
The Role of OsWRKY Genes in Rice When Faced with Single and Multiple Abiotic Stresses


 ,
,  , ,
, ,  ,
, 
 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. In Silico Mining and Meta-Analysis of WRKY TF Genes in O. sativa
2.2. Spatio-Temporal and Phytohormone Expression Analysis
2.3. Gene Features and Phylogenetic Analysis
2.4. Gene Structure Analysis and Gene Ontology Annotation
2.5. Molecular Interactome and Enrichment Analysis
2.6. Gene Synteny Analysis
3. Results
3.1. Identification of the WRKY Family Transcription Factors in Rice Genome
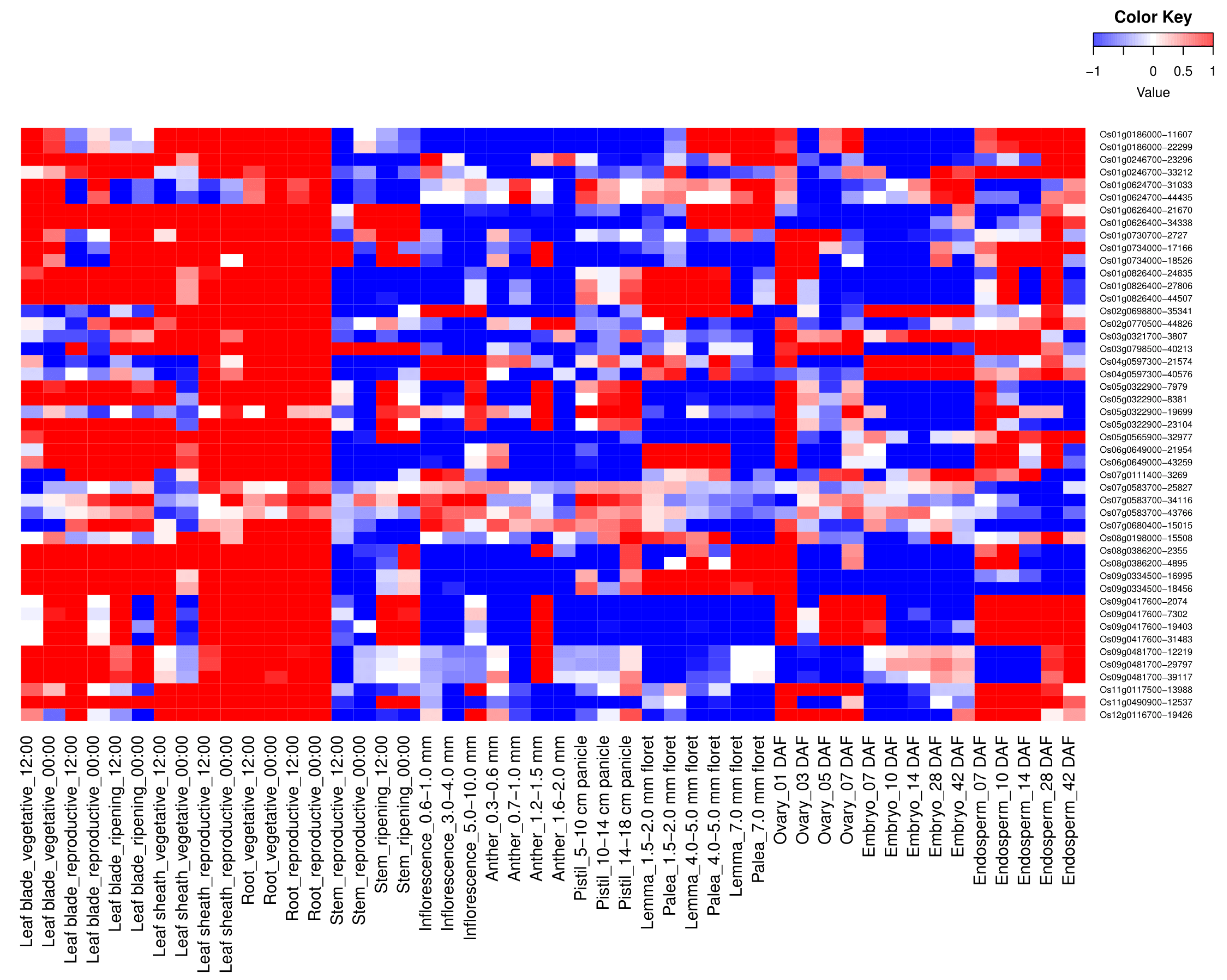
3.2. Spatio-Temporal Expression Analysis of OsWRKY
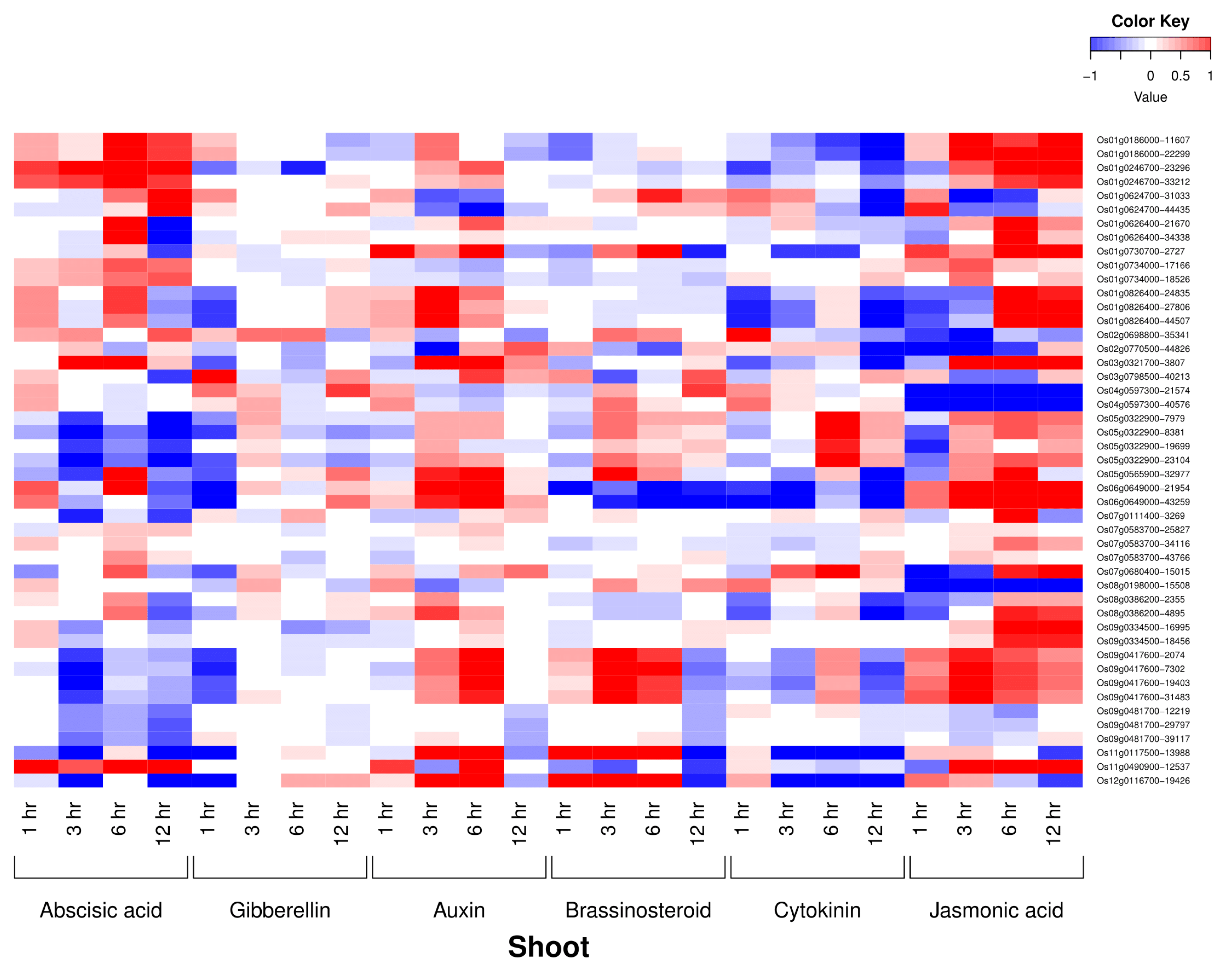
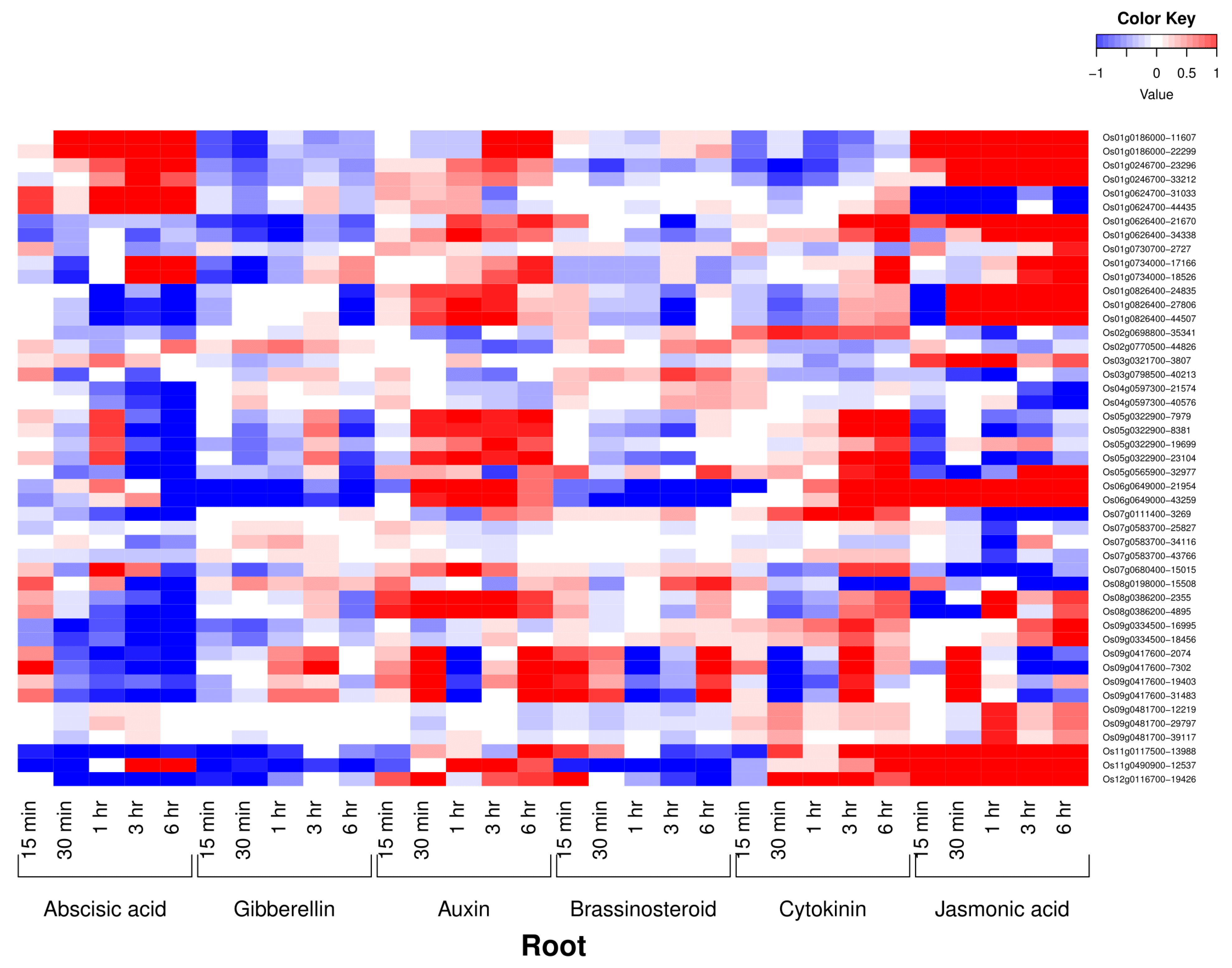
3.3. Phytohormone Expression Profiling
3.4. OsWRKY TF Genes with Their Properties
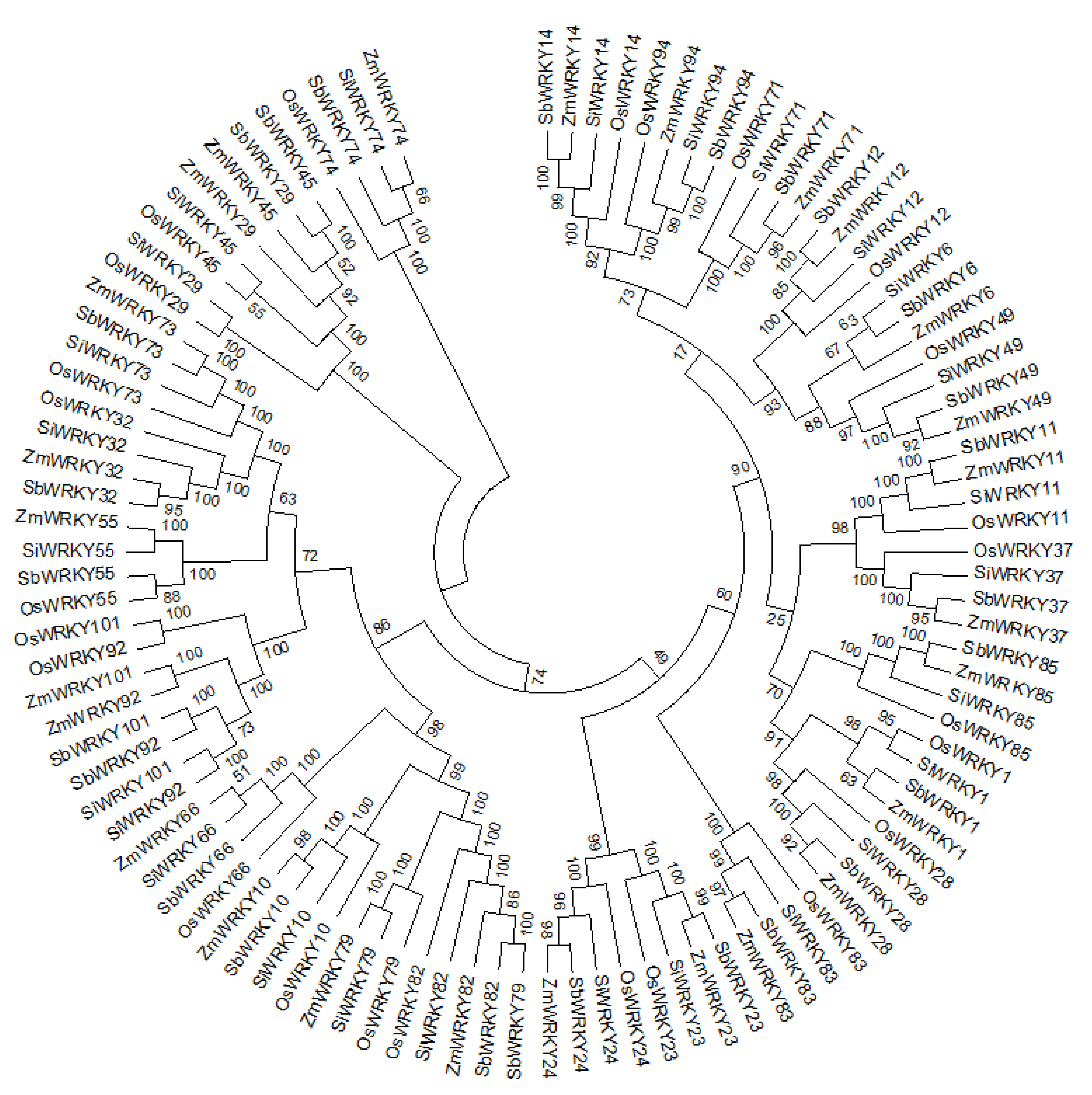
3.5. Phylogenetic Analysis of WRKY TFs
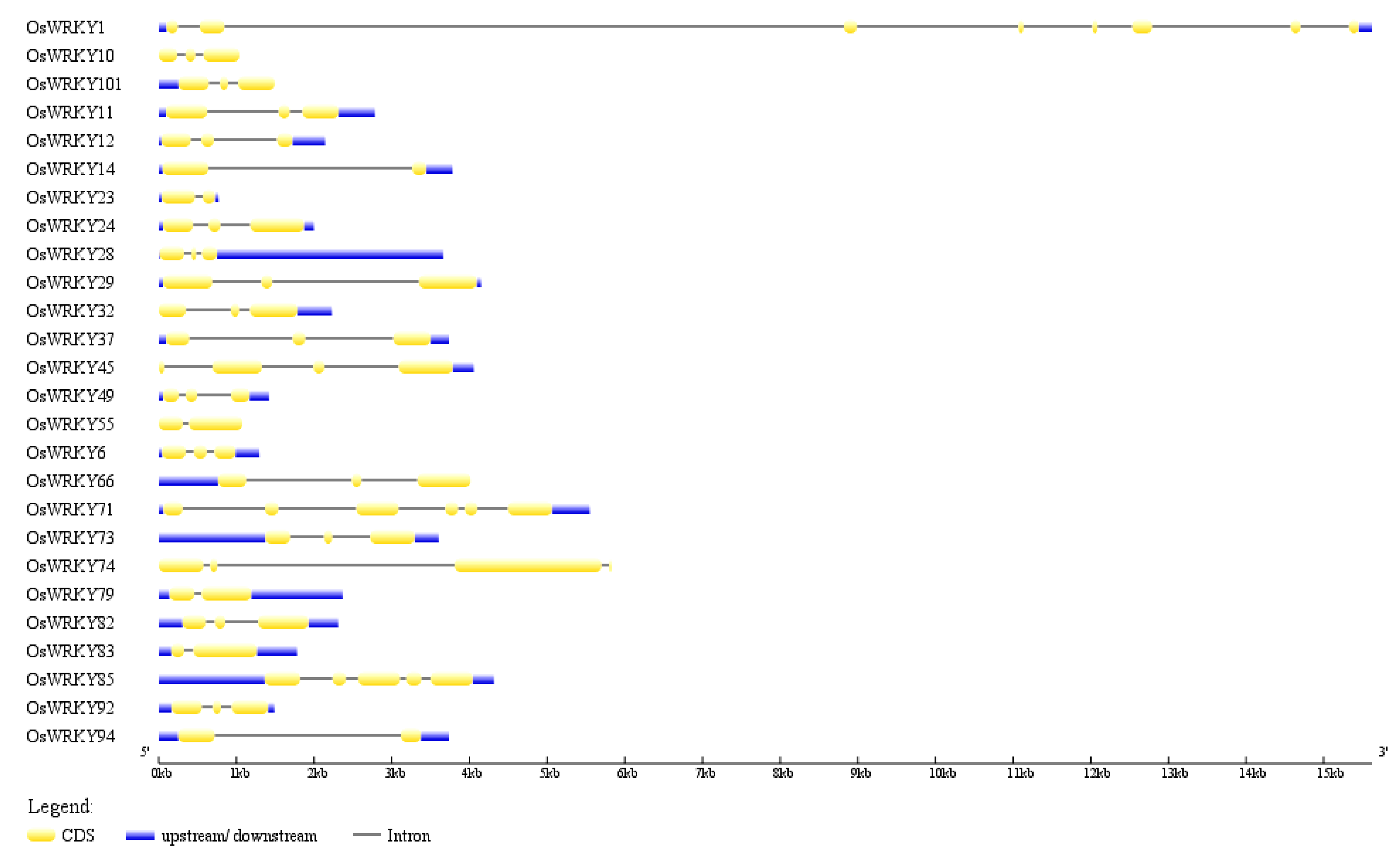
3.6. Gene Organization Analysis
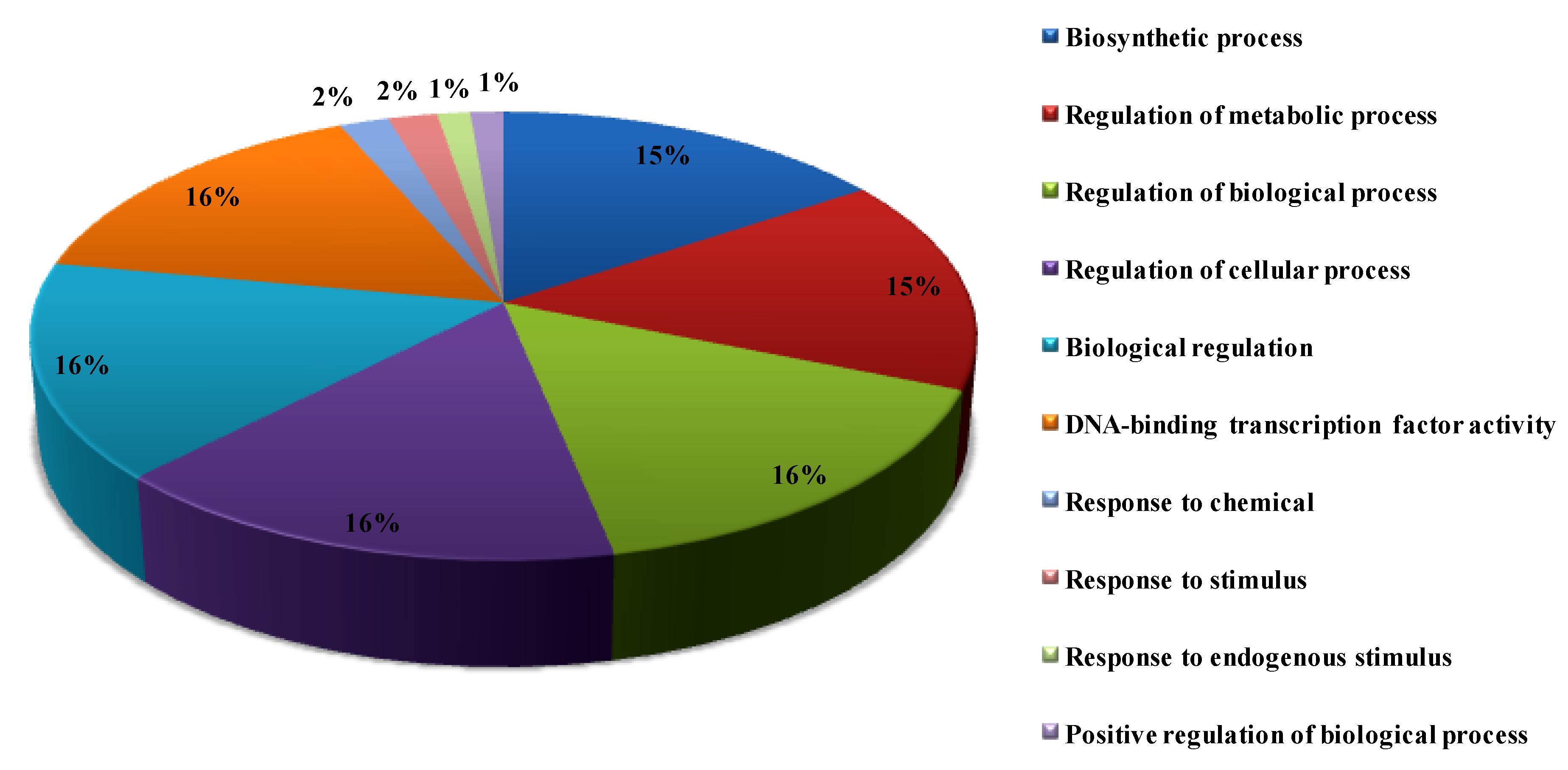
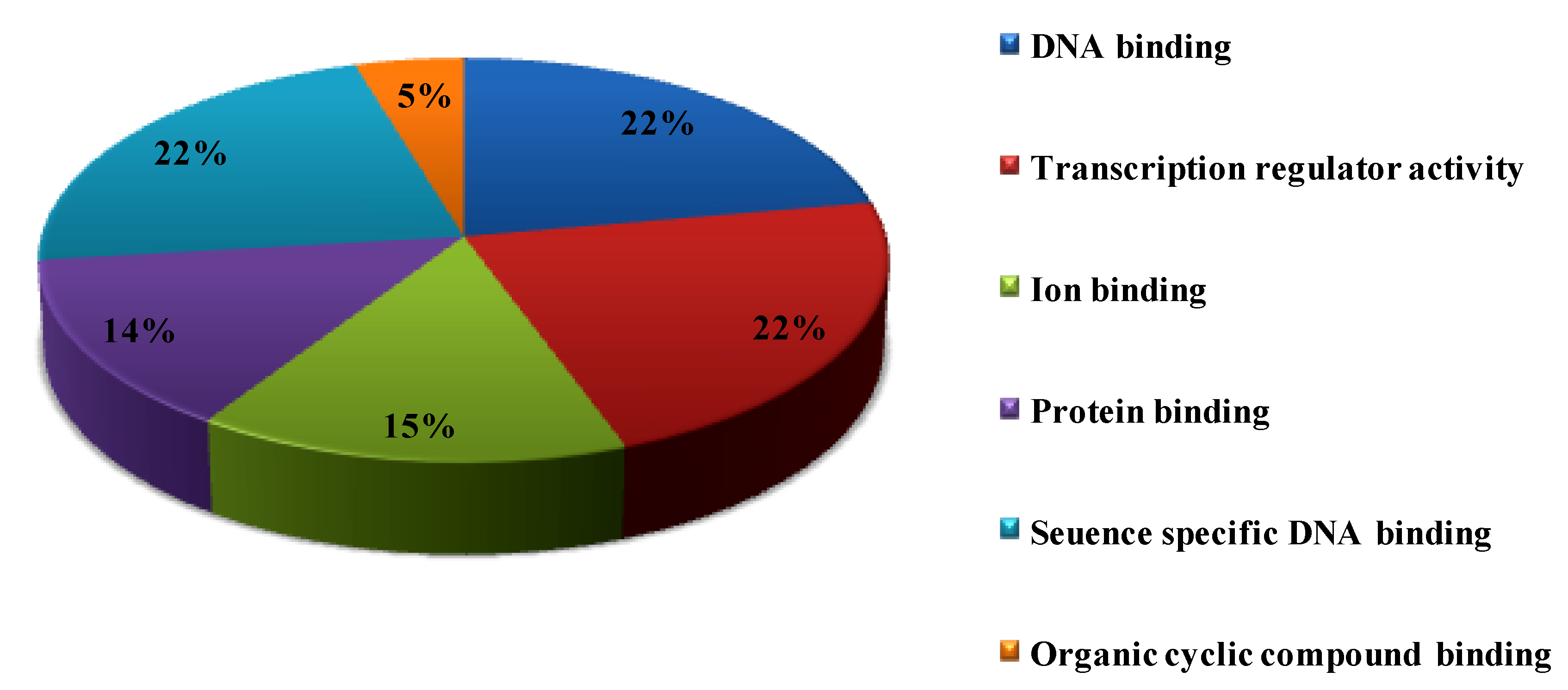
3.7. Functional GO Analysis of OsWRKY TFs
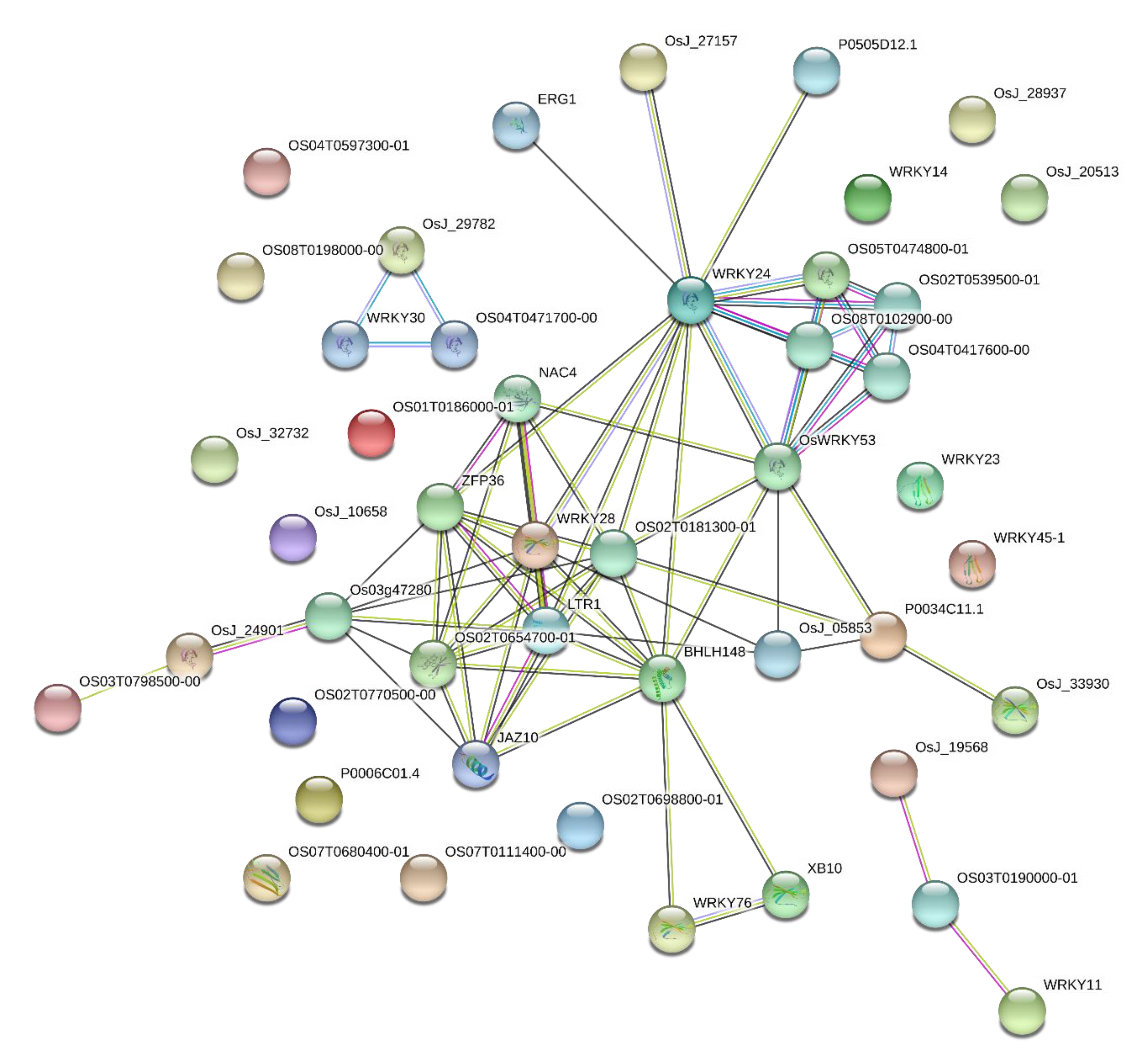
3.8. OsWRKY Gene Interaction Network Analysis
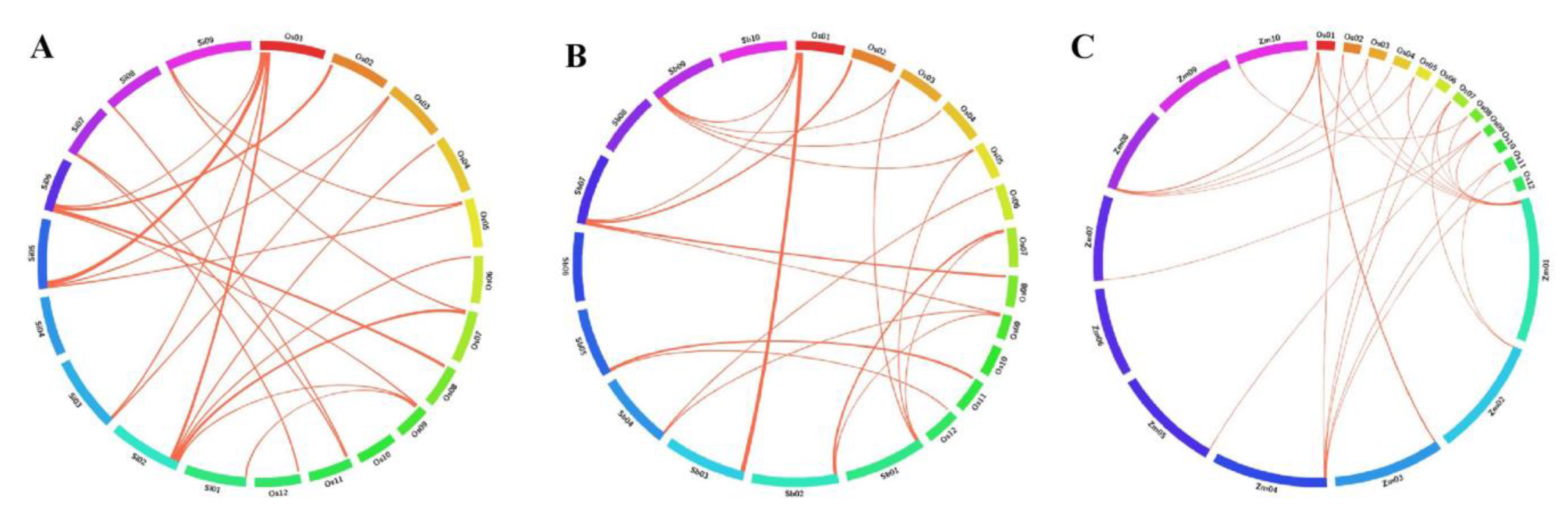
3.9. Orthologous Relationships of OsWRKY Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Calicioglu, O.; Flammini, A.; Bracco, S.; Bellù, L.; Sims, R. The Future Challenges of Food and Agriculture: An Integrated Analysis of Trends and Solutions. Sustainability 2019, 11, 222. [Google Scholar] [CrossRef] [Green Version]
- Mickelbart, M.V.; Hasegawa, P.M.; Bailey-Serres, J. Genetic Mechanisms of Abiotic Stress Tolerance that Translate to Crop Yield Stability. Nat. Rev. Genet. 2015, 16, 237. [Google Scholar] [CrossRef]
- Muthuramalingam, P.; Jeyasri, R.; Kalaiyarasi, D.; Pandian, S.; Krishnan, S.R.; Satish, L.; Pandian, S.K.; Ramesh, M. Emerging Advances in Computational Omics Tools for Systems Analysis of Gramineae Family Grass Species and Their Abiotic Stress Responsive Functions. OMICS-Based Approach Plant. Biotechnol. 2019, 185, 185. [Google Scholar]
- Muthuramalingam, P.; Krishnan, S.R.; Pandian, S.; Mareeswaran, N.; Aruni, W.; Pandian, S.K.; Ramesh, M. Global Analysis of Threonine Metabolism Genes Unravel Key Players in Rice to Improve the Abiotic Stress Tolerance. Sci. Rep. 2018, 8, 9270. [Google Scholar] [CrossRef]
- Muthuramalingam, P.; Jeyasri, R.; Bharathi, R.K.A.S.; Suba, V.; Pandian, S.T.K.; Ramesh, M. Global Integrated Omics Expression Analyses of Abiotic Stress Signaling HSF Transcription Factor Genes in Oryza Sativa L.: An in Silico Approach. Genomics 2020, 112, 908–918. [Google Scholar] [CrossRef]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Transcriptional Regulatory Networks in Cellular Responses and Tolerance to Dehydration and Cold Stresses. Annu. Rev. Plant. Biol. 2006, 57, 781–803. [Google Scholar] [CrossRef] [Green Version]
- Tran, L.S.; Nakashima, K.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Plant Gene Networks in Osmotic Stress Response: From Genes to Regulatory Networks. Methods Enzymol. 2007, 428, 109–128. [Google Scholar]
