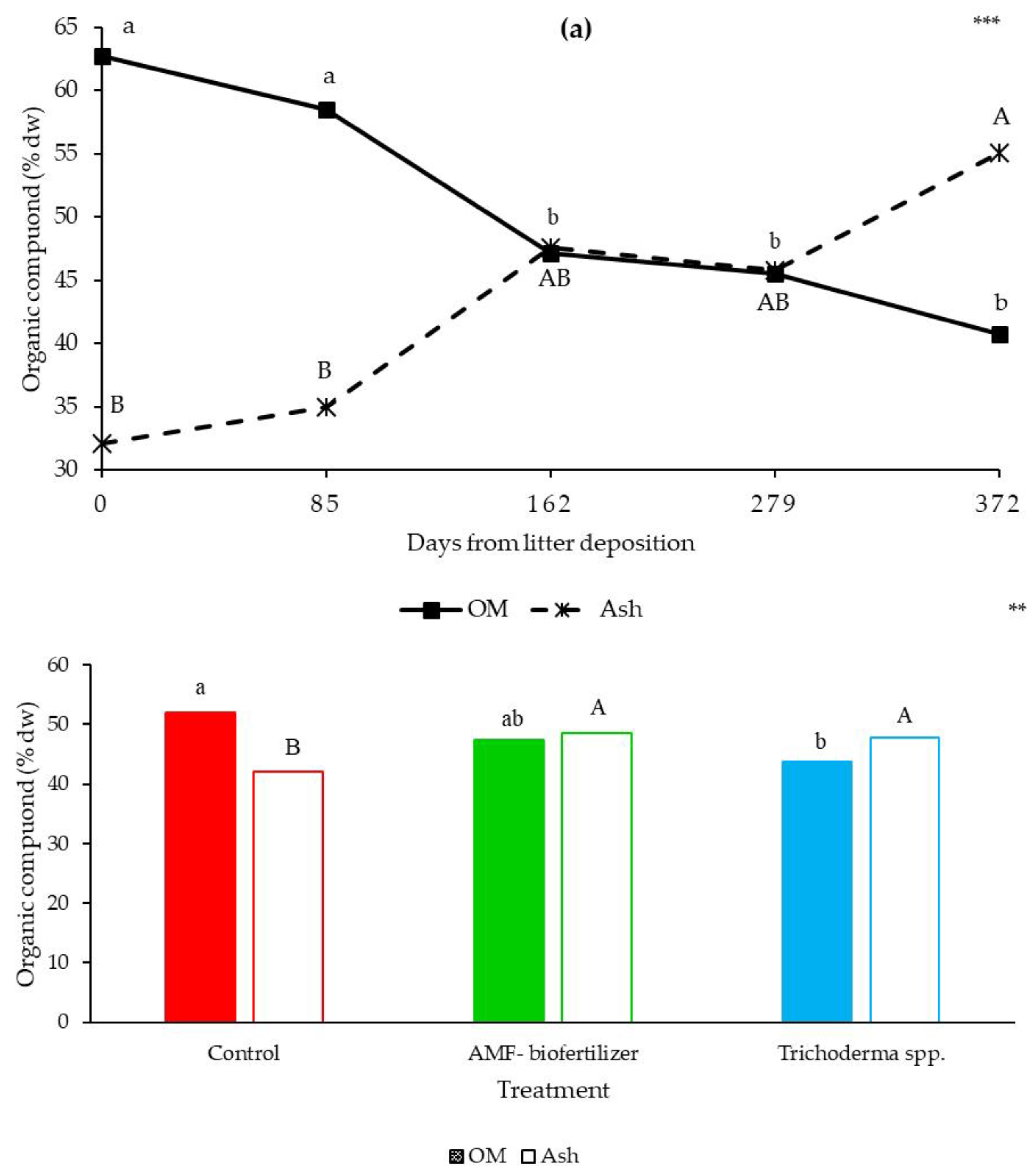
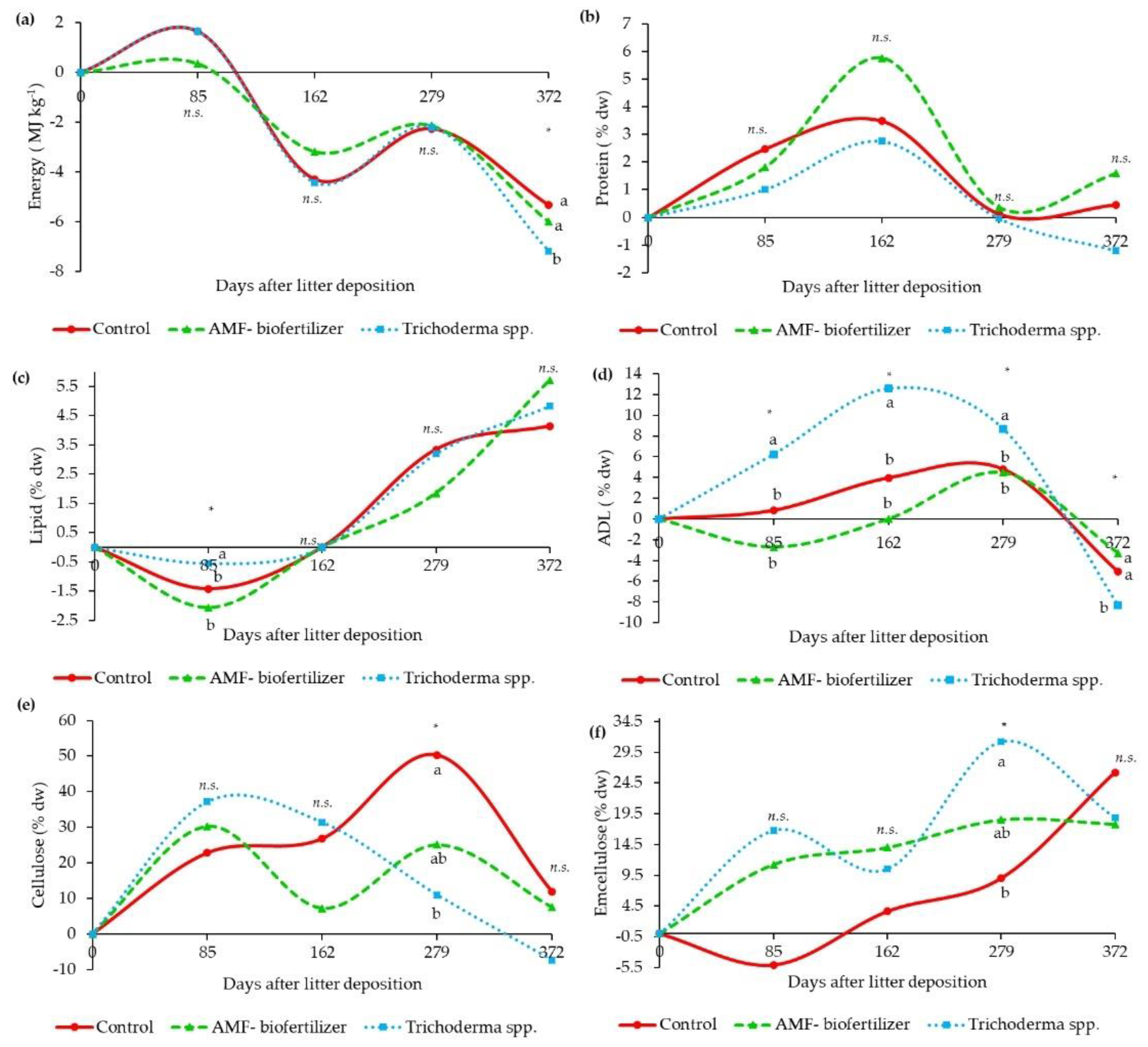
3.4. Litterbags Composition—NIR
4. Discussion
4.1. Microbial Community of Soil and Litterbags
4.2. Litterbags Decomposition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Raja, N. Biopesticides and biofertilizers: Ecofriendly sources for sustainable agriculture. J. Biofertil. Biopestici. 2013, 4, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Diacono, M.; Montemurro, F. Long-term effects of organic amendments on soil fertility. A review. Agron. Sustain. Dev. 2010, 30, 401–422. [Google Scholar] [CrossRef] [Green Version]
- Baldi, E.; Marcolini, G.; Quartieri, M.; Sorrenti, G.; Toselli, M. Effect of organic fertilization on nutrient concentration and accumulation in nectarine (Prunus persica var. nucipersica) trees: The effect of rate of application. Sci. Hort. 2014, 179, 174–179. [Google Scholar] [CrossRef]
- Megali, L.; Glauser, G.; Rasmann, S. Fertilization with beneficial microorganisms decreases tomato defenses against insect pests. Agron. Sustain. Dev. 2014, 34, 649–656. [Google Scholar] [CrossRef] [Green Version]
- Vessey, J.K. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil 2003, 255, 571–586. [Google Scholar] [CrossRef]
- Lopes, M.J.D.S.; Dias-Filho, M.B.; Gurgel, E.S.C. Successful plant growth-promoting microbes: Inoculation methods and abiotic factors. Front. Sustain. Food Syst. 2021, 5, 48–61. [Google Scholar] [CrossRef]
- Abhilash, P.C.; Dubey, R.K.; Tripathi, V.; Gupta, V.K.; Singh, H.B. Plant growth-promoting microorganisms for environmental sustainability. Trends Biotechnol. 2016, 34, 847–850. [Google Scholar] [CrossRef]
- Etesami, H. Plant–microbe interactions in plants and stress tolerance. In Plant Life under Changing Environ; Durgesh, K.T., Vijay, P.S., Devendra, K.C., Shivesh, S., Sheo, M.P., Nawal, K.D., Naleeni, R., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 355–396. [Google Scholar]
