
Carbohydrate and Amino Acid Dynamics during Grain Growth in Four Temperate Cereals under Well-Watered and Water-Limited Regimes

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growing Conditions
2.2. Physiological Traits
2.3. Carbohydrate Determination in the Stems, Flag Leaves and Grain
2.4. Free Amino Acid (AA) Determination
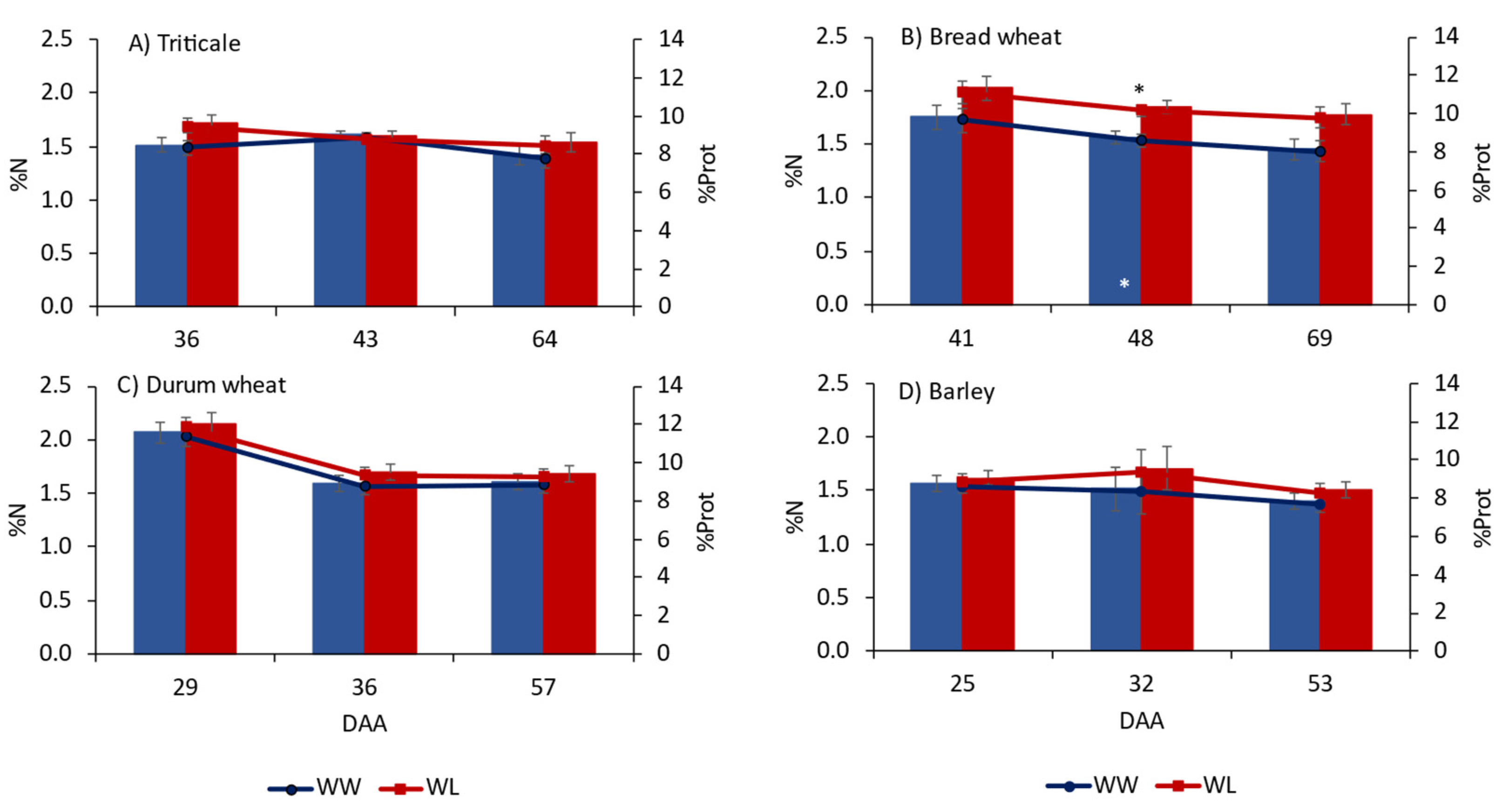
2.5. Nitrogen and Carbon Determination in Grain
2.6. Productivity Traits
2.7. Statistical Analysis
3. Results
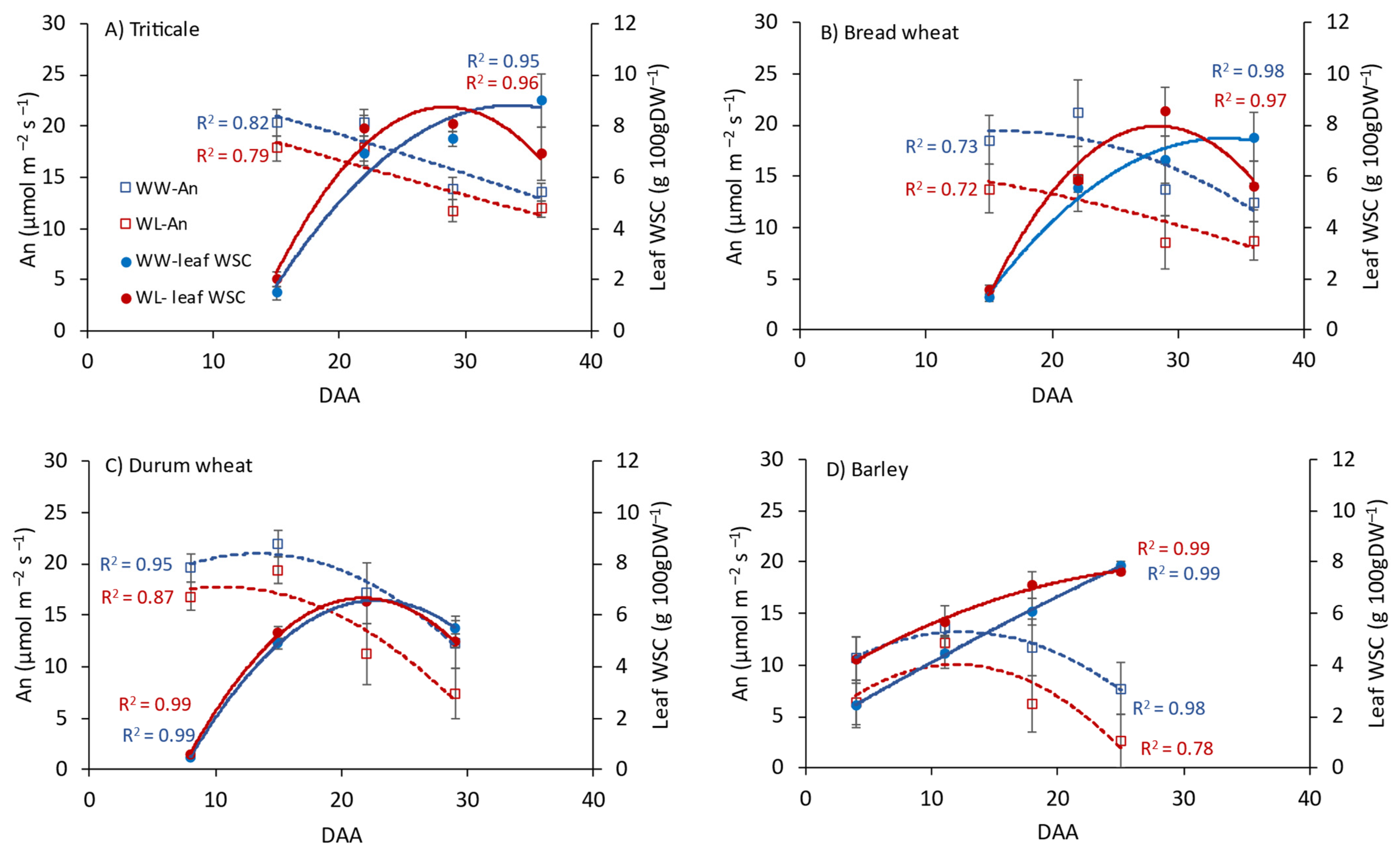
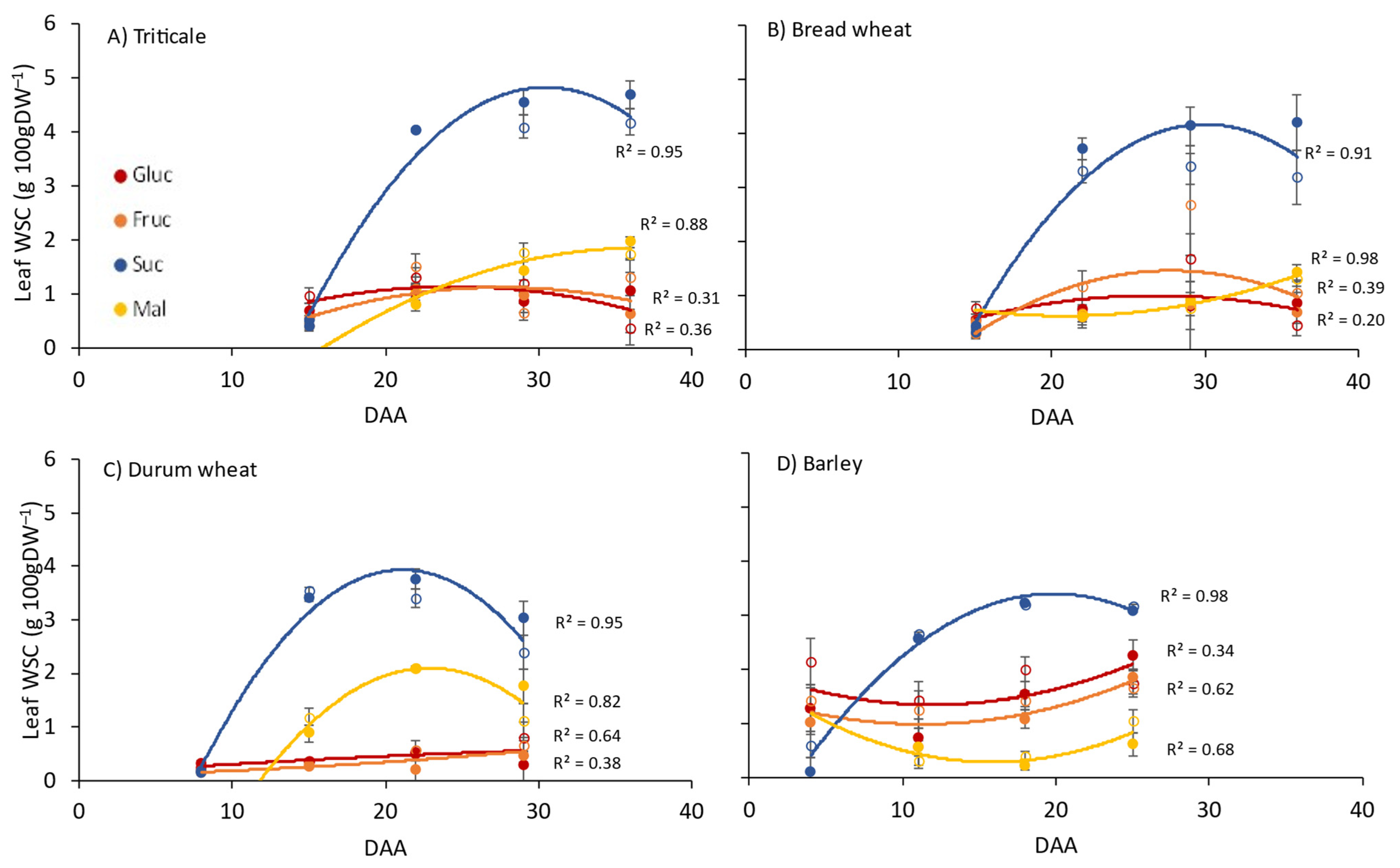
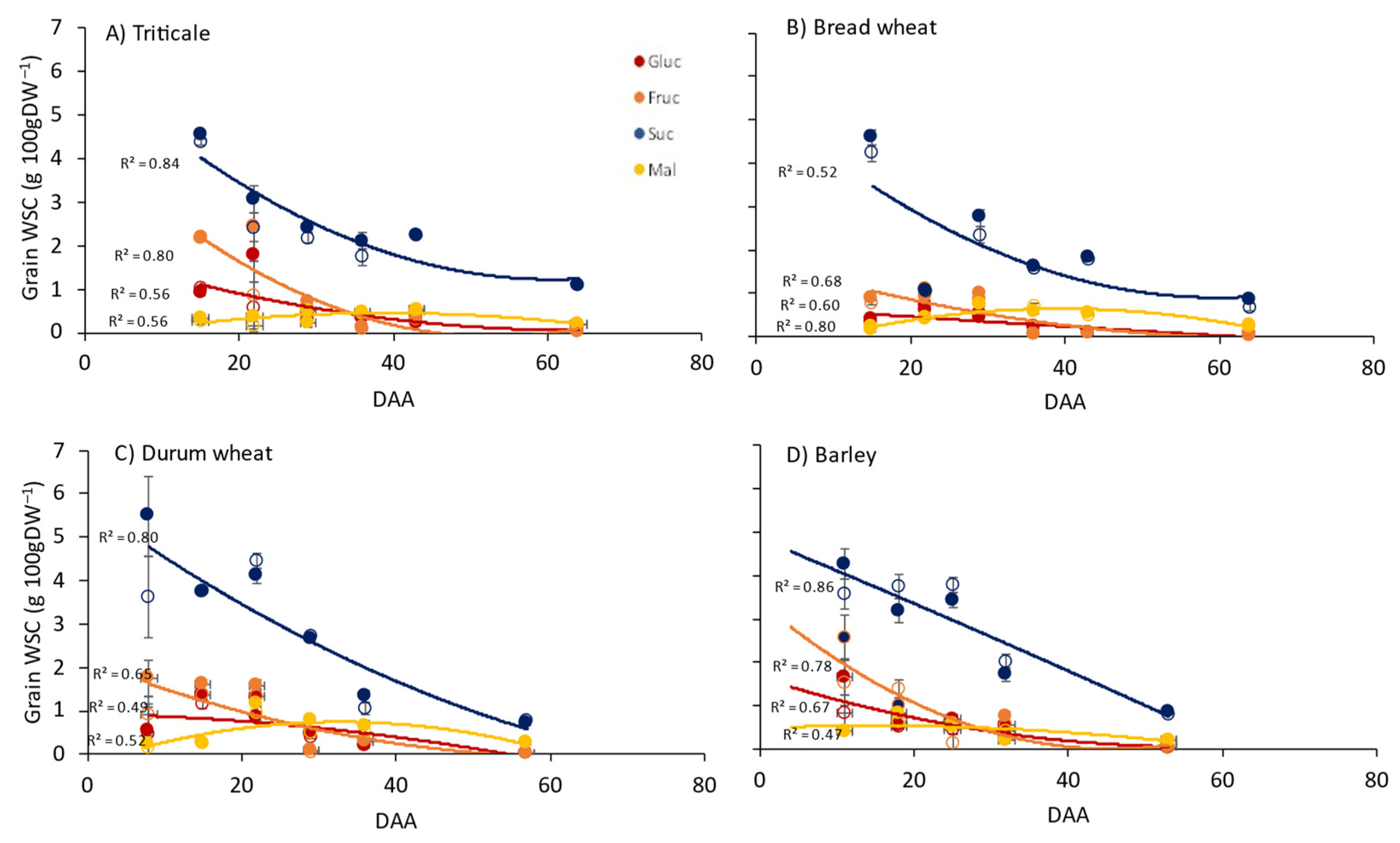
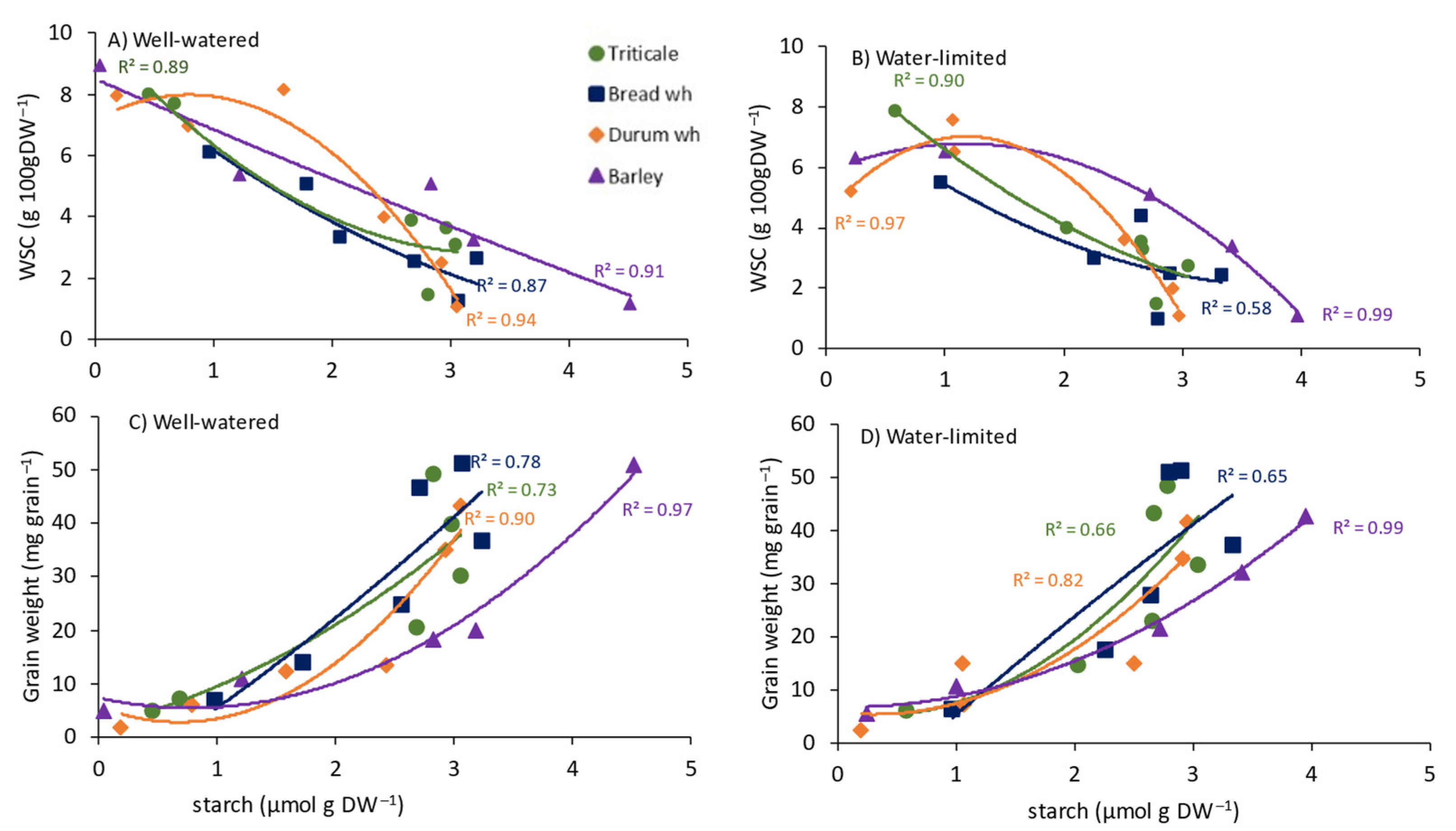
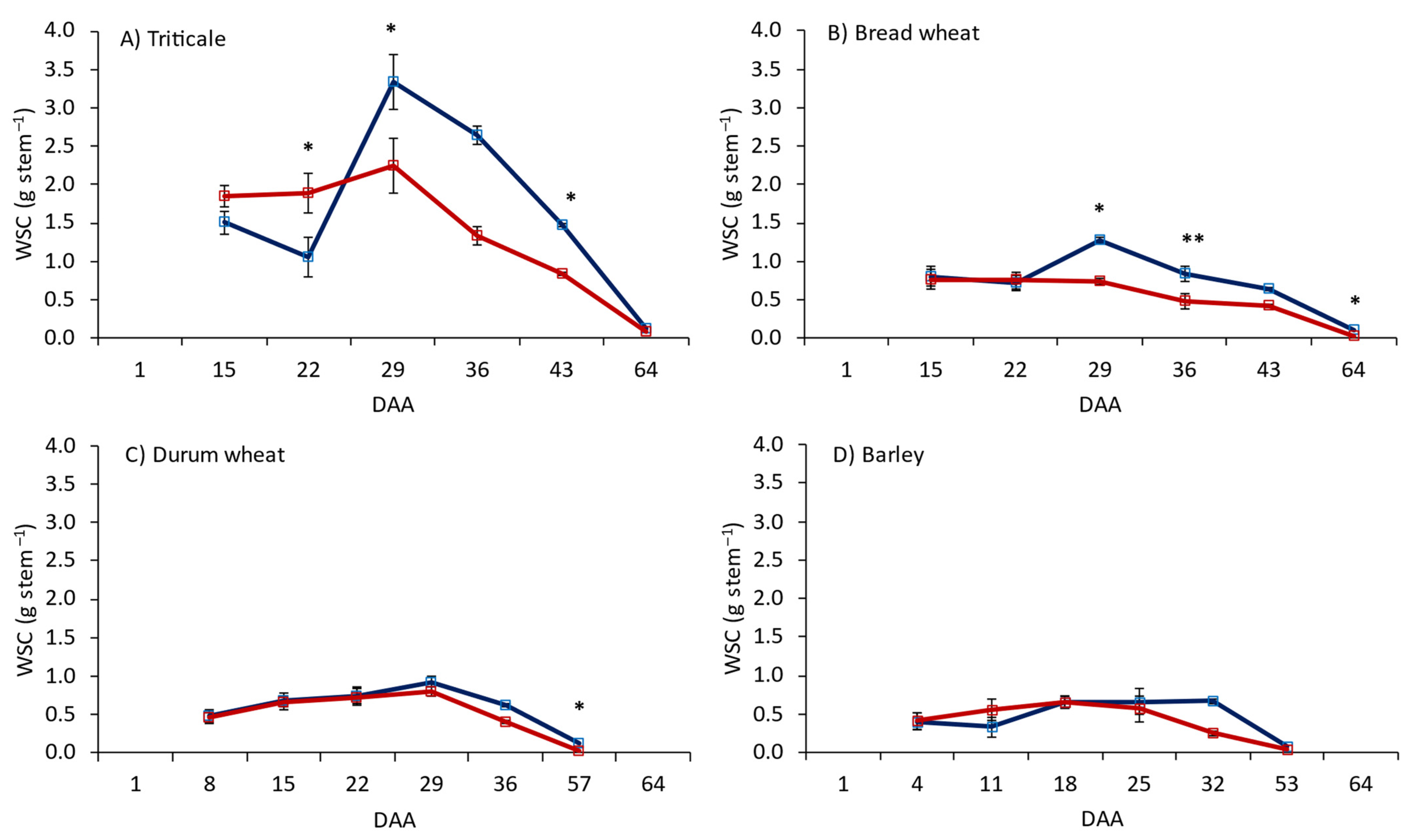
3.1. Leaf Photosynthesis and WSCs in Flag Leaves, Stems and Grain
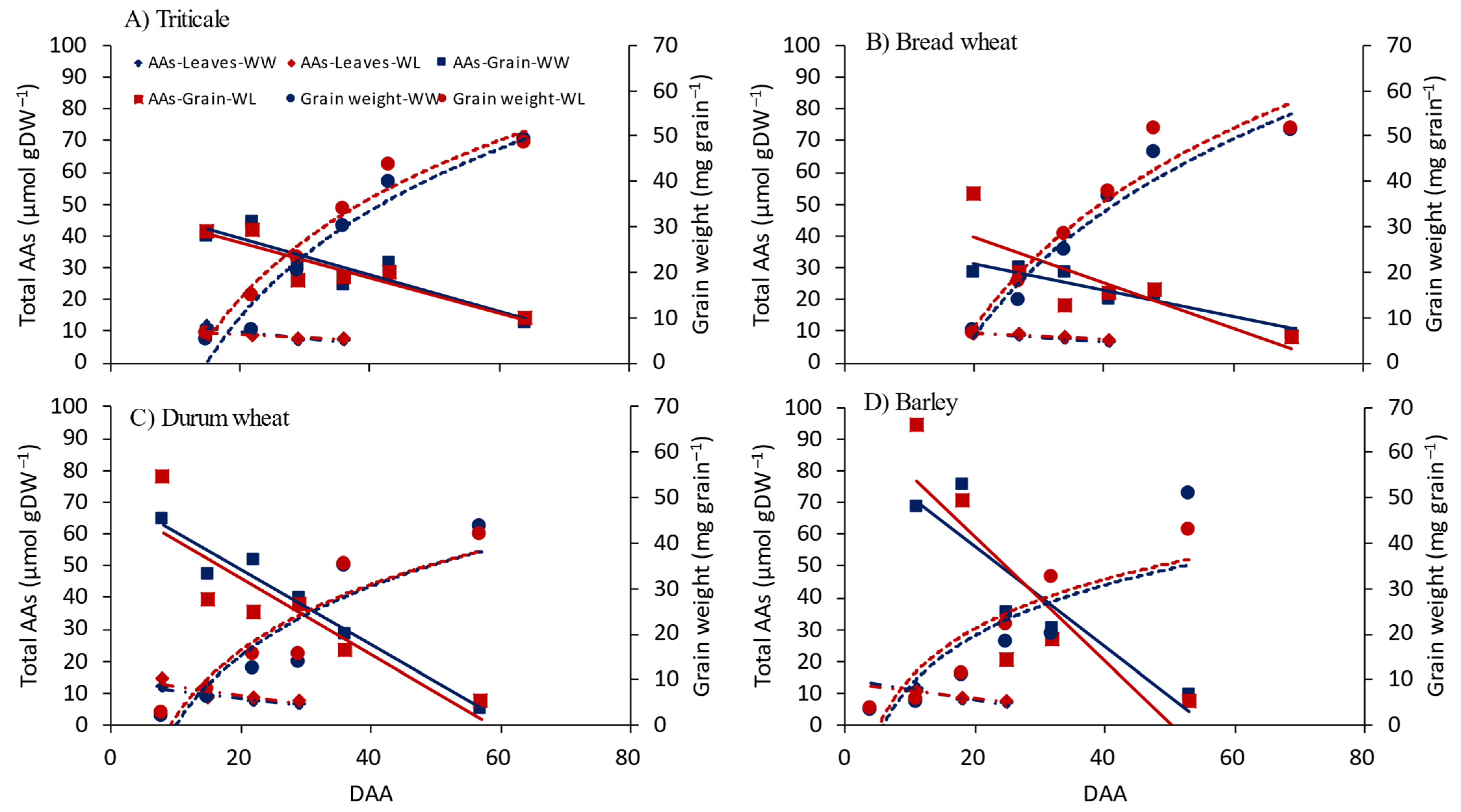
3.2. Amino Acids in Flag Leaves and Grain
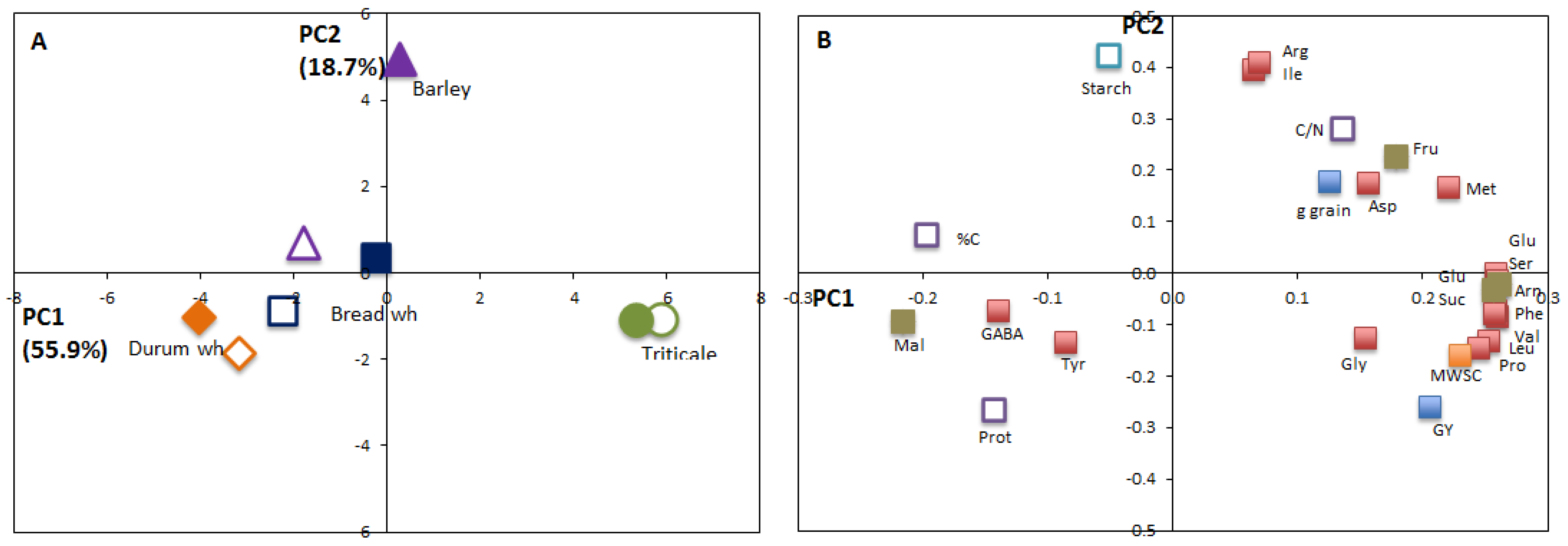
3.3. PCA Analysis
4. Discussion
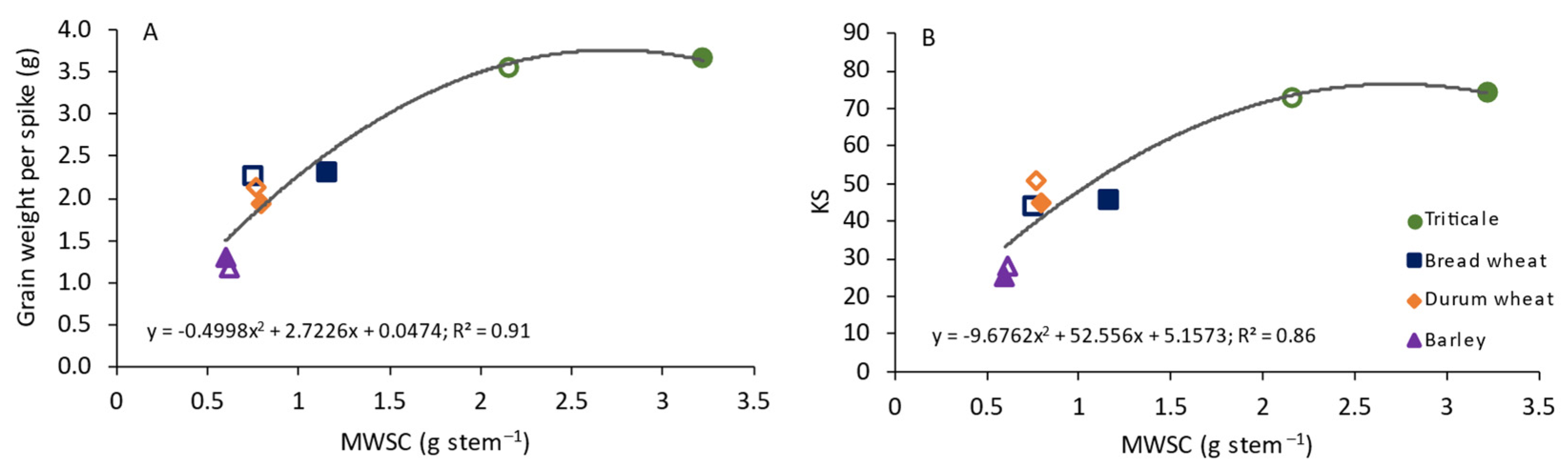
4.1. Carbohydrate Dynamics between Source and Sink Organs
4.2. Amino Acids in Flag Leaves and Grain
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Dolferus, R.; Powell, N.; Ji, X.; Ravash, R.; Edlington, J.; Oliver, S.; van Dongen, J.; shiran, B. Chapter 8: The physiology of reproductive-stage abiotic stress tolerance in cereals. In Molecular Stress Physiology of Plants; Rout, G., Das, A., Eds.; Springer: Chennai, India, 2013; pp. 193–218. [Google Scholar]
- Sanchez-Bragado, R.; Elazab, A.; Zhou, B.; Serret, M.D.; Bort, J.; Nieto-Taladriz, M.T.; Araus, J.L. Contribution of the ear and the flag leaf to grain filling in durum wheat inferred from the carbon isotope signature: Genotypic and growing conditions effects. J. Integr. Plant Biol. 2014, 56, 444–454. [Google Scholar] [CrossRef]
- Méndez-Espinoza, A.M.; Romero-Bravo, S.; Estrada, F.; Garriga, M.; Lobos, G.A.; Castillo, D.; Matus, I.; Aranjuelo, I.; del Pozo, A. Exploring agronomic and physiological traits associated with the differences in productivity between triticale and bread wheat in Mediterranean environments. Front. Plant Sci. 2019, 10, 404. [Google Scholar] [CrossRef] [PubMed]
- del Pozo, A.; Yañez, A.; Matus, I.A.; Tapia, G.; Castillo, D.; Sanchez-Jardón, L.; Araus, J.L. Physiological traits associated with wheat yield potential and performance under water-stress in a Mediterranean environment. Front. Plant Sci. 2016, 7, 987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanca, A.M.; Romagosa, I.; Takeda, K.; Lundborg, T.; Terzi, V.; Cattivelli, L. Chapter 9: Diversity in abiotic stress tolerances. In Diversity in Barley (Hordeum Vulgare); Von Bothmer, R., Van Hintum, T., Knüpffer, H., Sato, K., Eds.; Elsevier: Amsterdam, The Netherlands, 2003; Volume 7, pp. 179–199. [Google Scholar]
