
Strip Tillage and Crop Residue Retention Decrease the Size but Increase the Diversity of the Weed Seed Bank under Intensive Rice-Based Crop Rotations in Bangladesh

 ,
, 
 , ,
, ,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
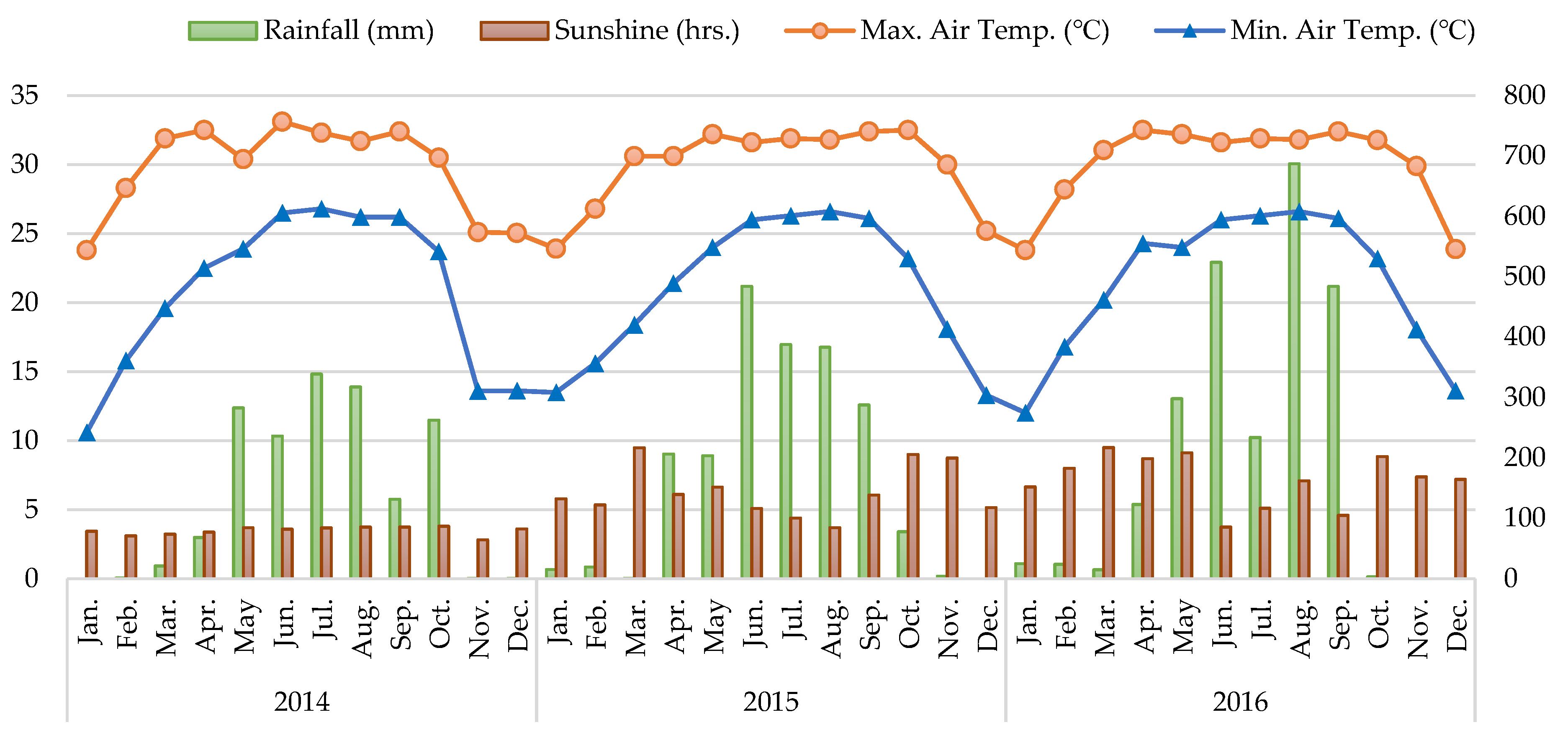
2.1. The Glimpse of the Field, Crops, and Climate of on-Farm CA Experiments
2.2. Methods of Land Preparation
2.3. Mulching of Crop Residues
2.4. Weeding Methods
2.5. Analysis of Soil Weed Seed Bank
2.6. Indicators of Diversity, Dominance, and Similarity
2.7. Data Analysis
3. Results
3.1. Analysis of Weed Species Composition in the Seed Bank
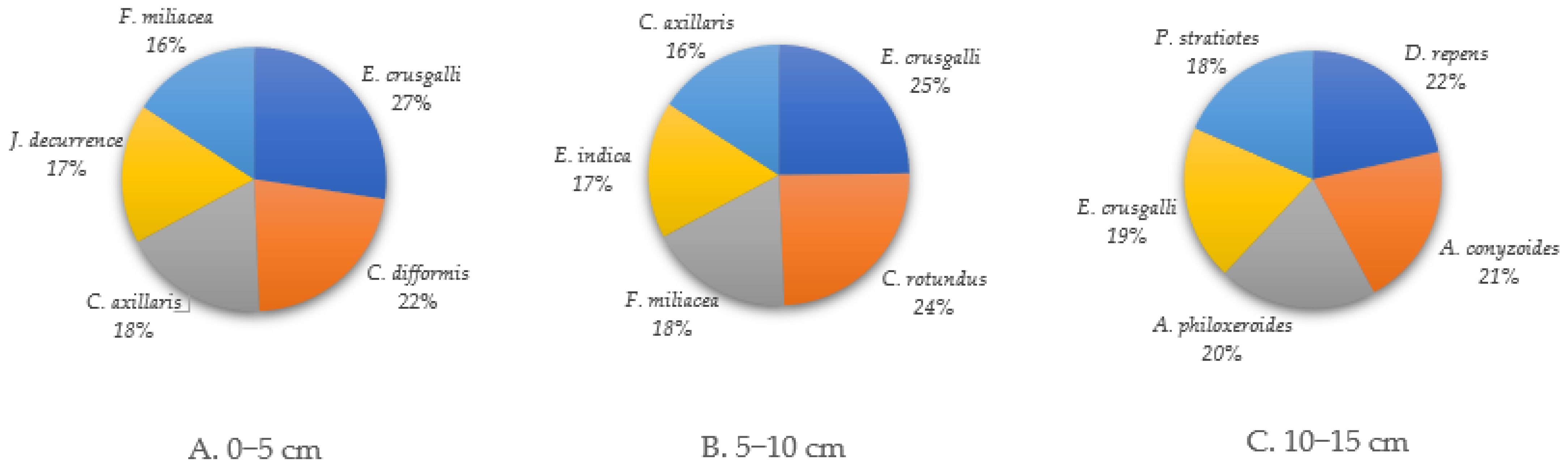
3.1.1. Weed Species Composition at 0–5 cm Soil Depth
3.1.2. Weed Species Composition at 5–10 cm Soil Depth
3.1.3. Weed Species Composition at 10–15 cm Soil Depth
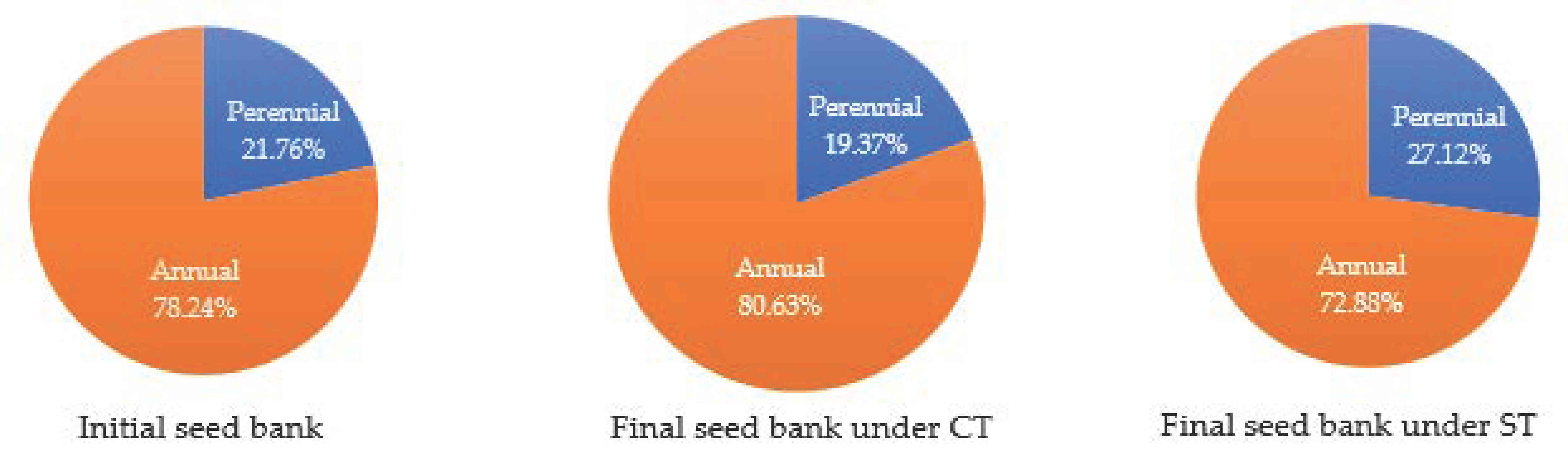
3.2. Effect of Tillage Practices and Residue Levels on Shannon’s Diversity Index (H’), Simpson’s Dominance Index (SI), and SØrensen’s Similarity Index of the Seed Bank
3.3. Effect of Tillage and Residue Levels on the Weed Density (Plant m−2) and Biomass (g m−2)
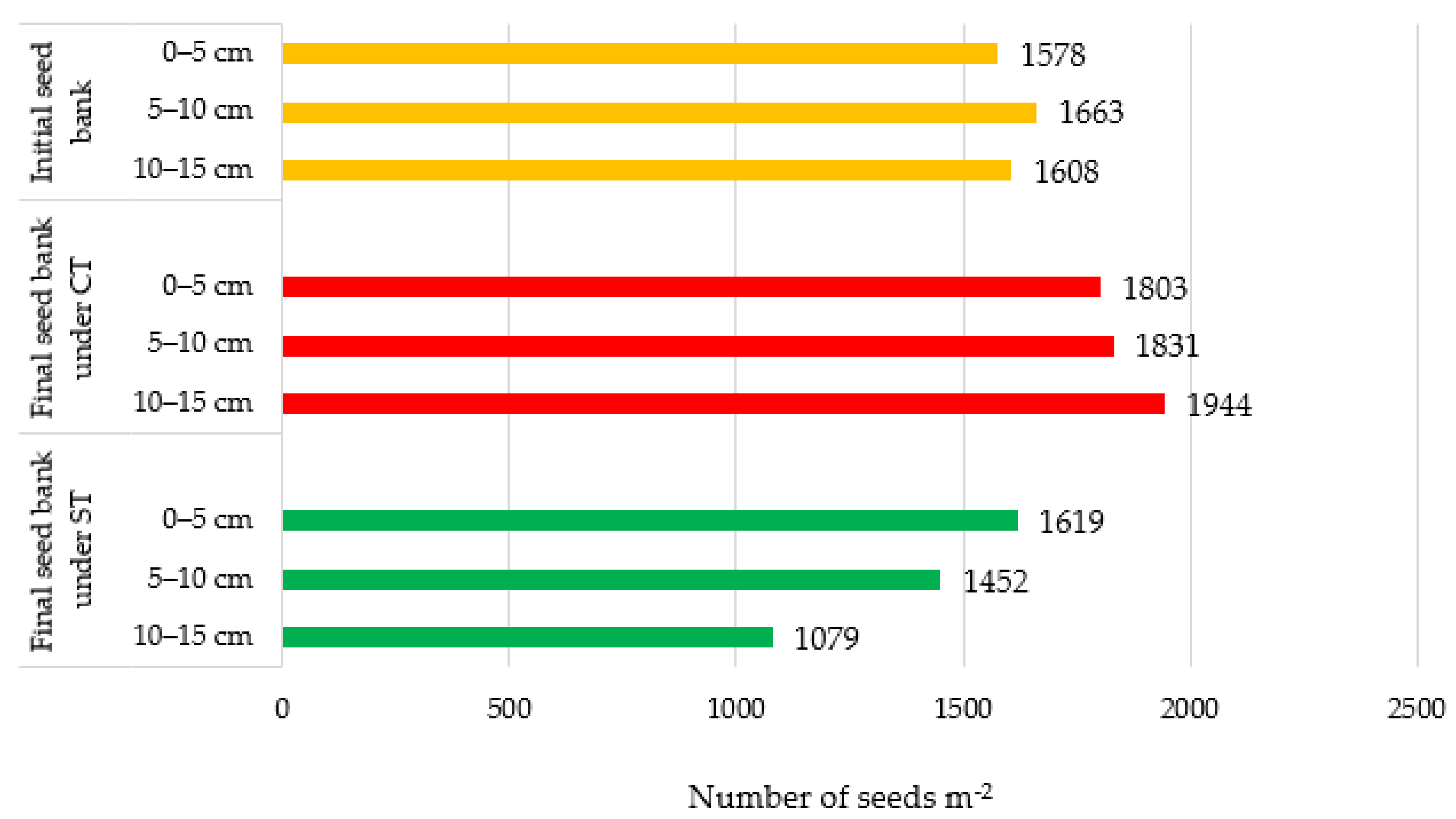
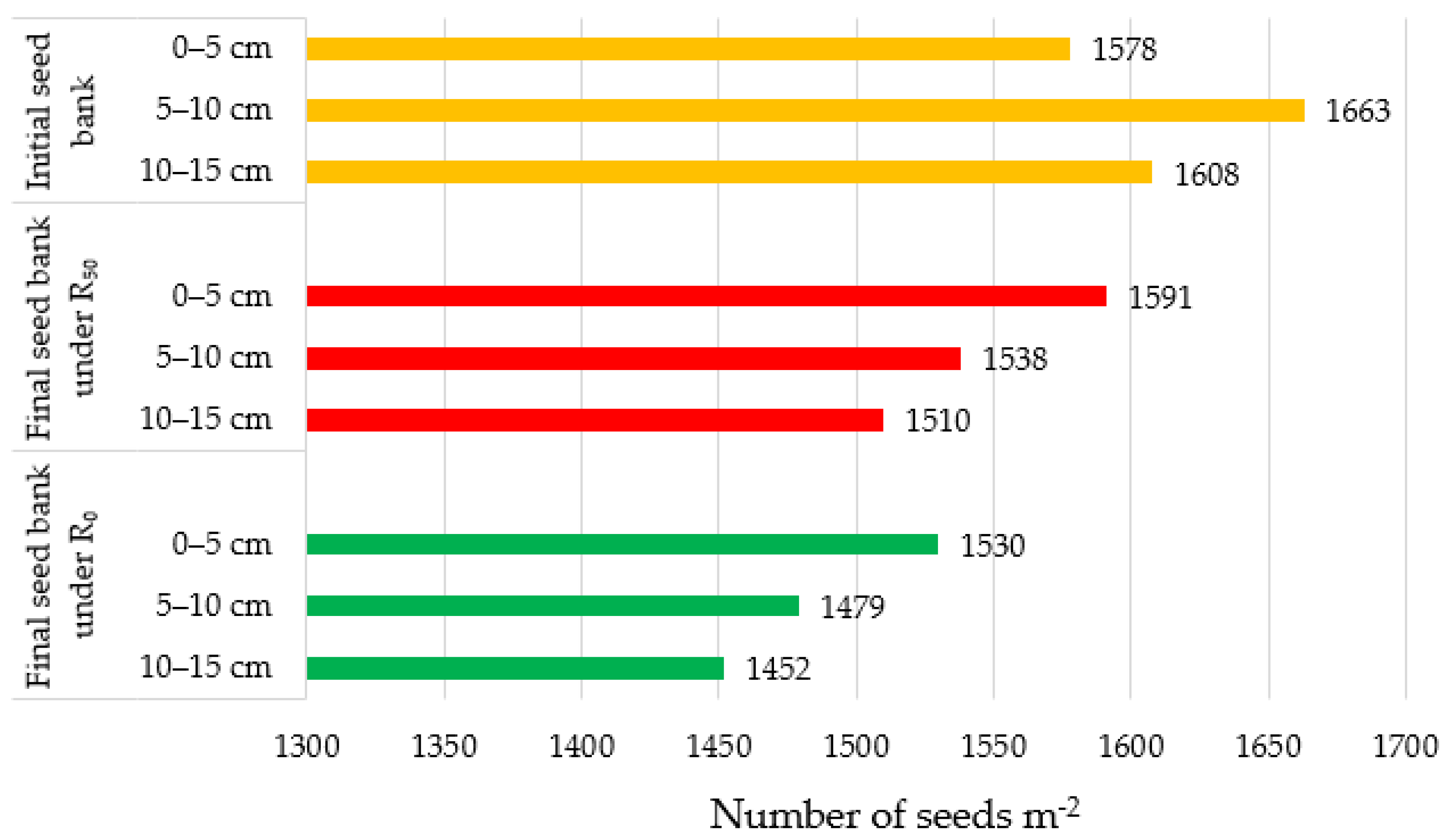
3.4. Effect of Tillage and Residue Levels on the Vertical Distribution of Weed Seeds
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tuesca, D.; Puricelli, E.; Papa, J.C. A Long-Term Study of Weed Flora Shifts in Different Tillage Systems. Weed Res. 2001, 41, 369–382. [Google Scholar] [CrossRef]
- Cardina, J.; Herms, C.P.; Doohan, D.J. Crop Rotation and Tillage System Effects on Weed Seedbanks. Weed Sci. 2002, 50, 448–460. [Google Scholar] [CrossRef]
- Murphy, S.D.; Clements, D.R.; Belaoussoff, S.; Kevan, P.G.; Swanton, C.J. Promotion of Weed Species Diversity and Reduction of Weed Seedbanks with Conservation Tillage and Crop Rotation. Weed Sci. 2006, 54, 69–77. [Google Scholar] [CrossRef]
- Sosnoskie, L.M.; Herms, C.P.; Cardina, J. Weed Seedbank Community Composition in a 35-Yr-Old Tillage and Rotation Experiment. Weed Sci. 2006, 54, 263–273. [Google Scholar] [CrossRef]
- Patterson, D.T. Effects of Environmental Stress on Weed/Crop Interactions. Weed Sci. 1995, 43, 483–490. [Google Scholar] [CrossRef]
- Buhler, D.D.; Kohler, K.A.; Thompson, R.L. Weed Seed Bank Dynamics During a Five-Year Crop Rotation. Weed Technol. 2001, 15, 170–176. [Google Scholar] [CrossRef]
- Sekutowski, T.R.; Smagacz, J. Share of Anthropophytes in the Crop Sequence: Winter Wheat-Maize-Spring Wheat Depending on Tillage System. Acta Agrobot. 2014, 67, 117–122. [Google Scholar] [CrossRef] [Green Version]
- Fracchiolla, M.; Terzi, M.; Frabboni, L.; Caramia, D.; Lasorella, C.; Giorgio, D.D.; Montemurro, P.; Cazzato, E. Influence of Different Soil Management Practices on Ground-Flora Vegetation in an Almond Orchard. Renew. Agric. Food Syst. 2016, 31, 300–308. [Google Scholar] [CrossRef]
- Singh, M.; Bhullar, M.S.; Chauhan, B.S. Influence of Tillage, Cover Cropping, and Herbicides on Weeds and Productivity of Dry Direct-Seeded Rice. Soil Tillage Res. 2015, 147, 39–49. [Google Scholar] [CrossRef]
- Aweto, A. Ecological Succession Theory and Models. In Shifting Cultivation and Secondary Succession in the Tropics; Aweto, A., Ed.; CABI: Wallingford, UK, 2013; pp. 116–130. [Google Scholar] [CrossRef]
- Woźniak, A. Effect of Tillage System on the Structure of Weed Infestation of Winter Wheat. Span. J. Agric. Res. 2018, 16, 14. [Google Scholar] [CrossRef]
- Nichols, V.; Verhulst, N.; Cox, R.; Govaerts, B. Weed Dynamics and Conservation Agriculture Principles: A Review. Field Crop. Res. 2015, 183, 56–68. [Google Scholar] [CrossRef] [Green Version]
