
Monitoring of Emerging Water Stress Situations by Thermal and Vegetation Indices in Different Almond Cultivars

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Irrigation Treatments
2.3. Plant Measurements and Indices Calculations
2.4. Experimental Design and Statistical Analysis
3. Results and Discussion
3.1. Weather Conditions
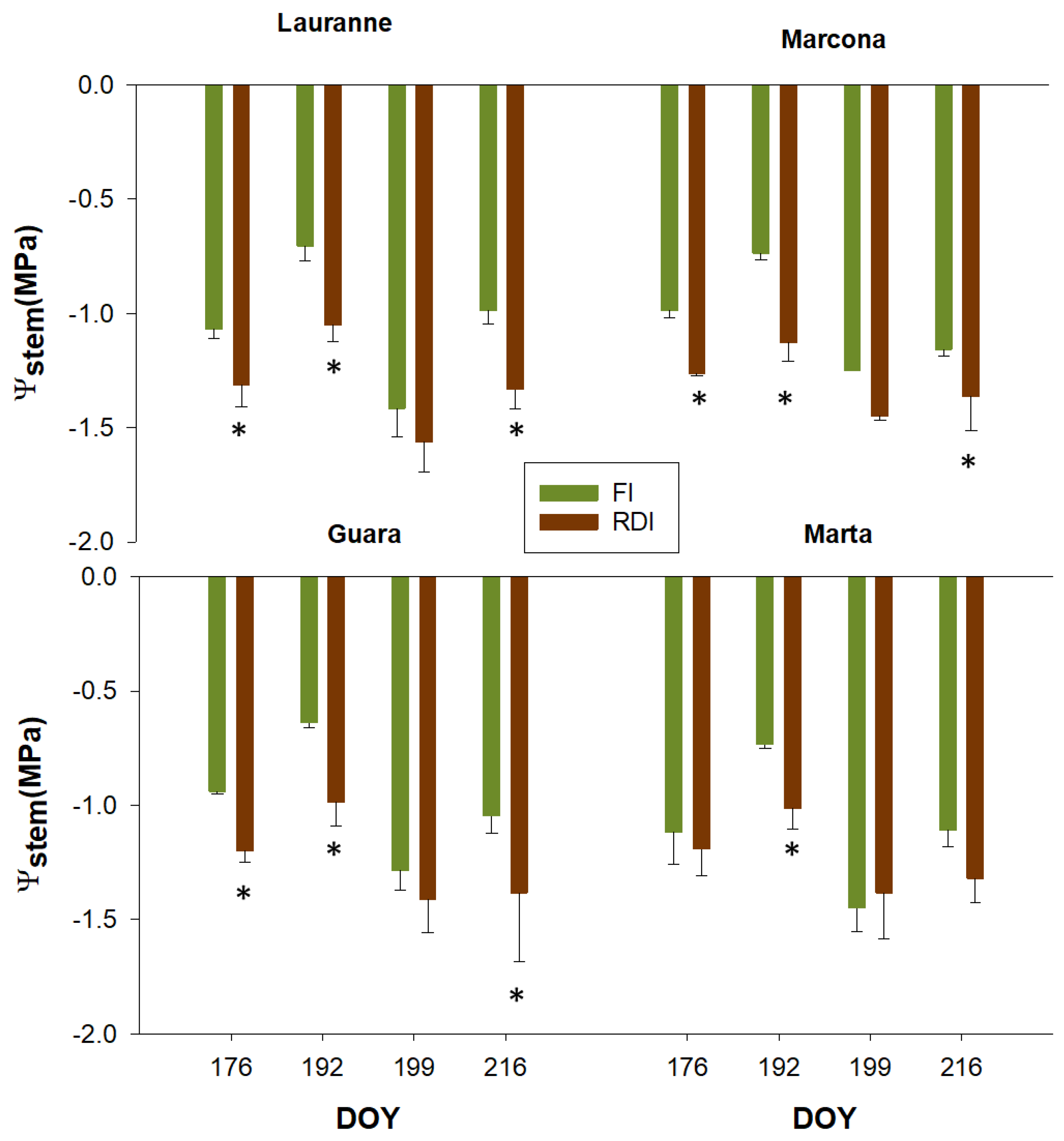
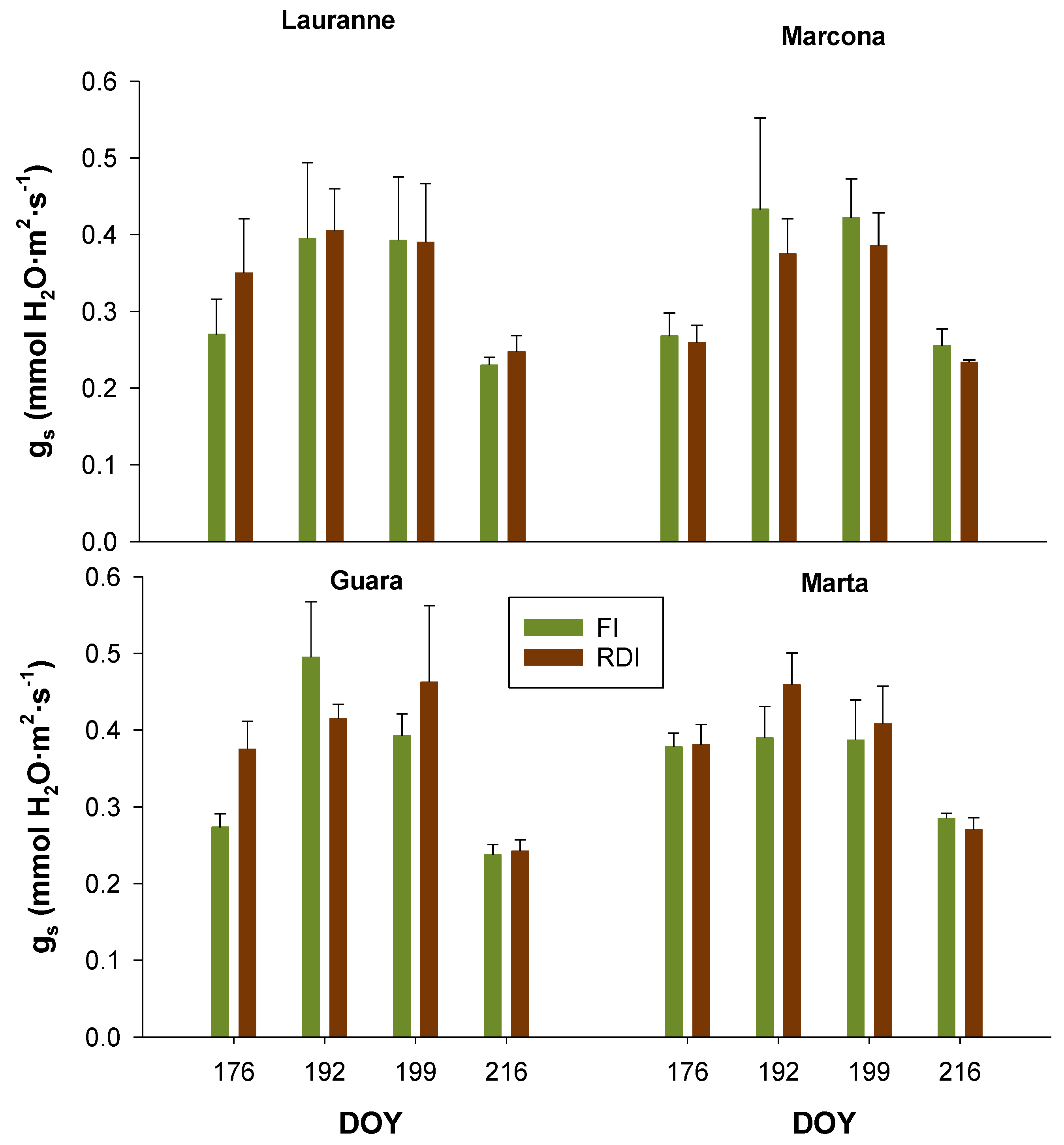
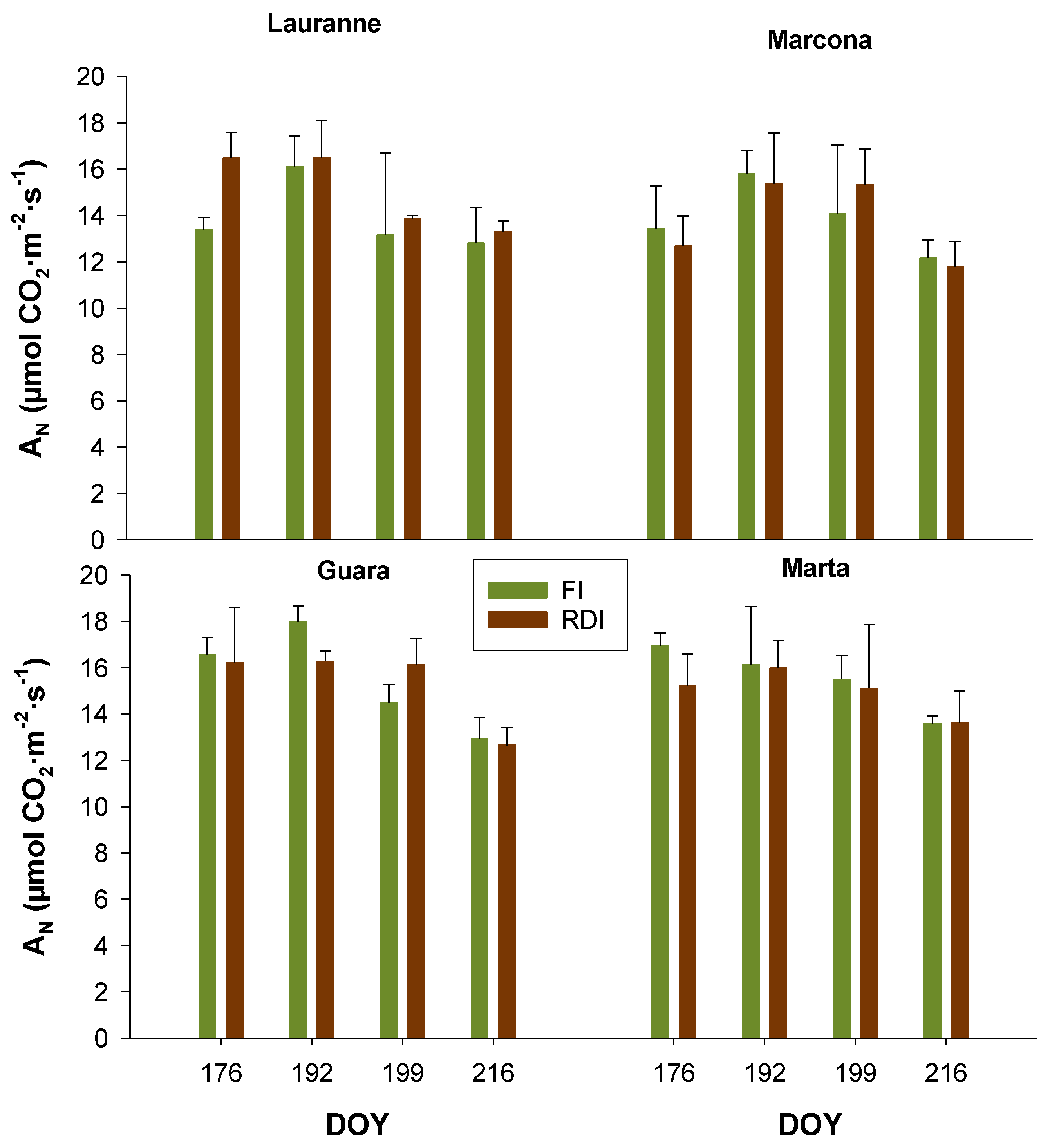
3.2. Charactetization of Cultivars Water Status Through Leaf-Level Physiological Measurements
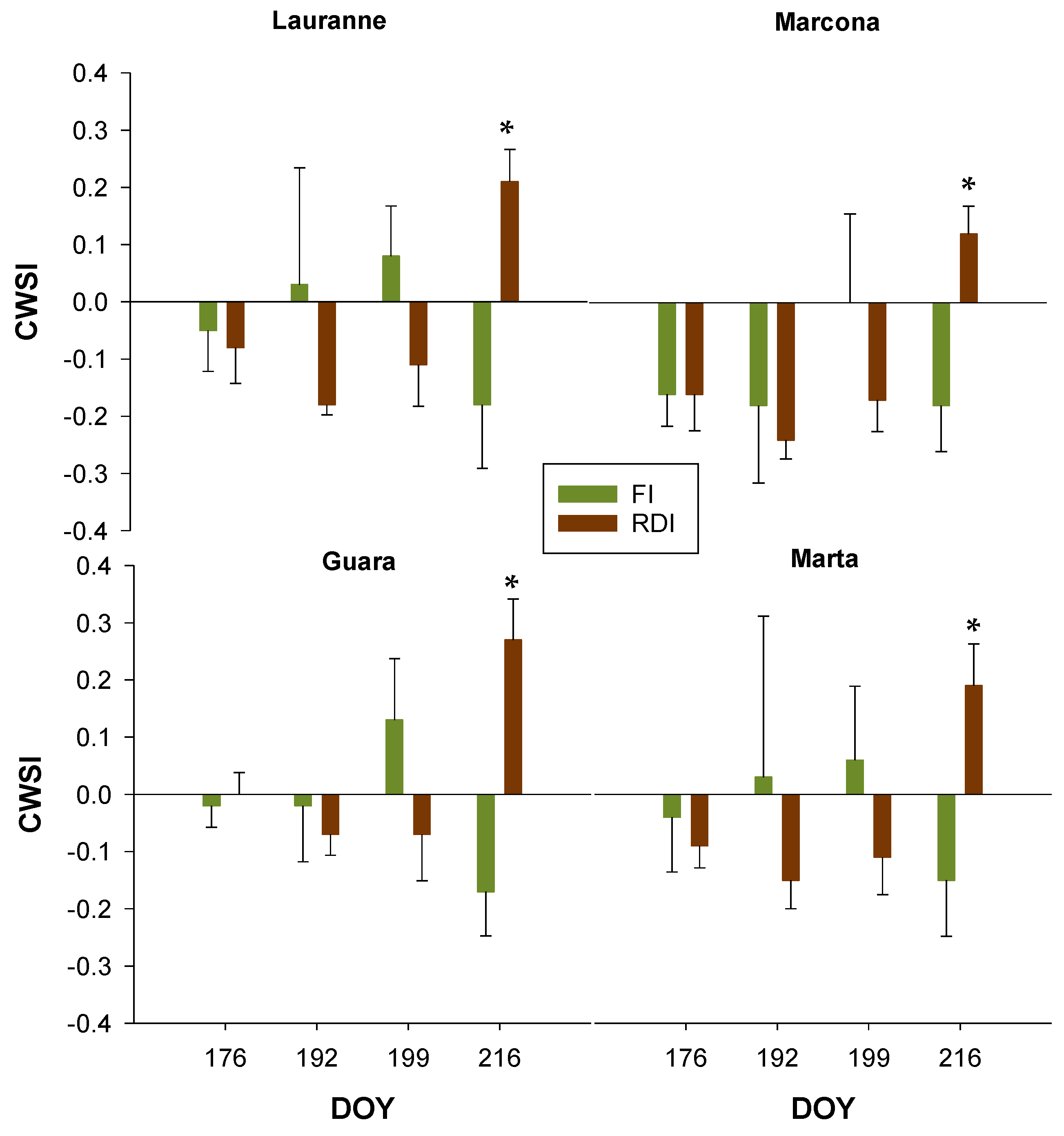
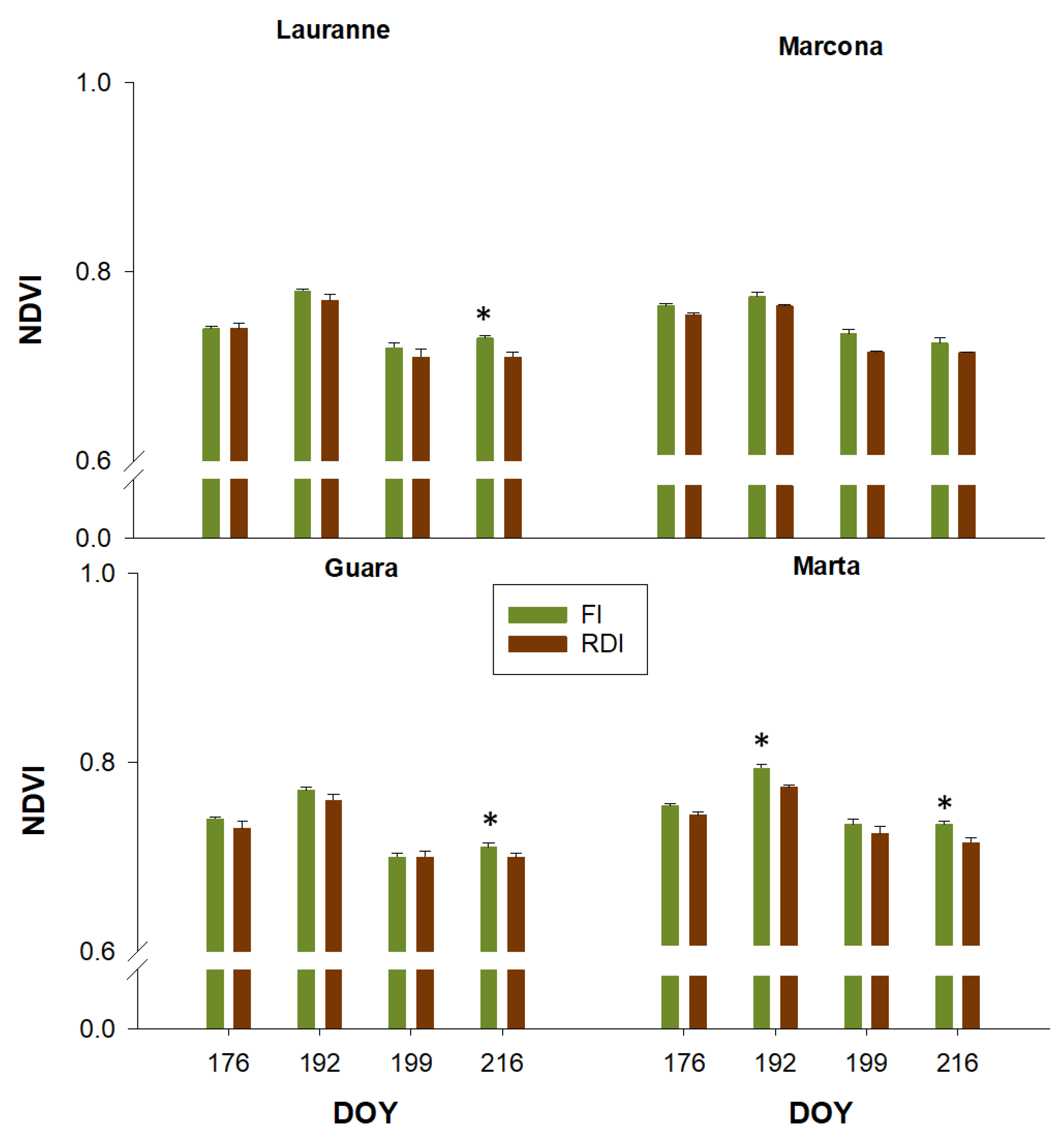
3.3. Charactetization of Cultivars Water Stress Levels Through Remote Sensing Measurements
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- ESYRCE. Encuesta Sobre Superficies y Rendimientos de Cultivos. Ministerio de Agricultura, Alimentación y medio Ambiente, 1st ed.; ESYRCE: Madrid, Spain, 2020. [Google Scholar]
- Goldhamer, D.A.; Fereres, E. Establishing an Almond Water Production Function for California Using Long-Term Yield Response to Variable Irrigation. Irrig. Sci. 2017, 35, 169–179. [Google Scholar] [CrossRef]
- López-López, M.; Espadador, M.; Testi, L.; Lorite, I.J.; Orgaz, F.; Fereres, E. Water Use of Irrigated Almond Trees When Subjected to Water Deficits. Agric. Water Manag. 2018, 195, 84–93. [Google Scholar] [CrossRef]