- Ehdaie, B.; Alloush, G.A.; Madore, M.A.; Waines, J.G. Genotypic variation for stem reserves and mobilization in wheat: I. Postanthesis changes in internode dry matter. Crop. Sci. 2006, 46, 735–746. [Google Scholar] [CrossRef]
- Schnyder, H. The role of carbohydrate storage and redistribution in the source-sink relations of wheat and barley during grain filling-a review. New Phytol. 1993, 123, 233–245. [Google Scholar] [CrossRef]
- Evans, L.; Wardlaw, I.; Fischer, R. Wheat; Cambridge University Press: London, UK, 1975; pp. 101–149. [Google Scholar]
- Maydup, M.L.; Antonietta, M.; Guiamet, J.J.; Tambussi, E.A. The contribution of green parts of the ear to grain filling in old and modern cultivars of bread wheat (Triticum aestivum L.): Evidence for genetic gains over the past century. Field Crops Res. 2012, 134, 208–215. [Google Scholar] [CrossRef]
- Tambussi, E.A.; Maydup, M.L.; Carrión, C.A.; Guiamet, J.J.; Araus, J.L. Ear photosynthesis in C3 cereals and its contribution to grain yield: Methodologies, controversies, and perspectives. J. Exp. Bot. 2021, 72, 3956–3970. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Bragado, R.; Vicente, R.; Molero, G.; Serret, M.D.; Maydup, M.L.; Araus, J.L. New avenues for increasing yield and stability in C3 cereals: Exploring ear photosynthesis. Curr. Opin. Plant Biol. 2020, 56, 223–234. [Google Scholar] [CrossRef]
- Teulat, B.; Borries, C.; This, D. New QTLs identified for plant water status, water-soluble carbohydrate and osmotic adjustment in a barley population grown in a growth-chamber under two water regimes. Theo. Appl. Genet. 2001, 103, 161–170. [Google Scholar] [CrossRef]
- Cramer, G.; Ergül, A.; Grimplet, J.; Tillett, R.; Tattersall, E.R.; Bohlman, M.; Vincent, D.; Sonderegger, J.; Evans, J.; Osborne, C.; et al. Water and salinity stress in grapevines: Early and late changes in transcript and metabolite profiles. Funct. Integr. Genomics 2007, 7, 111–134. [Google Scholar] [CrossRef]
- Bogeat-Triboulot, M.-B.; Brosché, M.; Renaut, J.; Jouve, L.; Le Thiec, D.; Fayyaz, P.; Vinocur, B.; Witters, E.; Laukens, K.; Teichmann, T. Gradual soil water depletion results in reversible changes of gene expression, protein profiles, ecophysiology, and growth performance in Populus euphratica, a poplar growing in arid regions. Plant Physiol. 2007, 143, 876–892. [Google Scholar] [CrossRef] [Green Version]
- Méndez, A.M.; Castillo, D.; Del Pozo, A.; Matus, I.; Morcuende, R. Differences in stem soluble carbohydrate contents among recombinant chromosome substitution lines (RCSLs) of barley under drought in a Mediterranean-type environment. Agron. Res. 2011, 9, 433. [Google Scholar]
- del Pozo, A.; Castillo, D.; Inostroza, L.; Matus, I.; Méndez, A.M.; Morcuende, R. Physiological and yield responses of recombinant chromosome substitution lines of barley to terminal drought in a Mediterranean-type environment. Ann. Appl. Biol. 2012, 160, 157–167. [Google Scholar] [CrossRef]