- Udvardi, M.K.; Kakar, K.; Wandrey, M.; Montanari, O.; Murray, J.; Andriankaja, A.; Zhang, J.Y.; Benedito, V.; Hofer, J.M.I.; Chueng, F.; et al. Legume Transcription Factors: Global Regulators of Plant Development and Response to the Environment. Plant Physiol. 2007, 144, 538–549. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Zhang, H.; Kong, L.; Gao, G.; Luo, J. PlantTFDB 3.0: A Portal for the Functional and Evolutionary Study of Plant Transcription Factors. Nucleic Acids Res. 2013, 42, 182–187. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Chen, C.; Chen, Z. Expression Profiles of the Arabidopsis WRKY Gene Superfamily during Plant Defense Response. Plant. Mol. Biol. 2003, 51, 21–37. [Google Scholar] [CrossRef]
- Eulgem, T.; Rushton, P.J.; Robatzek, S.; Somssich, I.E. The WRKY Superfamily of Plant Transcription Factors. Trends Plant. Sci. 2000, 5, 199–206. [Google Scholar] [CrossRef]
- Xie, Z.; Zhang, Z.L.; Zou, X.; Huang, J.; Ruas, P.; Thompson, D.; Shen, Q.J. Annotations and Functional Analyses of the Rice WRKY Gene Superfamily Reveal Positive and Negative Regulators of Abscisic Acid Signaling in Aleurone Cells. Plant Physiol. 2005, 137, 176–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, C.; Palmqvist, S.; Olsson, H.; Borén, M.; Ahlandsberg, S.; Jansson, C. A Novel WRKY Transcription Factor, SUSIBA2, Participates in Sugar Signaling in Barley by Binding to the Sugar-Responsive Elements of the Iso1 Promoter. Plant Cell 2003, 15, 2076–2092. [Google Scholar] [CrossRef] [Green Version]
- Guan, Y.; Meng, X.; Khanna, R.; LaMontagne, E.; Liu, Y.; Zhang, S. Phosphorylation of a WRKY Transcription Factor by MAPKs Is Required for Pollen Development and Function in Arabidopsis. PLoS Genet. 2014, 10, e1004384. [Google Scholar] [CrossRef] [Green Version]
- Ding, Z.J.; Yan, J.Y.; Xu, X.Y.; Yu, D.Q.; Li, G.X.; Zhang, S.Q.; Zheng, S.J. Transcription Factor WRKY 46 Regulates Osmotic Stress Responses and Stomatal Movement Independently in Arabidopsis. Plant J. 2014, 79, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.S.; Kolevski, B.; Smyth, D.R. TRANSPARENT TESTA GLABRA2, a Trichome and Seed Coat Development Gene of Arabidopsis, Encodes a WRKY Transcription Factor. Plant J. 2002, 14, 1359–1375. [Google Scholar] [CrossRef] [Green Version]
- Luo, M.; Dennis, E.S.; Berger, F.; Peacock, W.J.; Chaudhury, A. MINISEED3 (MINI3), a WRKY Family Gene, and HAIKU2 (IKU2), a Leucine-Rich Repeat (LRR) KINASE Gene, Are Regulators of Seed Size in Arabidopsis. Proc. Natl. Acad. Sci. USA 2005, 102, 17531–17536. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Chen, X.; Xie, K.; Xing, Q.; Wu, Y.; Li, J.; Du, C.; Sun, Z.; Guo, Z. Dlf1, a WRKY Transcription Factor, Is Involved in the Control of Flowering Time and Plant Height in Rice. PLoS ONE 2014, 9, e102529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexandrova, K.S.; Conger, B.V. Isolation of Two Somatic Embryogenesis-Related Genes from Orchardgrass (Dactylis glomerata). Plant. Sci. 2002, 162, 301–307. [Google Scholar] [CrossRef]
- Lagacé, M.; Matton, D.P. Characterization of a WRKY Transcription Factor Expressed in Late Torpedo-Stage Embryos of Solanum Chacoense. Planta 2004, 219, 185–189. [Google Scholar] [CrossRef]
- Wang, H.; Avci, U.; Nakashima, J.; Hahn, M.G.; Chen, F.; Dixon, R.A. Mutation of WRKY Transcription Factors Initiates Pith Secondary Wall Formation and Increases Stem Biomass in Dicotyledonous Plants. Proc. Natl. Acad. Sci. USA 2010, 107, 22338–22343. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Hu, R.; Wang, H.; Cao, Y.; He, G.; Fu, C.; Zhou, G. MlWRKY12, a Novel Miscanthus Transcription Factor, Participates in Pith Secondary Cell Wall Formation and Promotes Flowering. Plant. Sci. 2013, 212, 1–9. [Google Scholar] [CrossRef]