- Naresh, R.K.; Singh, D.K.; Singh, P.K. Crop Establishment with Conservation Tillage on Viable Weed Seed Density and Diversity in Soil, Crop and Water Productivity under RWCS in North-West IGP: A Review. Int. J. Curr. Microbiol. Appl. Sci. 2020, 9, 676–690. [Google Scholar] [CrossRef]
- Hashem, A.; Bowran, D.; Piper, T.; Dhammu, H. Resistance of Wild Radish (Raphanus raphanistrum) to Acetolactate Synthase-Inhibiting Herbicides in the Western Australia Wheat Belt. Weed Technol. 2001, 15, 68–74. [Google Scholar] [CrossRef]
- Busi, R.; Powles, S.B. Inheritance of 2,4-D Resistance Traits in Multiple Herbicide-Resistant Raphanus raphanistrum Populations. Plant Sci. 2017, 257, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Curran, W.S. Persistence of Herbicides in Soil. Crop. Soils 2016, 49, 16–21. [Google Scholar] [CrossRef]
- Liebman, M.; Mohler, C. Weeds and the Soil Environment. In Ecological Management of Agricultural Weeds; Liebman, M., Mohler, C., Staver, C.P., Eds.; Cambridge University Press: Cambridge, UK, 2001; pp. 210–268. [Google Scholar]
- Wilson, B.J.; Wright, K.J.; Brain, P.; Clements, M.; Stephens, E. Predicting the Competitive Effects of Weed and Crop Density on Weed Biomass, Weed Seed Production and Crop Yield in Wheat. Weed Res. 1995, 35, 265–278. [Google Scholar] [CrossRef]
- Manici, L.M.; Caputo, F.; Babini, V. Effect of Green Manure on Pythium Spp. Population and Microbial Communities in Intensive Cropping Systems. Plant Soil 2004, 263, 133–142. [Google Scholar] [CrossRef]
- Liebman, M.; Dyck, E. Crop Rotation and Intercropping Strategies for Weed Management. Ecol. Appl. 1993, 3, 92–122. [Google Scholar] [CrossRef] [PubMed]
- Sparks, D.L.; Page, A.L.; Helmke, P.A.; Loeppert, R.H.; Soltanpour, P.N.; Tabatabai, M.A.; Johnston, C.T.; Sumner, M.E. Methods of Soil Analysis; John Wiley & Sons Inc.: Toronto, ON, Canada, 1996. [Google Scholar] [CrossRef]
- Bell, R.W.; Enamul Haque, M.; Jahiruddin, M.; Moshiur Rahman, M.; Begum, M.; Monayem Miah, M.A.; Ariful Islam, M.; Anwar Hossen, M.; Salahin, N.; Zahan, T.; et al. Conservation Agriculture for Rice-Based Intensive Cropping by Smallholders in the Eastern Gangetic Plain. Agriculture 2019, 9, 5. [Google Scholar] [CrossRef] [Green Version]
- Chancellor, R.J. The Identification of Weed Seedlings of Farm and Garden; Blackwell Science Inc.: Oxford, UK, 1981. [Google Scholar]
- Shannon, C.E. A Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Simpson, E.H. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Curtis, J.T. The Vegetation of Wisconsin: An Ordination of Plant Communities; The University of Wisconsin Press: Madison, WI, USA, 1959. [Google Scholar]
- Sørensen, T. A Method of Establishing Groups of Equal Amplitude in Plant Sociology Based on Similarity of Species and Its Application to Analyses of the Vegetation on Danish Commons. R. Dan. Acad. Sci. Lett. 1948, 5, 1–34. [Google Scholar]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research, 2nd ed.; John Wiley and Sons Inc.: New York, NY, USA, 1984. [Google Scholar]
- Dahlin, A.S.; Rusinamhodzi, L. Yield and Labor Relations of Sustainable Intensification Options for Smallholder Farmers in Sub-Saharan Africa. A Meta-analysis. Agron. Sustain. Dev. 2019, 39, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Kleemann, S.G.L.; Gill, G.S. Seed Dormancy and Seedling Emergence in Ripgut Brome (Bromus diandrus) Populations in Southern Australia. Weed Sci. 2013, 61, 222–229. [Google Scholar] [CrossRef]