- de Roover, J.; Vandenbranden, K.; van Laere, A.; van den Ende, W. Drought induces fructan synthesis and 1-SST (sucrose: Sucrose fructosyltransferase) in roots and leaves of chicory seedlings (Cichorium intybus L.). Planta 2000, 210, 808–814. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, J. Grain filling of cereals under soil drying. New Phytol. 2006, 169, 223–236. [Google Scholar] [CrossRef] [PubMed]
- Yáñez, A.; Tapia, G.; Guerra, F.; del Pozo, A. Stem carbohydrate dynamics and expression of genes involved in fructan accumulation and remobilization during grain growth in wheat (Triticum aestivum L.) genotypes with contrasting tolerance to water stress. PLoS ONE 2017, 12, e0177667. [Google Scholar] [CrossRef] [Green Version]
- Maydup, M.L.; Antonietta, M.; Guiamet, J.J.; Graciano, C.; López, J.R.; Tambussi, E.A. The contribution of ear photosynthesis to grain filling in bread wheat (Triticum aestivum L.). Field Crops Res. 2010, 119, 48–58. [Google Scholar] [CrossRef]
- Rolland, F.; Baena-Gonzalez, E.; Sheen, J. Sugar sensing and signaling in plants: Conserved and novel mechanisms. Annu. Rev. Plant Biol. 2006, 57, 675–709. [Google Scholar] [CrossRef] [Green Version]
- Ramel, F.; Sulmon, C.; Gouesbet, G.; Couée, I. Natural variation reveals relationships between pre-stress carbohydrate nutritional status and subsequent responses to xenobiotic and oxidative stress in Arabidopsis thaliana. Ann. Bot. 2009, 104, 1323–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majumder, A.L.; Sengupta, S.; Goswami, L. Osmolyte regulation in abiotic stress. In Abiotic Stress Adaptation in Plants; Pareek, A., Sopory, S.K., Bohnert, H.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 349–370. [Google Scholar]
- Díaz, P.; Borsani, O.; Monza, J. Acumulación de prolina en plantas en respuesta al estrés osmótico. Agrociencia 1999, 3, 1–10. [Google Scholar]
- Lea, P.J.; Sodek, L.; Parry, M.A.J.; Shewry, P.R.; Halford, N.G. Asparagine in plants. Ann. Appl. Biol. 2007, 150, 1–26. [Google Scholar] [CrossRef]
- Kirma, M.; Araújo, W.L.; Fernie, A.R.; Galili, G. The multifaceted role of aspartate-family amino acids in plant metabolism. J. Exp. Bot. 2012, 63, 4995–5001. [Google Scholar] [CrossRef] [Green Version]
- Filiz, E.; Cetin, D.; Akbudak, M.A. Aromatic amino acids biosynthesis genes identification and expression analysis under salt and drought stresses in Solanum lycopersicum L. Sci. Hortic. 2019, 250, 127–137. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Szabados, L.; Kovács, H.; Zilberstein, A.; Bouchereau, A. Plants in extreme environments: Importance of protective compounds in stress tolerance. In Advances in Botanical Research; Ismail, T., Ed.; Academic Press: Cambridge, MA, USA, 2011; Volume 57, pp. 105–150. [Google Scholar]