- Xu, Y.H.; Wang, J.W.; Wang, S.; Wang, J.Y.; Chen, X.Y. Characterization of GaWRKY1, a Cotton Transcription Factor that Regulates the Sesquiterpene Synthase Gene (+)-δ-cadinene Synthase-A. Plant Physiol. 2004, 135, 507–515. [Google Scholar] [CrossRef] [Green Version]
- Ma, D.; Pu, G.; Lei, C.; Ma, L.; Wang, H.; Guo, Y.; Chen, J.; Du, Z.; Wang, H.; Li, G.; et al. Isolation and Characterization of AaWRKY1, an Artemisia Annua Transcription Factor that Regulates the Amorpha-4, 11-Diene Synthase Gene, a Key Gene of Artemisinin Biosynthesis. Plant. Cell Physiol. 2009, 50, 2146–2161. [Google Scholar] [CrossRef] [Green Version]
- Suttipanta, N.; Pattanaik, S.; Kulshrestha, M.; Patra, B.; Singh, S.K.; Yuan, L. The Transcription Factor Crwrky1 Positively Regulates the Terpenoid Indole Alkaloid Biosynthesis in Catharanthus Roseus. Plant Physiol. 2011, 157, 2081–2093. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.L.; Xie, Z.; Zou, X.; Casaretto, J.; Ho, T.H.D.; Shen, Q.J. A Rice WRKY Gene Encodes a Transcriptional Repressor of the Gibberellin Signaling Pathway in Aleurone Cells. Plant Physiol. 2004, 134, 1500–1513. [Google Scholar] [CrossRef] [Green Version]
- Robatzek, S.; Somssich, I.E. A New Member of the Arabidopsis WRKY Transcription Factor Family, Atwrky6, Is Associated with Both Senescence-and Defence-Related Processes. Plant J. 2001, 28, 123–133. [Google Scholar] [CrossRef]
- Robatzek, S.; Somssich, I.E. Targets of AtWRKY6 Regulation during Plant Senescence and Pathogen Defense. Genes Dev. 2002, 16, 1139–1149. [Google Scholar] [CrossRef] [Green Version]
- Miao, Y.; Laun, T.; Zimmermann, P.; Zentgraf, U. Targets of the WRKY53 Transcription Factor and Its Role during Leaf Senescence in Arabidopsis. Plant. Mol. Biol. 2004, 55, 853–867. [Google Scholar] [CrossRef]
- Muthuramalingam, P.; Jeyasri, R.; Krishnan, S.R.; Pandian, S.T.K.; Sathishkumar, R.; Ramesh, M. Integrating the Bioinformatics and Omics Tools for Systems Analysis of Abiotic Stress Tolerance in Oryza sativa (L.). In Advances in Plant Transgenics: Methods and Applications; Sathishkumar, R., Kumar, S., Hema, J., Baskar, V., Eds.; Springer: Singapore, 2019; pp. 59–77. [Google Scholar]
- Tang, J.; Wang, F.; Wang, Z.; Huang, Z.; Xiong, A.; Hou, X. Characterization and Co-Expression Analysis of WRKY Orthologs Involved in Responses to Multiple Abiotic Stresses in Pak-Choi (Brassica campestris Ssp. Chinensis). BMC Plant Biol. 2013, 13, 188. [Google Scholar] [CrossRef] [Green Version]
- Muthuramalingam, P.; Krishnan, S.R.; Saravanan, K.; Mareeswaran, N.; Ramesh, M. Genome Wide Identification of Effective Transcription Factor Gene Families in Rice Identifies Potential Candidates Involved in Abiotic Stress Signaling. J. Plant. Biochem. Biot. 2018, 27, 300–317. [Google Scholar] [CrossRef]
- Kim, C.Y.; Lee, S.H.; Park, H.C.; Bae, C.G.; Cheong, Y.H.; Choi, Y.J.; Han, C.D.; Lee, S.Y.; Lim, C.O.; Cho, M.J. Identification of Rice Blast Fungal Elicitor-Responsive Genes by Differential Display Analysis. Mol. Plant. Microbe Interact. 2000, 13, 470–474. [Google Scholar] [CrossRef] [Green Version]
- Asai, T.; Tena, G.; Plotnikova, J.; Willmann, M.R.; Chiu, W.L.; Gomez-Gomez, L.; Boller, T.; Ausubel, F.M.; Sheen, J. MAP Kinase Signalling Cascade in Arabidopsis Innate Immunity. Nature 2002, 415, 977. [Google Scholar] [CrossRef] [PubMed]