- Fereres, E.; Soriano, M.A. Deficit Irrigation for Reducing Agricultural Water Use. J. Exp. Bot. 2007, 58, 147–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cano-Lamadrid, M.; Hernández, F.; Corell, M.; Burló, F.; Legua, P.; Moriana, A.; Carbonell-Barrachina, Á.A. Antioxidant Capacity, Fatty Acids Profile, and Descriptive Sensory Analysis of Table Olives as Affected by Deficit Irrigation. J. Sci. Food Agric. 2017, 97, 444–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbonell-Barrachina, Á.A.; Memmi, H.; Noguera-Artiaga, L.; Gijón-López, M.D.C.; Ciapa, R.; Pérez-López, D. Quality Attributes of Pistachio Nuts as Affected by Rootstock and Deficit Irrigation. J. Sci. Food Agric. 2015, 95, 2866–2873. [Google Scholar] [CrossRef] [PubMed]
- Lipan, L.; Moriana, A.; López Lluch, D.B.; Cano-Lamadrid, M.; Sendra, E.; Hernández, F.; Vázquez-Araújo, L.; Corell, M.; Carbonell-Barrachina, Á.A. Nutrition Quality Parameters of Almonds as Affected by Deficit Irrigation Strategies. Molecules 2014, 24, 2646. [Google Scholar] [CrossRef] [Green Version]
- Fernández, J.E.; Perez-Martin, A.; Torres-Ruiz, J.M.; Cuevas, M.V.; Rodriguez-Dominguez, C.M.; Elsayed-Farag, S.; Morales-Sillero, A.; García, J.M.; Hernandez-Santana, V.; Diaz-Espejo, A. A Regulated Deficit Irrigation Strategy for Hedgerow Olive Orchards with High Plant Density. Plant Soil 2013, 372, 279–295. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Bello, M.A.; Ballester, C.; Castel, J.R.; Intrigliolo, D.S. Development and Validation of an Automatic Thermal Imaging Process for As-Sessing Plant Water Status. Agric. Water Manag. 2011, 98, 1497–1504. [Google Scholar] [CrossRef] [Green Version]
- Egea, G.; Diaz-Espejo, A.; Fernández, J.E. Soil Moisture Dynamics in a Hedgerow Olive Orchard under Well-Watered and Deficit Irrigation Regimes: Assessment, Prediction and Scenario Analysis. Agric. Water Manag. 2016, 164, 197–211. [Google Scholar] [CrossRef]
- Park, S.; Ryu, D.; Fuentes, S.; Chung, H.; Hernández-Montes, E.; O’Connell, M. Adaptive Estimation of Crop Water Stress in Nectarine and Peach Orchards Using High-Resolution Imagery from an Unmanned Aerial Vehicle (UAV). Remote Sens. 2017, 9, 828. [Google Scholar] [CrossRef] [Green Version]
- Ojeda-Bustamante, W.; González-Sánchez, A.; Mauricio-Pérez, A.; Flores-Velázquez, J. Aplicaciones de los Vehículos AÉREOS No Tripulados en la Ingeniería Hidroagrícola. Tecnol. Y Cienc. Del Agua. 2017, 8, 157–166. [Google Scholar] [CrossRef]
- García-Tejero, I.F.; Gutiérrez Gordillo, S.; Souza, L.; Cuadros-Tavira, S.; Durán Zuazo, V.H. Fostering Sustainable Water Use in Almond (Prunus dulcis Mill.) Orchards in a Semiarid Mediterranean Environment. Arch. Agron. Soil Sci. 2019, 65, 164–181. [Google Scholar] [CrossRef]