- Gallandt, E.R.; Fuerst, E.P.; Kennedy, A.C. Effect of Tillage, Fungicide Seed Treatment, and Soil Fumigation on Seed Bank Dynamics of Wild Oat (Avena fatua). Weed Sci. 2004, 52, 597–604. [Google Scholar] [CrossRef]
- Chauhan, B.S.; Singh, R.G.; Mahajan, G. Ecology and Management of Weeds under Conservation Agriculture: A Review. Crop Prot. 2012, 38, 57–65. [Google Scholar] [CrossRef]
- Oziegbe, M.; Faluyi, J.O.; Oluwaranti, A. Effect of Seed Age and Soil Texture on Germination of Some Ludwigia Species (Onagraceae) in Nigeria. Acta Bot. Croat. 2010, 69, 249–257. [Google Scholar]
- White, S.S.; Renner, K.A.; Menalled, F.D.; Landis, D.A. Feeding Preferences of Weed Seed Predators and Effect on Weed Emergence. Weed Sci. 2007, 55, 606–612. [Google Scholar] [CrossRef]
- Barroso, J.; Navarrete, L.; Sánchez Del Arco, M.J.; Fernandez-Quintanilla, C.; Lutman, P.J.W.; Perry, N.H.; Hull, R.I. Dispersal of Avena fatua and Avena sterilis Patches by Natural Dissemination, Soil Tillage and Combine Harvesters. Weed Res. 2006, 46, 118–128. [Google Scholar] [CrossRef]
- Gauvrit, C.; Chauvel, B. Sensitivity of Ambrosia artemisiifolia to Glufosinate and Glyphosate at Various Developmental Stages. Weed Res. 2010, 50, 503–510. [Google Scholar] [CrossRef]
- Tanveer, A.; Nadeem, M.A.; Ali, A.; Tahir, M.; Zamir, M.S.I. Germination Behaviour of Seeds from Herbicide Treated Plants of Chenopodium album L. Ann. Braz. Acad. Sci. 2009, 81, 873–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Opeña, J.L.; Chauhan, B.S.; Baltazar, A.M. Seed Germination Ecology of Echinochloa Glabrescens and Its Implication for Management in Rice (Oryza sativa L.). PLoS ONE 2014, 9, e92261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javaid, M.M.; Tanveer, A.; Ahmad, R.; Yaseen, M.; Khaliq, A. Optimizing Activity of Herbicides at Reduced Rate on Emex spinosa Campd. with Adjuvants. Planta Daninha 2012, 30, 425–435. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Pramanick, B.; Singh, A.; Kumar, S.; Kumar, A.; Singh, G. Bio-Efficacy of Fenoxaprop-p-ethyl for Grassy Weed Control in Onion and Its Residual Effect on Succeeding Maize Crop. Indian J. Weed Sci. 2017, 49, 63–66. [Google Scholar]
- Owen, M.J.; Goggin, D.E.; Powles, S.B. Intensive Cropping Systems Select for Greater Seed Dormancy and Increased Herbicide Resistance Levels in Lolium rigidum (Annual Ryegrass). Pest Manag. Sci. 2015, 71, 966–971. [Google Scholar] [CrossRef]
- Hossain, M.; Begum, M.; Hashem, A.; Rahman, M.; Bell, R. Weed Control in Strip Planted Wheat under Conservation Agriculture Practice Is More Effective than Conventional Tillage. Sci. J. Crop Sci. 2020, 9, 438–450. [Google Scholar] [CrossRef]
- Zahan, T.; Hashem, A.; Rahman, M.; W Bell, R.; Begum, M. Efficacy of Herbicides in Non-Puddled Transplanted Rice under Conservation Agriculture Systems and Their Effect on Establishment of the Succeeding Crops. Acta Sci. Malays. 2018, 2, 17–25. [Google Scholar] [CrossRef]
- Zhang, J.; Wu, L.F. Impact of Tillage and Crop Residue Management on the Weed Community and Wheat Yield in a Wheat–Maize Double Cropping System. Agriculture 2021, 11, 265. [Google Scholar] [CrossRef]
- Chhokar, R.S.; Sharma, R.K.; Gill, S.C.; Singh, R.K.; Joon, V.; Kajla, M.; Chaudhary, A. Suitable Wheat Cultivars and Seeding Machines for Conservation Agriculture in Rice-Wheat and Sugarcane-Wheat Cropping Systems. Wheat Barley Res. 2018, 10, 78–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chauhan, B.S.; Abugho, S.B. Effect of Crop Residue on Seedling Emergence and Growth of Selected Weed Species in a Sprinkler-Irrigated Zero-Till Dry-Seeded Rice System. Weed Sci. 2013, 61, 403–409. [Google Scholar] [CrossRef]