- Van Oosten, M.J.V.; Pepe, O.; Pascale, S.; Silletti, S.; Maggio, A. The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol. Technol. Agric. 2017, 4, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Gouda, S.; Kerry, R.G.; Das, G.; Paramithiotis, S.; Shin, H.; Patra, J.K. Revitalization of plant growth promoting rhizobacteria for sustainable development in agriculture. Microbiol. Res. 2018, 206, 131–140. [Google Scholar] [CrossRef]
- Malusà, E.; Pinzari, F.; Canfora, L. Efficacy of biofertilizers: Challenges to improve crop production. In Microbial Inoculants in Sustainable Agricultural Productivity; Singh, D., Singh, H., Prabha, R., Eds.; Springer: New Delhi, India, 2016; pp. 17–40. [Google Scholar]
- Mitter, B.; Brader, G.; Pfaffenbichler, N.; Sessitsch, A. Next generation microbiome applications for crop production—limitations and the need of knowledge-based solutions. Curr. Opin. Microbiol. 2019, 49, 59–65. [Google Scholar] [CrossRef]
- Bashan, Y.; Prabhu, S.R.; de-Bashan, L.E.; Kloepper, J.W. Disclosure of exact protocols of fermentation, identity of microorganisms within consortia, formation of advanced consortia with microbe-based products. Biol. Fert. Soil. 2020, 56, 443–445. [Google Scholar] [CrossRef]
- Forchetti, G.; Masciarelli, O.; Alemano, S.; Alvarez, D.; Abdala, G. Endophytic bacteria in sunflower (Helianthus annuus L.): Isolation, characterization, and production of jasmonates and abscisic acid in culture medium. Appl. Microbiol. Biotechnol. 2007, 76, 1145–1152. [Google Scholar] [CrossRef]
- Príncipe, A.; Alvarez, F.; Castro, M.G.; Zachi, L.; Fischer, S.E.; Mori, G.B.; Jofré, E. Biocontrol and PGPR features in native strains isolated from saline soils of Argentina. Curr. Microbiol. 2007, 55, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Sessitsch, A.; Pfaffenbichler, N.; Mitter, B. Microbiome applications from lab to field: Facing complexity. Trend Plant Sci. 2019, 24, 194–198. [Google Scholar] [CrossRef]
- Canfora, L.; Costa, C.; Pallottino, F.; Mocali, S. Trends in soil microbial inoculants research: A science mapping approach to unravel strengths and weaknesses of their application. Agriculture 2021, 11, 158. [Google Scholar] [CrossRef]