- Rizhsky, L.; Davletova, S.; Liang, H.; Mittler, R. The Zinc Finger Gene Zat12 Is Required for Cytosolic Ascorbate Peroxidase 1 Expression during Oxidative Stress in Arabidopsis. J. Biol. Chem. 2004, 279, 11736–11743. [Google Scholar] [CrossRef] [Green Version]
- Seki, M.; Narusaka, M.; Ishida, J.; Nanjo, T.; Fujita, M.; Oono, Y.; Kamiya, A.; Nakajima, M.; Enju, A.; Sakurai, T.; et al. Monitoring the Expression Profiles of 7000 Arabidopsis Genes under Drought, Cold and High-Salinity Stresses Using a Full-Length Cdna Microarray. Plant J. 2002, 31, 279–292. [Google Scholar] [CrossRef]
- Hara, K.; Yagi, M.; Kusano, T.; Sano, H. Rapid Systemic Accumulation of Transcripts Encoding a Tobacco WRKY Transcription Factor upon Wounding. Mol. Genet. Genom. 2000, 263, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Duman, J.G. Cloning and Characterization of a Thermal Hysteresis (Antifreeze) Gene with DNA-Binding Activity from Winter Bittersweet Nightshade, Solanum dulcamara. Plant. Mol. Biol. 2002, 48, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Provart, N.J.; Glazebrook, J.; Katagiri, F.; Chang, H.S.; Eulgem, T.; Mauch, F.; Luan, S.; Zou, G.; Whitham, S.A.; et al. Expression Profile Matrix of Arabidopsis Transcription Factor Genes Suggests Their Putative Functions in Response to Environmental Stresses. Plant Cell 2002, 14, 559–574. [Google Scholar] [CrossRef]
- Deslandes, L.; Olivier, J.; Theulières, F.; Hirsch, J.; Feng, D.X.; Bittner-Eddy, P.; Beynon, J.; Marco, Y. Resistance to Ralstonia Solanacearum in Arabidopsis Thaliana Is Conferred by the Recessive RRS1-R Gene, a Member of a Novel Family of Resistance Genes. Proc. Natl. Acad. Sci. USA 2002, 99, 2404–2409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.C.; Lai, Z.; Fan, B.; Chen, Z. Arabidopsis WRKY38 and WRKY62 Transcription Factors Interact with Histone Deacetylase 19 in Basal Defense. Plant Cell 2008, 20, 2357–2371. [Google Scholar] [CrossRef] [Green Version]
- Huh, S.U.; Choi, L.M.; Lee, G.J.; Kim, Y.J.; Paek, K.H. Capsicum Annuum WRKY Transcription Factor d (CaWRKYd) Regulates Hypersensitive Response and Defense Response upon Tobacco Mosaic Virus Infection. Plant. Sci. 2012, 197, 50–58. [Google Scholar] [CrossRef]
- Zheng, Z.; Qamar, S.A.; Chen, Z.; Mengiste, T. Arabidopsis WRKY33 Transcription Factor Is Required for Resistance to Necrotrophic Fungal Pathogens. Plant J. 2006, 48, 592–605. [Google Scholar] [CrossRef] [PubMed]
- Marchive, C.; Mzid, R.; Deluc, L.; Barrieu, F.; Pirrello, J.; Gauthier, A.; Corio-Costet, M.F.; Regad, F.; Cailleteau, B.; Hamdi, S.; et al. Isolation and Characterization of a Vitis Vinifera Transcription Factor, Vvwrky1, and Its Effect on Responses to Fungal Pathogens in Transgenic Tobacco Plants. J. Exp. Bot. 2007, 58, 1999–2010. [Google Scholar] [CrossRef] [Green Version]
- Beyer, K.; Binder, A.; Boller, T.; Collinge, M. Identification of Potato Genes Induced during Colonization by Phytophthora Infestans. Mol. Plant. Pathol. 2001, 2, 125–134. [Google Scholar] [CrossRef]
- Kalde, M.; Barth, M.; Somssich, I.E.; Lippok, B. Members of the Arabidopsis WRKY Group III Transcription Factors Are Part of Different Plant Defense Signaling Pathways. Mol. Plant. Microbe Interact. 2003, 16, 295–305. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Ni, Z.; Yao, Y.; Guo, G.; Sun, Q. Cloning and Expression Profiles of 15 Genes Encoding WRKY Transcription Factor in Wheat (Triticum aestivem L.). Prog. Nat. Sci. 2008, 18, 697–705. [Google Scholar] [CrossRef]