- Conn, J.S. Weed Seed Bank Affected by Tillage Intensity for Barley in Alaska. Soil Tillage Res. 2006, 90, 156–161. [Google Scholar] [CrossRef]
- Cardina, J.; Regnier, E.; Harrison, K. Long-Term Tillage Effects on Seed Banks in Three Ohio Soils. Weed Sci. 1991, 39, 186–194. [Google Scholar] [CrossRef]
- Borin, M.; Zanin, G.; Zuin, M.C. The Comparison of Seed Banks in Conventional and Ridge-Tilled Soils In North-Eastern Italy. In Soil Management in Sustainable Agriculture; Cook, H.F., Lee, H.C., Eds.; Wye College Press: Ashford, UK, 1995; pp. 544–555. [Google Scholar]
- Feldman, S.R.; Torres, C.A.P.S.; Lewis, P. The Effect of Different Tillage Systems on the Composition of the Seedbank. Weed Res. 1997, 37, 71–76. [Google Scholar] [CrossRef]
- Feledyn-Szewczyk, B.; Smagacz, J.; Kwiatkowski, C.A.; Harasim, E.; Woźniak, A. Weed Flora and Soil Seed Bank Composition as Affected by Tillage System in Three-Year Crop Rotation. Agriculture 2020, 10, 186. [Google Scholar] [CrossRef]
- Zanin, G.; Otto, S.; Riello, L.; Borin, M. Ecological Interpretation of Weed Flora Dynamics under Different Tillage Systems. Agric. Ecosyst. Environ. 1997, 66, 177–188. [Google Scholar] [CrossRef]
- Hossain, M.M.; Begum, M.; Hashem, A.; Moshiur, R.; Bell, R. Weed Populations of Intensive Rice Based Cropping System as Affected by Tillage and Increased Crop Residues in Bangladesh. J. Agric. Nat. Resour. Sci. 2021, 8, 1–12. [Google Scholar] [CrossRef]
- Buhler, D.D.; Stoltenberg, D.E.; Becker, R.L.; Gunsolus, J.L. Perennial Weed Populations After 14 Years of Variable Tillage and Cropping Practices. Weed Sci. 1994, 42, 205–209. [Google Scholar] [CrossRef]
- Hossain, M.M.; Begum, M.; Rahman, M. The Behavior of Weed Seed Bank to Different Tillage and Residue Mulch Treatments after Three Years of Cropping in Bangladesh. Azarian J. Agric. 2021, 8, 29–37. [Google Scholar] [CrossRef]
- Thomas, A.G.; Derksen, D.A.; Blackshaw, R.E.; Acker, R.C.V.; Légère, A.; Watson, P.R.; Turnbull, G.C. A Multistudy Approach to Understanding Weed Population Shifts in Medium- to Long-Term Tillage Systems. Weed Sci. 2004, 52, 874–880. [Google Scholar] [CrossRef]
- Piskier, T.; Sekutowski, T.R. Effect of Simplified Tillage on the Number and Distribution of Weed Seeds in Soil. J. Res. Appl. Agric. Eng. 2013, 58, 109–117. [Google Scholar]
- Clements, D.R.; Benoit, D.L.; Murphy, S.D.; Swanton, C.J. Tillage Effects on Weed Seed Return and Seedbank Composition. Weed Sci. 1996, 44, 314–322. [Google Scholar] [CrossRef]
- Hoffman, M.L.; Owen, M.D.K.; Buhler, D.D. Effects of Crop and Weed Management on Density and Vertical Distribution of Weed Seeds in Soil. Agron. J. 1998, 90, 793–799. [Google Scholar] [CrossRef]
- Bàrberi, P.; Bonari, E.; Mazzoncini, M. Weed Density and Composition in Winter Wheat as Influenced by Tillage Systems. In Proceedings of the Conservation Agriculture, a Worldwide Challenge, First World Congress on Conservation Agriculture, Madrid, Spain, 1–5 October 2001; Volume 2, pp. 451–455. [Google Scholar]
- Bàrberi, P.; Lo Cascio, B. Long-Term Tillage and Crop Rotation Effects on Weed Seedbank Size and Composition. Weed Res. 2001, 41, 325–340. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Properties | Values |
|---|---|
| pH | 6.69 |
| Total nitrogen (%) | 0.11 |
| Available phosphorus (mg kg−1) | 16.2 |
| Exchangeable potassium (cmol kg−1) | 0.31 |
| Available sulfur (mg kg−1) | 14.0 |