- Jackson, R.D.; Idso, S.B.; Reginato, R.J.; Pinter, P.J. Canopy Temperature as a Crop Water Stress Indicator. Water Resour. Res. 1981, 17, 1133–1138. [Google Scholar] [CrossRef]
- Idso, S.B.; Jackson, R.D.; Pinter, P.J.J.; Reginato, R.J.; Hatfield, J.L. Normalizing the Stress Degree-Day Parameter for Environmental Variability. Agric. Meteorol. 1981, 24, 45–55. [Google Scholar] [CrossRef]
- Egea, G.; Padilla-Díaz, C.M.; Martinez-Guanter, J.; Fernández, J.E.; Pérez-Ruiz, M. Assessing a Crop Water Stress Index Derived from Aerial Thermal Imaging and Infrared Thermometry in Super-High Density Olive Orchards. Agric. Water Manag. 2017, 187, 210–221. [Google Scholar] [CrossRef] [Green Version]
- Bellvert, J.; Marsal, J.; Girona, J.; González-Dugo, V.; Fereres, E.; Ustin, S.L.; Zarco-Tejada, P.J. Airborne Thermal Imagery to Detect the Seasonal Evolution of Crop Water Status in Peach, Nectarine and Saturn Peach Orchards. Remote Sens. 2016, 8, 39. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Dugo, V.; Zarco-Tejada, P.J.; Fereres, E. Applicability and Limitations of Using the Crop Water Stress Index as an Indicator of Water Deficits in Citrus Orchards. Agric. For. Meteorol. 2014, 198, 94–104. [Google Scholar] [CrossRef]
- Xue, J.; Su, B. Significant Remote Sensing Vegetation Indexes: A review of Developments and Applications. J. Sens. 2017. [Google Scholar] [CrossRef] [Green Version]