- Smirnoff, N.; Cumbes, Q.J. Hydroxyl radical scavenging activity of compatible solutes. Phytochemistry 1989, 28, 1057–1060. [Google Scholar] [CrossRef]
- Joshi, V.; Joung, J.-G.; Fei, Z.; Jander, G. Interdependence of threonine, methionine and isoleucine metabolism in plants: Accumulation and transcriptional regulation under abiotic stress. Amino Acids 2010, 39, 933–947. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, J.; Huang, Z.; Zhu, Q.; Wang, L. Remobilization of carbon reserves is improved by controlled soil-drying during grain filling of wheat. Crop. Sci. 2000, 40, 1645–1655. [Google Scholar] [CrossRef]
- Lunn, J.E. Compartmentation in plant metabolism. J. Exp. Bot. 2007, 58, 35–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, G.; McIntyre, C.L.; Jenkins, C.L.D.; Glassop, D.; van Herwaarden, A.F.; Shorter, R. Molecular dissection of variation in carbohydrate metabolism related to water-soluble carbohydrate accumulation in stems of wheat. Plant Physiol. 2008, 146, 441–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wingler, A.; Roitsch, T. Metabolic regulation of leaf senescence: Interactions of sugar signalling with biotic and abiotic stress responses. Plant Biol. 2008, 10, 50–62. [Google Scholar] [CrossRef]
- Aggarwal, P.K.; Singh, A.K.; Chaturvedi, G.S.; Sinha, S.K. Performance of wheat and triticale cultivars in a variable soil-water environment II. Evapotranspiration, water use efficiency, harvest index and grain yield. Field Crops Res. 1986, 13, 301–315. [Google Scholar] [CrossRef]
- Calderini, D.F.; Reynolds, M.P.; Slafer, G.A. Source-sink effects on grain weight of bread wheat, durum wheat, and triticale at different locations. Aust. J. Agric. Res. 2006, 57, 227–233. [Google Scholar] [CrossRef]
- Ugarte, C.; Calderini, D.F.; Slafer, G.A. Grain weight and grain number responsiveness to pre-anthesis temperature in wheat, barley and triticale. Field Crops Res. 2007, 100, 240–248. [Google Scholar] [CrossRef]
- Khan, M.; Khan, F.U.; Saleem, Z.; Khattak, S.G. Comparative suitability of wheat, triticale, barley, and oat in the ecology of Kohat, Pakistan. J. Biol. Agric. Healthc. 2015, 5, 202–204. [Google Scholar]
- Roohi, E.; Tahmasebi Sarvestani, Z.; Modarres-Sanavy, S.A.M.; Siosemardeh, A. Comparative Study on the effect of soil water stress on photosynthetic function of triticale, bread wheat, and barley. J. Agric. Sci. Technol. 2013, 15, 215–225. [Google Scholar]
- Planchon, C. Photosynthesis, transpiration, resistance to CO2 transfer, and water efficiency of flag leaf of bread wheat, durum wheat and triticale. Euphytica 1979, 28, 403–408. [Google Scholar] [CrossRef]
- Lonbani, M.; Arzani, A. Morpho-physiological traits associated with terminal droughtstress tolerance in triticale and wheat. Agron. Res. 2011, 9, 315–329. [Google Scholar]