- Djukic, I.; Kepfer-Rojas, S.; Schmidt, I.K.; Larsen, K.S.; Beier, C.; Berg, B.; Alatalo, J. Early stage litter decomposition across biomes. Sci. Total Environ. 2018, 628, 1369–1394. [Google Scholar] [CrossRef] [Green Version]
- Gioacchini, P.; Montecchio, D.; Ferrari, E.; Ciavatta, C.; Masia, A.; George, E.; Tonon, G. Litter quality changes during decomposition investigated by thermal analysis. iForest 2015, 8, 827–837. [Google Scholar] [CrossRef] [Green Version]
- Krishna, M.P.; Mohan, M. Litter decomposition in forest ecosystems: A review. Energy Ecol. Environ. 2017, 2, 236–249. [Google Scholar] [CrossRef]
- Dalias, P.; Mprezetou, I.; Troumbis, A.Y. Use of a modified litterbag technique for the study of litter mixtures. Eur. J. Soil Biol. 2003, 39, 57–64. [Google Scholar] [CrossRef]
- Montecchio, D.; Francioso, O.; Carletti, P.; Pizzeghello, D.; Chersich, S.; Previstali, F.; Nardi, S. Thermal analysis (TG-DTA) and drift spectroscopy applied to investigate the evolution of humic acids in forest soil at different vegetation stages. J. Therm. Anal. Calor. 2006, 83, 393–399. [Google Scholar] [CrossRef]
- Beall, F.C.; Merrill, W.; Baldwin, R.C.; Wang, J.H. Thermogravimetric evaluation of fungal degradation of wood. Wood Fiber Sci. 2007, 8, 159–167. [Google Scholar]
- Masoero, G.; Delmastro, M.; Cugnetto, A.; Giovannetti, G.; Nuti, M. NIRS footprint of bio-fertilizers from hay litter-bags. J. Agron. Res. 2018, 1, 22–33. [Google Scholar] [CrossRef] [Green Version]
- Genot, V.; Colinet, G.; Bock, L.; Vanvyve, D.; Reusen, Y.; Dardenne, P. Near infrared reflectance spectroscopy for estimating soil characteristics valuable in the diagnosis of soil fertility. J. Near Infrared Spectrosc. 2011, 19, 117–138. [Google Scholar] [CrossRef]
- Andueza, D.; Picard, F.; Martin-Rosset, W.; Aufrère, J. Near-infrared spectroscopy calibrations performed on oven-dried green forages for the prediction of chemical composition and nutritive value of preserved forage for ruminants. Appl. Spectrosc. 2016, 70, 1321–1327. [Google Scholar] [CrossRef]