- Qiu, Y.; Jing, S.; Fu, J.; Li, L.; Yu, D. Cloning and Analysis of Expression Profile of 13 WRKY Genes in Rice. Chin. Sci. Bull. 2004, 49, 2159–2168. [Google Scholar] [CrossRef]
- Jiang, Y.; Deyholos, M.K. Comprehensive Transcriptional Profiling of NaCl-stressed Arabidopsis Roots Reveals Novel Classes of Responsive Genes. BMC Plant. Biol. 2006, 6, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Fu, Q.; Chen, L.; Huang, W.; Yu, D. Arabidopsis thaliana WRKY25, WRKY26, and WRKY33 Coordinate Induction of Plant Thermotolerance. Planta 2011, 233, 1237–1252. [Google Scholar] [CrossRef]
- Singh, A.; Singh, P.K.; Sharma, A.K.; Singh, N.K.; Sonah, H.; Deshmukh, R.; Sharma, T.R. Understanding the Role of the WRKY Gene Family under Stress Conditions in Pigeonpea (Cajanus cajan, L.). Plants 2019, 8, 214. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Wang, P.; Nan, Z.; Wang, X. The WRKY Transcription Factor Genes in Lotus Japonicus. Int. J. Genom. 2014, 2014, 420128. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Wang, P.; Lin, J.Y.; Zhao, C.; Bi, Y.; Wang, X. Genome-Wide Identification and Characterization of WRKY Gene Family in Peanut. Front. Plant. Sci. 2016, 7, 534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Chi, Y.; Wang, Z.; Zhou, Y.; Fan, B.; Chen, Z. Functional Analysis of Structurally Related Soybean GmWRKY58 and GmWRKY76 in Plant Growth and Development. J. Exp. Bot. 2016, 67, 4727–4742. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, A.; Nishiyama, M.Y.; Fuentes, B.G.; Souza, G.M.; Janies, D.; Gray, J.; Grotewold, E. GRASSIUS: A Platform for Comparative Regulatory Genomics Across the Grasses. Plant Physiol. 2009, 149, 171–180. [Google Scholar] [CrossRef] [Green Version]
- Sakai, H.; Lee, S.S.; Tanaka, T.; Numa, H.; Kim, J.; Kawahara, Y.; Wakimoto, H.; Yang, C.C.; Iwamoto, M.; Abe, T.; et al. Rice Annotation Project Database (RAP-DB): An Integrative and Interactive Database for Rice Genomics. Plant. Cell Physiol. 2013, 54, e6. [Google Scholar] [CrossRef]
- Sato, Y.; Takehisa, H.; Kamatsuki, K.; Minami, H.; Namiki, N.; Ikawa, H.; Ohyanagi, H. RiceXPro Version 3.0: Expanding the Informatics Resource for Rice Transcriptome. Nucleic Acids Res. 2013, 41, D1206–D1213. [Google Scholar] [CrossRef] [Green Version]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Gene Identification and Analysis Tools on the ExPASy Server. In The Proteomics Protocols Handbook; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An Upgraded Gene Feature Visualization Server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING Database in 2017: Quality-Controlled Gene—Gene Association Networks, Made Broadly Accessible. Nucleic Acids Res. 2016, gkw937. [Google Scholar] [CrossRef]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An Information Aesthetic for Comparative Genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Chen, L.; Zhang, L.; Yu, D. Overexpression of OsWRKY72 Gene Interferes in the Abscisic Acid Signal and Auxin Transport Pathway of Arabidopsis. J. Biosci. 2010, 35, 459–471. [Google Scholar] [CrossRef]
- Zhou, X.; Jiang, Y.; Yu, D. WRKY22 Transcription Factor Mediates Dark-Induced Leaf Senescence in Arabidopsis. Mol. Cells. 2011, 31, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Bakshi, M.; Oelmüller, R. WRKY Transcription Factors: Jack of Many Trades in Plants. Plant. Signal Behav. 2014, 9, e27700. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Lai, Z.; Shi, J.; Xiao, Y.; Chen, Z.; Xu, X. Roles of Arabidopsis WRKY18, WRKY40 and WRKY60 Transcription Factors in Plant Responses to Abscisic Acid and Abiotic Stress. BMC Plant Biol. 2010, 10, 281. [Google Scholar] [CrossRef] [Green Version]