- Conesa, M.R.; Conejero, W.; Vera, J.; Ramírez-Cuesta, J.M.; Ruiz-Sánchez, M.C. Terrestrial and Remote Indexes to Assess Moderate Deficit Irrigation in Early-Maturing Nectarine Trees. Agronomy 2019, 9, 630. [Google Scholar] [CrossRef] [Green Version]
- Balota, M.; Oakes, J. UAV Remote Sensing for Phenotyping Drought tolerance in Peanuts. In Autonomous Air and Ground Sensing Systems for Agricultural Optimization and Phenotyping II; Alex Thomasson, J., McKee, M., Moorhead, R.J., Eds.; Society of Photo-Optical Instrumentation Engineers (SPIE) Conference Series; International Society for Optics and Photonics: Washington, DC, USA, 2017; p. 10218. [Google Scholar]
- Zhao, T.; Stark, B.; Chen, Y.; Ray, A.L.; Doll, D. Challenges in Water Stress Quantification Using Small Unmanned Aerial System (sUAS): Lessons from A Growing Season of Almond. J. Intell. Robot. Syst. 2017, 88, 721–735. [Google Scholar] [CrossRef]
- Espinoza, C.Z.; Khot, L.R.; Sankaran, S.; Jacoby, P.W. High Resolution Multispectral and Thermal Remote Sensing-Based Water Stress Assessment in Subsurface Irrigated Grapevines. Remote Sens. 2017, 9, 961. [Google Scholar] [CrossRef] [Green Version]
- Glenn, D.M.; Tabb, A. Evaluation of Five Methods to Measure Normalized Difference Vegetation Index (NDVI) in Apple and Citrus. Int. J. Fruit Sci. 2019, 19, 191–210. [Google Scholar] [CrossRef]