- Debusk, W.F.; Reddy, K.R. Litter decomposition and nutrient dynamics in a phosphorus enriched everglades marsh. Biogeochemistry 2005, 75, 217–240. [Google Scholar] [CrossRef]
- Anderson, J.; Domsch, K. A physiological method for the quantitative measurement of microbial biomass in soils. Soil Biol. Biochem. 1978, 10, 215–221. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [Green Version]
- Vainio, E.J.; Hantula, J. Direct analysis of wood-inhabiting fungi using denaturing gradient gel electrophoresis of amplified ribosomal DNA. Mycol. Res. 2000, 104, 927–936. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, S.; FASTQC. A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 9 September 2019).
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Bengtsson-Palme, J.; Thorell, K.; Wurzbacher, C.; Sjöling, Å.; Nilsson, R.H. Metaxa2 Diversity Tools: Easing microbial community analysis with Metaxa2. Ecol. Inform. 2016, 33, 45–50. [Google Scholar] [CrossRef]
- Bengtsson-Palme, J.; Ryberg, M.; Hartmann, M.; Branco, S.; Wang, Z.; Godhe, A.; De Wit, P.; Sánchez-García, M.; Ebersberger, I.; de Sousa, F.; et al. Improved software detection and extraction of ITS1 and ITS2 from ribosomal ITS sequences of fungi and other eukaryotes for analysis of environmental sequencing data. Methods Ecol. Evol. 2013, 4, 914–919. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Larsson, K.H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glöckner, O.F.; Tedersoo, L.; et al. The UNITE database for molecular identification of fungi: Handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 2019, 47, D259–D264. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Tassone, S.; Masoero, G.; Peiretti, P.G. Vibrational spectroscopy to predict in vitro digestibility and the maturity index of different forage crops during the growing cycle and after freeze- or oven-drying treatment. Anim. Feed Sci. Technol. 2014, 194, 12–25. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Simpson, J.; Levesque, C.A. RAM: R for Amplicon-Sequencing-Based Microbial-Ecology. 2018. Available online: CRAN.R-project.org/package=RAM (accessed on 12 March 2021).