- Zou, C.; Jiang, W.; Yu, D. Male Gametophyte-Specific WRKY34 Transcription Factor Mediates Cold Sensitivity of Mature Pollen in Arabidopsis. J. Exp. Bot. 2010, 61, 3901–3914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.F.; Li, L.Q.; Xu, Q.; Kong, Y.H.; Wang, H.; Wu, W.H. The WRKY6 Transcription Factor Modulates PHOSPHATE1 Expression in Response to Low Pi Stress in Arabidopsis. Plant Cell 2009, 21, 3554–3566. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Deyholos, M.K. Functional Characterization of Arabidopsis NaCl-inducible WRKY25 and WRKY33 Transcription Factors in Abiotic Stresses. Plant. Mol. Biol. 2009, 69, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.; Yan, L.; Liu, Z.Q.; Cao, Z.; Mei, C.; Xin, Q.; Wu, F.Q.; Wang, X.F.; Du, S.Y.; Jiang, T.; et al. The Mg-chelatase H Subunit of Arabidopsis Antagonizes a Group of WRKY Transcription Repressors to Relieve ABA-Responsive Genes of Inhibition. Plant Cell 2010, 22, 1909–1935. [Google Scholar] [CrossRef] [Green Version]
- Berri, S.; Abbruscato, P.; Faivre-Rampant, O.; Brasileiro, A.C.; Fumasoni, I.; Satoh, K.; Kikuchi, S.; Mizzi, L.; Morandini, P.; Pè, M.E.; et al. Characterization of WRKY Co-Regulatory Networks in Rice and Arabidopsis. BMC Plant. Biol. 2009, 9, 120. [Google Scholar] [CrossRef] [Green Version]
- Ramamoorthy, R.; Jiang, S.Y.; Kumar, N.; Venkatesh, P.N.; Ramachandran, S.A. Comprehensive Transcriptional Profiling of the WRKY Gene Family in Rice Under Various Abiotic and Phytohormone Treatments. Plant. Cell Physiol. 2008, 49, 865–879. [Google Scholar] [CrossRef] [PubMed]
- Qiu, D.; Xiao, J.; Xie, W.; Liu, H.; Li, X.; Xiong, L.; Wang, S. Rice Gene Network Inferred from Expression Profiling of Plants Overexpressing Oswrky13, a Positive Regulator of Disease Resistance. Mol. Plant. 2008, 1, 538–551. [Google Scholar]
- Wu, X.; Shiroto, Y.; Kishitani, S.; Ito, Y.; Toriyama, K. Enhanced Heat and Drought Tolerance in Transgenic Rice Seedlings Overexpressing Oswrky11 under the Control of HSP101 Promoter. Plant. Cell Rep. 2009, 28, 21–30. [Google Scholar] [CrossRef]
- Song, X.J.; Matsuoka, M. Bar the Windows: An Optimized Strategy to Survive Drought and Salt Adversities. Genes Dev. 2009, 23, 1709–1713. [Google Scholar] [CrossRef] [Green Version]
- Rushton, P.J.; Somssich, I.E.; Ringler, P.; Shen, Q.J. WRKY Transcription Factors. Trends Plant. Sci. 2010, 15, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Muthuramalingam, P.; Krishnan, S.R.; Pothiraj, R.; Ramesh, M. Global Transcriptome Analysis of Combined Abiotic Stress Signaling Genes Unravels Key Players in Oryza sativa L.: An in Silico Approach. Front. Plant. Sci. 2017, 8, 759. [Google Scholar] [CrossRef] [Green Version]
- Ning, P.; Liu, C.; Kang, J.; Lv, J. Genome-Wide Analysis of WRKY Transcription Factors in Wheat (Triticum aestivum L.) and Differential Expression under Water Deficit Condition. PeerJ. 2017, 5, e3232. [Google Scholar] [CrossRef] [Green Version]
- Imran, Q.M.; Hussain, A.; Mun, B.G.; Lee, S.U.; Asaf, S.; Ali, M.A.; Lee, I.J.; Yun, B.W. Transcriptome Wide Identification and Characterization of NO-Responsive WRKY Transcription Factors in Arabidopsis thaliana L. Environ. Exp. Bot. 2018, 148, 128–143. [Google Scholar] [CrossRef]