- Silva, A.d.N.; Ramos, M.L.G.; Ribeiro, W.Q.; de Alencar, E.R.; da Silva, P.C.; de Lima, C.A.; Vinson, C.C.; Silva, M.A.V. Water stress alters physical and chemical quality in grains of common bean, triticale and wheat. Agric. Water Manag. 2020, 231, 106023. [Google Scholar] [CrossRef]
- Ballesteros-Rodríguez, E.; Martínez-Rueda, C.G.; Morales-Rosales, E.J.; Estrada-Campuzano, G.; González, G.F. Changes in number and weight of wheat and triticale grains to manipulation in source-sink relationship. Int. J. Agron. 2019, 2019, 9. [Google Scholar] [CrossRef]
- López-Castañeda, C.; Richards, R.A. Variation in temperate cereals in rainfed environments III. Water use and water-use efficiency. Field Crops Res. 1994, 39, 85–98. [Google Scholar] [CrossRef]
- López-Castañeda, C.; Richards, R.A. Variation in temperate cereals in rainfed environments II. Phasic development and growth. Field Crops Res. 1994, 37, 63–75. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Circular. Calif. Agric. Exp. Stn. 1950, 347. [Google Scholar]
- Ehdaie, B.; Alloush, G.A.; Madore, M.A.; Waines, J.G. Genotypic variation for stem reserves and mobilization in wheat: II. Postanthesis changes in internode water-soluble carbohydrates. Crop. Sci. 2006, 46, 2093–2103. [Google Scholar] [CrossRef]
- Cohen, S.A.; Michaud, D.P. Synthesis of a fluorescent derivatizing reagent, 6-Aminoquinolyl-N-Hydroxysuccinimidyl carbamate, and its application for the analysis of hydrolysate amino acids via high-performance liquid chromatography. Anal. Biochem. 1993, 211, 279–287. [Google Scholar] [CrossRef]
- Ehdaie, B.; Alloush, G.A.; Waines, J.G. Genotypic variation in linear rate of grain growth and contribution of stem reserves to grain yield in wheat. Field Crops Res. 2008, 106, 34–43. [Google Scholar] [CrossRef]
- Hou, J.; Huang, X.; Sun, W.; Du, C.; Wang, C.; Xie, Y.; Ma, Y.; Ma, D. Accumulation of water-soluble carbohydrates and gene expression in wheat stems correlates with drought resistance. J. Plant Physiol. 2018, 231, 182–191. [Google Scholar] [CrossRef]
- Wardlaw, I.F.; Willenbrink, J. Mobilization of fructan reserves and changes in enzyme activities in wheat stems correlate with water stress during kernel filling. New Phytol. 2000, 148, 413–422. [Google Scholar] [CrossRef]
- Simkin, A.J.; Faralli, M.; Ramamoorthy, S.; Lawson, T. Photosynthesis in non-foliar tissues: Implications for yield. Plant J. 2020, 101, 1001–1015. [Google Scholar] [CrossRef]
- Foulkes, M.J.; Sylvester-Bradley, R.; Weightman, R.; Snape, J.W. Identifying physiological traits associated with improved drought resistance in winter wheat. Field Crops Res. 2007, 103, 11–24. [Google Scholar] [CrossRef]
- Rebetzke, G.J.; van Herwaarden, A.F.; Jenkins, C.; Weiss, M.; Lewis, D.; Ruuska, S.; Tabe, L.; Fettell, N.A.; Richards, R.A. Quantitative trait loci for water-soluble carbohydrates and associations with agronomic traits in wheat. Aust. J. Agric. Res. 2008, 59, 891–905. [Google Scholar] [CrossRef]