- Love, M.; Ahlmann-Eltze, C.; Forbes, K.; Anders, S.; Huber, W. DESeq2: Differential Gene Expression Analysis Based on the Negative Binomial Distribution. Bioconductor Version: Release (3.12). 2021. Available online: https://rdrr.io/bioc/DESeq2/ (accessed on 12 March 2021).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. 2019. Available online: CRAN.R-project.org/package=vegan (accessed on 28 August 2020).
- Leff, J.W. Leffj/Mctoolsr. 2020. Available online: https://github.com/leffj/mctoolsr (accessed on 12 March 2021).
- Simpson, G.L. Ggvegan: Ggplot2-Based Plots for Vegan. 2021. Available online: https://gavinsimpson.github.io/ggvegan (accessed on 12 March 2021).
- Hervé, M. RVAideMemoire: Testing and Plotting Procedures for Biostatistics. 2020. Available online: https://CRAN.R-project.org/package=RVAideMemoire (accessed on 28 August 2020).
- Foster, Z.; Grunwald, N.; Gilmore, R. Metacoder: Tools for Parsing, Manipulating, and Graphing Taxonomic Abundance Data. 2020. Available online: https://CRAN.R-project.org/package=metacoder (accessed on 12 March 2021).
- Coulis, M.; Hättenschwiler, S.; Fromin, N.; David, J.F. Macroarthropod-microorganism interactions during the decomposition of Mediterranean shrub litter at different moisture levels. Soil Biol. Biochem. 2013, 64, 114–121. [Google Scholar] [CrossRef]
- Austin, A.T.; Vivanco, L.; González-Arzac, A.; Pérez, L.I. There’s no place like home? An exploration of the mechanisms behind plant litter–decomposer affinity in terrestrial ecosystems. New Phytol. 2014, 204, 307–314. [Google Scholar] [CrossRef]
- Baldi, E.; Toselli, M.; Masoero, G.; Nuti, M. Organic and symbiotic fertilization of tomato plants monitored by Litterbag-NIRS and Foliar-NIRS rapid spectroscopic methods. J. Agron. Res. 2020, 3, 9–26. [Google Scholar] [CrossRef]
- Dehsheikh, A.B.; Sourestani, M.M.; Zolfaghari, M.; Enayatizamir, N. Changes in soil microbial activity, essential oil quantity, and quality of Thai basil as response to biofertilizers and humic acid. J. Clean. Prod. 2020, 256, 120439. [Google Scholar] [CrossRef]
- Wertheim, S.J.; Webster, A.D. Manipulation of growth and development by plant bioregulators. In Fundamentals of Temperate Zone Tree Fruit Production; Tromp, J., Webster, A.D., Wertheim, S.J., Eds.; Backhuys Publishers: Leiden, The Netherlands, 2005; pp. 267–294. [Google Scholar]
- Pirlak, L.; Turan, M.; Sahin, F.; Esitken, A. Floral and foliar application of plant growth promoting rhizobacteria (PGPR) to apples increases yield, growth, and nutrient element contents of leaves. J. Sustain. Agric. 2007, 30, 145–155. [Google Scholar] [CrossRef]
- Ortas, I.; Demirbas, A.; Akpinar, C. Time period and nutrient contents alter the mycorrhizal responsiveness of citrus seedlings Eur. J. Hortic. Sci. 2018, 83, 72–80. [Google Scholar]
- Ansari, M.H.; Hashemabadi, D.; Mahdavi, M.; Kaviani, B. The role of Pseudomonas strains and arbuscular mycorrhiza fungi as organic phosphate–solubilizing in the yield and quality improvement of strawberry (Fragaria × ananassa Duch cv. Selva) fruit. Acta Sci. Pol. Hortoru. 2018, 17, 93–107. [Google Scholar] [CrossRef]