- Fathi, A.; Tari, D.B. Effect of Drought Stress and Its Mechanism in Plants. Int. J. Life Sci. 2016, 10, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Cha, J.; Choi, C.; Choi, N.; Ji, H.S.; Park, S.R.; Lee, S.; Hwang, D.J. Rice WRKY11 Plays a Role in Pathogen Defense and Drought Tolerance. Rice 2018, 11, 1–12. [Google Scholar] [CrossRef]
- Dai, X.; Wang, Y.; Zhang, W.H. OsWRKY74, a WRKY Transcription Factor, Modulates Tolerance to Phosphate Starvation in Rice. J. Exp. Bot. 2016, 67, 947––960. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Liang, G.; Yu, D. Activated Expression of WRKY57 Confers Drought Tolerance in Arabidopsis. Mol. Plant. 2012, 5, 1375–1388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, X.; Chen, Z.; Liu, Y.; Zhang, H.; Zhang, M.; Liu, Q.; Hong, X.; Zhu, J.K.; Gong, Z. ABO3, a WRKY Transcription Factor, Mediates Plant Responses to Abscisic Acid and Drought Tolerance in Arabidopsis. Plant J. 2010, 63, 417–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.; Mishra, V.K.; Kumari, S.; Chand, R.; Varadwaj, P.K. Deciphering Genome-Wide WRKY Gene Family of Triticum aestivum L. and Their Functional Role in Response to Abiotic Stress. Genes Genom. 2019, 41, 79–94. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. NO | Gene Name | RAP ID | Start | End | Chr. No | UniProt ID |
|---|---|---|---|---|---|---|
| 1 | OsWRKY1 | Os01g0246700 | 4356383 | 4340849 | 1 | Q0JQ43 |
| 2 | OsWRKY6 | Os03g0798500 | 22731943 | 22733240 | 3 | Q94D50 |
| 3 | OsWRKY10 | Os01g0186000 | 26688416 | 26687377 | 1 | Q0JKQ9 |
| 4 | OsWRKY11 | Os01g0626400 | 25009453 | 25012236 | 1 | Q9FE35 |
| 5 | OsWRKY12 | Os01g0624700 | 29723065 | 29720923 | 1 | Q5JLU2 |
| 6 | OsWRKY14 | Os01g0730700 | 30604295 | 30608077 | 1 | Q942D2 |
| 7 | OsWRKY23 | Os01g0734000 | 36194611 | 36193840 | 1 | Q6IEM5 |
| 8 | OsWRKY24 | Os01g0826400 | 42946753 | 42948750 | 1 | Q5JM93 |
| 9 | OsWRKY28 | Os06g0649000 | 26283914 | 26280253 | 6 | Q0DZ26 |
| 10 | OsWRKY29 | Os07g0111400 | 28726783 | 28730933 | 7 | Q6Z8E9 |
| 11 | OsWRKY32 | Os02g0770500 | 12394669 | 12396898 | 2 | Q10LT9 |
| 12 | OsWRKY37 | Os04g0597300 | 31326926 | 31323190 | 4 | Q9AUV7 |
| 13 | OsWRKY45 | Os05g0322900 | 30132491 | 30136547 | 5 | Q0JAI8 |
| 14 | OsWRKY49 | Os05g0565900 | 4999626 | 4998210 | 5 | Q65WY5 |
| 15 | OsWRKY55 | Os03g0321700 | 23530499 | 23529423 | 3 | Q6IEN3 |
| 16 | OsWRKY66 | Os02g0698800 | 2958991 | 2963006 | 2 | Q5VMX9 |
| 17 | OsWRKY71 | Os07g0583700 | 23659625 | 23654076 | 7 | Q84ZS7 |
| 18 | OsWRKY73 | Os07g0680400 | 28832398 | 28828793 | 7 | Q7XHX5 |
| 19 | OsWRKY74 | Os08g0198000 | 5669406 | 5663578 | 8 | Q0J7F5 |
| 20 | OsWRKY79 | Os08g0386200 | 18220041 | 18222408 | 8 | Q6ZA22 |
| 21 | OsWRKY82 | Os09g0334500 | 10128825 | 10131136 | 9 | Q6ERI5 |
| 22 | OsWRKY83 | Os09g0417600 | 14977713 | 14975932 | 9 | Q6EPZ2 |
| 23 | OsWRKY85 | Os09g0481700 | 18501264 | 18496949 | 9 | Q0J0V4 |
| 24 | OsWRKY92 | Os11g0117500 | 789030 | 787542 | 11 | Q2RBB8 |
| 25 | OsWRKY94 | Os11g0490900 | 17352085 | 17355820 | 11 | Q2R432 |
| 26 | OsWRKY101 | Os12g0116700 | 825793 | 824302 | 12 | Q2QYJ6 |
| S. No | Gene Name | RAP ID | Nt L | aa L | M. Wt | pI | AI | II | GRAVY | SL |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | OsWRKY1 | Os01g0246700 | 1275 | 425 | 45,967.1 | 10.0694 | 68 | 40.54 | −0.383 | Ct |