| Herbicides | Crop | Application Rate of Product (ha−1) | Time of Application 1 |
|---|---|---|---|
| Glyphosate | All crops | 3.7 L | 3 DBS/T |
| Pendimethalin | All crops | 2.7 L | 3 DAT/S in rice and wheat IAS in mustard and mungbean |
| Ethoxysulfuron-ethyl | Rice | 100 g | 3 WAT |
| Carfentrazone-ethyl + isoproturon | Wheat | 1.25 kg | 3 WAS |
| Isoproturon | Mustard | 650 mL | 3 WAS |
| Fenoxaprop-p-ethyl | Mungbean | 650 mL | 3 WAS |
| Weed Species | Type | Ontogeny | Initial Seed Bank (Seeds m−2) | Final Seed Bank (Seeds m−2) | |||
|---|---|---|---|---|---|---|---|
| CT R0 | CT R50 | ST R0 | ST R50 | ||||
| Ageratum conyzoides L. | B | P | 170 | 194 | 169 | 239 | Absent |
| Alternanthera sessilis L. | B | P | 173 | 203 | 184 | 181 | 168 |
| A. philoxeroides L. | B | P | 160 | 190 | 165 | 157 | 146 |
| A. spinosus L. | B | A | 136 | 141 | 122 | 114 | Absent |
| Amaranthus viridis L. | B | A | 118 | 157 | 136 | Absent | Absent |
| Brassica kaber L. | B | A | 89 | 136 | 147 | Absent | Absent |
| Centipeda minima Lour. | B | P | Absent | 103 | 89 | 125 | 116 |
| Cyanotis axillaris Roem. | B | A | 288 | 94 | 167 | 186 | Absent |
| Cynodon dactylon L. | G | P | 129 | Absent | 201 | 234 v | 315 |
| Cyperus difformis L. | S | A | 348 | 381 iii | 337 | 148 | 137 |
| C. iria L. | S | A | 209 | 123 | Absent | Absent | 110 |
| C. rotundus L. | S | P | 161 | 259 | 112 | 303 ii | 381 |
| Dentella repens L. | B | P | 142 | 142 | 123 | Absent | Absent |
| Desmodium triflorum L. | B | P | 116 | 124 | 108 | Absent | Absent |
| Digitaria sanguinalis L. | G | A | 137 | 154 | 134 | Absent | 231 |
| Echinochloa crus-galli (L.) Beauv. | G | A | 351 | 512 i | 445 | 169 | 157 |
| E. colonum L. | G | A | Absent | 131 | 114 | Absent | 184 |
| Eclipta alba L. | B | A | 98 | 166 | 145 | Absent | Absent |
| Eichhornia crassipes Mart. | B | P | 102 | 188 | 163 | 135 | 146 |
| Eleocharis atropurpurea Ret. | S | A | 83 | 47 | Absent | Absent | Absent |
| Eleusine indica L. | G | A | 165 | 208 | 179 | Absent | 127 |
| Euphorbia parviflora L. | B | A | 104 | 150 | 129 | Absent | Absent |
| Fimbristylis miliacea (L.) Vahl. | S | A | 275 | 302 iv | 258 | 253 iv | 214 |
| Gnaphalium luteo-album L. | B | A | 99 | Absent | Absent | Absent | Absent |
| Hedyotis corymbosa L. | B | A | 144 | 202 | 175 | Absent | Absent |
| Jussia decurrence Walt. | B | P | 261 | 490 ii | 426 | 356 i | 330 |
| Leersia hexandra L. | G | P | 196 | 236 | 209 | 271 iii | 252 |
| Lindernia antipoda L. | B | A | 139 | 134 | Absent | Absent | 212 |
| L. hyssopifolia L. | B | A | 103 | 113 | Absent | Absent | 209 |
| Marsilea quadrifolia L. | B | A | 122 | 152 | Absent | 110 | 102 |
| Monochoria hastate L. | B | P | 115 | 163 | 187 | 157 | 146 |
| Nicotina plumbaginifolia L. | B | A | 174 | 217 | 198 | 141 | Absent |
| Panicum distichum L. | G | P | 73 | Absent | Absent | 158 | 133 |
| Physalis heterophylla Nees. | B | A | Absent | Absent | 167 | Absent | Absent |
| Pistia stratiotes L. | B | P | 163 | 132 | 114 | 123 | Absent |
| Polygonum coccineum L. | B | A | Absent | 173 | 150 | Absent | Absent |
| Rotala ramosior L. | B | A | Absent | 213 | 185 | Absent | Absent |
| Scirpus mucronatus (L.) Palla. | S | P | 199 | Absent | 121 | 198 | Absent |
| S. juncoides L. | S | P | Absent | 99 | 86 | 137 | Absent |
| S. supinus L. | S | P | Absent | 274 v | 238 | 119 | Absent |
| Solanum torvum Sw. | B | P | Absent | Absent | Absent | 171 | Absent |
| Spilanthes acmella L. | B | A | Absent | 227 | 198 | Absent | Absent |
| Total number of species | 33 | 36 | 34 | 24 | 19 | ||