- Todeschini, V.; AitLahmidi, N.; Mazzucco, E.; Marsano, F.; Gosetti, F.; Robotti, E.; Bona, E.; Massa, N.; Bonneau, L.; Marengo, E.; et al. Impact of beneficial microorganisms on strawberry growth, fruit production, nutritional quality, and volatilome. Front. Plant Sci. 2018, 9, 1611. [Google Scholar] [CrossRef]
- Basile, B.; Rouphael, Y.; Colla, G.; Soppelsa, S.; Andreotti, C. Appraisal of emerging crop management opportunities in fruit trees, grapevines and berry crops facilitated by the application of biostimulants. Sci. Hort. 2020, 267, 109330. [Google Scholar] [CrossRef]
- Bergmann, G.T.; Bates, S.T.; Eilers, K.G.; Lauber, C.L.; Caporaso, J.G.; Walters, W.A.; Knight, R.; Fierer, N. The under-recognized dominance of Verrucomicrobia in soil bacterial communities. Soil Biol. Biochem. 2011, 43, 1450–1455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldi, E.; Amadei, P.; Pelliconi, F.; Toselli, M. Use of Trichoderma spp. and arbuscular mycorrhizal fungi to increase soil beneficial population of bacteria in a nectarine commercial orchard: Effect on root growth, nutrient acquisition and replanting disease. J. Plant Nutr. 2016, 39, 1147–1155. [Google Scholar] [CrossRef]
- Kausar, H.; Sariah, M.; Saud, H.M.; Alam, M.Z.; Ismail, M.R. Isolation and screening of potential actinobacteria for rapid composting of rice straw. Biodegradation 2011, 22, 367–375. [Google Scholar] [CrossRef]
- Mei, J.; Shen, X.; Gang, L.; Xu, H.; Wu, F.; Sheng, L. A novel lignin degradation bacteria-Bacillus amyloliquefaciens SL-7 used to degrade straw lignin efficiently. Bioresour. Technol. 2020, 310, 123445. [Google Scholar] [CrossRef]
- Janusz, G.; Pawlik, A.; Sulej, J.; Świderska-Burek, U.; Jarosz-Wilkołazka, A.; Paszczyński, A. (Lignin degradation: Microorganisms, enzymes involved, genomes analysis and evolution. FEMS Microbiol. Rev. 2017, 41, 941–962. [Google Scholar] [CrossRef] [Green Version]
- Sigoillot, J.C.; Berrin, J.G.; Bey, M.; Lesage-Meessen, L.; Levasseur, A.; Lomascolo, A.; Record, E.; Uzan-Boukhris, E. Fungal strategies for lignin degradation. In Lignins: Biosynthesis, Biodegradation and Bioengineering; Jouanin, L., Lapierre, C., Eds.; Academic Press: London, UK; Elsevier: London, UK, 2012; pp. 263–308. [Google Scholar]
- Holker, U.; Dohse, J.; Hofer, M. Extracellular laccases in ascomycetes Trichoderma atroviride and Trichoderma harzianum. Folia Microbiol. 2002, 47, 423–437. [Google Scholar] [CrossRef] [PubMed]
- Velazquez-Cedeiio, M.A.; Farnet, A.M.; Ferre, E. Variations of lignocellulosic activities in dual cultures of >Pleurotus ostreatus and Trichoderma longibrachiatum on unsterilized wheat straw. Mycologia 2004, 96, 712–719. [Google Scholar] [CrossRef] [PubMed]
- Gochev, V.K.; Krastanov, A.I. Isolation of laccase producing Trichoderma spp. Bulg. J. Agric. Sci. 2007, 13, 171–176. [Google Scholar]