| 2 | OsWRKY6 | Os03g0798500 | 741 | 247 | 26,500.7 | 7.6528 | 68.21 | 55.29 | −0.549 | Cyto |
| 3 | OsWRKY10 | Os01g0186000 | 825 | 275 | 29,056.4 | 7.4157 | 60.33 | 43.9 | −0.4 | N |
| 4 | OsWRKY11 | Os01g0626400 | 1140 | 380 | 39,856 | 7.0634 | 49.45 | 63.38 | −0.731 | N |
| 5 | OsWRKY12 | Os01g0624700 | 738 | 246 | 25,832.6 | 8.2902 | 54.37 | 43.51 | −0.424 | Ct |
| 6 | OsWRKY14 | Os01g0730700 | 765 | 255 | 27,796.2 | 6.9031 | 66.14 | 47.02 | −0.656 | N |
| 7 | OsWRKY23 | Os01g0734000 | 588 | 196 | 21,158.8 | 9.4863 | 59.18 | 63.57 | −0.624 | N |
| 8 | OsWRKY24 | Os01g0826400 | 1233 | 411 | 45,109.9 | 4.6849 | 60 | 59.53 | −0.694 | N |
| 9 | OsWRKY28 | Os06g0649000 | 570 | 190 | 20,495.6 | 8.492 | 64.07 | 50.13 | −0.375 | Cyto |
| 10 | OsWRKY29 | Os07g0111400 | 1521 | 507 | 51,766.5 | 6.1521 | 57.67 | 48.26 | −0.354 | N |
| 11 | OsWRKY32 | Os02g0770500 | 1071 | 357 | 36,,383.7 | 4.9404 | 56.83 | 55.42 | −0.431 | N |
| 12 | OsWRKY37 | Os04g0597300 | 945 | 315 | 32733.4 | 5.869 | 60.83 | 69.16 | −0.416 | N |
| 13 | OsWRKY45 | Os05g0322900 | 1545 | 515 | 53,042.3 | 5.3217 | 57.43 | 47.7 | −0.415 | PM |
| 14 | OsWRKY49 | Os05g0565900 | 585 | 195 | 21,400.7 | 6.269 | 52.01 | 52.32 | −0.579 | Ct |
| 15 | OsWRKY55 | Os03g0321700 | 996 | 332 | 34,901.2 | 4.3811 | 57.73 | 65.5 | −0.507 | N |
| 16 | OsWRKY66 | Os02g0698800 | 1176 | 392 | 41,403.5 | 6.1909 | 51.33 | 52.47 | −0.646 | N |
| 17 | OsWRKY71 | Os07g0583700 | 1857 | 619 | 66,163.6 | 6.3317 | 58.75 | 51.59 | −0.683 | N |
| 18 | OsWRKY73 | Os07g0680400 | 1002 | 334 | 34,837.5 | 6.5945 | 54.26 | 70.97 | −0.471 | N |
| 19 | OsWRKY74 | Os08g0198000 | 2586 | 862 | 97,370.3 | 6.4311 | 80.14 | 41.56 | −0.41 | N |
| 20 | OsWRKY79 | Os08g0386200 | 960 | 320 | 33,550.3 | 6.6611 | 67.4 | 41.48 | −0.316 | N |
| 21 | OsWRKY82 | Os09g0334500 | 1086 | 362 | 37,944.1 | 6.4041 | 64.21 | 53.99 | −0.428 | N |
| 22 | OsWRKY83 | Os09g0417600 | 984 | 328 | 34,780.2 | 8.0535 | 68.69 | 50.66 | −0.496 | N |
| 23 | OsWRKY85 | Os09g0481700 | 1902 | 634 | 68,280.1 | 6.0112 | 51.66 | 64.68 | −0.712 | N |
| 24 | OsWRKY92 | Os11g0117500 | 963 | 321 | 35,915.9 | 6.417 | 72.25 | 58.59 | −0.672 | N |
| 25 | OsWRKY94 | Os11g0490900 | 729 | 243 | 25,857.2 | 9.3347 | 46.45 | 55.93 | −0.778 | N |
| 26 | OsWRKY101 | Os12g0116700 | 966 | 322 | 35,749.5 | 6.3705 | 69.31 | 60.38 | −0.723 | N |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeyasri, R.; Muthuramalingam, P.; Satish, L.; Adarshan, S.; Lakshmi, M.A.; Pandian, S.K.; Chen, J.-T.; Ahmar, S.; Wang, X.; Mora-Poblete, F.; et al. The Role of OsWRKY Genes in Rice When Faced with Single and Multiple Abiotic Stresses. Agronomy 2021, 11, 1301. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11071301
Jeyasri R, Muthuramalingam P, Satish L, Adarshan S, Lakshmi MA, Pandian SK, Chen J-T, Ahmar S, Wang X, Mora-Poblete F, et al. The Role of OsWRKY Genes in Rice When Faced with Single and Multiple Abiotic Stresses. Agronomy. 2021; 11(7):1301. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11071301
Chicago/Turabian StyleJeyasri, Rajendran, Pandiyan Muthuramalingam, Lakkakula Satish, Sivakumar Adarshan, Muthukannan Aishwarya Lakshmi, Shunmugiah Karutha Pandian, Jen-Tsung Chen, Sunny Ahmar, Xiukang Wang, Freddy Mora-Poblete, and et al. 2021. "The Role of OsWRKY Genes in Rice When Faced with Single and Multiple Abiotic Stresses" Agronomy 11, no. 7: 1301. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11071301