- Li, M.; Liu, Y.; Ma, J.; Zhang, P.; Wang, C.; Su, J.; Yang, D. Genetic dissection of stem WSC accumulation and remobilization in wheat (Triticum aestivum L.) under terminal drought stress. BMC Genet. 2020, 21, 1–14. [Google Scholar] [CrossRef]
- Ruuska, S.A.; Rebetzke, G.J.; van Herwaarden, A.F.; Richards, R.A.; Fettell, N.A.; Tabe, L.; Jenkins, C.L.D. Genotypic variation in water-soluble carbohydrate accumulation in wheat. Funct. Plant Biol. 2006, 33, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Ghahraman, B.; Sepaskhah, A.R. Use of a water deficit sensitivity index for partial irrigation scheduling of wheat and barley. Irrig. Sci. 1997, 18, 11–16. [Google Scholar] [CrossRef]
- Molero, G.; Reynolds, M.P. Spike photosynthesis measured at high throughput indicates genetic variation independent of flag leaf photosynthesis. Field Crops Res. 2020, 255, 107866. [Google Scholar] [CrossRef]
- Smeekens, S.; Ma, J.; Hanson, J.; Rolland, F. Sugar signals and molecular networks controlling plant growth. Curr. Opin. Plant Biol. 2010, 13, 273–278. [Google Scholar] [CrossRef]
- Müller, B.; Pantin, F.; Génard, M.; Turc, O.; Freixes, S.; Piques, M.; Gibon, Y. Water deficits uncouple growth from photosynthesis, increase C content, and modify the relationships between C and growth in sink organs. J. Exp. Bot. 2011, 62, 1715–1729. [Google Scholar] [CrossRef] [Green Version]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [Green Version]
- Hummel, I.; Pantin, F.; Sulpice, R.; Piques, M.; Rolland, G.; Dauzat, M.; Christophe, A.; Pervent, M.; Bouteillé, M.; Stitt, M.; et al. Arabidopsis plants acclimate to water deficit at low cost through changes of carbon usage: An integrated perspective using growth, metabolite, enzyme, and gene expression analysis. Plant Physiol. 2010, 154, 357–372. [Google Scholar] [CrossRef] [Green Version]
- Prathap, V.; Ali, K.; Singh, A.; Vishwakarma, C.; Krishnan, V.; Chinnusamy, V.; Tyagi, A. Starch accumulation in rice grains subjected to drought during grain filling stage. Plant Physiol. Biochem. 2019, 142, 440–451. [Google Scholar] [CrossRef]
- He, J.-F.; Goyal, R.; Laroche, A.; Zhao, M.-L.; Lu, Z.-X. Water stress during grain development affects starch synthesis, composition and physicochemical properties in triticale. J. Cereal Sci. 2012, 56, 552–560. [Google Scholar] [CrossRef]
- Halford, N.G.; Curtis, T.Y.; Muttucumaru, N.; Postles, J.; Mottram, D.S. Sugars in crop plants. Ann. Appl. Biol. 2011, 158, 1–25. [Google Scholar] [CrossRef]
- Singh, R.; Juliano, B.O. Free sugars in relation to starch accumulation in developing rice grain. Plant Physiol. 1977, 59, 417. [Google Scholar] [CrossRef] [Green Version]