- Sharma, B.L.; Singh, S.P.; Sharma, M.L. Bio-degradation of crop residues by Trichoderma species vis-a vis nutrient quality of the prepared compost. Sugar Tech. 2012, 14, 174–180. [Google Scholar] [CrossRef]
- Ahmed, A.S.; Ezziyyani, M.; Sánchez, C.P.; Candela, M.E. Effect of chitin on biological control activity of Bacillus spp. and Trichoderma harzianum against root rot disease in pepper (Capsicum annuum) plants. Eur. J. Plant Pathol. 2009, 109, 633–637. [Google Scholar] [CrossRef]
- Zafra, G.; Moreno-Montaño, A.; Absalón, Á.E.; Cortés-Espinosa, D.V. Degradation of polycyclic aromatic hydrocarbons in soil by a tolerant strain of Trichoderma asperellum. Environ. Sci. Pollut. R. 2015, 22, 1034–1042. [Google Scholar] [CrossRef]
- Zhang, W.; Yuan, S.; Hu, N.; Lou, Y.; Wang, S. Predicting soil fauna effect on plant litter decomposition by using boosted regression trees. Soil Biol. Biochem. 2015, 82, 81–86. [Google Scholar] [CrossRef]
- Wang, Y.; Li, F.Y.; Song, X.; Wang, X.; Suri, G.; Baoyin, T. Changes in litter decomposition rate of dominant plants in a semi-arid steppe across different land-use types: Soil moisture, not home-field advantage, plays a dominant role. Agric. Ecosyst. Environ. 2020, 303, 107119. [Google Scholar] [CrossRef]
- Toselli, M.; Baldi, E.; Cavani, L.; Sorrenti, G. Nutrient management in fruit crops: An organic way. In Fruit Crops: Diagnosis and Management of Nutrients Constraints; Elsevier: Amsterdam, The Netherlands, 2019; pp. 379–390. [Google Scholar]
- Rovira, P.; Vallejo, V.R. Examination of thermal and acid hydrolysis procedures in characterization of soil organic matter. Comm. Soil Sci. Plant Anal. 2000, 31, 81–100. [Google Scholar] [CrossRef]
- Kosheleva, Y.P.; Trofimov, S.Y. Characteristics of the biochemical composition of plant litter at different stages of decomposition (according to thermal analysis data). Biol. Bull. 2008, 35, 64–69. [Google Scholar] [CrossRef]
- Hamadi, Z.; Steinberger, Y.; Kutiel, P.; Lavee, H.; Barness, G. Decomposition of Avena sterilis litter under arid conditions. J. Arid Environ. 2000, 46, 281–293. [Google Scholar] [CrossRef]
- McKee, G.A.; Soong, J.L.; Caldéron, F.; Borch, T.; Cotrufo, M.F. An integrated spectroscopic and wet chemical approach to investigate grass litter decomposition chemistry. Biogeochemistry 2016, 128, 107–123. [Google Scholar] [CrossRef] [Green Version]
- Cortez, J.; Demard, J.M.; Bottner, P.; Monrozier, L.J. Decomposition of Mediterranean leaf litters: A microcosm experiment investigating relationships between decomposition rates and litter quality. Soil Biol. Biochem. 1996, 28, 443–452. [Google Scholar] [CrossRef]
- Klotzbücher, T.; Kaiser, K.; Guggenberger, G.; Gatzek, C.; Kalbitz, K. A new conceptual model for the fate of lignin in decomposing plant litter. Ecology 2011, 92, 1052–1062. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Unit | Value |
|---|---|---|
| C | % DW | 32.4 ± 0.091 1 |
| N | % DW | 0.823 ± 0.006 |
| Energy | MJ kg−1 | 18.8 ± 0.323 |
| Protein | % DW | 2.87 ± 0.425 |