- Zhuang, S.; Wang, P.; Jiang, B.; Li, M.; Gong, Z. Early detection of water stress in maize based on digital images. Comput. Electron. Agric. 2017, 140, 461–468. [Google Scholar] [CrossRef]
- López-López, M.; Espadafor, M.; Testi, L.; Lorite, I.J.; Orgaz, F.; Fereres, E. Yield Response of Almond Trees to Transpiration Deficits. Irrig. Sci. 2018, 36, 111–120. [Google Scholar] [CrossRef]
- Girona, J.; Mata, M.; Marsal, J. Regulated Deficit Irrigation during the Kernel-Filling Period and Optimal Irrigation Rates in Almond. Agric. Water Manag. 2005, 75, 152–167. [Google Scholar] [CrossRef]
- Gonzalez-Dugo, V.; Lopez-Lopez, M.; Espadafor, M.; Orgaz, F.; Testi, L.; Zarco-Tejada, P.; Lorite, I.J.; Fereres, E. Transpiration from Canopy Temperature: Implications for the Assessment of Crop Yield in Almond Orchards. Eur. J. Agron. 2019, 105, 78–85. [Google Scholar] [CrossRef]
- RIA. Red de Información Agroclimática de Andalucía. Instituto Andaluz de Investigación y Formación Agraria y Pesquera. Consejería de Agricultura, Ganadería, Pesca y Desarrollo Sostenible. Junta de Andalucía. Available online: https://ifapa.junta-419andalucia.es/agriculturaypesca/ifapa/ria/servlet/FrontController?action=Static&u420rl=coordenadas.jsp&c_provincia=41&c_estacion=19 (accessed on 1 February 2021).