- Gous, P.W.; Hasjim, J.; Franckowiak, J.; Fox, G.P.; Gilbert, R.G. Barley genotype expressing “stay-green”-like characteristics maintains starch quality of the grain during water stress condition. J. Cereal Sci. 2013, 58, 414–419. [Google Scholar] [CrossRef]
- Ahmadi, A.; Baker, D.A. The effect of water stress on the activities of key regulatory enzymes of the sucrose to starch pathway in wheat. Plant Growth Regul. 2001, 35, 81–91. [Google Scholar] [CrossRef]
- Distelfeld, A.; Avni, R.; Fischer, A. Senescence, nutrient remobilization, and yield in wheat and barley. J. Exp. Bot. 2014, 65, 3783–3798. [Google Scholar] [CrossRef] [Green Version]
- Koga, R.; Meng, T.; Nakamura, E.; Miura, C.; Irino, N.; Devkota, H.P.; Yahara, S.; Kondo, R. The effect of photo-irradiation on the growth and ingredient composition of young green barley (Hordeum vulgare). Agric. Sci. 2013, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Sharma, A.; Kaur, R.; Thukral, A.K.; Bhardwaj, R.; Ahmad, P. Differential distribution of amino acids in plants. Amino Acids 2017, 49, 821–869. [Google Scholar] [CrossRef]
- Siddiqi, R.A.; Singh, T.P.; Rani, M.; Sogi, D.S.; Bhat, M.A. Diversity in grain, flour, amino acid composition, protein profiling, and proportion of total flour proteins of different wheat cultivars of North India. Front. Nutri. 2020, 7, 141. [Google Scholar] [CrossRef]
- Raffan, S.; Halford, N.G. Cereal asparagine synthetase genes. Ann. Appl. Biol. 2021, 178, 6–22. [Google Scholar] [CrossRef]
- Hare, P.D.; Cress, W.A. Metabolic implications of stress-induced proline accumulation in plants. Plant Growth Regul. 1997, 21, 79–102. [Google Scholar] [CrossRef]
- Gregersen, P.L. Senescence and nutrient remobilization in crop plants. In The Molecular and Physiological Basis of Nutrient Use Efficiency in Crops; Hawkesford, M., Barraclough, P., Eds.; Wiley-Blackwell: Oxford, UK, 2011; pp. 83–102. [Google Scholar]
- Branković, G.; Pajić, V.; Zivanović, T.; Dodig, D.; Kandić, V.; Knežević, D.; Đurić, N. Genetic parameters of Triticum aestivum and Triticum durum for technological quality properties in Serbia. Zemdirb. Agric. 2018, 105, 39–48. [Google Scholar] [CrossRef]
- Guerrieri, N.; Cavaletto, M. 8-Cereals proteins. In Proteins in Food Processing, 2nd ed.; Yada, R.Y., Ed.; Woodhead Publishing: Cambridge, UK, 2018; pp. 223–244. [Google Scholar]
- Giunta, F.; Pruneddu, G.; Zuddas, M.; Motzo, R. Bread and durum wheat: Intra- and inter-specific variation in grain yield and protein concentration of modern Italian cultivars. Eur. J. Agron. 2019, 105, 119–128. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WSC (g stem−1) | MWSC (g stem−1) | ME (%) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Max | Min | ||||||||||||
| Cereal | WW | WL | ANOVA T | WW | WL | ANOVA T | WW | WL | ANOVA T | WW | WL | ANOVA T | |
| Triticale | 0.53 | 0.51 a | ns | 0.09 a | 0.03 | * | 0.44 | 0.47 a | ns | 81.83 | 94.16 | * | |