| Lipid | % DW | 3.69 ± 0.182 |
| ADL 2 | % DW | 12.2 ± 0.992 |
| Cellulose | % DW | 35.5 ± 3.14 |
| Hemicellulose | % DW | 5.65 ± 0.220 |
| Treatment | 85 | 162 | 279 | 372 |
|---|---|---|---|---|
| Control | 201 ± 44.1 1 | 401 ± 31.4 b | 276 ± 32.4 b | 641 ± 109 b |
| AMF-biofertilizer | 164 ± 35.8 | 617 ± 41.7 a | 361 ± 11.3 b | 854 ± 104 a |
| Trichoderma spp. | 249 ± 94.7 | 611 ± 62.7 a | 533 ± 43.2 a | 807 ± 124 a |
| Significance | n.s.2 | * | ** | * |
| Treatment | Mass Loss (%) | Temperature (°C) | ||||
|---|---|---|---|---|---|---|
| EXO1 1 | EXO2 | EXO3 | EXO1 | EXO2 | EXO3 | |
| Control | 31.3 ± 2.33 2 a 3 | 17.4 ± 0.980 | 3.56 ± 0.181 | 346 ± 1.02 | 442 ± 4.38 ab | 521 ± 1.87 |
| AMF- biofertilizer | 27.9 ± 1.53 ab | 16.7 ± 1.63 | 3.10 ± 0.424 | 345 ± 1.15 | 450 ± 4.55 a | 519 ± 0.810 |
| Trichoderma spp. | 25.8 ± 1.76 b | 15.5 ± 1.33 | 2.87 ± 0.169 | 345 ± 0.591 | 433 ± 4.99 b | 519 ± 0.169 |
| Significance | *3 | n.s. | n.s. | n.s. | ** | n.s. |
| Days | ||||||
| 0 | 41.6 ± 0.440 4 a | 11.5 ± 0.476 c | 4.30 ± 0.109 a | 327 ± 0.567 c | 410 ± 1.40 c | 471 ± 0.157 b |
| 85 | 36.3 ± 2.37 b | 21.3 ± 1.24 a | 2.40 ± 0.245 b | 344 ± 0.500 b | 460 ± 2.44 a | 518 ± 0.720 a |
| 162 | 27.3 ± 1.60 c | 15.8 ± 0.863 b | 3.91 ± 0.422 a | 345 ± 0.597 b | 444 ± 3.24 b | 522 ± 0.994 a |
| 279 | 26.4 ± 1.20 c | 16.0 ± 1.54 b | 2.58 ± 0.259 b | 348 ± 1.56 a | 433 ± 6.16 b | 519 ± 2.30 a |
| 372 | 24.0 ± 1.76 c | 13.5 ± 1.51 bc | 3.21 ± 0.215 ab | 343 ± 1.14 b | 429 ± 5.11 b | 517 ± 1.59 a |
| Significance | *** | *** | ** | *** | *** | * |
| Treatment*days | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. |
| Treatment | 0 | 85 | 162 | 279 | 372 |
|---|---|---|---|---|---|
| Control | 0.381 ± 0.020 1 | 0.446 ± 0.02 | 0.684 ± 0.05 | 0.767 ± 0.09 | 0.699 ± 0.04 |
| AMF- biofertilizer | 0.381 ± 0.020 | 0.750 ± 0.03 | 0.675 ± 0.02 | 0.674 ± 0.05 | 0.633 ±0.03 |
| Trichoderma spp. | 0.381 ± 0.020 | 0.617 ± 0.10 | 0.838 ± 0.04 | 0.532± 0.01 | 0.752 ±0.08 |
| Significance | 2SEM2= 0.030 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baldi, E.; Gioacchini, P.; Montecchio, D.; Mocali, S.; Antonielli, L.; Masoero, G.; Toselli, M. Effect of Biofertilizers Application on Soil Biodiversity and Litter Degradation in a Commercial Apricot Orchard. Agronomy 2021, 11, 1116. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11061116
Baldi E, Gioacchini P, Montecchio D, Mocali S, Antonielli L, Masoero G, Toselli M. Effect of Biofertilizers Application on Soil Biodiversity and Litter Degradation in a Commercial Apricot Orchard. Agronomy. 2021; 11(6):1116. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11061116
Chicago/Turabian StyleBaldi, Elena, Paola Gioacchini, Daniela Montecchio, Stefano Mocali, Livio Antonielli, Giorgio Masoero, and Moreno Toselli. 2021. "Effect of Biofertilizers Application on Soil Biodiversity and Litter Degradation in a Commercial Apricot Orchard" Agronomy 11, no. 6: 1116. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11061116