| Shannon’s diversity index (H’) | 3.41 | 3.47 | 3.44 | 3.07 | 2.92 | ||
| Simpson’s dominance index (SI) | 0.36 | 0.34 | 0.35 | 0.49 | 0.58 | ||
| Weed Species | Type | Ontogeny | Initial Seed Bank (Plants m−2) | Final Seed Bank (Plants m−2) | |||
|---|---|---|---|---|---|---|---|
| CT R0 | CT R50 | ST R0 | ST R50 | ||||
| Ageratum conyzoides L. | B | P | 89 | 128 | 110 | 137 | 127 |
| Alternanthera sessilis L. | B | P | 71 | 120 | 133 | 149 | 138 |
| A. philoxeroides L. | B | P | 64 | 115 | 115 | 115 | 92 |
| A. spinosus L. | B | A | 78 | 99 | 125 | 103 | 91 |
| Amaranthus viridis L. | B | A | 67 | 112 | 129 | 136 | Absent |
| Brassica kaber L. | B | A | 125 | 81 | 178 | Absent | Absent |
| Centipeda minima Lour. | B | P | Absent | 139 | 123 | 157 | 146 |
| Cyanotis axillaris Roem. | B | A | 117 | 234 iv | 195 | 116 | Absent |
| Cynodon dactylon L. | G | P | 105 | Absent | Absent | Absent | Absent |
| Cyperus difformis L. | S | A | 125 | 91 | 188 | 172 | 160 |
| C. iria L. | S | A | 185 | 108 | 80 | Absent | Absent |
| C. rotundus L. | S | P | 307 | 292 ii | 180 | 324 ii | 301 |
| Dentella repens L. | B | P | 136 | 84 | 103 | Absent | Absent |
| Desmodium triflorum L. | B | P | 116 | 97 | 115 | Absent | Absent |
| Digitaria sanguinalis L | G | A | 131 | 127 | 160 | Absent | Absent |
| Echinochloa crus-galli (L.) Beauv. | G | A | 352 | 363 i | 334 | 249 iii | 232 |
| E. colonum L. | G | A | Absent | 197 | 107 | Absent | Absent |
| Eclipta alba L. | B | A | 117 | 73 | 101 | 88 | 107 |
| Eichhornia crassipes Mart. | B | P | 124 | 111 | 189 | 99 | 111 |
| Eleusine indica L. | G | A | 225 | 263 iii | 152 | 235 | 218 |
| Euphorbia parviflora L. | B | A | 95 | 140 | 193 | Absent | Absent |
| Fimbristylis miliacea (L.) Vahl. | S | A | 253 | 224 v | 115 | 115 | 82 |
| Jussia decurrence Walt. | B | P | 227 | 120 | 93 | 179 | 166 |
| J. repens L. | B | P | 94 | 119 | 70 | 107 | 99 |
| Leersia hexandra L. | G | P | 110 | 135 | 107 | 371 i | 345 |
| Lindernia antipoda (L.) Aston. | B | A | 81 | 226 | Absent | 121 | Absent |
| L. hyssopifolia L. | B | A | 125 | 79 | Absent | 71 | Absent |
| Panicum distichum L. | S | P | 87 | Absent | Absent | Absent | Absent |
| Pistia stratiotes L. | B | P | 112 | 67 | 197 | Absent | Absent |
| Polygonum coccineum L. | B | A | 99 | 102 | 164 | Absent | Absent |
| Rotala ramosior (L.) Koch. | B | A | 219 | 97 | 134 | Absent | Absent |
| Scirpus mucronatus (L.) Palla. | S | P | Absent | Absent | Absent | 225 iv | 209 |
| S. juncoides L. | S | P | Absent | 119 | 147 | Absent | Absent |
| Solanum torvum Sw. | B | P | 77 | 63 | 122 | 215 v | 199 |
| Total number of species | 27 | 31 | 29 | 21 | 17 | ||
| Shannon’s diversity index (H’) | 3.18 | 3.32 | 3.31 | 2.94 | 2.77 | ||
| Simpson’s dominance index (SI) | 0.47 | 0.41 | 0.38 | 0.58 | 0.71 | ||
| Weed Species | Type | Ontogeny | Initial Seed Bank (Seeds m−2) | Final Seed Bank (Seeds m−2) | |||
|---|---|---|---|---|---|---|---|
| CT R0 | CT R50 | ST R0 | ST R50 | ||||
| Ageratum conyzoides L. | B | P | 117 | 95 ii | 100 | 97 v | 74 |
| A. philoxeroides L. | B | P | 92 | 19 | 64 | 114 iv | 73 |
| Alternanthera sessilis L. | B | P | 84 | 78 | 57 | 75 | 66 |
| Amaranthus viridis L. | B | A | 51 | 74 | 85 | Absent | Absent |
| C. rotundus L. | S | P | 82 | 74 | 58 | 109 i | 167 |
| Cynodon dactylon L. | G | P | 63 | 67 | 70 | 173 ii | 59 |
| Cyperus difformis L. | S | A | 99 | 88 | 69 | Absent | Absent |
| Dentella repens L. | B | P | 103 | 28 | 73 | 96 | 51 |
| Desmodium triflorum L. | B | P | 97 | 81 | 73 | Absent | 69 |
| E. colonum L. | G | A | 74 | 87 | 54 | 97 | Absent |