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration—Guidelines for Computing Crop Water Requirements-Irrigation and Drainage Paper 56; Food and Agriculture Organization (FAO): Rome, Italy, 1998. [Google Scholar]
- García-Tejero, I.F.; Hernandez, A.; Rodriguez, V.M.; Ponce, J.R.; Ramos, V.; Muriel, J.L.; Durán Zuazo, V.H. Estimating Almond Crop Coefficients and Physiological Response to Water Stress in Semiarid Environments (SW Spain). J. Agric. Sci. Technol. 2015, 17, 1255–1266. [Google Scholar]
- Steduto, P.; Hsiao, T.C.; Fereres, E.; Raes, D. Crop Yield Response to Water. FAO Irrigation and Drainage Paper No. 66; Food and Agriculture Organization of the United Nations: Rome, Italy, 2012. [Google Scholar]
- Bellvert, J.; Zarco-Tejada, P.J.; Girona, J.; Fereres, E. Mapping Crop Water Stress Index in a “Pinot-noir” Vineyard: Comparing Ground Measurements with Thermal Remote Sensing Imagery from an Unmanned Aerial Vehicle. Precis. Agric. 2014, 15, 361–376. [Google Scholar] [CrossRef]
- Testi, L.; Goldhamer, D.A.; Iniesta, F.; Salinas, M. Crop water stress index is a sensitive water stress indicator in pistachio trees. Irrig. Sci. 2008, 26, 395–405. [Google Scholar] [CrossRef] [Green Version]
- Goldhamer, D.A.; Fereres, E. Irrigation Scheduling Protocols Using Continuously Recorded Trunk Diameter Measurements. Irrig. Sci. 2001, 20, 115–1250. [Google Scholar] [CrossRef]
- Barzegar, K.; Yadollahi, A.; Imani, A.; Ahmadi, N. Influences of Severe Water Stress on Photosynthesis, Water Use Efficiency and Proline Content of Almond Cultivars. J. Appl. Hortic. 2012, 14, 33–39. [Google Scholar] [CrossRef]
- Larsen, F.E.; Higgins, S.S.; Al Wir, A. Diurnal Water Relations of Apple, Apricot, Grape, Olive and Peach in an Arid Environment (Jordan). Sci. Hortic. 1989, 39, 211–222. [Google Scholar] [CrossRef]
- Egea, G.; Dodd, I.C.; González-Real, M.M.; Domingo, R.; Baille, A. Partial Rootzone Drying Improves Almond Tree Leaf-Level Water Use Efficiency and Afternoon Water Status Compared with Regulated Deficit Irrigation. Funct. Plant Biol. 2011, 38, 372–385. [Google Scholar] [CrossRef] [PubMed]
- Wartinger, A.; Heilmeier, H.; Hartung, W.; Schulze, E.D. Daily and Seasonal Courses of Leaf Conductance and Abscisic Acid in the Xylem Sap of Almond Trees [Prunus dulcis (Miller) DA Webb] under Desert Conditions. New Phytol. 1990, 116, 581–587. [Google Scholar] [CrossRef]
- Steinberg, S.L.; Miller, J.C.; McFarland, M.J. Dry Matter Partitioning and Vegetative Growth of Young Peach Trees under Water Stress. Funct. Plant Biol. 1990, 17, 23–36. [Google Scholar] [CrossRef]
- Fernández, J.E. Understanding Olive Adaptation to Abiotic Stresses as a TOOL to increase Crop Performance. Environ. Exp. Bot. 2014, 103, 158–179. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.; Meinzer, F.C. Metrics and Proxies for Stringency of Regulation of Plant Water Status (Iso/Anisohydry): A Global Data Set Reveals Coordination and Trade-Offs among Water Transport Traits. Tree Physiol. 2018, 39, 122–134. [Google Scholar] [CrossRef] [PubMed]
- Tardieu, F.; Simonneau, T. Variability among Species of Stomatal Control under Fluctuating Soil Water Status and Evaporative Demand: Modelling Isohydric and Anisohydric Behaviours. J. Exp. Bot. 1998, 49, 419–432. [Google Scholar] [CrossRef] [Green Version]
- Hamlyn, G.J. Irrigation Scheduling-Comparison of Soil, Plant and Atmosphere Monitoring Approaches. Act. Hortic. 2008, 792, 16. [Google Scholar]
- Hamlyn, G.J.; Tardieu, F. Modeling Water Relations of horticultural CROPS: A Review. Sci. Hortic. 1998, 74, 21–46. [Google Scholar]
- Limpus, S. Isohydric and Anisohydric Characterization of Vegetable Crops. The Classification of Vegetables by Their Physiological Responses to Water Stress. Project Report, 1st ed.; The Department of Primary Industries and Fisheries: Brisbane, QLD, Australia, 2009. [Google Scholar]
- Espadafor, M.; Orgaz, F.; Testi, L.; Lorite, I.J.; González-Dugo, V.; Fereres, E. Responses of Transpiration and Transpiration Efficiency of Almond Trees to Moderate Water Deficits. Sci. Hortic. 2017, 225, 6–14. [Google Scholar] [CrossRef]