| Upper | Bread wheat | 0.28 | 0.18 b | ns | 0.05 b | 0.01 | * | 0.23 | 0.17 b | ns | 82.63 | 96.76 | * |
| stem | Durum wheat | 0.27 | 0.27 b | ns | 0.04 b | 0.01 | ** | 0.23 | 0.25 b | ns | 83.45 | 95.35 | *** |
| Barley | 0.23 | 0.24 b | ns | 0.03 b | 0.02 | ns | 0.2 | 0.23 b | ns | 88.54 | 93.37 | * | |
| ANOVA G | ns | * | * | ns | ns | * | ns | ns | |||||
| Triticale | 2.94 a | 1.74 a | * | 0.03 | 0.06 a | ns | 2.91 a | 1.68 a | * | 98.89 | 96.32 | ns | |
| Lower | Bread wheat | 0.99 b | 0.59 b | * | 0.06 | 0.01 b | ns | 0.93 b | 0.58 b | * | 93.94 | 97.98 | ns |
| stem | Durum wheat | 0.66 b | 0.54 b | ns | 0.09 | 0.02 b | ns | 0.56 b | 0.51 b | ns | 85.58 | 95.44 | ns |
| Barley | 0.48 b | 0.41 b | ns | 0.04 | 0.02 b | ns | 0.44 b | 0.39 b | ns | 91.26 | 92.85 | ns | |
| ANOVA G | *** | *** | ns | * | *** | *** | ns | ns | |||||
| Triticale | 3.34 a | 2.24 a | * | 0.12 | 0.09 a | ns | 3.22 a | 2.15 a | * | 96.25 | 95.74 | ns | |
| Bread wheat | 1.27 b | 0.76 b | * | 0.11 | 0.02 b | * | 1.16 b | 0.75 b | * | 91.42 | 97.59 | ns | |
| Stem | Durum wheat | 0.93 b | 0.80 b | ns | 0.14 | 0.04 b | * | 0.79 b | 0.77 b | ns | 85.14 | 95.39 | * |
| Barley | 0.67 b | 0.65 b | ns | 0.07 | 0.04 b | ns | 0.60 b | 0.62 b | ns | 90.1 | 93.28 | ns | |
| ANOVA G | *** | *** | ns | * | *** | *** | ns | ns | |||||
| Cereal | Trat | SN | GW | AB |
|---|---|---|---|---|
| Triticale | WW | 2 a | 6.69 ab | 13.63 a |
| WL | 2 a | 5.45 ab | 12.41 a | |
| Bread wheat | WW | 3 ab | 6.35 b | 13.19 a |
| WL | 3 ab | 5.09 b | 11.48 a | |
| Durum wheat | WW | 5 c | 6.73 b | 14.19 a |
| WL | 4 bc | 4.63 b | 11.42 a | |
| Barley | WW | 10 d | 7.49 a | 18.75 b |
| WL | 9 d | 5.43 a | 15.62 b | |
| ANOVA | ||||
| G | *** | * | *** | |
| T | * | *** | ** | |
| G × T | n.s. | n.s. | n.s. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Méndez-Espinoza, A.M.; Garriga, M.; Ben Mariem, S.; Soba, D.; Aranjuelo, I.; del Pozo, A. Carbohydrate and Amino Acid Dynamics during Grain Growth in Four Temperate Cereals under Well-Watered and Water-Limited Regimes. Agronomy 2021, 11, 1516. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081516
Méndez-Espinoza AM, Garriga M, Ben Mariem S, Soba D, Aranjuelo I, del Pozo A. Carbohydrate and Amino Acid Dynamics during Grain Growth in Four Temperate Cereals under Well-Watered and Water-Limited Regimes. Agronomy. 2021; 11(8):1516. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081516
Chicago/Turabian StyleMéndez-Espinoza, Ana María, Miguel Garriga, Sinda Ben Mariem, David Soba, Iker Aranjuelo, and Alejandro del Pozo. 2021. "Carbohydrate and Amino Acid Dynamics during Grain Growth in Four Temperate Cereals under Well-Watered and Water-Limited Regimes" Agronomy 11, no. 8: 1516. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081516