| Echinochloa crus-galli (L.) Beauv. | G | A | 68 | 81 i | 129 | 69 | 48 |
| Eichhornia crassipes Mart. | B | P | 62 | 55 | 34 | 85 | 31 |
| Euphorbia parviflora L. | B | A | 86 | 79 | 85 | Absent | Absent |
| Fimbristylis miliacea (L.) Vahl. | S | A | 69 | 93 | 44 | 67 | 43 |
| Jussia decurrence Walt. | B | P | 53 | 84v | 77 | 118 iii | 96 |
| Lindernia antipoda (L.) Aston. | B | A | 111 | 73 | 82 | Absent | Absent |
| Physalis heterophylla Nees. | B | A | 120 | 30 | Absent | Absent | Absent |
| Pistia stratiotes L. | B | P | 112 | 89 | 54 | 62 | Absent |
| Rotala ramosior (L.) Koch. | B | A | 75 | 79iv | 83 | Absent | Absent |
| Scirpus mucronatus (L.) Palla. | S | P | Absent | 62 iii | 119 | 72 | 62 |
| Total number of species | 19 | 20 | 19 | 14 | 12 | ||
| Shannon’s diversity index (H’) | 2.91 | 2.93 | 2.89 | 2.49 | 2.44 | ||
| Simpson’s dominance index (SI) | 0.55 | 0.54 | 0.57 | 0.91 | 0.90 | ||
| Weed Species | Type | Ontogeny | Wheat-Mungbean-Monsoon Rice Rotation | Monsoon Rice-Mustard-Winter Rice Rotation |
|---|---|---|---|---|
| Ageratum conyzoides L. | B | Perennial | 182 | 150 |
| Alternanthera sessilis L. | B | Perennial | 194 | 166 |
| A. philoxeroides L. | B | Perennial | 178 | 150 |
| A. spinosus L. | B | Annual | 132 | 120 |
| Amaranthus viridis L. | B | Annual | 147 | Absent |
| Brassica kaber L. | B | Annual | 142 | Absent |
| Centipeda minima Lour. | B | Perennial | 96 | 129 |
| Cyanotis axillaris Roem. | B | Annual | 131 | 328 |
| Cynodon dactylon L. | G | Perennial | 101 | 165 |
| Cyperus difformis L. | S | Annual | 359 | 31 |
| C. iria L. | S | Annual | 62 | 67 |
| C. rotundus L. | S | Perennial | 186 | 108 |
| Dentella repens L. | B | Perennial | 133 | Absent |
| Desmodium triflorum L. | B | Perennial | 116 | 120 |
| Digitaria sanguinalis L. | G | Annual | 144 | 120 |
| Echinochloa crus-galli (L.) Beauv. | G | Annual | 479 | 238 |
| E. colonum L. | G | Annual | 123 | 65 |
| Eclipta alba L. | B | Annual | 156 | 136 |
| Eichhornia crassipes Mart. | B | Perennial | 176 | 166 |
| Eleocharis atropurpurea Ret. | S | Annual | 24 | 236 |
| Eleusine indica L. | G | Annual | 194 | 164 |
| Euphorbia parviflora L. | B | Annual | 140 | 152 |
| Fimbristylis miliacea (L.) Vahl. | S | Annual | 280 | 310 |
| Gnaphalium luteo-album L. | B | Annual | Absent | 160 |
| Hedyotis corymbosa L. | B | Annual | 189 | 150 |
| Jussia decurrence Walt. | B | Perennial | 458 | 163 |
| Leersia hexandra L. | G | Perennial | 223 | 310 |
| Lindernia antipoda L. | B | Annual | 67 | 161 |
| L. hyssopifolia L. | B | Annual | 57 | 191 |
| Marsilea quadrifolia L. | B | Annual | 76 | 151 |
| Monochoria hastate L. | B | Perennial | 175 | 139 |
| Nicotina plumbaginifolia L. | B | Annual | 208 | 153 |
| Panicum distichum L. | G | Perennial | Absent | 121 |
| Physalis heterophylla Nees. | B | Annual | 84 | 190 |
| Pistia stratiotes L. | B | Perennial | 123 | 55 |
| Polygonum coccineum L. | B | Annual | 162 | 161 |
| Rotala ramosior L. | B | Annual | 199 | 108 |
| Scirpus mucronatus L. | S | Perennial | 61 | 120 |
| S. juncoides L. | S | Perennial | 93 | Absent |
| S. supinus L. | S | Perennial | 256 | Absent |
| Solanum torvum Sw. | B | Perennial | Absent | 55 |
| Spilanthes acmella L. | B | Annual | 213 | Absent |
| Total number of species | 39 | 33 | ||
| Total number of plants m−2 | 6506 | 5498 | ||
| Shannon’s diversity index (H’) | 3.51 | 3.47 | ||
| Simpson’s dominance index (SI) | 0.32 | 0.35 | ||
| Tillage at the Different Soil Depth | Initial Seed Bank | Final Seed Bank of CT | Final Seed Bank of ST | |