- Egea, G.; Verhoef, A.; Vidale, P.L. Towards an Improved and More Flexible Representation of Water Stress in Coupled Photosynthesis—Stomatal Conductance Models. Agric For. Meteorol. 2011, 151, 1370–1384. [Google Scholar] [CrossRef]
- Suarez, L.; González-Dugo, V.; Camino, C.; Hornero, A.; Zarco-Tejada, P.-J. Physical Model Inversion of the Green Spectral Region to Track Assimilation Rate in Almond Trees with an Airborne Nano-Hyperspectral Imager. Remote Sens. Environ. 2021, 252, 112147. [Google Scholar] [CrossRef]
- Caturegli, L.; Corniglia, M.; Gaetani, M.; Grossi, N.; Magni, S.; Migliazzi, M.; Angelini, L.; Mazzoncini, M.; Silvestri, N.; Fontanelli, M.; et al. Unmanned Aerial Vehicle to Estimate Nitrogen Status of Turfgrasses. PLoS ONE 2016, 11, e0158268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Julie, C.A.; Zinnert, C.; Polo, M.-J.; Young, D.-R. NDVI as An Indicator for Changes in Water Availability to Woody Vegetation. Ecol. Indic. 2012, 23, 290–300. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Coefficients | March | April | May | June | July | August | September | October |
|---|---|---|---|---|---|---|---|---|
| Kc | 0.4 | 0.6 | 0.9 | 1.1 | 1.2 | 1.1 | 0.8 | 0.7 |
| Kr | 0.4 | 0.6 | 0.7 | 0.7 | 0.7 | 0.7 | 0.7 | 0.4 |
| Month | Air Temperature | Relative Humidity | Radiation | Rainfall | ETo | ETc |
|---|---|---|---|---|---|---|
| March | 14.0 | 71.7 | 15.9 | 66.2 | 80.6 | 13.0 |
| April | 15.9 | 75.5 | 16.1 | 71.6 | 84.9 | 34.4 |
| May | 21.5 | 61.5 | 24.5 | 45.0 | 153.1 | 130.8 |
| June | 23.4 | 52.2 | 27.9 | 0.0 | 167.0 | 148.7 |
| July | 28.9 | 46.2 | 27.3 | 2.0 | 190.5 | 182.6 |
| August | 25.0 | 32.9 | 25.3 | 0.8 | 167.1 | 143.7 |
| September | 20.1 | 58.6 | 19.8 | 19.8 | 118.5 | 80.2 |
| October | 17.3 | 65.8 | 15.1 | 37.4 | 70.2 | 24.1 |
| DOY | Hour | RH | T | VPD |
|---|---|---|---|---|
| 176 | 11:00 | 27.6 | 34.47 | 3.94 |
| 11:30 | 25.03 | 34.82 | 4.16 | |
| 12:00 | 24.43 | 34.84 | 4.20 | |
| 12:30 | 24.52 | 34.98 | 4.23 | |
| 192 | 11:00 | 36.25 | 33.24 | 3.24 |
| 11:30 | 32.73 | 34.34 | 3.64 | |
| 12:00 | 30.96 | 35.1 | 3.89 | |
| 12:30 | 25.93 | 36.52 | 4.51 | |
| 199 | 11:00 | 18.72 | 38.06 | 5.39 |
| 11:30 | 18.45 | 38.22 | 5.45 | |
| 12:00 | 17.02 | 38.62 | 5.67 | |
| 12:30 | 16.52 | 39.08 | 5.84 | |
| 216 | 11:00 | 27.1 | 33.58 | 3.78 |
| 11:30 | 25.55 | 34.35 | 4.03 | |
| 12:00 | 23.51 | 34.9 | 4.26 | |
| 12:30 | 23.71 | 35.39 | 4.37 |
| DOY | VPD | Ψstem_REF | Ψstem_RDI | Water Stress Signal |
|---|---|---|---|---|
| 176 | 4.13 | −0.91 | −1.24 | 1.37 |
| 192 | 3.82 | −0.87 | −1.05 | 1.20 |
| 199 | 5.59 | −1.08 | −1.45 | 1.34 |
| 216 | 4.11 | −0.90 | −1.35 | 1.49 |
| Mild Water Stress | Ψstem | gs | AN | CWSI | NDVI |
|---|---|---|---|---|---|
| Cultivar | 0.675 | 0.026 * | 0.598 | 0.063 | 0.087 |
| Treatment | <0.001 * | 0.609 | 0.507 | 0.005 * | 0.086 |
| Cultivar × Treatment | 0.403 | 0.581 | 0.585 | 0.890 | 0.938 |
| Moderate Water Stress | |||||
| Cultivar | 0.765 | 0.008 * | 0.489 | 0.805 | 0.180 |
| Treatment | <0.001 * | 0.737 | 0.957 | <0.001 * | <0.001 * |
| Cultivar × Treatment | 0.809 | 0.548 | 0.940 | 0.824 | 0.991 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutiérrez-Gordillo, S.; de la Gala González-Santiago, J.; Trigo-Córdoba, E.; Rubio-Casal, A.E.; García-Tejero, I.F.; Egea, G. Monitoring of Emerging Water Stress Situations by Thermal and Vegetation Indices in Different Almond Cultivars. Agronomy 2021, 11, 1419. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11071419
Gutiérrez-Gordillo S, de la Gala González-Santiago J, Trigo-Córdoba E, Rubio-Casal AE, García-Tejero IF, Egea G. Monitoring of Emerging Water Stress Situations by Thermal and Vegetation Indices in Different Almond Cultivars. Agronomy. 2021; 11(7):1419. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11071419
Chicago/Turabian StyleGutiérrez-Gordillo, Saray, Javier de la Gala González-Santiago, Emiliano Trigo-Córdoba, Alfredo Emilio Rubio-Casal, Iván Francisco García-Tejero, and Gregorio Egea. 2021. "Monitoring of Emerging Water Stress Situations by Thermal and Vegetation Indices in Different Almond Cultivars" Agronomy 11, no. 7: 1419. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11071419