|---|---|---|---|---|
| 0–5 cm | Initial seed pool | - | 82 | 67 |
| Final seed pool of CT | - | - | 60 | |
| 5–10 cm | Initial seed pool | - | 96 | 74 |
| Final seed pool of CT | - | - | 80 | |
| 10–15 cm | Initial seed pool | - | 98 | 71 |
| Final seed pool of CT | - | - | 75 | |
| Crop residue at the different soil depth | Final seed bank of CT R0 | Final seed bank of ST R0 | ||
| 0–5 cm | Final seed bank of R50 | 83 | 31 | |
| 5–10 cm | Final seed bank of R50 | 96 | 89 | |
| 10–15 cm | Final seed bank of R50 | 97 | 86 | |
| Weed Type | Soil Depth (cm) | Initial Seed Bank (Seeds m−2) | Final Seed Bank (Seeds m−2) | |||
|---|---|---|---|---|---|---|
| CT R0 | CT R50 | ST R0 | ST R50 | |||
| Broadleaf | 0–5 | 3016 a | 850 g,h | 896 g,h | 2195 a | 1708 b |
| 5–10 | 2233 b | 2406 c,d | 2589 c | 1793 b | 1276 c | |
| 10–15 | 1163 c,d | 4204 a | 3647 b | 585 g | 517 g | |
| Grass | 0–5 | 1051 d | 249 j | 224 j,k | 832 e | 1266 c |
| 5–10 | 1010 d | 1014 g | 706 h | 855 e | 795 e,f | |
| 10–15 | 205 f | 1241 e,f | 1282 e,f | 339 h | 107 j | |
| Sedge | 0–5 | 1275 c | 317 i | 290 i | 1158 c,d | 842 e |
| 5–10 | 870 e | 905 g | 864 g,h | 836 e | 752 f | |
| 10–15 | 250 f | 1485 e | 1152 f | 248 i | 272 i | |
| Standard Deviation | 896.1 | 1229.8 | 1122.2 | 645.15 | 512.21 | |
| Standard Error | 298.6 | 409.9 | 374.08 | 215.05 | 170.73 | |
| Coefficient of Variance (%) | 72.83 | 87.35 | 86.69 | 65.67 | 61.18 | |
| Weed Type | Soil Depth (cm) | Initial Seed Bank (g m−2) | Final Seed Bank (g m−2) | |||
|---|---|---|---|---|---|---|
| CT R0 | CT R50 | ST R0 | ST R50 | |||
| Broadleaf | 0–5 | 1719 a | 484 e | 510 d,e | 1251 a | 973 a |
| 5–10 | 1272 b | 1371 c | 1475 c | 1022 b | 727 b | |
| 10–15 | 663 c | 2396 a | 2078 b | 333 d,e | 294 d,e | |
| Grass | 0–5 | 462 d | 109 g,f | 98 f | 366 d | 557 c |
| 5–10 | 444 d | 446 e,f | 310 f | 376 d | 349 d | |
| 10–15 | 90 f | 546 d,e | 564 d | 149 e,f | 47 f | |
| Sedge | 0–5 | 599 c,d | 148 g | 136 g | 544 c | 395 d |
| 5–10 | 408 d | 425 e | 406 e,f | 392 d | 353 d | |
| 10–15 | 117 e | 697 d | 541 d,e | 127 e,f | 116 e,f | |
| Standard Deviation | 532.16 | 722.71 | 661.08 | 384.21 | 289.28 | |
| Standard Error | 177.38 | 240.90 | 220.36 | 128.07 | 96.42 | |
| Coefficient of Variance (%) | 82.89 | 98.16 | 97.18 | 75.96 | 68.04 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hossain, M.M.; Begum, M.; Hashem, A.; Rahman, M.M.; Ahmed, S.; Hassan, M.M.; Javed, T.; Shabbir, R.; Hadifa, A.; Sabagh, A.E.; et al. Strip Tillage and Crop Residue Retention Decrease the Size but Increase the Diversity of the Weed Seed Bank under Intensive Rice-Based Crop Rotations in Bangladesh. Agronomy 2021, 11, 1164. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11061164
Hossain MM, Begum M, Hashem A, Rahman MM, Ahmed S, Hassan MM, Javed T, Shabbir R, Hadifa A, Sabagh AE, et al. Strip Tillage and Crop Residue Retention Decrease the Size but Increase the Diversity of the Weed Seed Bank under Intensive Rice-Based Crop Rotations in Bangladesh. Agronomy. 2021; 11(6):1164. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11061164
Chicago/Turabian StyleHossain, Mohammad Mobarak, Mahfuza Begum, Abul Hashem, Md. Moshiur Rahman, Sharif Ahmed, Montaser M. Hassan, Talha Javed, Rubab Shabbir, Adel Hadifa, Ayman EL Sabagh, and et al. 2021. "Strip Tillage and Crop Residue Retention Decrease the Size but Increase the Diversity of the Weed Seed Bank under Intensive Rice-Based Crop Rotations in Bangladesh" Agronomy 11, no. 6: 1164. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11061164